Are There Differences in the Reaction of the Light-Tolerant Subgenus Pinus spp. Biomass to Climate Change as Compared to Light-Intolerant Genus Picea spp.?

 ,
,
Abstract
:1. Introduction
2. Material and Methods
→lnV = f [lnA, lnN, (lnA)(lnN), ln(Tm + 40), lnPRm, [ln(Tm + 40)]·(lnPRm)} →
→lnPi = f {lnA, lnV, lnN, (lnA)(lnN), ln(Tm + 40), lnPRm, [ln(Tm + 40)]·(lnPRm)}
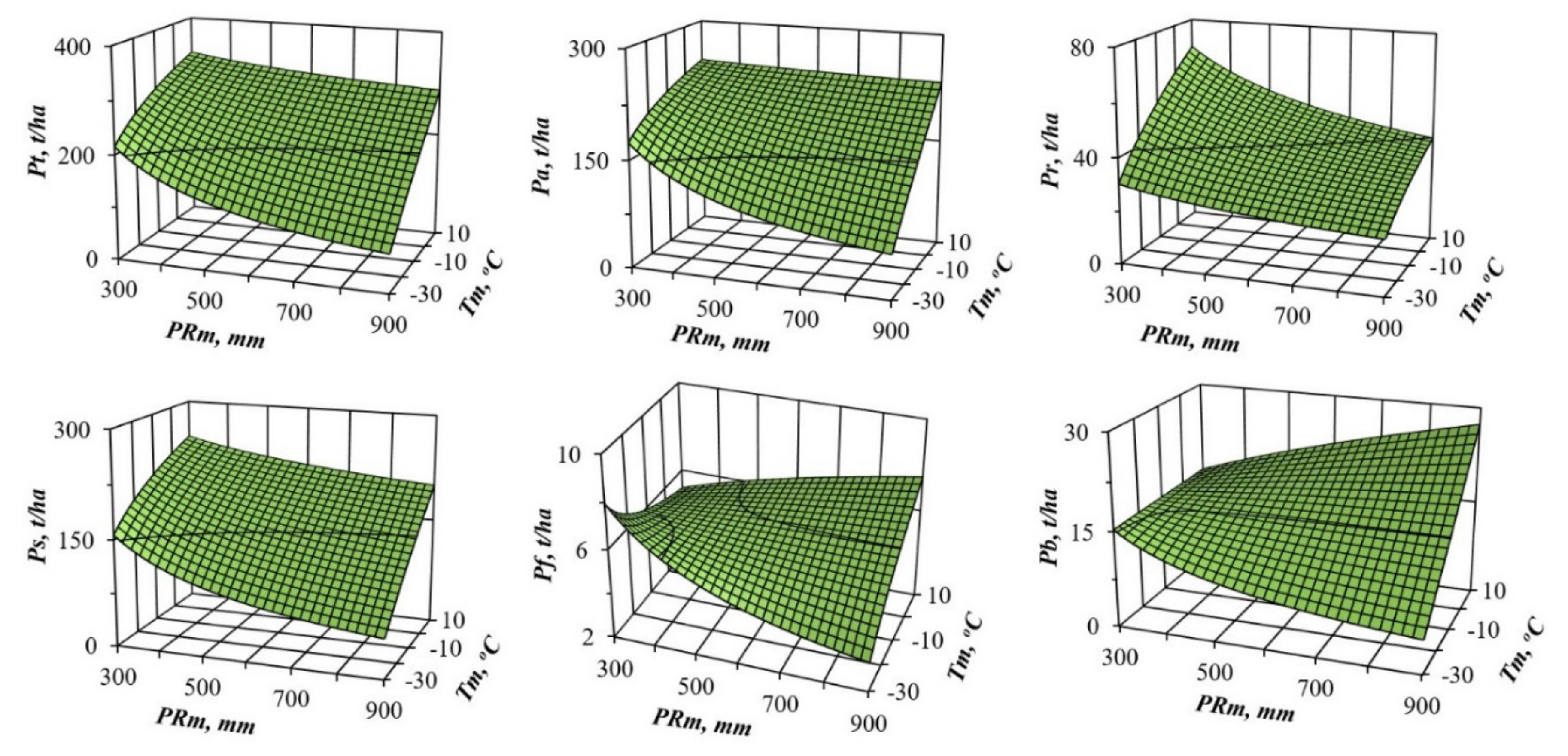
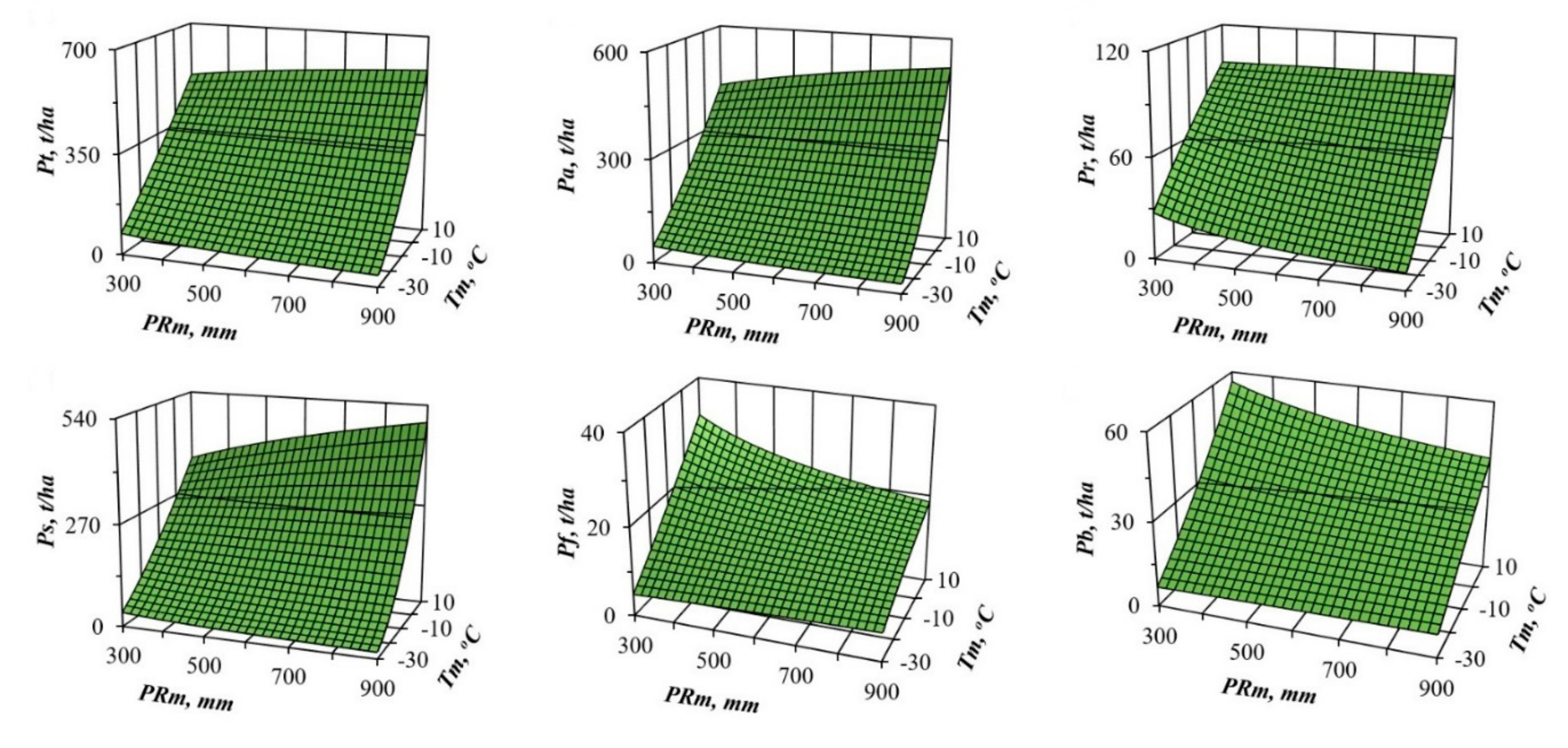
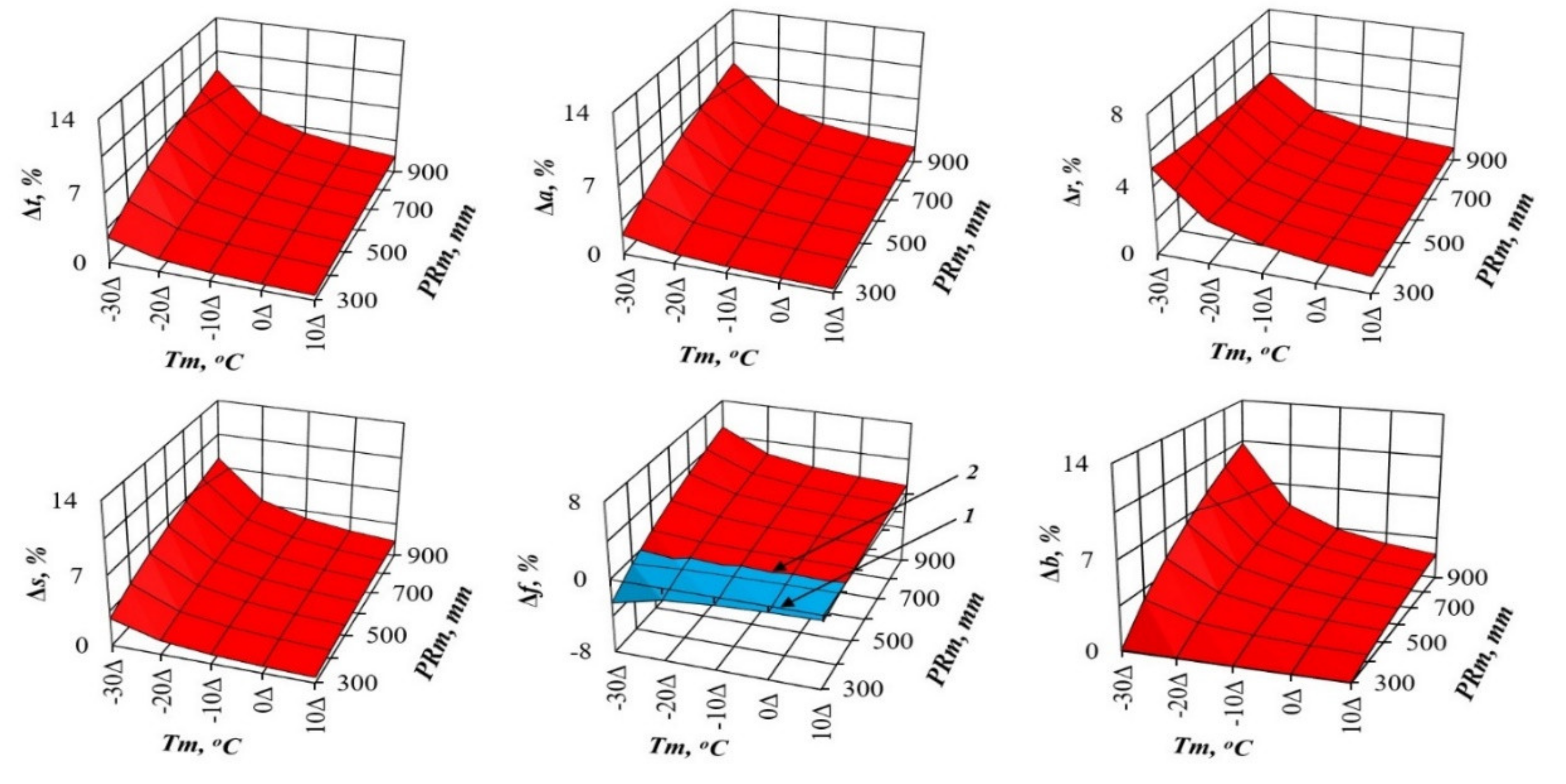
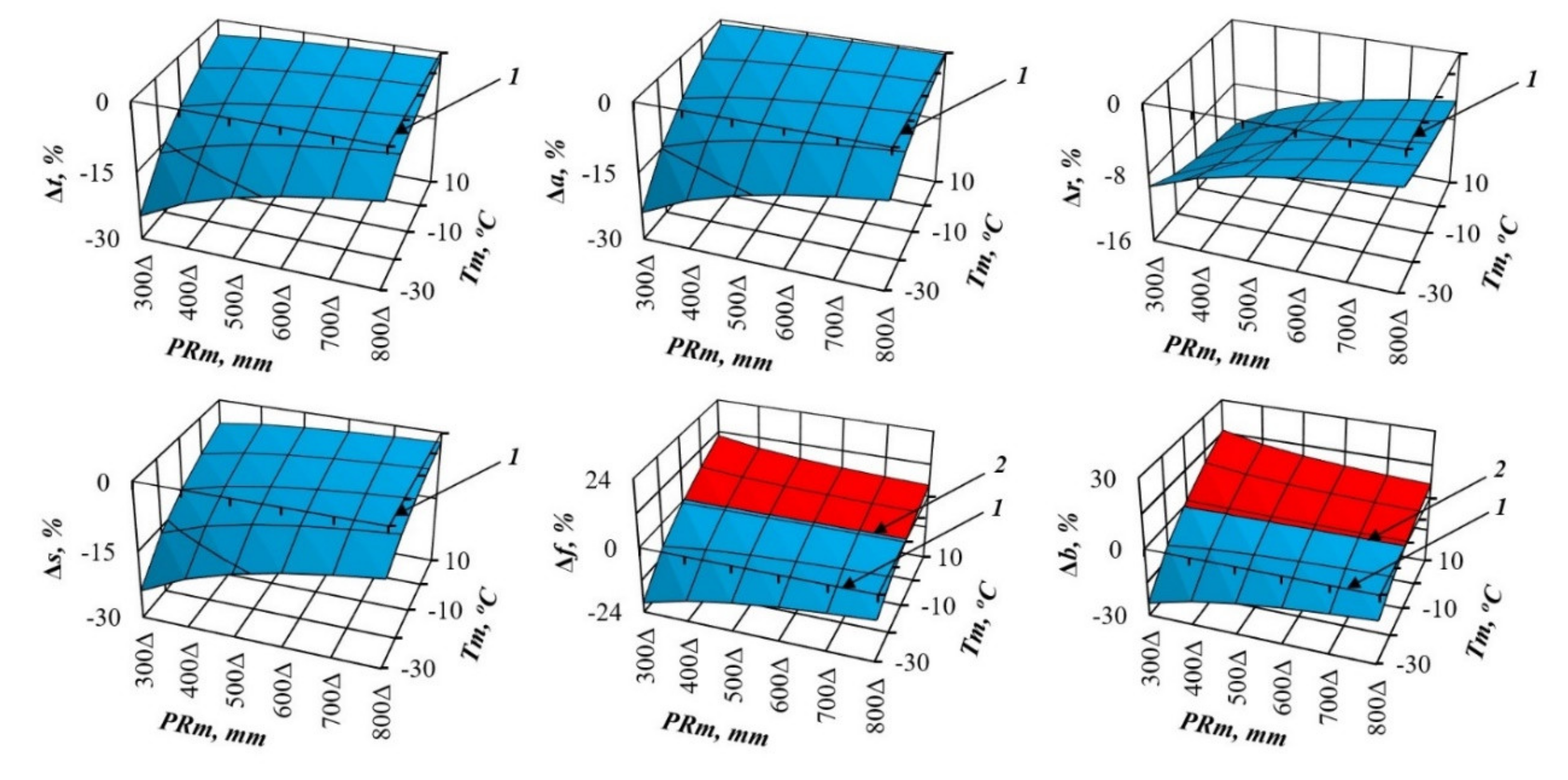
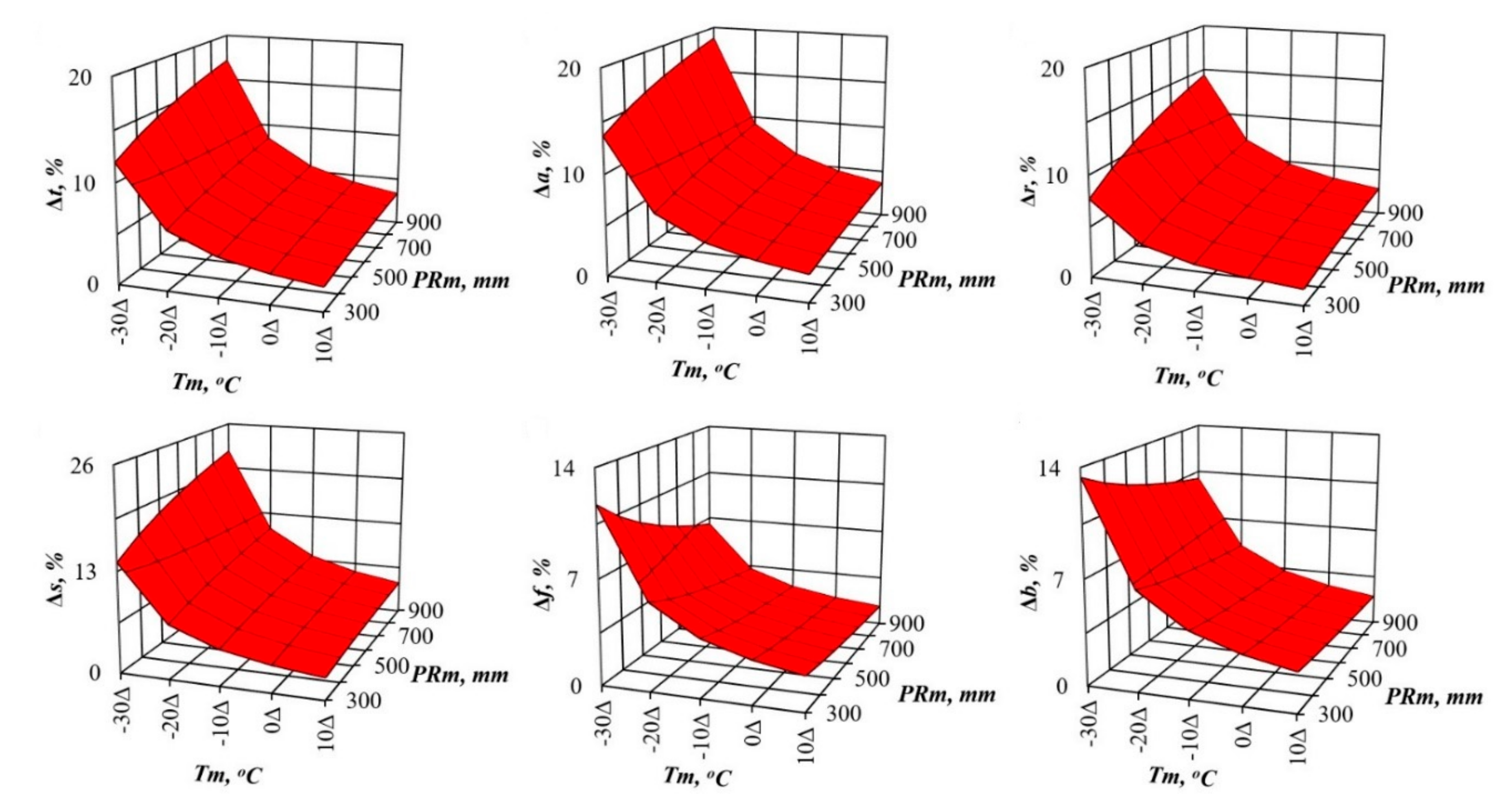
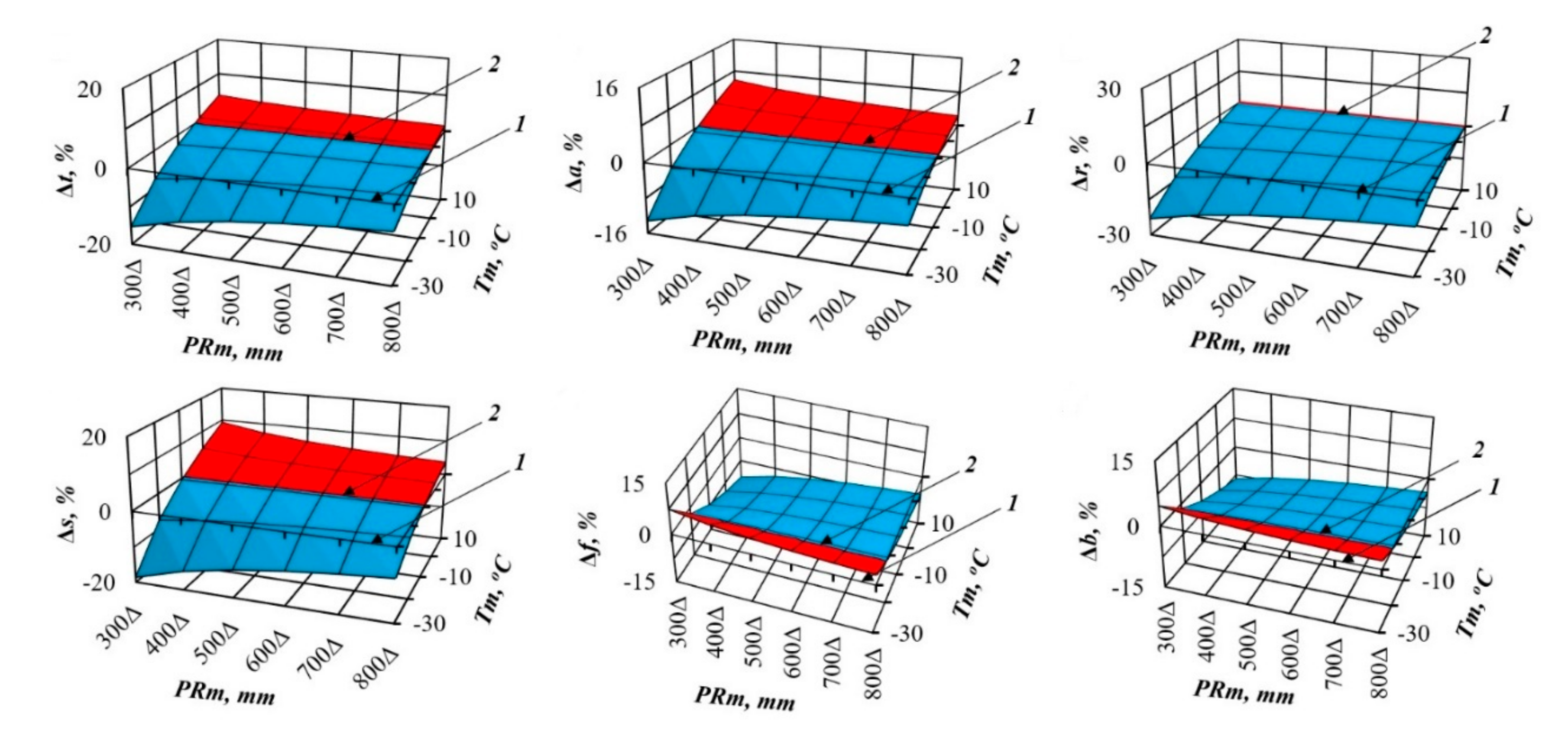
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Morin, X.; Fahse, L.; Jactel, H.; Scherer-Lorenzen, M.; García-Valdés, R.; Bugmann, H. Long-term response of forest productivity to climate change is mostly driven by change in tree species composition. Sci. Rep. 2018, 8, 5627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauhkonen, J.; Packalen, T. Uncertainties related to climate change and forest management with implications on climate regulation in Finland. Ecosyst. Serv. 2018, 33, 103–246. [Google Scholar] [CrossRef]
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest adaptation to climate change—Is non-management an option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef] [Green Version]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellomäki, S.; Strandman, H.; Nuutinen, T.; Peltola, H.; Korhonen, K.T.; Väisänen, H. Adaptation of Forest Ecosystems, Forests and Forestry to Climate Change; FINADAPT Working Paper 4; Finnish environmental Institute: Mimeographs, Helsinki, 2005; p. 44. [Google Scholar]
- Santini, M.; Collalti, A.; Valentini, R. Climate change impacts on vegetation and water cycle in the Euro-Mediterranean region, studied by a likelihood approach. Reg. Environ. Chang. 2014, 14, 1405–1418. [Google Scholar] [CrossRef]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehman, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Behrensmeyer, A. Atmosphere. Climate change and human evolution. Science 2006, 311, 476–478. [Google Scholar]
- Laczko, F.; Aghazarm, C. Migration, Environment and Climate Change: Assessing the Evidence; International Organization for Migration: Geneva, The Switzerland, 2009; p. 441. [Google Scholar]
- Miles-Novelo, A.; Anderson, C.A. Climate change and psychology: Effects of rapid global warming on violence and aggression. Curr. Clim. Chang. Rep. 2019, 5, 36–46. [Google Scholar] [CrossRef]
- Xu, C.; Kohler, T.A.; Lenton, T.M.; Svenning, J.-C.; Scheffer, M. Future of the human climate niche. Proc. Natl. Acad. Sci. USA 2020, 117, 11350–11355. [Google Scholar] [CrossRef]
