
Diversity of Leaf Fungal Endophytes from Two Coffea arabica Varieties and Antagonism towards Coffee Leaf Rust
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Sampling, Isolation, and Endophyte Identification
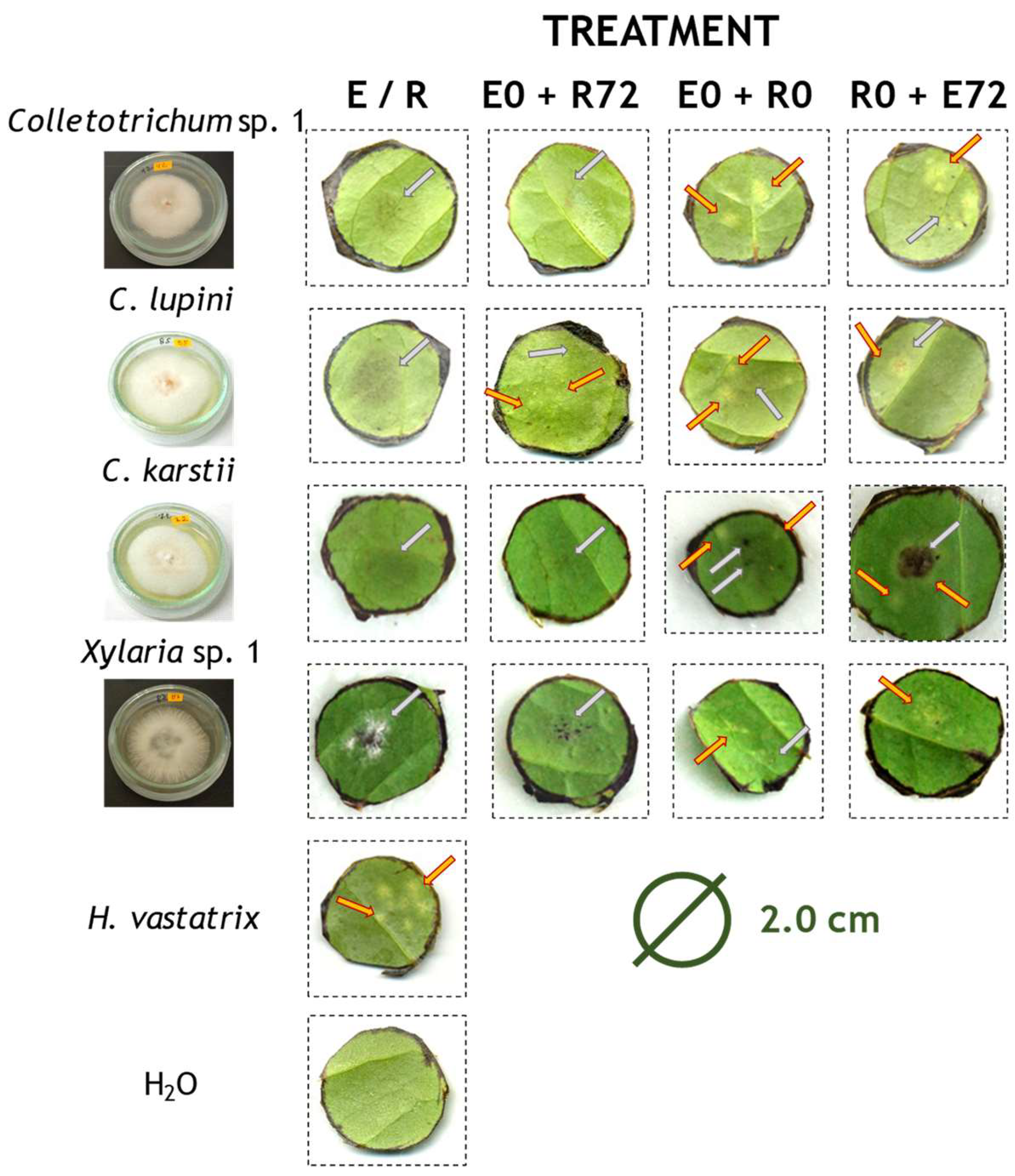
2.2. Cross-Inoculation Essays in Leaf Discs
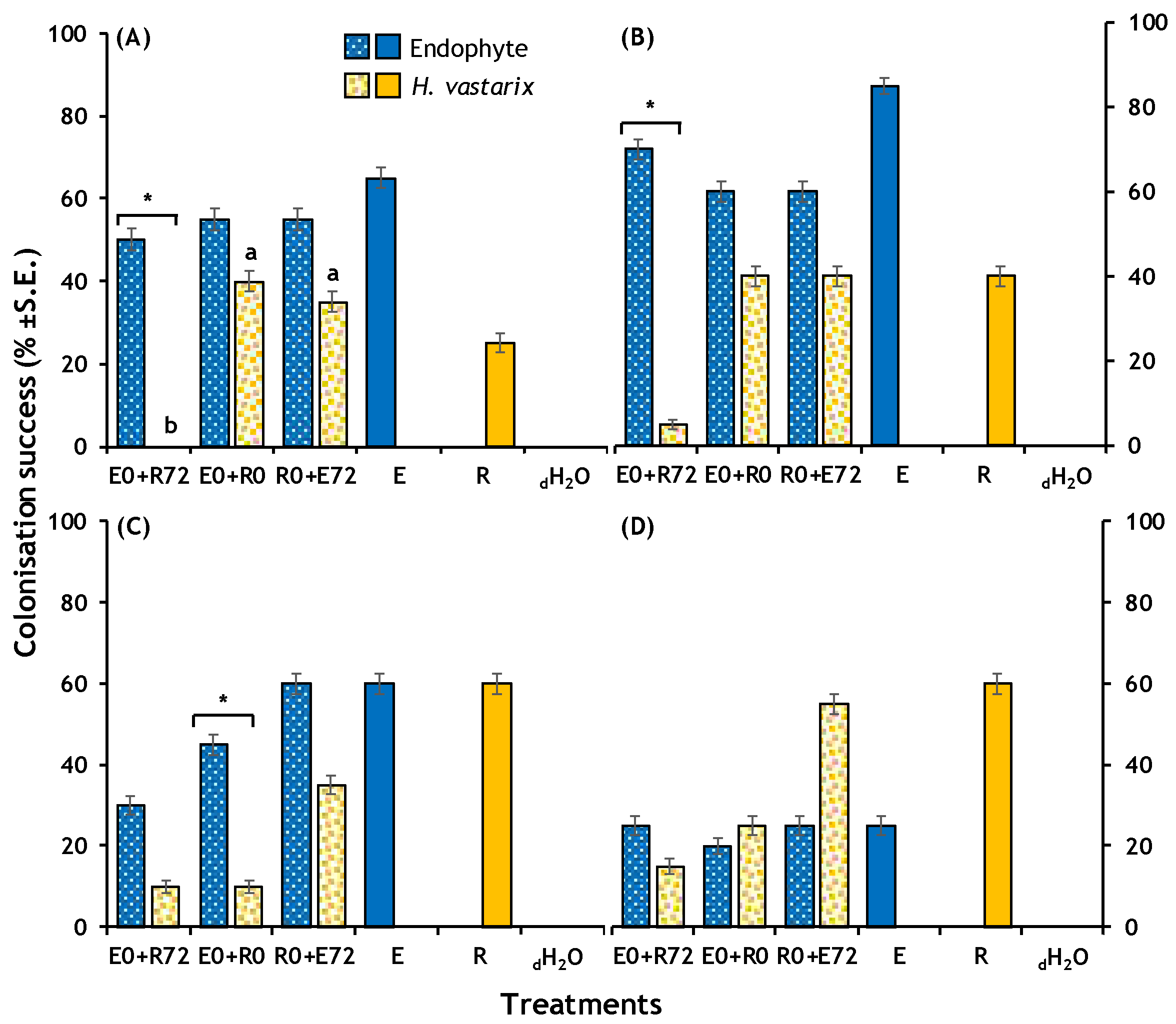
2.2.1. Colonisation Success
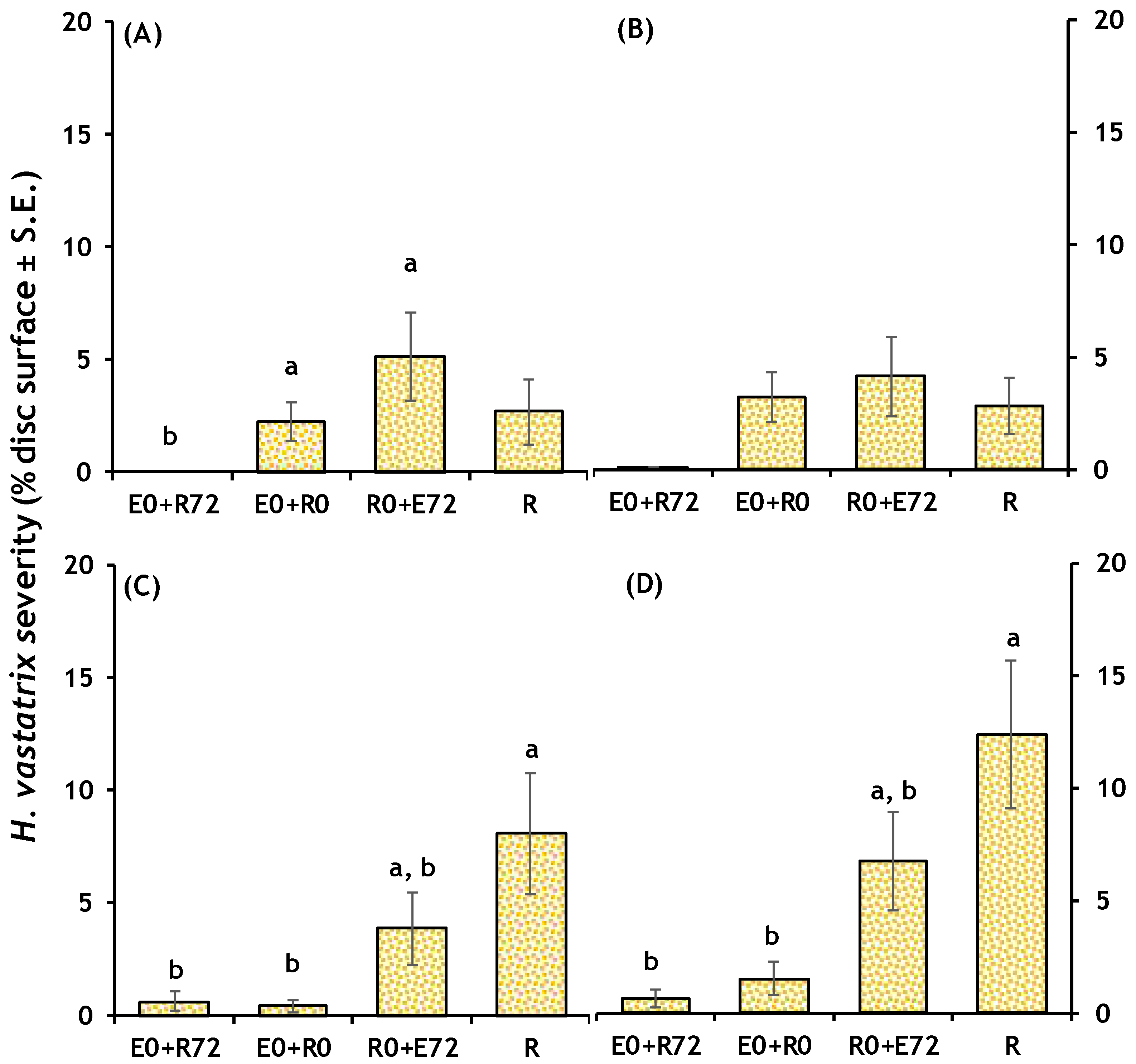
2.2.2. CLR Severity in Leaf Discs Infection
3. Discussion
4. Materials and Methods
4.1. Sampling, Isolation, and Endophyte Identification
4.2. Endophyte Biodiversity
4.3. Cross-Inoculation Assays in Leaf Discs
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Statistical Pocketbook: Coffee 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; 194p. [Google Scholar]
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The impact of climate change and variability on coffee production: A systematic review. Clim. Chang. 2019, 156, 609–630. [Google Scholar] [CrossRef]
- Magrach, A.; Ghazoul, J. Climate and pest-driven geographic shifts in global coffee production: Implications for forest cover, biodiversity and carbon storage. PLoS ONE 2015, 10, e0133071. [Google Scholar] [CrossRef] [PubMed]
- Alègre, G. Climats et caféiers d’Arabie. L’Agronomie Trop. 1959, 14, 23–58. [Google Scholar]
- Descroix, F.; Snoeck, J. Environmental factors suitable for coffee cultivation. In Coffee: Growing, Processing, Sustainable Production; Wintgens, J.N., Ed.; Wiley-VCH GmbH & Co. KGaA: Weinheim, Germany, 2004; pp. 164–177. [Google Scholar]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle-Rivera, O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Clim. Chang. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Jimenez, J.G.P.; Montagnon, C.; Schilling, T. Multiclass classification of agro-ecological zones for Arabica coffee: An improved understanding of the impacts of climate change. PLoS ONE 2015, 10, e0140490. [Google Scholar] [CrossRef] [PubMed]
- Toniutti, L.; Breitler, J.-C.; Etienne, H.; Campa, C.; Doulbeau, S.; Urban, L.; Lambot, C.; Pinilla, J.-C.H.; Bertrand, B. Influence of environmental conditions and genetic background of Arabica coffee (C. arabica L.) on leaf rust (Hemileia vastatrix) pathogenesis. Front. Plant Sci. 2017, 8, 2025. [Google Scholar] [CrossRef]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Secur. 2015, 7, 303–321. [Google Scholar] [CrossRef]
- Bebber, D.P.; Castillo, Á.D.; Gurr, S.J. Modelling coffee leaf rust risk in Colombia with climate reanalysis data. Phil Trans. R. Soc. B 2016, 371, 20150458. [Google Scholar] [CrossRef]
- Brown, J.S.; Whan, J.H.; Kenny, M.K.; Merriman, P.R. The effect of coffee leaf rust on foliation and yield of coffee in Papua New Guinea. Crop Prot. 1995, 14, 589–592. [Google Scholar] [CrossRef]
- McCook, S. Global rust belt: Hemileia vastatrix and the ecological integration of world coffee production since 1850. J. Glob. Hist. 2006, 1, 177–195. [Google Scholar] [CrossRef]
