
Plant Essential Oils: Dual Action of Toxicity and Egg-Laying Inhibition on Tetranychus urticae (Acari: Tetranychidae), Unveiling Their Potential as Botanical Pesticides
 , and
, and
Abstract
:1. Introduction
2. Results
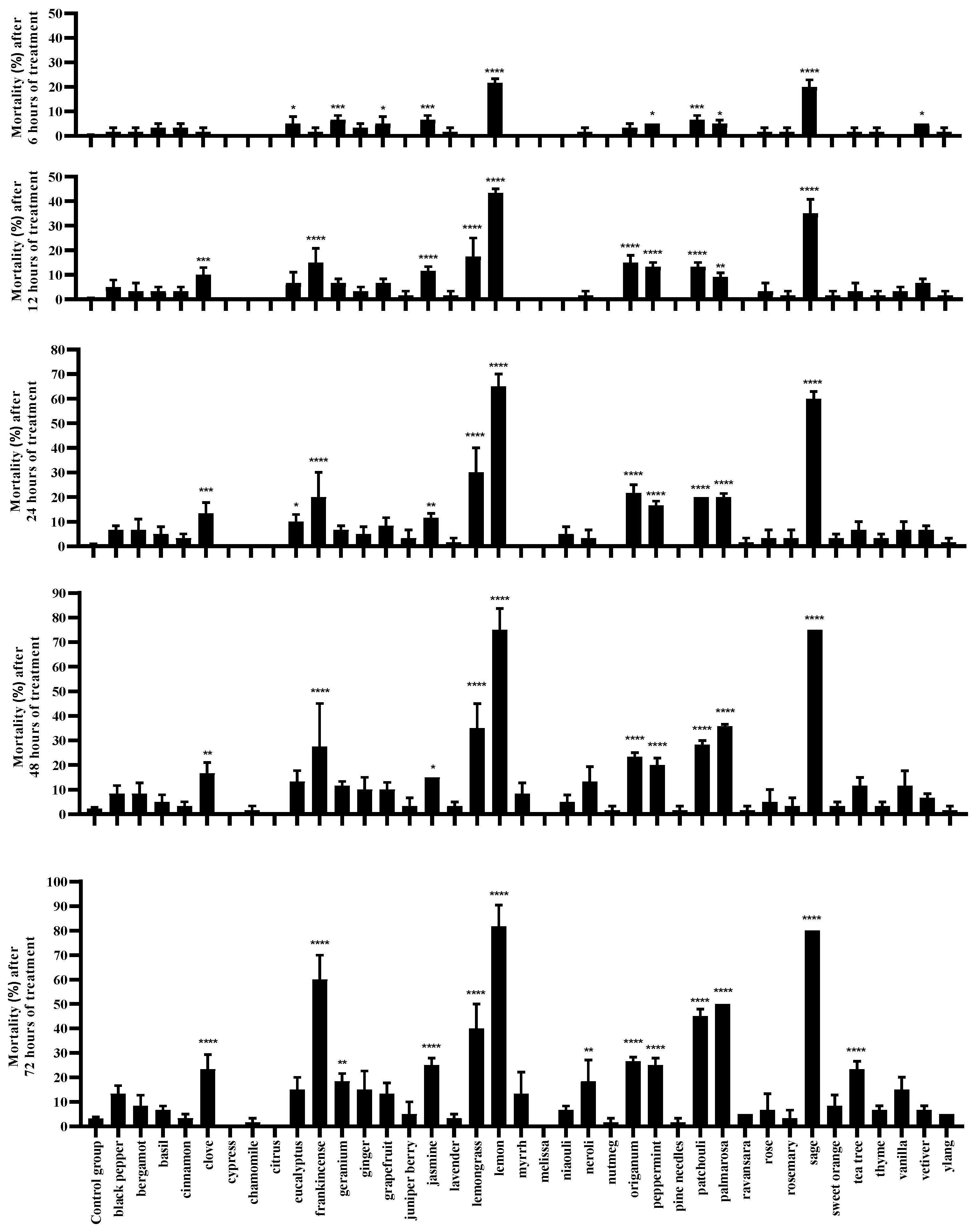
2.1. Preliminary Screening of Acaricidal Activity of EOs against T. urticae
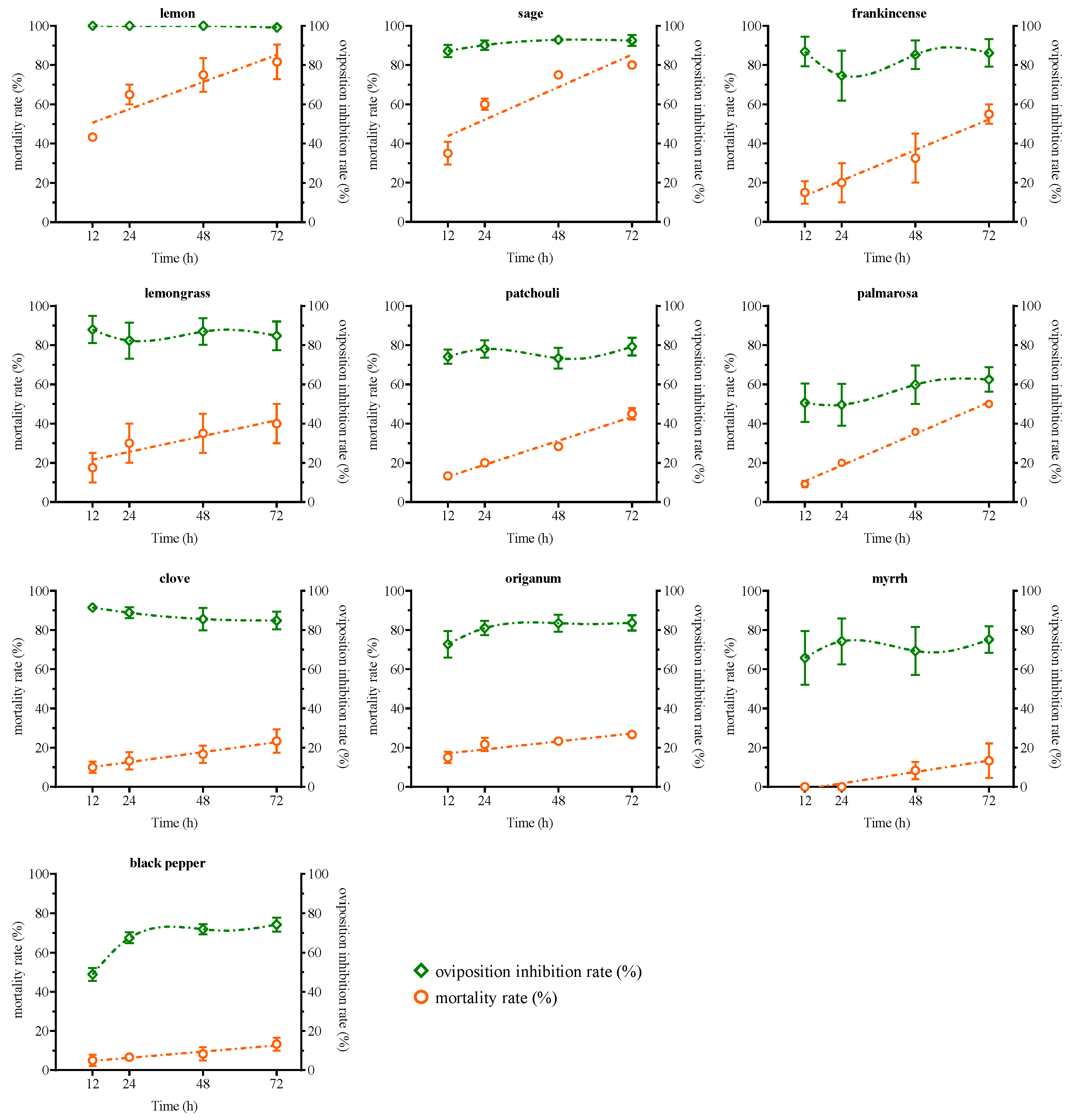
2.2. Toxicity of Seven Selected EOs on T. urticae
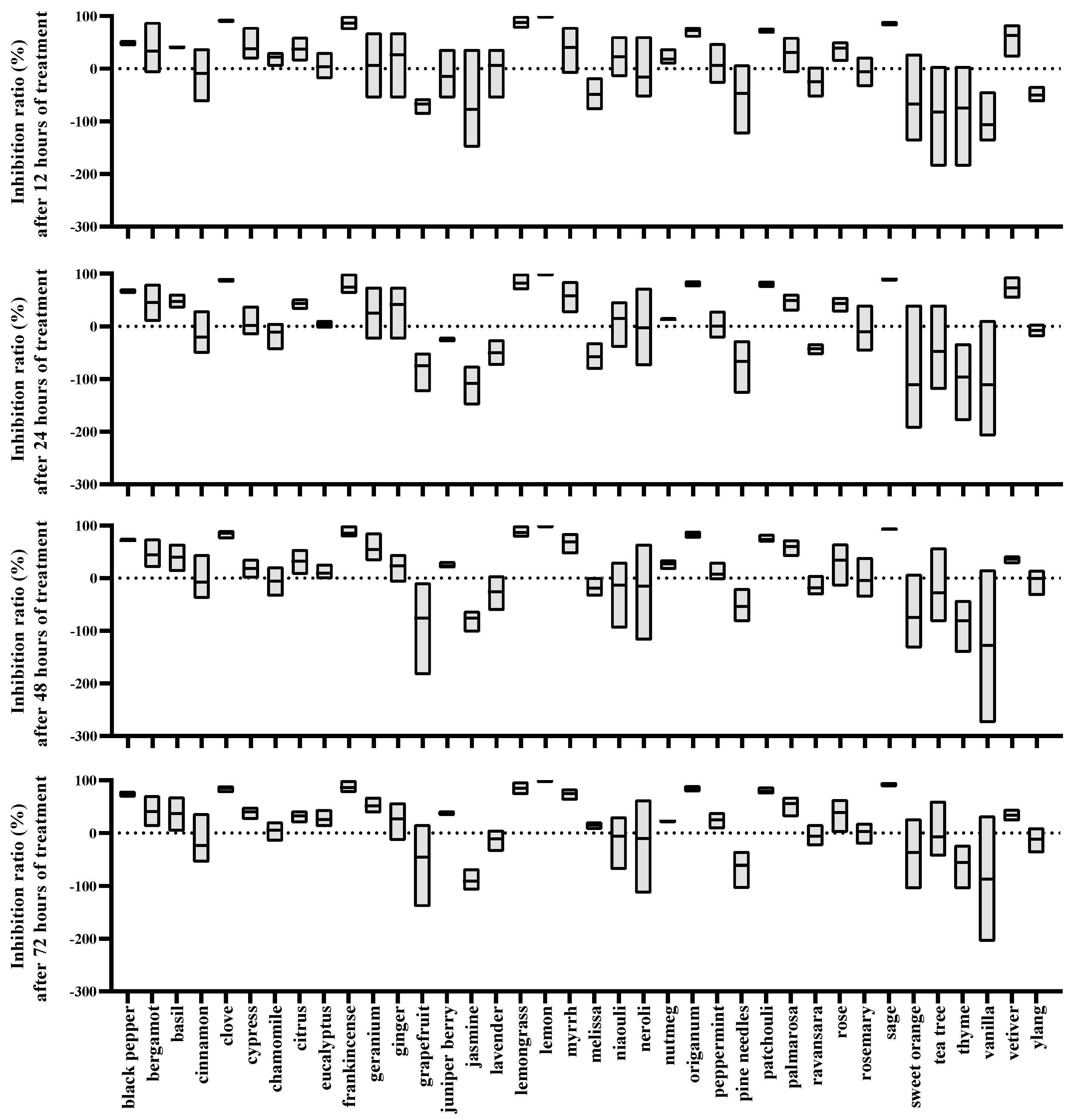
2.3. Effects of EOs on Oviposition of Female T. urticae
3. Discussion
4. Materials and Methods
4.1. Plants, Pest Mites, and Essential Oils
4.2. Toxicity Determination of EOs against T. urticae
4.3. Determination of Oviposition Inhibitory Effects of EOs on T. urticae