- Beketov, A.N. On the influence of climate on the growth of pine and spruce. In Proceedings of the 1st Congress of naturalists in St. Petersburg. Botan. Part, St. Petersburg, Russia; 1868; pp. 111–163. [Google Scholar]
- Shvedov, F.N. Tree as a chronicle of droughts. Meteorol. Vestn. 1892, 5, 163–178. [Google Scholar]
- Klyuchnikov, V.F. The course of oak growth in the Krapivenskoye forest farm of the Tula province. Lesn. Zhurnal 1902, 2, 21–30. [Google Scholar]
- Douglas, A.E. Climatic Cycles and Trees-Growth. A Study of the Annual Rings of Trees in Relation to Climate and Solar Activity; Carnegie Institution of Washington: Washington, WA, USA, 1919; Publication No. 289; p. 127. [Google Scholar]
- Zaozersky, S.G. On the method of retrospective identification of climatic conditions by studying the course of tree growth. Irrig. Probl. 1934, 1, 75–86. [Google Scholar]
- Tolsky, A.P. On the issue of detecting climate fluctuations by analyzing the growth of trees. Work. Agric. Meteorol. 1936, 24, 117–123. [Google Scholar]
- Bitvinskas, T.T. Dendroclimatic Research; Hydrometeoizdat: Leningrad, Russia, 1974; p. 172. [Google Scholar]
- Shiyatov, S.G. Dendrochronology of the Upper Border of the Forest in the Urals; “Nauka” Publishing: Moscow, Russia, 1986; p. 136. [Google Scholar]
- Kostin, S.I. Solar activity and its influence on the growth of trees and the state of forests in the central part of the forest-steppe of the Russian plain. In Proceedings of the Main Geophysical Observatory Named after A. I. Voeikov, St. Petersburg, Russia; 1961; Volume 111, pp. 108–117. [Google Scholar]
- Olenin, S.M. Dynamics of Radial Growth of Stands of Pine Phytocenoses of the Middle Taiga Subzone of the Pre-Urals. Ph.D. Thesis, The Institute of Plant and Animal Ecology of the Ural branch of the Russian Academy of Sciences, Sverdlovsk, Russia, 1982; p. 18.
- Ricklefs, R.E. Fundamentals of Common Ecology; “Mir” Publishing: Moscow, Russia, 1979; p. 424. [Google Scholar]
- Babst, F.; Bodesheim, P.; Charney, N.; Friend, A.D.; Girardin, M.P.; Klesse, S.; Moore, D.J.P.; Seftigen, K.; Björklund, J.; Bouriaud, O.; et al. When tree rings go global: Challenges and opportunities for retro- and prospective insight. Quat. Sci. Rev. 2018, 197, 1–20. [Google Scholar] [CrossRef]
