
Effect of Plant Growth Regulators on Different Explants of Artemisia ludoviciana under Photoperiod and Darkness Conditions and Their Influence on Achillin Production

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Equipment
2.2. Plant Material
2.2.1. Extraction and Isolation of Achillin from Wild Plant
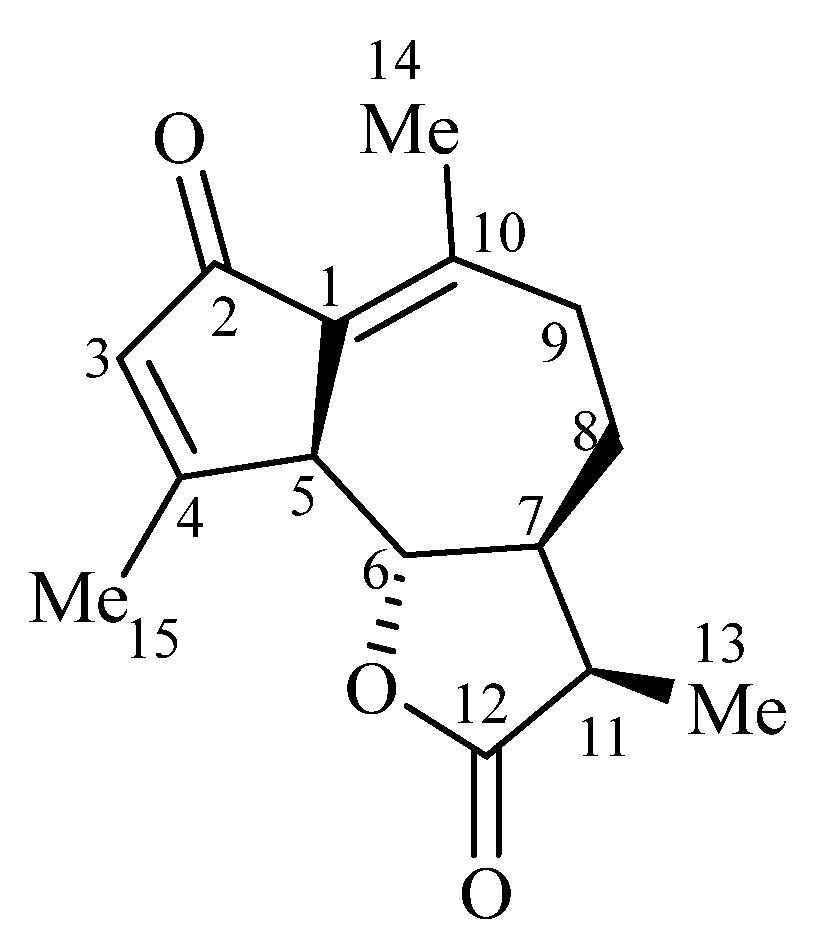
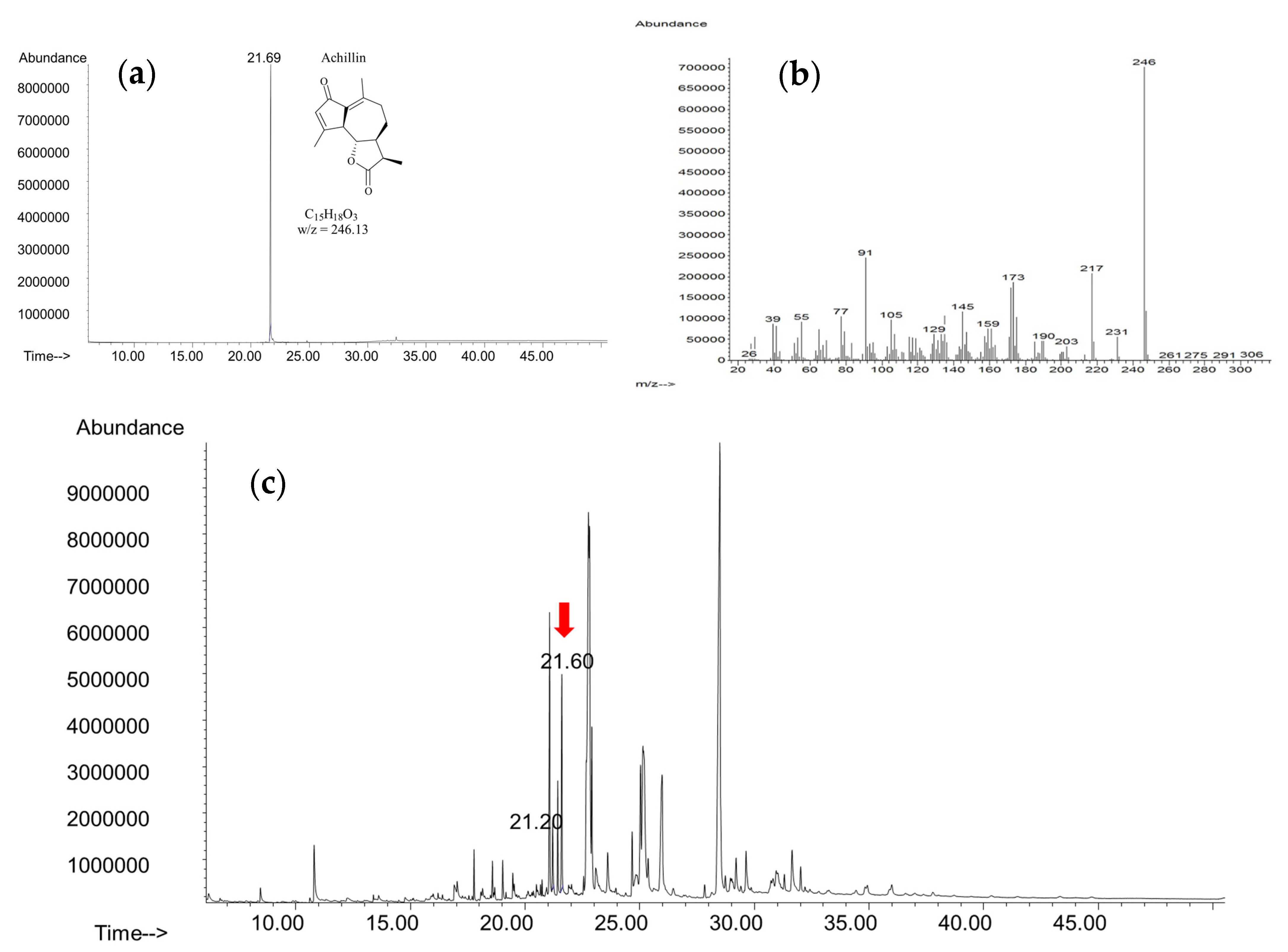
2.2.2. Characterization of Achillin Compound
2.3. Obtaining In Vitro Plantlets of A. ludoviciana
2.3.1. Evaluation of PGRs in Explants
2.3.2. Evaluation of Photoperiod and Total Darkness in Explants
2.4. Extraction and Quantification of Achillin from In Vitro Cultures
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effect of PGRs in Leaf Explants
3.2. Effect of PGRs in Nodal Explants
3.3. Effect of PGRs in Internodal Explants
3.4. Effect of PGRs in Root Explants
3.5. Effect of Photoperiod on Morphogenetic Response
3.6. Achillin Quantification in In Vitro Cultures Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ivanescu, B.; Miron, A.; Corciova, A. Sesquiterpene lactones from Artemisia Genus: Biological activities and methods of analysis. J. Anal. Methods Chem. 2015, 2015, 247685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merfort, I. Perspectives on sesquiterpene lactones in inflammation and cancer. Curr. Drug Targets 2011, 12, 1560–1573. [Google Scholar] [CrossRef] [PubMed]
- Moujir, L.; Callies, O.; Sousa, P.M.C.; Sharopov, F.; Seca, A.M.L. Applications of sesquiterpenes lactones: A review of some potential success cases. Appl. Sci. 2020, 10, 3001. [Google Scholar] [CrossRef]
- Palacios-Espinosa, J.F.; Núñez-Aragón, P.N.; Gomez-Chang, E.; Linares, E.; Bye, R.; Romero, I. Anti-helicobacter pylori activity of Artemisia ludoviciana subsp. mexicana and two of its bioactive components, estafiatin and eupatilin. Molecules 2021, 26, 3654. [Google Scholar] [CrossRef]
- Abad, M.M.J.; Bedoya, D.O.L.M.; Apaza, T.L.; Bermejo, B.P. The Artemisia L. Genus: A review of bioactive sesquiterpene lactones. Stud. Nat. Prod. Chem. 2012, 37, 43–65. [Google Scholar]
- Zhang, S.; Won, Y.-K.; Ong, C.N.; Shen, H.-M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem. Anticancer Agents 2005, 5, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Argueta, A.; Cano, L.M.; Rodarte, M.E. Atlas de las Plantas de la Medicina Tradicional Mexicana, Tomo I–III, 1st ed.; Instituto Nacional Indigenista: Mexico City, Mexico, 1994. [Google Scholar]
- Epstein, W.W.; Ubben, J.E.E. Anthemidin, a new sesquiterpene lactone from Artemisia ludoviciana. J. Nat. Prod. 1979, 42, 279–281. [Google Scholar] [CrossRef]
- Jakupovic, J.; Tan, R.X.; Bohlmann, F.; Boldt, P.E.; Jia, Z.J. Sesquiterpene lactones from Artemisia ludoviciana. Phytochemistry 1991, 30, 1573–1577. [Google Scholar] [CrossRef]
- Ruiz-Cancino, A.; Cano, A.E.; Delgado, G. Sesquiterpene lactones and flavonoids from Artemisia ludoviciana ssp. Mexicana. Phytochemistry 1993, 33, 1113–1115. [Google Scholar] [CrossRef]
- Said, F.S.; Ramos, G.M.C.; Mata, C.B.D.; Vargas, V.J.; Villarreal, T.L. In vitro antiprotozoal activity of the leaves of Artemisia ludoviciana. Fitoterapia 2005, 76, 466–468. [Google Scholar] [CrossRef]