- McCook, S.; Vandermeer, J. The big rust and the Red Queen: Long-term perspectives on coffee rust research. Phytopathology 2015, 105, 1164–1173. [Google Scholar] [CrossRef]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The Coffee Leaf Rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef]
- Helfer, S. Rust fungi and global change. New Phytol. 2013, 201, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Avelino, J.; Zelaya, H.; Merlo, A.; Pineda, A.; Ordoñez, M.; Savary, S. The intensity of a coffee rust epidemic is dependent on production situations. Ecol. Model. 2006, 197, 431–447. [Google Scholar] [CrossRef]
- Becker, S.; Kranz, J. Vergleichende Untersuchungen zur Verbreitung von Hemileia vastatrix in Kenia. Z. Pflanzenkrankh. Pflanzenschutz 1977, 84, 526–539. [Google Scholar]
- Lacava, P.T.; Bogas, A.C.; Nogeuira Cruz, F.P. Plant Growth Promotion and Biocontrol by Endophytic and Rhizospheric Microorganisms from the Tropics: A Review and Perspectives. Front. Sustain. Food Syst. 2022, 6, 796113. [Google Scholar] [CrossRef]
- Carroll, G. Fungal Endophytes in Stems and Leaves: From Latent Pathogen to Mutualistic Symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. PNAS 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Exploring the potentialities of beneficial endophytes for improved plant growth. Saudi J. Biol. Sci. 2020, 27, 3622–3633. [Google Scholar] [CrossRef]
- Hynes, R.K.; Boyetchko, S.M. Research initiatives in the art and science of biopesticide formulations. Soil Biol. Biochem. 2006, 38, 845–849. [Google Scholar] [CrossRef]
- Singh, D.P.; Singh, H.B.; Prabha, R. Plant-Microbe Interactions in Agro-Ecological Perspectives. Volume 1: Fundamental Mechanisms, Methods and Functions; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Azevedo, J.L. Endophytic fungi from Brazilian tropical hosts and their biotechnological applications. In Microbial Diversity and Biotechnology in Food Security; Kharwar, R.N., Upadhyay, R., Dubey, N., Raghuwanshi, R., Eds.; Springer: New Delhi, India, 2014; pp. 17–22. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, E. Development, registration and commercialization of microbial pesticides for plant protection. Int. Microbiol. 2003, 6, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Surnayarayanan, T.S.; Shaanker, R.U. Can fungal endophytes fast-track plant adaptations to climate change? Fungal Ecol. 2021, 50, 101039. [Google Scholar] [CrossRef]
- Haddad, F.; Saraiva, R.M.; Mizubuti, E.S.G.; Romeiro, R.S.; Maffia, L.A. Antifungal compounds as a mechanism to control Hemileia vastatrix by antagonistic bacteria. Trop. Plant Pathol. 2013, 38, 398–405. [Google Scholar] [CrossRef]
- Lu, L.; Karunarathna, S.C.; Hyde, K.D.; Suwannarach, N.; Elgorban, A.M.; Stephenson, S.L.; Al-Rejaie, S.; Jayawardena, R.S.; Tibpromma, S. Endophytic Fungi Associated with Coffee Leaves in China Exhibited In Vitro Antagonism against Fungal and Bacterial Pathogens. J. Fungi 2022, 8, 698. [Google Scholar] [CrossRef]