- Röhle, H.; Gerold, D.; Gemballa, R. Beziehungen zwischen Klima und Zuwachs, dargestellt am Beispiel von Fichte, Kiefer und Buche in Sachsen. Allg. Forst-Und Jagdztg. 2010, 181, 21–35. [Google Scholar]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Fang, O.; Wang, Y.; Shao, X. The effect of climate on the net primary productivity (NPP) of Pinus koraiensis in the Changbai Mountains over the past 50 years. Trees 2016, 30, 281–294. [Google Scholar] [CrossRef]
- Stegen, J.C.; Swenson, N.G.; Enquist, B.J.; White, E.P.; Phillips, O.L.; Jorgensen, P.M.; Weiser, M.D.; Mendoza, A.M.; Vargas, P.N. Variation in above-ground forest biomass across broad climatic gradients. Glob. Ecol. Biogeogr. 2011, 20, 744–754. [Google Scholar] [CrossRef]
- Fu, L.; Lei, X.; Hu, Z.; Zeng, W.; Tang, S.; Marshall, P.; Cao, L.; Song, X.; Li, Y.; Liang, J. Integrating regional climate change into allometric equations for estimating tree aboveground biomass of Masson pine in China. Ann. For. Sci. 2017, 74, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Duo, H.; Lei, X.; Chen, X.; Wang, X.; Pu, Y.; Zou, W. Individual tree biomass equations and growth models sensitive to climate variables for Larix spp. in China. Eur. J. For. Res. 2017, 136, 233–249. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Zeller, L.; Liang, J.; Pretzsch, H. Tree species richness enhances stand productivity while stand structure can have opposite effects, based on forest inventory data from Germany and the United States of America. For. Ecosyst. 2018, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Usoltsev, V.A. On the application of regression analysis in forestry problems. Lesn. Taksatsiya Lesoustroistvo 2004, 1, 49–55. [Google Scholar]
- Clutter, J.L. Compatible growth and yield models for loblolly pine. For. Sci. 1963, 9, 354–371. [Google Scholar]
- Furnival, G.M.; Wilson, R.W. Systems of equations for predicting forest growth and yield. Stat. Ecol. 1971, 3, 43–55. [Google Scholar]
- Warren, W.G. Record of preplanned and spontaneous discussions concerning the paper by Furnival G.M. and Wilson R.W. “Systems of equations for predicting forest growth and yield”. Stat. Ecol. 1971, 3, 56–57. [Google Scholar]
- Borders, B.E. Systems of equations in forest stand modelling. For. Sci. 1989, 35, 548–556. [Google Scholar]
- Draper, N.; Smith, H. Prikladnoy Regressionnyi Analiz; Statistika: Moscow, Russia, 1973; p. 362. [Google Scholar]
- Usoltsev, V.A.; Koltunova, A.I.; Kajimoto, T.; Osawa, A.; Koike, T. Geographical gradients of annual biomass production from larch forests in Northern Eurasia. Eurasian J. For. Res. 2002, 5, 55–62. [Google Scholar]