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect. Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider Mites Web: A Comprehensive Database for the Tetranychidae[EB/OL]. Available online: https://www1.montpellier.inra.fr/CBGP/spmweb (accessed on 25 October 2022).
- Grbic, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouze, P.; Grbic, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef]
- Jonckheere, W.; Dermauw, W.; Zhurov, V.; Wybouw, N.; Van den Bulcke, J.; Villarroel, C.A.; Greenhalgh, R.; Grbic, M.; Schuurink, R.C.; Tirry, L.; et al. The Salivary Protein Repertoire of the Polyphagous Spider Mite Tetranychus urticae: A Quest for Effectors. Mol. Cell Proteom. 2016, 15, 3594–3613. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef]
- Niu, J.Z.; Dou, W.; Ding, T.B.; Shen, G.M.; Zhang, K.; Smagghe, G.; Wang, J.J. Transcriptome analysis of the citrus red mite, Panonychus citri, and its gene expression by exposure to insecticide/acaricide. Insect. Mol. Biol. 2012, 21, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, C.; Jiang, J.; Song, C.; Wang, C.; Feng, K.; Wei, P.; He, L. Cross resistance, inheritance and fitness advantage of cyetpyrafen resistance in two-spotted spider mite, Tetranychus urticae. Pestic. Biochem. Physiol. 2022, 183, 105062. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect. Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef]
- Mota-Sanchez, D.; Wise, J.C. Arthropod Pesticide Resistance Database (APRD)[EB/OL]. Available online: https://www.pesticideresistance.org/ (accessed on 13 March 2023).