- Anaya-Eugenio, G.D.; Rivero-Cruz, I.; Rivera-Chávez, J.; Mata, R. Hypoglycemic properties of some preparations and compounds from Artemisia ludoviciana Nutt. J. Ethnopharmacol. 2014, 155, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Anaya-Eugenio, G.D.; Rivero-Cruz, I.; Bye, R.; Linares, E.; Mata, R. Antinociceptive activity of the essential oil from Artemisia ludoviciana. J. Ethnopharmacol. 2016, 179, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Soto, S.; Sánchez-Rencillas, A.; Navarrete-Vázquez, G.; Castillo-España, P.; Villalobos-Molina, R.; Ibarra-Barajas, M. Relaxant effects of Artemisia ludoviciana on isolated rat smooth muscle tissues. J. Ethnopharmacol. 2012, 139, 513–518. [Google Scholar] [CrossRef]
- Ohno, N.; Gershenzon, J.; Roane, C.; Mabry, T.J. 11,13-dehydrodesacetylmatricarin and other sesquiterpene lactones from Artemisia ludoviciana Var. Ludoviciana and the identity of artecanin and chryrsartemin B. Phytochemistry 1980, 19, 103–106. [Google Scholar] [CrossRef]
- Rivero-Cruz, I.; Anaya-Eugenio, G.; Pérez-Vásquez, A.; Martínez, L.; Mata, R. Quantitative analysis and pharmacological effects of Artemisia ludoviciana aqueous extract and compounds. Nat. Prod. Commun. 2017, 12, 1531–1534. [Google Scholar] [CrossRef] [Green Version]
- Romero, J.L.; Sosa, C.M.; Burgoa, G.L.; Leal, A.C.; El Kassis, E.G.; Rodríguez, E.B.; Juárez, G.A.; Hernández, L.R.; Bach, H.; Juárez, Z.N. Antimycobacterial, cytotoxic, and anti-inflammatory activities of Artemisia ludoviciana. J. Ethnopharmacol. 2022, 293, 115249. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Carranza, J.N.; González-Maya, L.; Razo-Hernández, R.S.; Salas-Vidal, E.; Nolasco-Quintana, N.Y.; Clemente-Soto, A.F.; García-Arizmendi, L.; Sánchez-Ramos, M.; Marquina, S.; Alvarez, L. Achillin increases chemosensitivity to paclitaxel, overcoming resistance and enhancing apoptosis in human hepatocellular carcinoma cell line resistant to paclitaxel (Hep3B/PTX). Pharmaceutics 2019, 11, 512. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar]
- Rhodes, M.J.C. Physiological roles for secondary metabolites in plants: Some progress, many outstanding problems. Plant Mol. Biol. 1994, 24, 1–20. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Soni, U.; Brar, S.; Gauttam, V.K. Effect of seasonal variation on secondary metabolites of medicinal plants. Int. J. Pharm. Sci. Res. 2015, 6, 3654–3662. [Google Scholar]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Mishra, T. Climate change and production of secondary metabolites in medicinal plants: A review. Int. J. Herb. Med. 2016, 4, 27–30. [Google Scholar]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of deveplomental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Wawrosch, C.; Zotchev, S.B. Production of bioactive plant secondary metabolites through in vitro technologies-status and outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649–6668. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Dwivedi, P. Methyl-jasmonate and salicylic acid as potent elicitors for secondary metabolite production in medicinal plants: A review. J. Pharmacog. Phytochem. 2018, 7, 750–757. [Google Scholar]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- González, I.F.M.; González, D.E.F.; Morena, V.C. Secondary metabolites in plants: Main classes, phytochemical analysis and pharmacological activities. Bionatura 2019, 4, 1000–1009. [Google Scholar] [CrossRef]