- Fu, L.Y.; Zeng, W.S.; Tang, S.Z.; Sharma, R.P.; Li, H.K. Using linear mixed model and dummy variable model approaches to construct compatible single-tree biomass equations at different scales—A case study for Masson pine in Southern China. J. For. Sci. 2012, 58, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Sun, W.; Wang, G. A climate-sensitive aboveground biomass model for three larch species in northeastern and northern China. Trees 2017, 31, 557–573. [Google Scholar] [CrossRef]
- Zheng, D.; Zeng, W. Using dummy variable approach to construct segmented aboveground biomass models for larch and oak in northeastern China. J. Beijing For. Univ. 2013, 35, 34–36. [Google Scholar]
- Zeng, W.S. Developing tree biomass models for eight major tree species in China. In Biomass Volume Estimation and Valorization for Energy; Tumuluri, J.S., Ed.; Intech Publisher: Beijing, China, 2017; pp. 3–21. [Google Scholar]
- Gernandt, D.S.; López, G.G.; García, S.O.; Liston, A. Phylogeny and classification of Pinus. Taxon 2005, 54, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Bobrov, E.G. Forest-Forming Conifers of the USSR; Nauka Publishing: Leningrad, Russia, 1978; p. 188. [Google Scholar]
- Mamaev, S.A. Types of Coniferous in the Urals and Their Use in Planting of Greenery; Institute of Plants and Animals Ecology of the Ural Scientific Centre of the Academy of Sciences of the USSR: Sverdlovsk, Russia, 1983; p. 111. [Google Scholar]
- Sukachev, V.N. Dendrology with the Fundamentals of Forest of Geobotany, 2nd ed; Roslestekhizdat Publishing: Leningrad, Russia, 1938; p. 576. [Google Scholar]
- Usoltsev, V.A.; Shobairi, S.O.R.; Tsepordey, I.S.; Chasovskikh, V.P. Modeling the additive structure of stand biomass equations in climatic gradients of Eurasia. Environ. Qual. Manag. 2018, 28, 55–61. [Google Scholar] [CrossRef]
- Paquette, A.; Vayreda, J.; Coll, L.; Messier, C.; Retana, J. Climate change could negate positive tree diversity effects on forest productivity: A study across five climate types in Spain and Canada. Ecosystems 2018, 21, 960–970. [Google Scholar] [CrossRef] [Green Version]
- Usoltsev, V.A. Forest Biomass and Primary Production Database for Eurasia: Digital Version, 3rd ed.; Ural State Forest Engineering University: Yekaterinburg, Russia, 2020. [Google Scholar] [CrossRef]
- World Weather Maps. 2007. Available online: https://www.mapsofworld.com/referrals/weather/ (accessed on 17 January 2018).
- Berner, L.T.; Beck, P.S.A.; Bunn, A.G.; Goetz, S.J. Plant response to climate change along the forest-tundra ecotone in northeastern Siberia. Glob. Chang. Biol. 2013, 19, 3449–3462. [Google Scholar] [CrossRef]
