
Multilocus Molecular Phylogeny of the Umbilicaria aprina Group (Umbilicariaceae, Lichenized Ascomycota) Supports Species Level and Neo-Endemic Status of Umbilicaria krascheninnikovii
 , , and
, , and
Abstract
:1. Introduction
2. Results
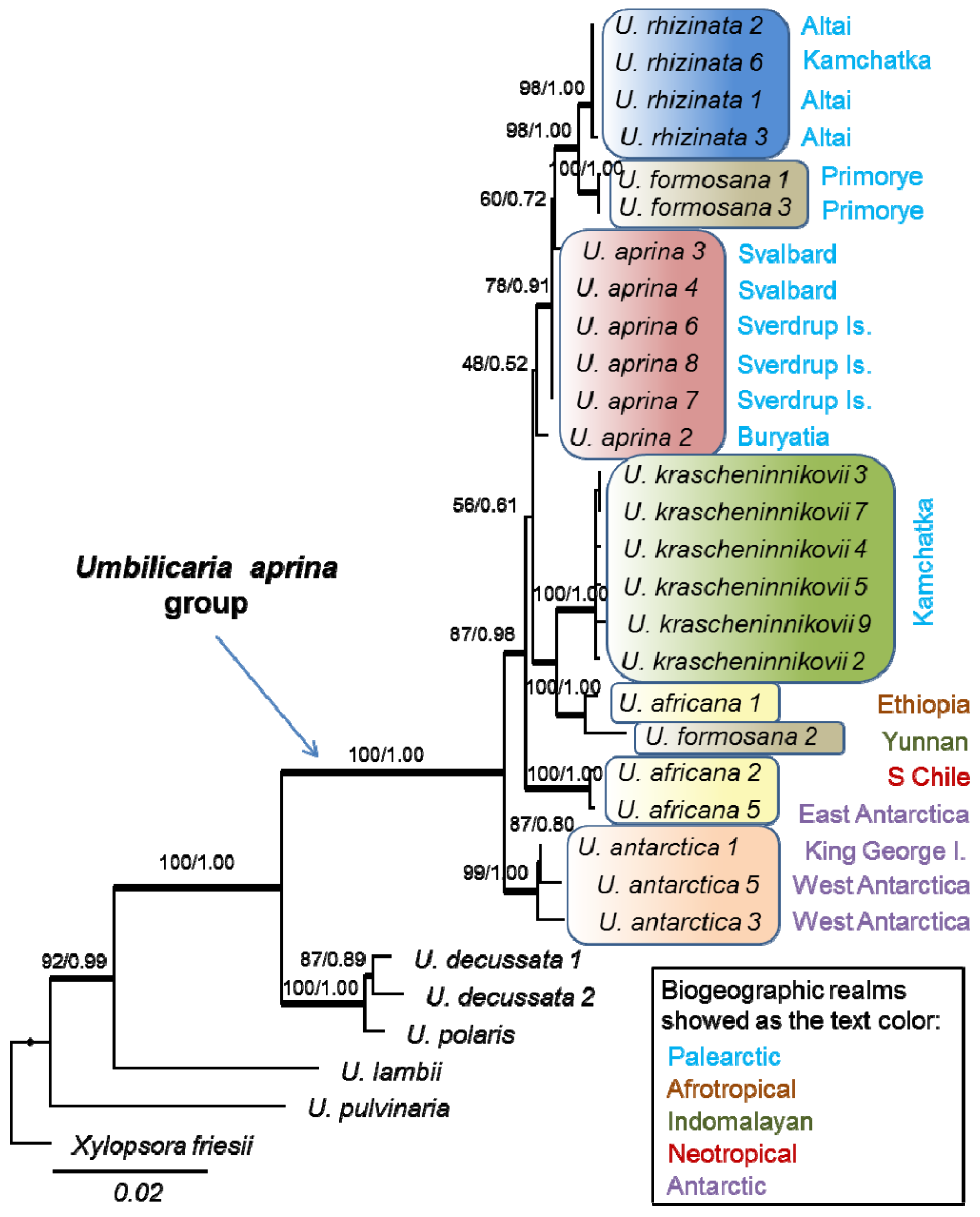