- Rincon, R.A.; Rodriguez, D.; Coy-Barrera, E. Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites. Plants 2019, 8, 272. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends. Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Chaudhari, A.K.; Singh, V.K.; Kedia, A.; Das, S.; Dubey, N.K. Essential oils and their bioactive compounds as eco-friendly novel green pesticides for management of storage insect pests: Prospects and retrospects. Environ. Sci. Pollut. Res. 2021, 28, 18918–18940. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Jalali Sendi, J.; Aliakbar, A.; Razmjou, J. Chemical Composition and Acaricidal Effects of Essential Oils of Foeniculum vulgare Mill. (Apiales: Apiaceae) and Lavandula angustifolia Miller (Lamiales: Lamiaceae) against Tetranychus urticae Koch (Acari: Tetranychidae). Psyche J. Entomol. 2014, 2014, 424078. [Google Scholar] [CrossRef]
- Wagan, T.A.; Cai, W.; Hua, H. Repellency, toxicity, and anti-oviposition of essential oil of Gardenia jasminoides and its four major chemical components against whiteflies and mites. Sci. Rep. 2018, 8, 9375. [Google Scholar] [CrossRef]
- Lazarević, J.; Jevremović, S.; Kostić, I.; Kostić, M.; Vuleta, A.; Manitašević Jovanović, S.; Šešlija Jovanović, D. Toxic, Oviposition Deterrent and Oxidative Stress Effects of Thymus vulgaris Essential Oil against Acanthoscelides obtectus. Insects 2020, 11, 563. [Google Scholar] [CrossRef]
- Stepanycheva, E.; Petrova, M.; Chermenskaya, T.; Pavela, R. Fumigant effect of essential oils on mortality and fertility of thrips Frankliniella occidentalis Perg. Environ. Sci. Pollut. Res. 2019, 26, 30885–30892. [Google Scholar] [CrossRef] [PubMed]
- Antonious, G.F.; Snyder, J.C. Repellency and oviposition deterrence of wild tomato leaf extracts to spider mites, Tetranychus urticae Koch. J. Environ. Sci. Health B 2015, 50, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.I.; Lee, S.G.; Park, H.M.; Ahn, Y.J. Toxicity of plant essential oils to Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae). J. Econ. Entomol. 2004, 97, 553–558. [Google Scholar] [CrossRef]
- Awad, S.E.; Salah, K.B.H.; Jghef, M.M.; Alkhaibari, A.M.; Shami, A.A.; Alghamdi, R.A.; El-Ashry, R.M.; Ali, A.A.I.; El-Maghraby, L.M.M.; Awad, A.E. Chemical Characterization of Clove, Basil and Peppermint Essential Oils; Evaluating Their Toxicity on the Development Stages of Two-Spotted Spider Mites Grown on Cucumber Leaves. Life 2022, 12, 1751. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Tak, J.-H. Miticidal and repellent activity of thirty essential oils and their synergistic interaction with vanillin against Tetranychus urticae Koch (Acari: Tetranychidae). Ind. Crops Prod. 2022, 182, 114872. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, T.; Xie, Y.; Wu, Y.; Deng, J.; Cao, L.; Zhang, Z.; Rao, Q. Screening of the Acaricidal Activity of Essential Oils against Panonychus citri (McGregor) (Acari: Tetranychidae). Agronomy 2023, 13, 397. [Google Scholar] [CrossRef]