- Bouriaud, O.; Bréda, N.; Dupouey, J.-L.; Granier, A. Is ring width a reliable proxy for stem-biomass increment? A case study in European beech. Can. J. For. Res. 2005, 35, 2920–2933. [Google Scholar] [CrossRef]
- Khan, D.; Muneer, M.A.; Nisa, Z.-U.; Shah, S.; Saeed, S.; Uddin, S.; Munir, M.Z.; Lushuang, G.; Huang, H. Effect of climatic factors on stem biomass and carbon stock of Larix gmelinii and Betula platyphylla in Daxing’anling Mountain of Inner Mongolia, China. Adv. Meteorol. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zubairov, B.; Heußner, K.-U.; Schröder, H. Searching for the best correlation between climate and tree rings in the Trans-Ili Alatau, Kazakhstan. Dendrobiology 2018, 79, 119–130. [Google Scholar] [CrossRef]
- Bocharnikov, M.V. Bioclimate as a factor of the phytocenotic diversity’s formation of the mountain taiga forests in the Stanovoy Highland. In Proceedings of the Forest Ecosystems of Boreal Zone: Biodiversity, Bioeconomy, Ecological Risks, Krasnoyarsk, Russia, 26–31 August 2019; pp. 57–59. [Google Scholar]
- Morley, J.W.; Batt, R.D.; Pinsky, M.L. Marine assemblages respond rapidly to winter climate variability. Glob. Chang. Biol. 2017, 23, 2590–2601. [Google Scholar] [CrossRef]
- Emanuel, W.R.; Shugart, H.H.; Stevenson, M.P. Climate change and the broad scale distribution of terrestrial ecosystem complexes. Clim. Chang. 1985, 7, 29–43. [Google Scholar] [CrossRef]
- Folland, C.K.; Palmer, T.N.; Parker, D.E. Climate change 2001: The scientific basis. In Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., Van der Linden, P.J., Dai, X., Maskell, K., Johnson, C.A., Eds.; Cambridge University Press: Cambridge, UK, 2001; p. 1032. [Google Scholar]
- Laing, J.; Binyamin, J. Climate change effect on winter temperature and precipitation of Yellowknife, Northwest Territories, Canada from 1943 to 2011. Am. J. Clim. Chang. 2013, 2, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.-M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J.; et al. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usoltsev, V.A. Some methodological and conceptual uncertainties in estimating the in-come component of the forest carbon cycle. Russ. J. Ecol. 2007, 38, 1–10. [Google Scholar] [CrossRef]
- Baskerville, G.L. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox. Silva Fenn. 2002, 36, 703–743. [Google Scholar] [CrossRef]
- Schaphoff, S.; Reyer, C.P.; Schepaschenko, D.; Gerten, D.; Shvidenko, A. Tamm review: Observed and projected climate change impacts on Russia’s forests and its carbon balance. For. Ecol. Manag. 2016, 361, 432–444. [Google Scholar] [CrossRef] [Green Version]