- Savithramma, N.; Linga, M.R.; Suhrulatha, D. Screening of medicinal plants for secondary metabolites. Middle East J. Sci. Res. 2011, 8, 579–584. [Google Scholar]
- Jain, C.; Khatana, S.; Vijayvergia, R. Bioactivity of secondary metabolites of various plants: A review. Int. J. Pharm. Sci. Res. 2018, 10, 494–504. [Google Scholar]
- Vanisree, M.; Lee, C.-Y.; Lo, S.-F.; Nalawade, S.M.; Lin, C.Y.; Tsay, H.-S. Studies on the production of some important secondary metabolites from medicinal plants by tissue cultures. Bot. Bull. Acad. Sin. 2004, 45, 1–22. [Google Scholar]
- Yue, W.; Ming, Q.-L.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L.-P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2014, 1, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Isah, T.; Umar, S.; Mujib, A.; Prasad, M.S.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 132, 239–265. [Google Scholar] [CrossRef]
- Mulabagal, V.; Tsay, H.S. Plant cell cultures—An alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng./Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Ahuja, A.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Tiwari, G.; Mishra, N.; Bhargav, S.; Tiwari, S. Recent advancements on callus and cell suspension cultures: An effectual reserve for the production of pharmaceutically significant metabolites. Curr. Asp. Pharm. Res. Dev. 2021, 6, 96–111. [Google Scholar]
- Chattopadhyay, S.; Farkya, S.; Srivastava, A.K.; Bisaria, V.S. Bioprocess considerations for production of secondary metabolites by plant cell suspension cultures. Biotechnol. Bioprocess. Eng. 2002, 7, 138–149. [Google Scholar] [CrossRef]
- Moscatiello, R.; Baldan, B.; Navazio, L. Plant cell suspension cultures. In Plant Mineral Nutrients Methods and Protocols; Maathius, F.J.M., Ed.; Springer Protocols: Berlin/Heidelberg, Germany, 2012; pp. 77–93. [Google Scholar]
- Wu, T.; Kerbler, S.M.; Fernie, A.R.; Zhang, Y. Plant cell cultures as heterologous bio-factories for secondary metabolite production. Plant Commun. 2021, 2, 100235. [Google Scholar] [CrossRef]
- Marx, J.N.; White, E.H. The stereochemistry and synthesis of Achillin. Tetrahedron 1969, 25, 2117–2120. [Google Scholar] [CrossRef]
- Ho, C.; Ju, E.C.; Soo, G.Y.; Kim, K.-M.; Yong, S.I. Desacetylmatricarin, an Anti-allergic component from Taraxacum platycarpum. Planta Med. 1998, 64, 557–578. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.M.; Muñoz-Zamora, A.; Joseph-Nathan, P. Conformational analysis of achillin and leukodin. J. Nat. Prod. 1988, 51, 221–228. [Google Scholar] [CrossRef]
- Balboul, B.A.A.A.; Ahmed, A.A.; Otsuka, H.; Bando, M.; Kido, M.; Takeda, Y. A guaianolide and a germacranolide from Achillea santolina. Phytochemistry 1997, 46, 1045–1049. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio Assays with Tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Tariq, M.; Ali, G.; Hadi, F.; Ahmad, S.; Ali, N.; Ali, S.A. Callus induction and in vitro plant regeneration of Rice (Oryza sativa L.) under various conditions. Pak. J. Biol. Sci. 2008, 11, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Ahmed, M.; Mahaldar, D. In vitro callus induction and plant regeneration in seed explants of Rice (Oryza Sativa L.). J. Agric. Biol. Sci. 2005, 1, 72–75. [Google Scholar]
- Farjaminezhad, R.; Garoosi, G.-A. New biological trends on cell and callus growth and azadirachtin production in Azadirachta indica. Biotech 2019, 9, 309. [Google Scholar] [CrossRef]