- Silva, H.; Tozzi, J.; Terrasan, C.; Bettiol, W. Endophytic microorganisms from coffee tissues as plants growth promoters and biocontrol agents of coffee leaf rust. Biol. Control 2012, 63, 62–67. [Google Scholar] [CrossRef]
- Mejía, L.C.; Rojas, E.I.; Maynard, Z.; Van Bael, S.V.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Moonjely, S.; Barelli, L.; Bidochka, M.J. Insect Pathogenic Fungi as Endophytes. Adv. Genet. 2016, 94, 107–135. [Google Scholar] [CrossRef]
- Vandermeer, J.; Perfecto, I.; Liere, H. Evidence for hyperparasitism of coffee rust (Hemileia vastatrix) by the entomogenous fungus, Lecanicillium lecanii, through a complex ecological web. Plant Pathol. 2009, 58, 636–641. [Google Scholar] [CrossRef]
- Shiomi, H.F.; Alves Silva, H.S.; Soares de Melo, I.; Vieira Nunes, F.; Bettiol, W. Bioprospecting endophytic bacteria for biological control of coffee leaf rust. Sci. Agric. 2006, 63, 32–39. [Google Scholar] [CrossRef]
- James, T.Y.; Marino, J.A.; Perfecto, I.; Vandermeer, J. Identification of putative coffee rust mycoparasites via single-molecule DNA sequencing of infected pustules. Appl. Environ. Microbiol. 2016, 82, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.J.V.; Souza, R.G.; Lima, T.E.F.; Cavalcanti, M.A.Q. Endophytic fungal diversity in coffee leaves (Coffea arabica) cultivated using organic and conventional crop management systems. Mycosphere 2014, 5, 523–530. [Google Scholar] [CrossRef]
- Santamaría, J.; Bayman, P. Fungal epiphytes and endophytes of coffee leaves (Coffea arabica). Microb. Ecol. 2005, 50, 1–8. [Google Scholar] [CrossRef]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal endophyte diversity in coffee plants from Colombia, Hawai’i, Mexico, and Puerto Rico. Fungal Ecol. 2010, 3, 122–138. [Google Scholar] [CrossRef]
- Bongiorno, V.A.; Rhoden, S.A.; Garcia, A.; Polonio, J.C.; Azevedo, J.L.; Pereira, J.O.; Pamphile, J.A. Genetic Diversity of Endophytic Fungi from Coffea arabica Cv. IAPAR-59 in Organic Crops. Ann. Microbiol. 2016, 66, 855–865. [Google Scholar] [CrossRef]
- Saucedo-García, A.; Anaya, A.L.; Espinosa-García, F.J.; González, M.C. Diversity and Communities of Foliar Endophytic Fungi from Different Agroecosystems of Coffea arabica L. in Two Regions of Veracruz, Mexico. PLoS ONE 2014, 9, e98454. [Google Scholar] [CrossRef] [PubMed]
- Alkemade, J.A.; Messmer, M.M.; Voegele, R.T.; Finckh, M.R.; Hohmann, P. Genetic diversity of Colletotrichum lupini and its virulence on white and Andean lupin. Sci. Rep. 2021, 11, 13547. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2010, 73, 37–113. [Google Scholar] [CrossRef]
- Hladki, A.I.; Romero, A.I. primeras citas del género Xylaria (ascomycota, Xylariaceae) para la república argentina. Darwiniana 2007, 45, 28–44. [Google Scholar]