- Spathelf, P.; Stanturf, J.; Kleine, M.; Jandl, R.; Chiatante, D.; Bolte, A. Adaptive measures: Integrating adaptive forest management and forest landscape restoration. Ann. For. Sci. 2018, 75, 55. [Google Scholar] [CrossRef] [Green Version]
- Vasseur, F.; Exposito-Alonso, M.; Ayala-Garay, O.J.; Wang, G.; Enquist, B.J.; Vile, D.; Violle, C.; Weigel, D. Adaptive diversification of growth allometry in the plant Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, 3416–3421. [Google Scholar] [CrossRef] [Green Version]
- DeLeo, V.L.; Menge, D.N.; Hanks, E.M.; Juenger, T.E.; Lasky, J.R. Effects of two centuries of global environmental variation on phenology and physiology of Arabidopsis thaliana. Glob. Chang. Biol. 2020, 26, 523–538. [Google Scholar] [CrossRef]
- Denney, D.A.; Anderson, J.T. Natural history collections document biological responses to climate change: A commentary on DeLeo et al. (2019), Effects of two centuries of global environmental variation on phenology and physiology of Arabidopsis thaliana. Glob. Chang. Biol. 2020, 26, 340–342. [Google Scholar] [CrossRef] [Green Version]
- Liebig, J. Organic chemistry in its application to agriculture and physiology Braunschweig. In German Text Archive; 1840; Available online: http://www.deutschestextarchiv.de/liebig_agricultur_1840 (accessed on 22 May 2020).
- Shelford, V.E.; Metcalf, C. Animal communities in temperate America, as illustrated in the Chicago region: A study in animal ecology. Geogr. Soc. Chic. Bull. 1913, 13, 362. [Google Scholar]
- Odum, E.P. Fundamentals of Ecology, 3rd ed.; W.B. Saunders Company: London, UK, 1971. [Google Scholar]
- Du, J.; Watts, J.D.; Jiang, L.; Lu, H.; Cheng, X.; Duguay, C.; Farina, M.; Qiu, Y.; Kim, Y.; Kimball, J.S.; et al. Remote Sensing of Environmental Changes in Cold Regions: Methods, Achievements and Challenges. Remote Sens. 2019, 11, 1952. [Google Scholar] [CrossRef] [Green Version]
- Rößiger, G.; Kulla, L.; Bošeľa, M. Changes in growth caused by climate change and other limiting factors in time affect the optimal equilibrium of close-to-nature forest management. Cent. Eur. For. J. 2019, 65, 180–190. [Google Scholar] [CrossRef]
- Nowakowska, J.A.; Hsiang, T.; Patynek, P.; Stereńczak, K.; Olejarski, I.; Oszako, T. Health assessment and genetic structure of monumental Norway spruce trees during a bark beetle (Ips typographus L.) outbreak in the Białowieża Forest District, Poland. Forests 2020, 11, 647. [Google Scholar] [CrossRef]
- Eggers, J.; Lindner, M.; Zudin, S.; Zaehle, S.; Liski, J. Impact of changing wood de-mand, climate and land use on European forest resources and carbon stocks during the 21st century. Glob. Chang. Biol. 2008, 14, 2288–2303. [Google Scholar] [CrossRef]