- Bernabé-Antonio, A.; Maldonado-Magaña, A.; Ramírez-López, C.B.; Salcedo-Pérez, E.; Meza-Contreras, J.C.; González-García, Y.; López-Dellamary, T.F.A.; Cruz-Sosa, F. Establishment of callus and cell suspension cultures of Eysenhardtia polystachya (Ortega) and fungistatic activity of their extracts. S. Afr. J. Bot. 2017, 112, 40–47. [Google Scholar] [CrossRef]
- Rasool, R.; Ganai, B.A.; Kamili, A.N.; Akbar, S. Antioxidant potential in callus culture of Artemisia amygdalina Decne. Nat. Prod. Res. 2012, 26, 2103–2106. [Google Scholar]
- Thomas, T.D.; Yoichiro, H. In vitro propagation for the conservation of a rare medicinal plant Justicia gensarussa Burm. f. by nodal explants and shoot regeneration from callus. Acta Physiol. Plant 2010, 32, 943–950. [Google Scholar] [CrossRef]
- Sakpere, A.M.A.; Ajayi, S.A.; Adelusi, A.A. Effect of growth regulators and explant types on callus induction in Telfairia occidentalis Hook F. Afr. J. Biotechnol. 2014, 13, 2015–2021. [Google Scholar] [CrossRef] [Green Version]
- Osman, N.I.; Awal, A.; Sidik, N.J.; Abdullah, S. Callus induction and somatic embryogenesis from leaf and nodal explants of Lycium barbarum L. (Goji). Biotechnology 2013, 12, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.D.; Maseena, E.A. Callus induction and plant regeneration in Cardiospermum halicacabum Linn. an important medicinal plant. Sci. Hortic. 2006, 108, 332–336. [Google Scholar] [CrossRef]
- Kumlay, A.M.; Ercisli, S. Callus induction, shoot proliferation and root regeneration of potato (Solanum tuberosum L.) stem node and leaf explants under long-day conditions. Biotechnol. Biotecnol. Equip. 2015, 29, 1075–1084. [Google Scholar] [CrossRef] [Green Version]
- Pandey, V.P.; Kudakasseril, J.; Cherian, E.; Patani, G. Comparison of two methods for in vitro propagation of Rauwolfia serpentina from nodal explants. Indian Drugs 2007, 44, 514–519. [Google Scholar] [CrossRef]
- Dharmapal, S.P.; Najla, M.; Swetha, E.S.; Udayan, P.S.; Elyas, K.K. Callus induction and organogenesis from Tinospora formanii Udayan and Pradeep: A rare endemic plant. Trop. Plant Res. 2017, 4, 71–76. [Google Scholar] [CrossRef]
- Kumar, M.S.; Nandi, S.C. High frequency plant regeneration with histological analysis of organogenic callus from internode explants of Asteracantha longifolia Nees. J. Genet. Eng. Biotechnol. 2015, 13, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Bakhtiar, Z.; Mirjalili, M.H.; Sonboli, A. In vitro callus induction and micropropagation of Thymus persicus (Lamiaceae), an endangered medicinal plant. Crop. Breed. App. Biotechnol. 2016, 16, 48–54. [Google Scholar] [CrossRef]
- Arif, M.; Rauf, S.; Din, A.U.; Rauf, M.; Afrasiab, H. High frequency plant regeneration from leaf derived callus of Dianthus caryophyllus L. Am. J. Plant Sci. 2014, 5, 2454–2463. [Google Scholar] [CrossRef] [Green Version]
- Shirin, F.; Hossain, M.; Kabir, M.F.; Roy, M.; Sarker, S.R. Callus induction and plant regeneration from internodal and leaf explants of four potato (Solanum tuberosum L.) cultivars. World J. Agric. Sci. 2007, 3, 01–06. [Google Scholar]
- Khan, H.; Siddique, I.; Anis, M.; Khan, P.R. In vitro organogenesis from internode derived callus cultures of Capsicum annuum L. J. Plant Biochem. Biotechnol. 2011, 20, 84–89. [Google Scholar] [CrossRef]