- Wang, T.; Ren, Y.; Zhao, J.; Liu, Y.; Xu, B.; Yang, M.; Zhao, W.; Zheng, X.; Wang, J.; Deng, L. Research on the Bioactivity of Plant Essential Oils on Armyworm [Mythimna separata (Walker)] Larvae. Front. Chem. 2022, 10, 936873. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Nassrallah, A.A.; Abdel-Raheem, M.A.; Elbehery, H.H. Lemon peel essential oil and its nano-formulation to control Agrotis ipsilon (Lepidoptera: Noctuidae). Sci. Rep. 2023, 13, 17922. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Soltani, A.; Ben Jemâa, J.M.; Laruelle, F.; Sahraoui, A.L.H. In Vitro Potential of Clary Sage and Coriander Essential Oils as Crop Protection and Post-Harvest Decay Control Products. Foods 2022, 11, 312. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Maggi, F.; Giordani, C.; Cappellacci, L.; Petrelli, R.; Canale, A. Insecticidal activity of two essential oils used in perfumery (ylang ylang and frankincense). Nat. Prod. Res. 2021, 35, 4746–4752. [Google Scholar] [CrossRef]
- Mario, M.B.; Astuti, L.P.; Hsu, J.L.; Kafle, L.; Fernando, I. Bioefficacy of eight different plant powders applied as fumigants against the adzuki bean weevil, Callosobruchus chinensis. Crop Prot. 2023, 167, 106200. [Google Scholar] [CrossRef]
- Afiunizadeh, M.; Karimzadeh, J.; Imani, S.; Moharramipour, S. Insecticidal and oviposition deterrent effects of five medicinal plant extracts on the diamondback moth. J. Plant Dis. Protect. 2022, 129, 805–817. [Google Scholar] [CrossRef]
- Susurluk, H. Potential use of essential oils from Origanum vulgare and Syzygium aromaticum to control Tetranychus urticae Koch (Acari: Tetranychidae) on two host plant species. PeerJ 2023, 11, e14475. [Google Scholar] [CrossRef]
- Roh, H.S.; Lim, E.G.; Kim, J.; Park, C.G. Acaricidal and oviposition deterring effects of santalol identified in sandalwood oil against two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). J. Pest Sci. 2011, 84, 495–501. [Google Scholar] [CrossRef]
- Huang, S.H.; Xian, J.D.; Kong, S.Z.; Li, Y.C.; Xie, J.H.; Lin, J.; Chen, J.N.; Wang, H.F.; Su, Z.R. Insecticidal activity of pogostone against Spodoptera litura and Spodoptera exigua (Lepidoptera: Noctuidae). Pest Manag. Sci. 2014, 70, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Cetin, H.; Erler, F.; Yanikoglu, A. A comparative evaluation of Origanum onites essential oil and its four major components as larvicides against the pine processionary moth, Thaumetopoea wilkinsoni Tams. Pest Manag. Sci. 2007, 63, 830–833. [Google Scholar] [CrossRef]
- Gaspar-Pintiliescu, A.; Mihai, E.; Ciucan, T.; Popescu, A.F.; Luntraru, C.; Tomescu, J.; Craciunescu, O. Antioxidant and acetylcholinesterase inhibition capacity of hyrosols from lamiaceae plants for biopesticide use: Role of phenolics. Int. J. Food Prop. 2022, 25, 996–1008. [Google Scholar] [CrossRef]
- Gad, A.F.; Abdelgalil, G.M.; Radwan, M.A. Bio-molluscicidal potential and biochemical mechanisms of clove oil and its main component eugenol against the land snail, Theba pisana. Pestic. Biochem. Physiol. 2023, 192, 105407. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhou, H.Y.; Tian, L.C. Study on the Synergetic Effect of Avermectins and Plant Essential Oils Used in Combination against Tetranychus cinnabarinus. Plant Dr. 2021, 34, 57–61. (In Chinese) [Google Scholar]
- Francikowski, J.; Baran, B.; Cup, M.; Janiec, J.; Krzyzowski, M. Commercially Available Essential Oil Formulas as Repellents Against the Stored-Product Pest Alphitobius diaperinus. Insects 2019, 10, 96. [Google Scholar] [CrossRef]