- Shuman, J.K.; Shugart, H.H. Evaluating the sensitivity of Eurasian forest biomass to climate change using a dynamic vegetation model. Environ. Res. Lett. 2009, 4, 1–7. [Google Scholar] [CrossRef]
- Schulze, E.D. The carbon and nitrogen cycle of forest ecosystems. In Carbon and Nutrient Cycling in European Forest Ecosystems; Schulze, E.-D., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2000; Volume 142, pp. 3–13. [Google Scholar]
- Anderegg, W.R.L.; Anderegg, L.D.L.; Kerr, K.L.; Trugman, A.T. Widespread drought-induced tree mortality at dry range edges indicates that climate stress exceeds species’ compensating mechanisms. Glob. Chang. Biol. 2019, 25, 3793–3802. [Google Scholar] [CrossRef]
- Zhou, W.N. Phytomass and Features of Nitrogen and Ash Elements Cycling in the Ecosystems of the Cherry Oak of Various Phenological Varieties in the Conditions of the Central Chernozem Region. Ph.D. Thesis, Voronezh Forest Engineering Institute, Voronezh, Russia, 1992; p. 23. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ln(Y) (1) | a0 (2) | lnA | lnV | lnN | (lnA)·(lnN) | ln(Tm + 40) | lnPRm | [ln(Tm + 40)]·(lnPRm) | adjR2 (3) | SE (4) |
|---|---|---|---|---|---|---|---|---|---|---|
| Pinus | ||||||||||
| ln(N) | 2.8168− | −1.0696 | - | - | - | 1.9165 | 0.5011 | -0.3577 | 0.566 | 0.72 |
| ln(V) | 16.4304 | 0.7200 | - | −0.7996 | 0.2065 | −3.3579 | −2.5007 | 0.6225 | 0.472 | 0.69 |
| ln(Pt) | 1.8338 | 0.0782 | 0.8291 | 0.0367 | −0.0031 | −0.5645 | −0.3541 | 0.1157 | 0.944 | 0.16 |
| ln(Pa) | 3.4809 | 0.0441 | 0.8221 | 0.0924 | −0.0240 | −1.1030 | −0.5944 | 0.1931 | 0.952 | 0.17 |
| ln(Pr) | −12.6886 | 0.1991 | 0.7013 | −0.1314 | 0.0597 | 3.2874 | 1.7271 | −0.5006 | 0.770 | 0.33 |
| ln(Ps) | 0.2311 | 0.1283 | 0.9346 | −0.0173 | 0.0137 | −0.4483 | −0.2603 | 0.0901 | 0.966 | 0.17 |
| ln(Pf) | 11.8492 | −0.3495 | 0.4313 | 0.1311 | −0.0289 | −3.6140 | −1.6841 | 0.5555 | 0.424 | 0.36 |
| ln(Pb) | 9.6761 | −0.1607 | 0.5734 | −0.0327 | −0.0154 | −3.3278 | −1.6096 | 0.5548 | 0.650 | 0.39 |
| Picea | ||||||||||
| ln(N) | −11.5190 | −1.0091 | - | - | - | 5.2191 | 3.0637 | −0.9733 | 0.569 | 0.61 |
| ln(V) | −11.2252 | 0.8455 | - | −1.1457 | 0.2699 | −2.9798 | −2.4759 | 0.7765 | 0.642 | 0.59 |
| ln(Pt) | 0.0180 | −0.0646 | 0.8128 | −0.0875 | 0.0208 | 0.6708 | 0.1668 | −0.1113 | 0.970 | 0.19 |
| ln(Pa) | −1.2565 | −0.0524 | 0.8145 | −0.0796 | 0.0224 | 0.8107 | 0.2335 | −0.1077 | 0.972 | 0.17 |
| ln(Pr) | 4.1101 | −0.0090 | 0.7970 | −0.0973 | 0.0296 | −0.7176 | −0.6121 | 0.0666 | 0.884 | 0.39 |