- Sridhar, T.M.; Naidu, C.V. An efficient callus induction and plant regeneration of Solanum nigrum (L.)—An important antiulcer medicinal plant. J. Phytol. 2011, 3, 23–28. [Google Scholar]
- Arockiasamy, D.I.; Muthukumar, B.; Natarajan, E.; Britto, S.J. Plant regeneration from node and internode explants of Solanum trilobatum L. Plant Tissue Cult. 2002, 12, 93–97. [Google Scholar]
- Rani, G.; Virk, G.S.; Nagpal, A. Callus induction and plantlet regeneration in Withania somnifera (L.) dunal. In Vitro. Cell. Dev. Biol. Plant 2003, 39, 468–474. [Google Scholar] [CrossRef]
- Soorni, J.; Kahrizi, D. Effect of genotype, explant type and 2,4-D on cell differentiation and callus induction in Cumin (Cuminum cyminum L.) medicinal plant. J. Appl. Biotechnol. Rep. 2015, 2, 265–270. [Google Scholar]
- Bano, S.; Jabeen, M.; Rahim, F.; Ilahi, I. Callus induction and regeneration in seed explants of rice (Oryza sativa CV. Swat-II). Pak. J. Bot. 2005, 37, 829–836. [Google Scholar]
- Roy, P.K.; Barat, G.K.; Mehta, S.L. In vitro plant regeneration from callus derived from root explants of Lathyrus sativus. Plant Cell Tissue Organ Cult. 1992, 29, 135–138. [Google Scholar] [CrossRef]
- Zhong, J.J.; Bai, Y.; Wang, S.J. Effects of plant growth regulators on cell growth and ginsenoside saponin production by suspension cultures of Panax quinquefolium. J. Biotechnol. 1996, 45, 227–234. [Google Scholar] [CrossRef]
- Farhadi, N.; Panahandeh, J.; Azar, A.M.; Salte, S.A. Effects of explant type, growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of Persian shallot (Allium hirtifolium). Sci. Hortic. 2017, 218, 80–86. [Google Scholar] [CrossRef]
- Gao, F.; Peng, C.; Wang, H.; Shen, H.; Yang, L. Selection of culture conditions for callus induction and proliferation by somatic embryogenesis of Pinus koraiensis. J. For. Res. 2021, 32, 483–491. [Google Scholar] [CrossRef]
- Guo, G.; Ryong, B.J. Explant, medium, and plant growth regulator (PGR) affect induction and proliferation of callus in Abies koreana. Forests 2021, 12, 1388. [Google Scholar] [CrossRef]
- Fulzele, D.P.; Sipahimalani, A.T.; Heble, M.R. Tissue cultures of Artemisia annua: Organogenesis and Artemisinin production. Phytother Res. 1991, 5, 149–153. [Google Scholar] [CrossRef]
- Nin, S.; Morosi, E.; Schiff, S.; Bennici, A. Callus cultures of Artemisia absinthium L.: Initiation, growth optimization and organogenesis. Plant Cell Tissue Organ Cult. 1996, 45, 67–72. [Google Scholar] [CrossRef]
- Tariq, U.; Ali, M.; Abbasi, B.H. Morphogenic and biochemical variations under different spectral lights in callus cultures of Artemisia absinthium L. J. Photochem. Photobiol. B: Biol. 2014, 130, 264–271. [Google Scholar] [CrossRef] [PubMed]
- André, S.B.; Mongimaké, K.; Modeste, K.K.; Edmond, K.K.; Tchoa, K.; Hilare, K.T.; Yatty, K.Y. Effects of plant growth regulators and carbohydrates on callus induction and proliferation from leaf explant of Lippia multiflora Moldenke (Verbenacea). Int. J. Agric. Crop Sci. 2015, 8, 118–127. [Google Scholar]
- Ali, M.; Haider, B.A. Thidiazuron-induced changes in biomass parameters, total phenolic content, and antioxidant activity in callus cultures of Artemisia absinthium L. Appl. Biochem. Biotechnol. 2014, 172, 2363–2376. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.K.; Ahmed, S.A. Efficient callus induction, plant regeneration and estragole estimation in Tarragon (Artemisia dracunculus L.). J. Essent. Oil Res. 2011, 23, 16–20. [Google Scholar] [CrossRef]