- Macías-Rubalcava, M.L.; Sánchez-Fernández, R.E. Secondary metabolites of endophytic Xylaria species with potential applications in medicine and agriculture. World J. Microbiol. Biotechnol. 2017, 33, 15. [Google Scholar] [CrossRef]
- Monteiro, M.C.P.; Alves, N.M.; Queiroz, M.V.; Pinho, D.B.; Pereira, O.L.; Souza, S.M.C.; Cardoso, P.G. Antimicrobial activity of endophytic fungi from coffee plants. Biosci. J. Uberlândia 2017, 33, 381–389. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A. Fungal endophytes in green coffee seeds. Mycosystema 2008, 27, 75–84. [Google Scholar]
- Hoang, M.T.V.; Irinyi, L.; Chen, S.C.A.; Sorrell, T.C.; ISHAM Barcoding of Medical Fungi Working Group; Meyer, W. Dual DNA Barcoding for the molecular identification of the agents of invasive fungal infections. Front. Microbiol. 2019, 10, 1647. [Google Scholar] [CrossRef] [PubMed]
- Veloso, T.G.R.; da Silva, M.C.S.; Moreira, T.R.; da Luz, J.M.R.; Moreli, A.P.; Kasuya, M.C.M.; Pereira, L.L. Microbiomes associated with Coffea arabica and Coffea canephora in four different floristic domains of Brazil. Sci. Rep. 2023, 13, 18477. [Google Scholar] [CrossRef]
- Carrión, G.; Rico-Gray, V. Mycoparasites on the coffee rust in Mexico. Fungal Divers. 2002, 11, 49–60. [Google Scholar]
- Adame-Álvarez, R.M.; Mendiola-Soto, J.; Heil, M. Order of arrival shifts endophyte-pathogen interactions in bean from resistance induction to disease facilitation. FEMS Microbiol. Lett. 2014, 355, 100–107. [Google Scholar] [CrossRef]
- Xu, D.; Li, N.; Gu, Y.-Q.; Huang, J.; Hu, B.-S.; Zheng, J.-Y.; Hu, J.-W.; Du, Q. Endophytic fungus Colletotrichum sp. AP12 promotes growth physiology and andrographolide biosynthesis in Andrographis paniculata (Burm. f.) Nees. Front. Plant Sci. 2023, 14, 1166803. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef]
- Silva, M.d.C.; Guerra-Guimarães, L.; Diniz, I.; Loureiro, A.; Azinheira, H.; Pereira, A.P.; Tavares, S.; Batista, D.; Várzea, V. An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives. Agronomy 2022, 12, 326. [Google Scholar] [CrossRef]
- Priyashantha, A.K.H.; Dai, D.-Q.; Bhat, D.J.; Stephenson, S.L.; Promputtha, I.; Kaushik, P.; Tibpromma, S.; Karunarathna, S.C. Plant–Fungi Interactions: Where It Goes? Biology 2023, 12, 809. [Google Scholar] [CrossRef] [PubMed]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San José, Costa Rica, 1967; 149p. [Google Scholar]
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S. Fungal endophytes in dicotyledonous neotropical trees: Patterns of abundance and diversity. Mycol. Res. 2001, 105, 1502–1507. [Google Scholar] [CrossRef]
- Wilson, D. Endophyte: The Evolution of a Term, and Clarification of Its Use and Definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a Medium for Simple Extraction of DNA for PCR-Based Typing from Forensic Material. BioTechniques 1991, 10, 506–513. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Chao, A.; Shen, T.-J. Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample. Environ. Ecol. Stat. 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence–absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef]