- Shlosman, K.; Rein, D.M.; Shemesh, R.; Koifman, N.; Caspi, A.; Cohen, Y. Encapsulation of Thymol and Eugenol Essential Oils Using Unmodified Cellulose: Preparation and Characterization. Polymers 2023, 15, 95. [Google Scholar] [CrossRef]
- Abdelaal, K.; Essawy, M.; Quraytam, A.; Abdallah, F.; Mostafa, H.; Shoueir, K.; Fouad, H.; Hassan, F.A.S.; Hafez, Y. Toxicity of Essential Oils Nanoemulsion against and Their Inhibitory Activity on Insect Enzymes. Processes 2021, 9, 624. [Google Scholar] [CrossRef]
- Hikal, W.M.; Baz, M.M.; Alshehri, M.A.; Bahattab, O.; Baeshen, R.S.; Selim, A.M.; Alhwity, L.; Bousbih, R.; Alshourbaji, M.S.; Ahl, H. Sustainable Pest Management Using Novel Nanoemulsions of Honeysuckle and Patchouli Essential Oils against the West Nile Virus Vector, Culex pipiens, under Laboratory and Field Conditions. Plants 2023, 12, 3682. [Google Scholar] [CrossRef] [PubMed]
- Hogenbom, J.; Jones, A.; Wang, H.V.; Pickett, L.J.; Faraone, N. Synthesis and Characterization of beta-Cyclodextrin-Essential Oil Inclusion Complexes for Tick Repellent Development. Polymers 2021, 13, 1892. [Google Scholar] [CrossRef]
- Peres, M.C.; Costa, G.C.D.; dos Reis, L.E.L.; da Silva, L.D.; Peixoto, M.F.; Alves, C.C.F.; Forim, M.R.; Quintela, E.D.; Araújo, W.L.; Cazal, C.D. In natura and nanoencapsulated essential oils from Xylopia aromatica reduce oviposition of Bemisia tabaci in Phaseolus vulgari. J. Pest Sci. 2020, 93, 807–821. [Google Scholar] [CrossRef]
- Zulu, L.; Gao, H.S.; Zhu, Y.J.; Wu, H.M.; Xie, Y.J.; Liu, X.Y.; Yao, H.F.; Rao, Q. Antifungal effects of seven plant essential oils against Penicillium digitatum. Chem. Biol. Technol. Agric. 2023, 10, 82. [Google Scholar] [CrossRef]
- Subaharan, K.; Senthamarai Selvan, P.; Subramanya, T.M.; Senthoorraja, R.; Manjunath, S.; Das, T.; Pragadheesh, V.S.; Bakthavatsalam, N.; Mohan, M.G.; Senthil-Nathan, S.; et al. Ultrasound-assisted nanoemulsion of Trachyspermum ammi essential oil and its constituent thymol on toxicity and biochemical aspect of Aedes aegypti. Environ. Sci. Pollut. Res. 2022, 29, 71326–71337. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| EOs | 6 h | 12 h | 24 h | 48 h | 72 h | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| LC50 (μL/mL) 95% CL | Regression Equation | LC50 (μL/mL) 95% CL | Regression Equation | LC50 (μL/mL) 95% CL | Regression Equation | LC50 (μL/mL) 95% CL | Regression Equation | LC50 (μL/mL) 95% CL | Regression Equation | |
| frankincense | 25.064 13.012~817.163 | y = 1.0906ln(x) + 1.4438 | 14.994 8.886~163.901 | y = 1.0761ln(x) + 2.2349 | 9.435 6.607~31.513 | y = 1.3868ln(x) + 1.9405 | 6.067 4.270~12.090 | y = 1.8452ln(x) + 1.6412 | 4.611 3.206~7.105 | y = 2.3934ln(x) + 1.073 |
| lemongrass | - | - | 12.225 8.170~36.978 | y = 1.2095ln(x) + 2.1351 | 8.205 5.708~23.423 | y = 1.377ln(x) + 2.1884 | 7.142 5.116~15.579 | y = 1.4857ln(x) + 2.118 | 6.196 4.973~8.538 | y = 1.4856ln(x) + 2.2852 |
| lemon | 13.036 8.890~26.508 | y = 1.1005ln(x) + 2.2964 | 6.203 4.164~10.083 | y = 1.3344ln(x) + 2.4365 | 3.539 2.332~5.142 | y = 1.1263ln(x) + 3.3387 | 2.851 1.562~4.470 | y = 1.2371ln(x) + 3.4333 | 2.310 1.117~3.692 | y = 1.1837ln(x) + 3.841 |