- Dangash, A.; Ram, M.; Niranjan, R.; Bharillya, A.; Misra, H.; Pandya, N.; Jain, D.C. In vitro selection and hormonal regulation in cell culture of Artemisia annua L. Plant. JSM Cell Dev. Biol. 2015, 3, 1013. [Google Scholar]
- Aslam, N.; Zia, M.; Chaudhary, M.F. Callogenesis and direct organogenesis of Artemisia scoparia. Pak. J. Biol. Sci. 2006, 9, 1783–1786. [Google Scholar] [CrossRef]
- Ouyang, J.; Wang, X.; Zhao, B.; Wang, Y. Light intensity and spectral quality influencing the callus growth of Cistanche deserticola and biosynthesis of phenylethanoid glycosides. Plant Sci. 2003, 165, 657–661. [Google Scholar] [CrossRef]
- Wahyuni, D.K.; Huda, A.; Faizah, S.; Purnobasuki, H.; Wardojo, B.P.E. Effects of light, sucrose concentration and repetitive subculture on callus growth and medically important production in Justicia gendarussa Burm. f. Biotechnol. Rep. 2020, 27, e00473. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, S.; Kosari-Nasab, M.; Mahjouri, S.; Talebpour, A.H.; Movafeghi, A.; Maggi, F. Enhancement of in vitro production of volatile organic compounds by shoot differentiation in Artemisia spicigera. Plants 2021, 10, 208. [Google Scholar] [CrossRef]
- Morini, S.; D’Onofrio, C.; Bellocchi, G.; Fisichella, M. Effect of 2,4-D and light quality on callus production and differentiation from in vitro cultured quince leaves. Plant Cell Tissue Organ Cult. 2000, 63, 47–55. [Google Scholar] [CrossRef]
- Mohlakola, E.M.; Cheng, C.; Lin, Y.; Guo, R.; Min, K.T.; Chen, Y.; Lai, Z. Effects of 2,4-Dichlorophenoxy acetic acid and light on growth of Gerbera (Gerbera jamesonii cv. Daxueju) callus. ACS Agric. Sci. Technol. 2017, 18, 385–393. [Google Scholar]
- Irshad, M.; Debnath, B.; Mitra, S.; Arafat, Y.; Li, M.; Sun, Y.; Qiu, D. Accumulation of anthocyanin in callus cultures of red-pod okra [Abelmoschus esculentus (L.) Hongjiao] in response to light and nitrogen levels. Plant Cell Tissue Organ Cult. 2018, 134, 29–39. [Google Scholar] [CrossRef]
- Yousaf, R.; Khan, M.A.; Ullah, N.; Khan, I.; Hayat, O.; Shehzad, M.A.; Khan, I.; Taj, F.; Din, N.U.; Khan, A.; et al. Biosynthesis of anti-leishmanial natural products in callus cultures of Artemisia Scoparia. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1122–1131. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGRs | (mg/L) | Photoperiod | Darkness | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Leaf | Internode | Nodes | Root | Leaf | Internode | Nodes | Root | ||
| Control | 0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| KIN | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| BAP | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| NAA | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| 2,4-D | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| IAA | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| PIC | 0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| KIN/NAA | 0.1/0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 0 ± 0.00 e | 46.67 ± 2.89 b | 80 ± 10.00 b | 80.00 ± 5.00 b | 0 ± 0.00 d | 40 ± 5.00 c | 26.67 ± 2.89 c | 70 ± 5.00 b | |