- Lu, L.; Tibpromma, S.; Karunarathna, S.C.; Jayawardena, R.S.; Lumyong, S.; Xu, J.; Hyde, K.D. Comprehensive Review of Fungi on Coffee. Pathogens 2022, 11, 411. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Higgins, K.L.; Coley, P.D.; Kursar, T.A.; Arnold, A.E. Culturing and direct PCR suggest prevalent host generalism among diverse fungal endophytes of tropical forest grasses. Mycologia 2011, 103, 247–260. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L. Seasonally dynamic fungal communities in the Quercus macrocarpa phyllosphere differ between urban and nonurban environments. New Phytol. 2010, 186, 496–513. [Google Scholar] [CrossRef]
- Rojas-Jimenez, K.; Tamayo-Castillo, G. Fungal Endophytes and Their Bioactive Compounds in Tropical Forests of Costa Rica. In Neotropical Endophytic Fungi; Rosa, L.H., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 109–130. [Google Scholar]
- Pereira, J.S.; Costa, R.R.; Nagamoto, N.S.; Forti, L.C.; Pagnocca, F.C.; Rodrigues, A. Comparative analysis of fungal communities in colonies of two leaf-cutting ant species with different substratum preferences. Fungal Ecol. 2016, 21, 68–75. [Google Scholar] [CrossRef]
- Akinfala, T.O.; Houbraken, J.; Sulyok, M.; Adedeji, A.R.; Odebode, A.C.; Krska, R.; Ezekiel, C.N. Moulds and their secondary metabolites associated with the fermentation and storage of two cocoa bean hybrids in Nigeria. Int. J. Food Microbiol. 2020, 316, 108490. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Miadlikowska, J.; Higgins, K.L.; Sarvate, S.D.; Gugger, P.; Way, A.; Hofstetter, V.; Kauff, F.; Lutzoni, F. A phylogenetic estimation of trophic transition networks for ascomycetous fungi: Are lichens cradles of symbiotrophic fungal diversification? Syst. Biol. 2009, 58, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.90045598 (accessed on 27 December 2023).
- Martin, P.L.; Krawczyk, T.; Khodadadi, F.; Aćimović, S.G.; Peter, K.A. Bitter rot of apple in the mid-Atlantic United States: Causal species and evaluation of the impacts of regional weather patterns and cultivar susceptibility. Phytopathology 2021, 111, 966–981. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, Z.; Jiang, X.; Fang, L.; Qiao, M. Endophytic Colletotrichum species from aquatic plants in southwest China. J. Fungi 2022, 8, 87. [Google Scholar] [CrossRef]
- Silva, A.A.; Polonio, J.C.; de Oliveira, J.A.D.S.; Bulla, A.M.; Golias, H.C.; Polli, A.D.; Soares, L.C.; Vicentini, V.E.P.; de Oliveira, A.J.B.; Gonçalves, J.E.; et al. Multilocus sequence analysis of endophytic fungi from Justicia brandegeana with the culture-dependent method and their bioprospection for health field. S. Afr. J. Bot. 2020, 134, 359–368. [Google Scholar] [CrossRef]