2.1. The Phylogenetic Study
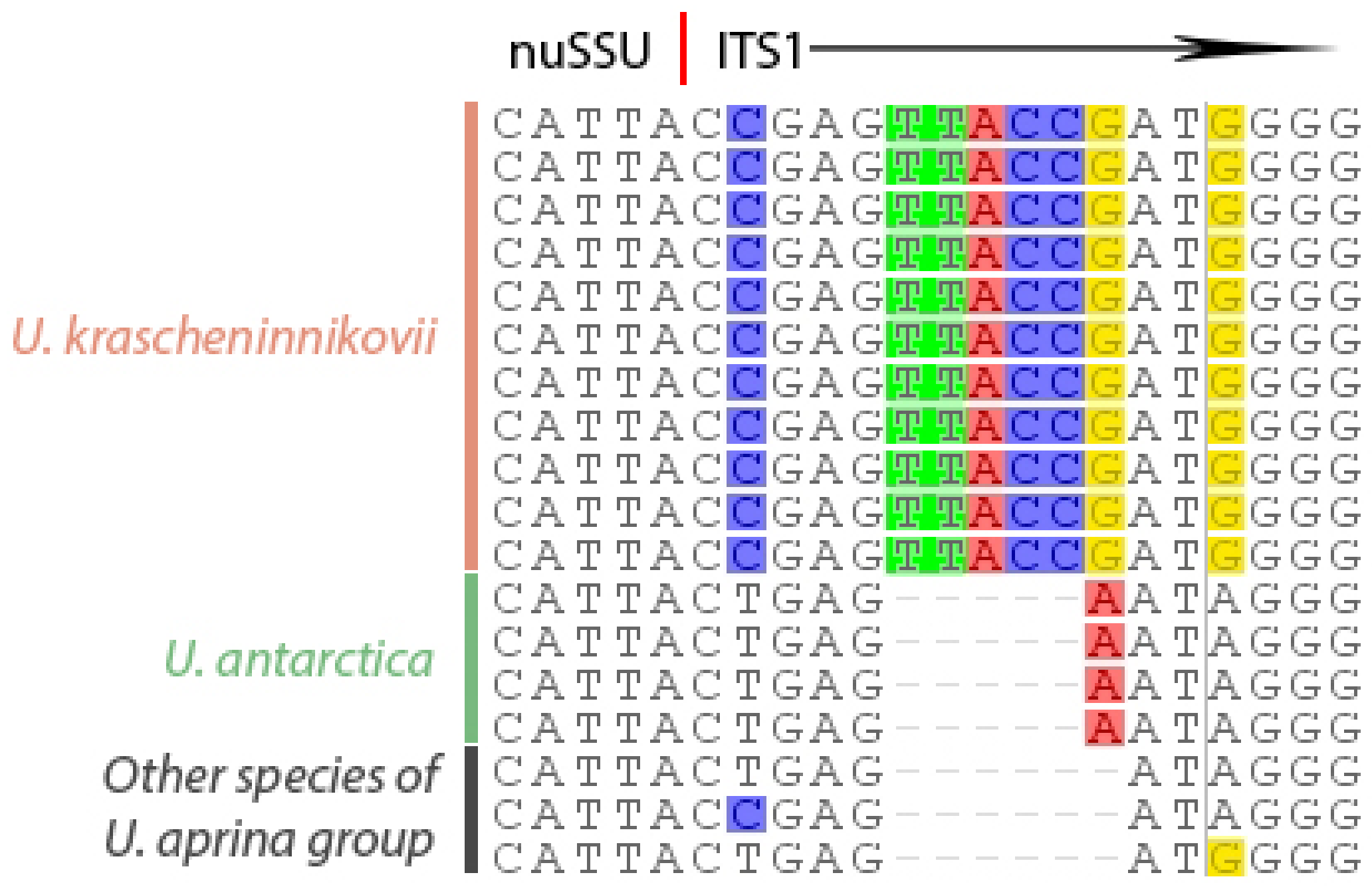
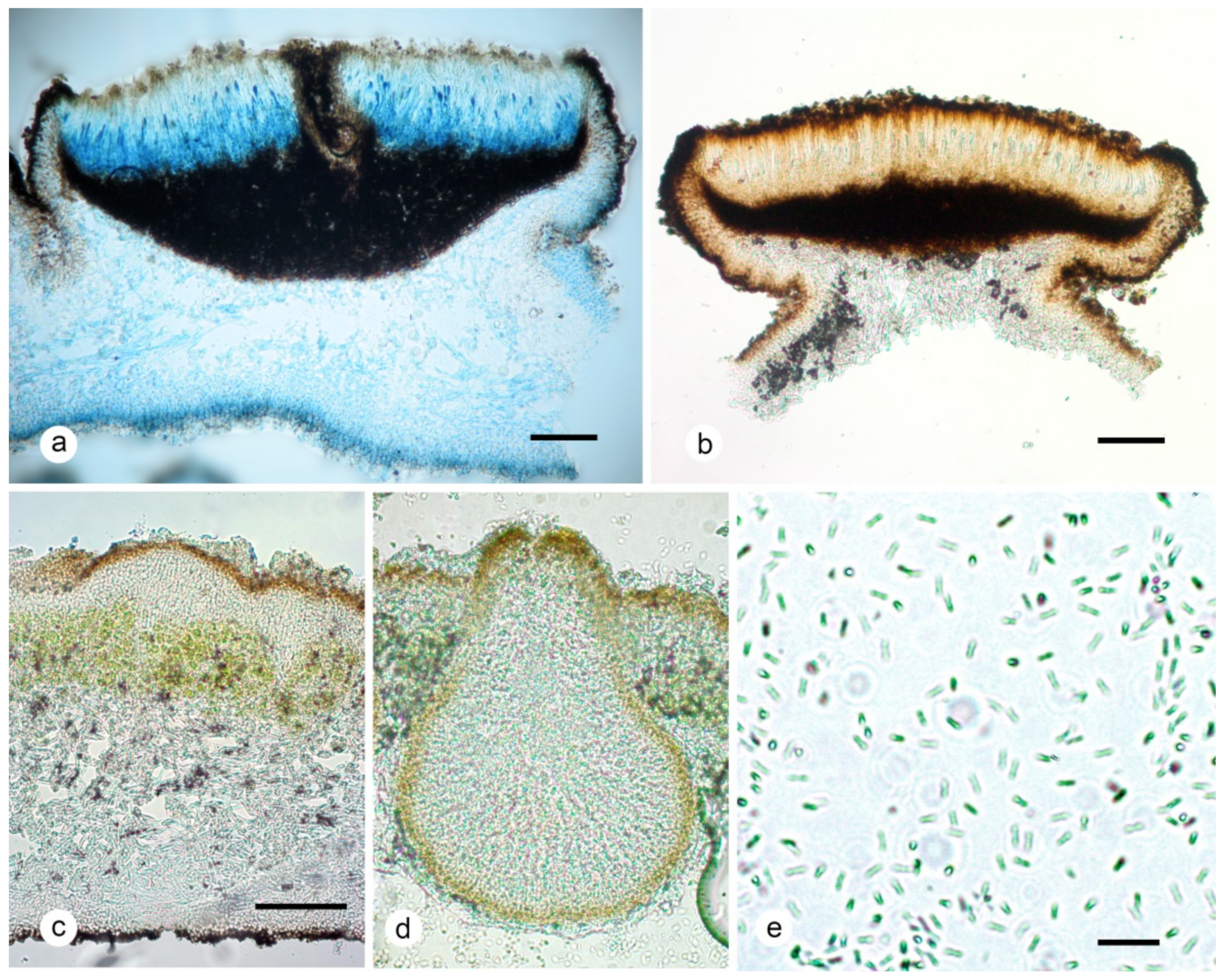
2.2. The Description of Umbilicaria krascheninnikovii