| KIN/2,4-D | 0.1/0.1 | 0 ± 0.00 e | 71 ± 7.64 a | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 50 ± 5.00 c | 45 ± 5.00 b | 81.67 ± 2.89 b | 100 ± 0.00 a | 70.00 ± 5.00 b | 66.67 ± 2.89 b | 0 ± 0.00 d | 98.33 ± 2.86 a | |
| KIN/PIC | 0.1/0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 90 ± 5.0 a | 0 ± 0.00 c | 0 ± 0.00 d | 80.00 ± 5.00 b | 100 ± 0.00 a | 55 ± 5.00 b | 71.67 ± 5.77 ab | 93.33 ± 2.89 a | |
| KIN/IAA | 0.1/0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| BAP/NAA | 0.1/0.1 | 75 ± 5.0 b | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 95 ± 5.0 a | 0 ± 0.00 c | 100 ± 0.00 a | 0 ± 0.00 c | 95 ± 5.00 a | 0 ± 0.00 d | 83.33 ± 7.64 a | 0 ± 0.00 c | |
| BAP/2,4-D | 0.1/0.1 | 53.33 ± 5.77 c | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 16.67 ± 2.89 d | 31.67 ± 2.89 b | 33.33 ± 5.77 c | 0 ± 0.00 c | 58.33 ± 2.89 c | 33.33 ± 5.77 c | 30 ± 5.00 c | 0 ± 0.00 c | |
| BAP/PIC | 0.1/0.1 | 16.67 ± 2.89 d | 40 ± 10.00 b | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 85 ± 5.0 ab | 83.33 ± 7.64 a | 81.67 ± 2.89 b | 80 ± 5.00 b | 80 ± 5.00 b | 85.00 ± 5.00 a | 60 ± 10.00 b | 0 ± 0.00 c | |
| BAP/IAA | 0.1/0.1 | 0 ± 0.00 e | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c |
| 0.1/1.0 | 0 ± 0.00 e | 0 ± 0.00 c | 75 ± 5.00 b | 0 ± 0.00 c | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 c | |
| PGRs | (mg/L) | Photoperiod | Darkness | |
|---|---|---|---|---|
| Leaf | Leaf | Root | ||
| Control | 0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| KIN | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| BAP | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| NAA | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| 2,4-D | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| IAA | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| PIC | 0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| KIN/NAA | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 73.33 ± 5.77 b | 0 ± 0.00 c | 0 ± 0.00 b | |
| KIN/2,4-D | 0.1/0.1 | 98.33 ± 2.89 a | 95 ± 5.00 a | 100 ± 0.00 a |
| 0.1/1.0 | 100 ± 0.00 a | 0 ± 0.00 c | 0 ± 0.00 b | |
| KIN/PIC | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| KIN/IAA | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| BAP/NAA | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 96.67 ± 2.89 a | 0 ± 0.00 b | |
| BAP/2,4-D | 0.1/0.1 | 0 ± 0.00 c | 23.33 ± 2.89 b | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| BAP/PIC | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| BAP/IAA | 0.1/0.1 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 c | 0 ± 0.00 c | 0 ± 0.00 b | |
| PGR | (mg/L) | Nodes | Internodes | ||
|---|---|---|---|---|---|
| Photoperiod | Darkness | Photoperiod | Darkness | ||
| Control | 0.0 | 23.33 ± 5.77 c | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| KIN | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| BAP | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| NAA | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| 2,4-D | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| PIC | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| IAA | 0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| 1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| KIN/NAA | 0.1/0.1 | 0 ± 0.00 d | 23.33 ± 2.87 c | 0 ± 0.00 e | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| KIN/2,4-D | 0.1/0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b |