- Douanla-Meli, C.; Unger, J.G. Phylogenetic study of the Colletotrichum species on imported citrus fruits uncovers a low diversity and a new species in the Colletotrichum gigasporum complex. Fungal Biol. 2017, 121, 858–868. [Google Scholar] [CrossRef]
- Higginbotham, S.J.; Arnold, A.E.; Ibañez, A.; Spadafora, C.; Coley, P.D.; Kursar, T.A. Bioactivity of fungal endophytes as a function of endophyte taxonomy and the taxonomy and distribution of their host plants. PLoS ONE 2013, 8, e73192. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Yang, J.H.; Woo, J.J.; Oh, S.O.; Hur, J.S. Diversity and distribution patterns of endolichenic fungi in Jeju Island, South Korea. Sustainability 2020, 12, 3769. [Google Scholar] [CrossRef]
- Thomas, D.C.; Vandegrift, R.; Ludden, A.; Carroll, G.C.; Roy, B.A. Spatial ecology of the fungal genus Xylaria in a tropical cloud forest. Biotropica 2016, 48, 381–393. [Google Scholar] [CrossRef]
- U'ren, J.M.; Dalling, J.W.; Gallery, R.E.; Maddison, D.R.; Davis, E.C.; Gibson, C.M.; Arnold, A.E. Diversity and evolutionary origins of fungi associated with seeds of a neotropical pioneer tree: A case study for analysing fungal environmental samples. Mycol. Res. 2009, 113, 432–449. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef]
- Del Olmo-Ruiz, M.; Arnold, A.E. Community structure of fern-affiliated endophytes in three neotropical forests. J. Trop. Ecol. 2016, 33, 60–73. [Google Scholar] [CrossRef]
- Gonzaga, L.L.; Costa, L.E.O.; Santos, T.T.; Araújo, E.F.; Queiroz, M.V. Endophytic fungi from the genus Colletotrichum are abundant in the Phaseolus vulgaris and have high genetic diversity. J. Appl. Microbiol. 2015, 118, 485–496. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Fournier, J.; Rogers, J.D.; Voglmayr, H. Phylogenetic and taxonomic revision of Lopadostoma. Persoonia-Mol. Phylogeny Evol. Fungi 2014, 32, 52–82. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Taxonomic Group | Sample ID (GenBank ID) | Closest Taxa (GenBank ID: % ID) | RF (%) in Caturra | RF (%) in Colombia |
|---|---|---|---|---|
| Ascomycota | ||||
| Dothideomycetes | ||||
| Incertae sedis | ||||
| Fungal endophyte sp. 1 | F112 (MK005037) | EU687041: 90.22 | 0.00 | 12.50 |
| Capnodiales; Cladosporiaceae | ||||
| Uncultured Cladosporiaceae | F129 ** | GQ517141: 86.88 | 12.50 | 0.00 |
| Pleosporales; Sporormiaceae | ||||
| Preussia pseudominima | F041 (MK005019) | KU204603: 99.24 | 0.00 | 12.50 |
| Preussia sp. | F134 (MK005045) | KR093940: 95.20 | 12.50 | 0.00 |
| Eurotiomycetes | ||||
| Eurotiales; Aspergillaceae | ||||
| Penicillium sp. | F096, F126 (MK005040) | KF498874/MN788117/MN788106: 91.12 | 12.50 | 12.50 |
| Trichocomaceae | ||||
| Talaromyces sp. | F022 (MK005060), F081, F099 | OM791640/MH934969/MT530189: 89.88 | 12.50 | 25.00 |
| Sordariomycetes | ||||
| Fungal endophyte sp. 2 | F065 (MK005023) | KT289540: 96.19 | 0.00 | 12.50 |
| Diaporthales; Diaporthaceae | ||||
| Diaporthe sp. | F118 (MK005038) | MF280391/OM975589: 95.34 | 12.50 | 0.00 |