| patchouli | - | - | 15.721 9.831~92.783 | y = 1.0997ln(x) + 1.8758 | 6.795 5.259~10.178 | y = 1.6683ln(x) + 1.6055 | 4.853 4.482~5.320 | y = 2.657ln(x) + 1.2572 | 4.202 3.810~4.604 | y = 2.7609ln(x) + 1.3018 |
| palmarosa | 82.958 33.573~2565.803 | y = 0.7385ln(x) + 1.7545 | 18.586 13.787~36.748 | y = 1.0735ln(x) + 1.5465 | 11.387 8.762~16.731 | y = 1.369ln(x) + 1.412 | 7.865 6.057~10.216 | y = 1.8096ln(x) + 1.3555 | 6.887 5.193~8.551 | y = 1.7287ln(x) + 2.0848 |
| sage | 11.320 9.945~13.176 | y = 1.8253ln(x) + 0.4772 | 6.804 3.994~14.357 | y = 1.4205ln(x) + 2.0342 | 6.588 4.161~9.037 | y = 1.4949ln(x) + 2.3663 | 4.116 2.710~5.743 | y = 1.9369ln(x) + 2.1153 | 3.684 2.059~5.728 | y = 1.904ln(x) + 2.4796 |
| oregano | 37.022 21.843~143.552 | y = 0.8207ln(x) + 1.895 | 12.358 9.672~17.793 | y = 1.2328ln(x) + 1.9051 | 9.733 7.055~15.860 | y = 1.3429ln(x) + 2.2048 | 9.551 6.490~14.821 | y = 1.2911ln(x) + 2.527 | 9.123 6.245~13.072 | y = 1.4264ln(x) + 2.2606 |
| EOs | 12 h | 24 h | 48 h | 72 h |
|---|---|---|---|---|
| black pepper | 0.33 ± 0.05 | 0.51 ± 0.06 | 0.56 ± 0.06 | 0.59 ± 0.08 |
| bergamot | 0.29 ± 0.45 | 0.34 ± 0.32 | 0.32 ± 0.25 | 0.29 ± 0.25 |
| basil | 0.26 ± 0.03 | 0.32 ± 0.13 | 0.27 ± 0.22 | 0.26 ± 0.26 |
| cinnamon | 0.00 ± 0.24 | −0.06 ± 0.21 | 0.00 ± 0.25 | −0.07 ± 0.26 |
| clove | 0.59 ± 0.14 | 0.60 ± 0.13 | 0.69 ± 0.17 | 0.73 ± 0.12 |
| cypress | 0.24 ± 0.34 | 0.06 ± 0.18 | 0.06 ± 0.12 | 0.00 ± 0.11 |
| chamomile | 0.13 ± 0.10 | −0.04 ± 0.12 | −0.01 ± 0.14 | 0.03 ± 0.10 |
| citrus | 0.25 ± 0.18 | 0.29 ± 0.09 | 0.21 ± 0.18 | 0.20 ± 0.09 |
| eucalyptus | 0.03 ± 0.14 | 0.02 ± 0.04 | 0.06 ± 0.09 | 0.15 ± 0.12 |
| frankincense | 0.78 ± 0.21 | 0.63 ± 0.32 | 0.76 ± 0.21 | 0.77 ± 0.20 |
| geranium | 0.11 ± 0.38 | 0.21 ± 0.36 | 0.42 ± 0.30 | 0.36 ± 0.15 |
| ginger | 0.27 ± 0.43 | 0.36 ± 0.41 | 0.15 ± 0.17 | 0.19 ± 0.24 |
| grapefruit | −0.25 ± 0.05 | −0.26 ± 0.10 | −0.22 ± 0.23 | −0.13 ± 0.26 |
| juniper berry | −0.03 ± 0.23 | −0.11 ± 0.00 | 0.13 ± 0.05 | 0.23 ± 0.00 |
| jasmine | −0.19 ± 0.36 | −0.34 ± 0.08 | −0.27 ± 0.06 | −0.31 ± 0.05 |
| lavender | 0.08 ± 0.26 | −0.19 ± 0.08 | −0.10 ± 0.14 | −0.05 ± 0.09 |
| lemongrass | 0.80 ± 0.20 | 0.72 ± 0.25 | 0.78 ± 0.19 | 0.75 ± 0.20 |
| lemon | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | 0.98 ± 0.03 |
| myrrh | 0.32 ± 0.36 | 0.45 ± 0.31 | 0.56 ± 0.24 | 0.61 ± 0.15 |
| melissa | −0.24 ± 0.10 | −0.20 ± 0.08 | −0.12 ± 0.08 | −0.18 ± 0.05 |
| niaouli | 0.17 ± 0.26 | 0.13 ± 0.26 | 0.00 ± 0.28 | 0.01 ± 0.24 |
| neroli | 0.01 ± 0.38 | 0.09 ± 0.44 | 0.05 ± 0.43 | 0.07 ± 0.42 |
| nutmeg | 0.11 ± 0.12 | 0.07 ± 0.02 | 0.16 ± 0.07 | 0.13 ± 0.02 |
| oregano | 0.58 ± 0.14 | 0.69 ± 0.09 | 0.72 ± 0.11 | 0.73 ± 0.10 |
| peppermint | 0.06 ± 0.23 | 0.02 ± 0.15 | 0.05 ± 0.12 | 0.15 ± 0.11 |
| pine needles | −0.15 ± 0.21 | −0.23 ± 0.14 | −0.20 ± 0.10 | −0.23 ± 0.11 |
| patchouli | 0.59 ± 0.08 | 0.64 ± 0.11 | 0.59 ± 0.12 | 0.66 ± 0.11 |
| palmarosa | 0.40 ± 0.20 | 0.48 ± 0.14 | 0.55 ± 0.13 | 0.54 ± 0.14 |
| ravansara | −0.10 ± 0.12 | −0.17 ± 0.04 | −0.08 ± 0.09 | −0.02 ± 0.10 |
| rose | 0.26 ± 0.16 | 0.29 ± 0.12 | 0.26 ± 0.29 | 0.28 ± 0.25 |
| rosemary | −0.02 ± 0.14 | −0.01 ± 0.24 | 0.01 ± 0.22 | 0.02 ± 0.11 |