| ln(Ps) | 0.3868 | 0.0371 | 0.9481 | −0.1493 | 0.0393 | −0.0652 | −0.2143 | 0.0208 | 0.984 | 0.15 |
| ln(Pf) | −10.8564 | −0.3423 | 0.5893 | −0.2302 | 0.1047 | 3.9064 | 1.8125 | −0.6126 | 0.669 | 0.42 |
| ln(Pb) | −11.3469 | −0.1765 | 0.6888 | −0.2263 | 0.0903 | 3.7022 | 1.7557 | −0.5780 | 0.785 | 0.40 |
| ln(Y) | Independent Variables 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| lnA(I) | lnV(II) | lnN(III) | (lnA)·(lnN) (IV) | I + II + III + IV | ln(Tm + M) (V) | lnPRm (VI) | [ln(Tm + M)]·(lnPRm) (VII) | V + VI + VII | |
| Pinus | |||||||||
| ln(Pt) | 5.5 | 88.8 | 1.1 | 0.3 | 95.7 | 1.3 | 1.4 | 1.6 | 4.3 |
| ln(Pa) | 2.7 | 84.4 | 2.9 | 2.8 | 92.8 | 2.3 | 2.3 | 2.6 | 7.2 |
| ln(Pr) | 11.4 | 62.4 | 3.2 | 5.5 | 82.5 | 6.0 | 5.7 | 5.8 | 17.5 |
| ln(Ps) | 7.1 | 88.0 | 0.5 | 1.5 | 97.1 | 0.9 | 0.9 | 1.1 | 2.9 |
| ln(Pf) | 22.4 | 47.3 | 4.3 | 3.6 | 77.6 | 7.9 | 6.8 | 7.7 | 22.4 |
| ln(Pb) | 10.5 | 64.1 | 1.1 | 2.0 | 77.7 | 7.6 | 6.7 | 8.0 | 22.3 |
| X ± σ 1 | 9.9 ± 6.9 | 72.5 ± 17.1 | 2.2 ± 1.5 | 2.6 ± 1.8 | 87.2 ± 9.0 | 4.3 ± 3.2 | 4.0 ± 2.7 | 4.5 ± 3.1 | 12.8 ± 9.0 |
| Picea | |||||||||
| ln(Pt) | 3.3 | 89.7 | 2.1 | 2.0 | 97.1 | 1.2 | 0.5 | 1.2 | 2.9 |
| ln(Pa) | 2.8 | 89.2 | 2.2 | 2.4 | 96.6 | 1.4 | 0.8 | 1.2 | 3.4 |
| ln(Pr) | 0.5 | 90.4 | 2.3 | 2.9 | 96.1 | 1.3 | 1.9 | 0.7 | 3.9 |
| ln(Ps) | 1.7 | 90.2 | 3.5 | 3.7 | 99.1 | 0.1 | 0.6 | 0.2 | 0.9 |
| ln(Pf) | 15.1 | 53.9 | 5.2 | 9.5 | 83.7 | 5.8 | 4.9 | 5.6 | 16.3 |
| ln(Pb) | 7.8 | 63.4 | 5.2 | 8.2 | 84.6 | 5.5 | 4.8 | 5.3 | 15.4 |
| X ± σ | 5.2 ± 5.4 | 79.5 ± 16.4 | 3.4 ± 1.5 | 4.8 ± 3.2 | 92.9 ± 6.8 | 2.6 ± 2.4 | 2.3 ± 2.1 | 2.4 ± 2.4 | 7.1 ± 6.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usoltsev, V.A.; Lin, H.; Shobairi, S.O.R.; Tsepordey, I.S.; Ye, Z. Are There Differences in the Reaction of the Light-Tolerant Subgenus Pinus spp. Biomass to Climate Change as Compared to Light-Intolerant Genus Picea spp.? Plants 2020, 9, 1255. https://doi.org/10.3390/plants9101255
Usoltsev VA, Lin H, Shobairi SOR, Tsepordey IS, Ye Z. Are There Differences in the Reaction of the Light-Tolerant Subgenus Pinus spp. Biomass to Climate Change as Compared to Light-Intolerant Genus Picea spp.? Plants. 2020; 9(10):1255. https://doi.org/10.3390/plants9101255
Chicago/Turabian StyleUsoltsev, Vladimir A., Hui Lin, Seyed Omid Reza Shobairi, Ivan S. Tsepordey, and Zilin Ye. 2020. "Are There Differences in the Reaction of the Light-Tolerant Subgenus Pinus spp. Biomass to Climate Change as Compared to Light-Intolerant Genus Picea spp.?" Plants 9, no. 10: 1255. https://doi.org/10.3390/plants9101255