| Glomerellales; Glomerellaceae | ||||
| Colletotrichum acutatum complex | ||||
| C. lupini | F072 (MK005027), F073, F075, F082 | MH178095: 99.14 | 0.00 | 25.00 |
| C. scovillei | F043 (MK017761), F045, F057 | LC488868: 99.15 | 0.00 | 12.50 |
| Colletotrichum sp. 1 | F042 (MK005049), F058 | MK005027 (F072): 91.53 MK005048 (F040)/MH865411/MH854629: 91.45 | 0.00 | 25.00 |
| Colletotrichum sp. 2 | F036, F039, F040 (MK005048), F046, F102 | EF687919: 92.65 | 0.00 | 50.00 |
| Colletotrichum sp. 3 | F127 (MK005055) | JQ894656: 91.74 | 12.50 | 0.00 |
| Colletotrichum sp. 4 | F026, F132 (MK005044) | ON329227: 97.95 | 25.00 | 0.00 |
| Colletotrichum sp. 5 | F113 (MK005058) | ON368204: 89.00 | 0.00 | 12.50 |
| C. boninense species complex | ||||
| Colletotrichum sp. 6 | F066 (MK005051), F080 (MK005052) | MK005052 (F080): 97.70/MN458530: 97.30/MT464454: 97.32 | 0.00 | 12.50 |
| Colletotrichum sp. 7 | F011 * (MK005046), F078, F084, F133 | OL842171: 90.40 | 12.50 | 37.50 |
| C. gloeosporioides species complex | ||||
| C. karstii | F059 (MK005022), F083 (MK005031), F085 (MK005032), F050 (MK005020), F079 (MK005030), F077 (MK005029), F095 (MK005035), F031, F068, F089, F098, F100 | OM436864: 96.95/MN842791: 97.66/OP445269: 98.14/KX578788: 98.80/KX578788: 98.80/OM436864: 98.51/OP782678: 93.77/KX578788/MK005029: 93.52 | 0.00 | 87.50 |
| Colletotrichum sp. 8 | F053 (MK005050), F094 | MK005055 (F127): 91.21/OK030873: 89.36 | 0.00 | 25.00 |
| Colletotrichum sp. 9 | F007 * (MK005009), F029 | OW988162: 94.84/KX069828: 94.66 | 0.00 | 12.50 |
| Glomerellales; Plectosphaerellaceae | ||||
| Musicillium sp. | F017 *, F019 * (MK005015), F103 | MK579179: 95.84 | 0.00 | 12.50 |
| Hypocreales; Clavicipitaceae | ||||
| Beauveria sp. | F117 | 12.50 | 0.00 | |
| Sordariales | ||||
| Fungal endophyte sp. 3 | F104 ** | KF435260: 86.97 | 0.00 | 12.50 |
| Sordariales; Chaetomiaceae | ||||
| Chaetomium sp. | F060, F074 (MK005028) | MF495440/KF435950/KF435726/KF435552/KF435385: 87.70 | 0.00 | 25.00 |
| Trichosphaeriales; Trichosphaeriaceae | ||||
| Nigrospora sp. 1 | F025 (MK005047), F030, F034 ** | MT123068: 91.94 | 12.50 | 25.00 |
| Nigrospora sp. 2 | F128 (MK005041) | MN341467: 89.74 | 12.50 | 0.00 |
| Xylariales; Hypoxylaceae | ||||
| Annulohypoxylon cf. stygium | F015 (MK005013) | KP133169: 98.75 | 12.50 | 0.00 |
| Hypoxylon sp. 1 | F071 (MK005026) | FJ612775: 96.97 | 0.00 | 12.50 |
| Xylariales; Xylariaceaea | ||||
| Fungal endophyte sp. 4 | F106 (MK005036) | EU687119: 97.13 | 0.00 | 12.50 |
| Fungal endophyte sp. 5 | F064 ** | KU747690/FJ612923: 75.39 | 0.00 | 12.50 |
| Anthostomella sp. | F121 (MK005059) | JQ754021: 91.14 | 12.50 | 0.00 |
| Lopadostoma sp. | F001 *, F002 * (MK005007) | KC774600: 91.11 | 12.50 | 0.00 |
| Nemania sp. | F125 (MK005039) | KF435731: 97.88; MF770851: 97.38 | 12.50 | 0.00 |