| sage | 0.77 ± 0.08 | 0.82 ± 0.07 | 0.87 ± 0.05 | 0.86 ± 0.09 |
| sweet orange | −0.18 ± 0.31 | −0.23 ± 0.43 | −0.23 ± 0.24 | −0.11 ± 0.26 |
| tea tree | −0.24 ± 0.25 | −0.12 ± 0.34 | −0.04 ± 0.39 | 0.03 ± 0.35 |
| thyme | −0.21 ± 0.25 | −0.30 ± 0.17 | −0.27 ± 0.12 | −0.20 ± 0.13 |
| vanilla | −0.33 ± 0.13 | −0.28 ± 0.30 | −0.29 ± 0.34 | −0.21 ± 0.37 |
| vetiver | 0.53 ± 0.35 | 0.61 ± 0.27 | 0.22 ± 0.06 | 0.21 ± 0.09 |
| ylang | −0.20 ± 0.05 | −0.03 ± 0.06 | 0.01 ± 0.13 | −0.04 ± 0.11 |
| EOs | Species | Family | EOs | Species | Family |
|---|---|---|---|---|---|
| black pepper | Piper nigrum | Piperaceae | melissa | Melissa officinalis | Lamiaceae |
| bergamot | Citrus medica ‘Fingered’ | Rutaceae | niaouli | Melaleuca viridiflora | Myrtaceae |
| basil | Ocimum basilicum | Lamiaceae | neroli | Citrus × aurantium | Rutaceae |
| cinnamon | Cinnamomum cassia | Lauraceae | nutmeg | Alpinia katsumadai | Zingiberaceae |
| clove | Syringa oblata | Oleaceae | oregano | Origanum vulgare | Lamiaceae |
| cypress | Cupressus sempervirens | Cupressaceae | peppermint | Mentha canadensis | Lamiaceae |
| chamomile | Matricaria chamomilla | Asteraceae | pine needles | Pinus | Pinaceae |
| citrus | Citrus reticulata | Rutaceae | patchouli | Pogostemon cablin | Lamiaceae |
| eucalyptus | Eucalyptus robusta | Myrtaceae | palmarosa | Cymbopogon martini | Poaceae |
| frankincense | Boswellia carteri | Burseraceae | ravansara | Ravensara aromatica | Lauraceae |
| geranium | Pelargonium hortorum | Geraniaceae | rose | Rosa rugosa | Rosaceae |
| ginger | Zingiber officinale | Zingiberaceae | rosemary | Rosmarinus officinalis | Lamiaceae |
| grapefruit | Citrus × aurantium | Rutaceae | sage | Salvia japonica | Lamiaceae |
| juniper berry | Juniperus rigida | Cupressaceae | sweet orange | Citrus sinensis | Rutaceae |
| jasmine | Jasminum sambac | Oleaceae | tea tree | Camellia sinensis | Theaceae |
| lavender | Lavandula angustifolia | Lamiaceae | thyme | Thymus mongolicus | Lamiaceae |
| lemongrass | Cymbopogon citratus | Poaceae | vanilla | Vanilla planifolia | Orchidaceae |
| lemon | Citrus × limon | Rutaceae | vetiver | Chrysopogon zizanioides | Poaceae |
| myrrh | Eucalyptus robusta | Burseraceae | ylang | Cananga odorata | Annonaceae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Wu, T.; Hu, Q.; He, W.; Zheng, Y.; Xie, Y.; Rao, Q.; Liu, X. Plant Essential Oils: Dual Action of Toxicity and Egg-Laying Inhibition on Tetranychus urticae (Acari: Tetranychidae), Unveiling Their Potential as Botanical Pesticides. Plants 2024, 13, 763. https://doi.org/10.3390/plants13060763
Zhu Y, Wu T, Hu Q, He W, Zheng Y, Xie Y, Rao Q, Liu X. Plant Essential Oils: Dual Action of Toxicity and Egg-Laying Inhibition on Tetranychus urticae (Acari: Tetranychidae), Unveiling Their Potential as Botanical Pesticides. Plants. 2024; 13(6):763. https://doi.org/10.3390/plants13060763
Chicago/Turabian StyleZhu, Yijing, Taoqi Wu, Qianyu Hu, Wenze He, Yushi Zheng, Yongjian Xie, Qiong Rao, and Xunyue Liu. 2024. "Plant Essential Oils: Dual Action of Toxicity and Egg-Laying Inhibition on Tetranychus urticae (Acari: Tetranychidae), Unveiling Their Potential as Botanical Pesticides" Plants 13, no. 6: 763. https://doi.org/10.3390/plants13060763