| 0.1/1.0 | 0 ± 0.00 d | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| KIN/PIC | 0.1/0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 91.67 ± 2.89 a | 0 ± 0.00 b |
| 0.1/1.0 | 95 ± 5.00 a | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| KIN/IAA | 0.1/0.1 | 0 0 ± 0.00 d | 23.33 ± 2.89 c | 0 ± 0.00 e | 0 ± 0.00 b |
| 0.1/1.0 | 26.67 ± 2.89 c | 93.33 ± 2.89 a | 0 ± 0.00 e | 0 ± 0.00 b | |
| BAP/NAA | 0.1/0.1 | 28.33 ± 2.89 c | 23.33 ± 2.89 c | 0 ± 0.00 e | 0 ± 0.00 b |
| 0.1/1.0 | 41.67 ± 2.89 b | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| BAP/2,4-D | 0.1/0.1 | 96.67 ± 2.89 a | 33.33 ± 2.89 b | 0 ± 0.00 e | 0 ± 0.00 b |
| 0.1/1.0 | 33.33 ± 2.89 bc | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| BAP/PIC | 0.1/0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 16.67 ± 5.78 d | 0 ± 0.00 b |
| 0.1/1.0 | 98.33 ± 2.89 a | 0 ± 0.00 d | 0 ± 0.00 e | 0 ± 0.00 b | |
| BAP/IAA | 0.1/0.1 | 0 ± 0.00 d | 0 ± 0.00 d | 36.67 ± 5.78 c | 100 ± 0.00 a |
| 0.1/1.0 | 91.67 ± 2.89 a | 0 ± 0.00 d | 53.33 ± 2.89 b | 0 ± 0.00 b | |
| Plant Resource | Morphogenetic Response | Growing Conditions | PGRs (mg/L) | Achillin (µg/g Dry Biomass) | |
|---|---|---|---|---|---|
| Auxin | Cytokinin | ||||
| Leaves | Callus | Photoperiod | PIC (1.0) | KIN (0.1) | 1703.05 ± 40.92 |
| Nodes | Callus | Total darkness | NAA (1.0) | BAP (0.1) | 1041.15 ± 16.47 |
| Leaves | Callus | Photoperiod | 2,4-D (1.0) | BAP (0.1) | 88.34 ± 8.15 |
| Roots | Callus | Total darkness | 2,4-D (1.0) | BAP (0.1) | 12.79 ± 13.98 |
| Nodes | Plantlets | Photoperiod | PIC (0.1) | BAP (0.1) | 1880.01 ± 42.67 |
| Internodes | Plantlets | Photoperiod | PIC (0.1) | KIN (0.1) | 334.03 ± 13.29 |
| Internodes | Plantlets | Photoperiod | NAA (0.1) | BAP (0.1) | 28.11 ± 1.03 |
| Internodes | Plantlets | Photoperiod | PIC (0.1) | BAP (0.1) | 1482.17 ± 53.44 |
| Internodes | Plantlets | Total darkness | IAA (0.1) | BAP (0.1) | 53.46 ± 12.95 |
| Internodes | Plantlets | Photoperiod | IAA (0.1) | BAP (0.1) | 1589.25 ± 30.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Ramos, M.; Berman-Bahena, S.; Alvarez, L.; Sánchez-Carranza, J.N.; Bernabé-Antonio, A.; Román-Guerrero, A.; Marquina-Bahena, S.; Cruz-Sosa, F. Effect of Plant Growth Regulators on Different Explants of Artemisia ludoviciana under Photoperiod and Darkness Conditions and Their Influence on Achillin Production. Processes 2022, 10, 1439. https://doi.org/10.3390/pr10081439
Sánchez-Ramos M, Berman-Bahena S, Alvarez L, Sánchez-Carranza JN, Bernabé-Antonio A, Román-Guerrero A, Marquina-Bahena S, Cruz-Sosa F. Effect of Plant Growth Regulators on Different Explants of Artemisia ludoviciana under Photoperiod and Darkness Conditions and Their Influence on Achillin Production. Processes. 2022; 10(8):1439. https://doi.org/10.3390/pr10081439
Chicago/Turabian StyleSánchez-Ramos, Mariana, Samantha Berman-Bahena, Laura Alvarez, Jessica Nayelli Sánchez-Carranza, Antonio Bernabé-Antonio, Angélica Román-Guerrero, Silvia Marquina-Bahena, and Francisco Cruz-Sosa. 2022. "Effect of Plant Growth Regulators on Different Explants of Artemisia ludoviciana under Photoperiod and Darkness Conditions and Their Influence on Achillin Production" Processes 10, no. 8: 1439. https://doi.org/10.3390/pr10081439