| Xylaria sp. 1 | F086 (MK005033), F035 (MK005017), F032 (MK005016), F087 (MK005034), F097 (MK005054), F131 (MK005043), F047, F048, F055 | KP133288: 96.92, 95.79, 95.07, 95.60, 93.68, and 95.18 | 12.50 | 87.50 |
| Xylaria sp. 2 | F008 * (MK005010), F021 | KF467102: 94.59 | 12.500 | 0.00 |
| Xylaria sp. 3 | F016 * (MK005014), F114 | MH003490: 96.34 | 0.00 | 12.50 |
| Xylaria sp. 4 | F067 (MK005024), F037 (MK005018), F010 *, F063 | KF435704: 98.54,/MK005018 (F037) 97.24 | 0.00 | 25.00 |
| Xylaria sp. 5 | F023 (MK005056) | MH003401: 92.02 | 12.50 | 0.00 |
| Xylaria sp. 6 | F130 (MK005042) | MT992054: 94.33 | 12.50 | 0.00 |
| Xylaria sp. 7 | F027 **, F044, F056 (MK005021), F061, F062, F076, F101 | JQ341084: 98.23 | 0.00 | 87.50 |
| Xylaria sp. 8 | F088 (MK005053), F012 *, F051, F054, F124 | MK334005/MK247857: 98.75 | 12.50 | 37.50 |
| Xylaria sp. 9 | F070 (MK005025) | KJ883611: 97.21 | 0.00 | 12.50 |
| Xylaria sp. 10 | F009 * (MK005011), F052 | MN833802/KT289626/KP13343: 94.22 | 0.00 | 12.50 |
| Basidiomycota | ||||
| Agaricomycetes | ||||
| Hymenochaetales; Schizoporaceae | ||||
| Xylodon sp. | F107 ** | OM891735: 87.3 | 0.00 | 12.50 |
| 44 taxonomic groups | 20 spp. | 30 spp. | ||
| Caturra (± C.I.) | Colombia (± C.I.) | |
|---|---|---|
| Taxa | 20 | 30 |
| Individuals | 21 | 62 |
| Simpson’s D | 0.005 ± 0.029 | 0.043 ± 0.013 |
| Unbiased Shannon’s HCS | 7.955 ± 1.120 | 3.491 ± 0.100 |
| Margalef’s K | 6.241 ± 0.655 | 7.027 ± 0.127 |
| Whitakker’s β-diversity | 0.76 | |
| βsim | 0.70 | |
| Pearson’s χ2 | d.f. | p-Value | |
|---|---|---|---|
| Colletotrichum sp. 1 (F042) | 0.134 | 2 | 0.935 |
| CLR | 10.133 | 2 | 0.006 |
| C. lupini (F072) | 0.574 | 2 | 0.750 |
| CLR | 8.044 | 2 | 0.018 |
| C. karstii (F085) | 3.636 | 2 | 0.162 |
| CLR | 5.566 | 2 | 0.062 |
| Xylaria sp. 1 (F131) | 0.186 | 2 | 0.911 |
| CLR | 8.010 | 2 | 0.018 |
| N | H | d.f. | p-Value | |
|---|---|---|---|---|
| Treatment | 320 | 36.790 | 3 | <0.001 |
| Colletotrichum sp. 1 × Treatment | 80 | 9.531 | 3 | 0.023 |
| C. lupini × Treatment | 80 | 8.969 | 3 | 0.030 |
| C. karstii × Treatment | 80 | 17.330 | 3 | 0.001 |
| Xylaria sp. 1 × Treatment | 80 | 16.121 | 3 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poma-Angamarca, R.A.; Rojas, J.R.; Sánchez-Rodríguez, A.; Ruiz-González, M.X. Diversity of Leaf Fungal Endophytes from Two Coffea arabica Varieties and Antagonism towards Coffee Leaf Rust. Plants 2024, 13, 814. https://doi.org/10.3390/plants13060814
Poma-Angamarca RA, Rojas JR, Sánchez-Rodríguez A, Ruiz-González MX. Diversity of Leaf Fungal Endophytes from Two Coffea arabica Varieties and Antagonism towards Coffee Leaf Rust. Plants. 2024; 13(6):814. https://doi.org/10.3390/plants13060814
Chicago/Turabian StylePoma-Angamarca, Ruth A., Jacqueline R. Rojas, Aminael Sánchez-Rodríguez, and Mario X. Ruiz-González. 2024. "Diversity of Leaf Fungal Endophytes from Two Coffea arabica Varieties and Antagonism towards Coffee Leaf Rust" Plants 13, no. 6: 814. https://doi.org/10.3390/plants13060814