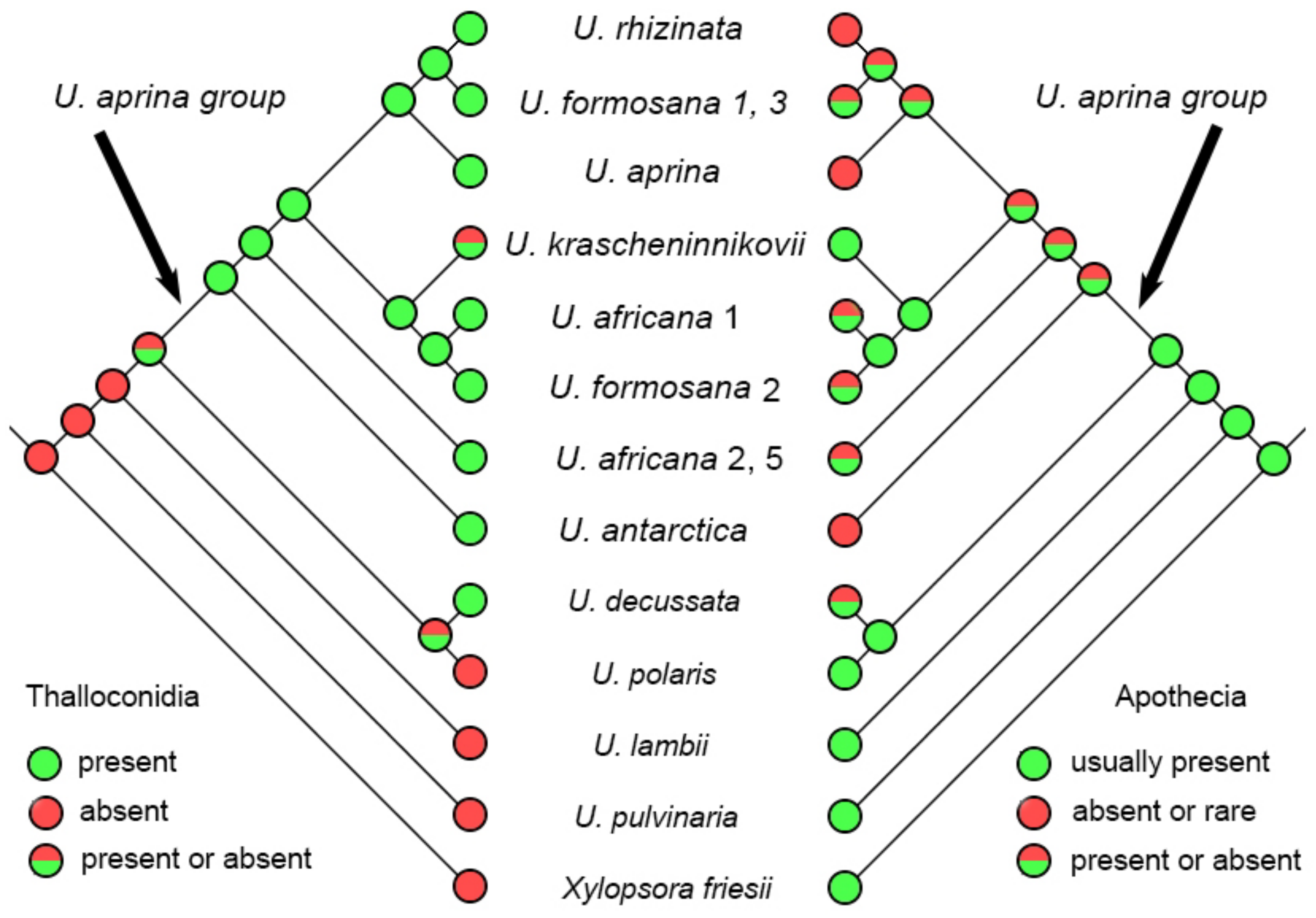
2.3. Reconstruction of the Character Evolution in the Umbilicaria aprina Group
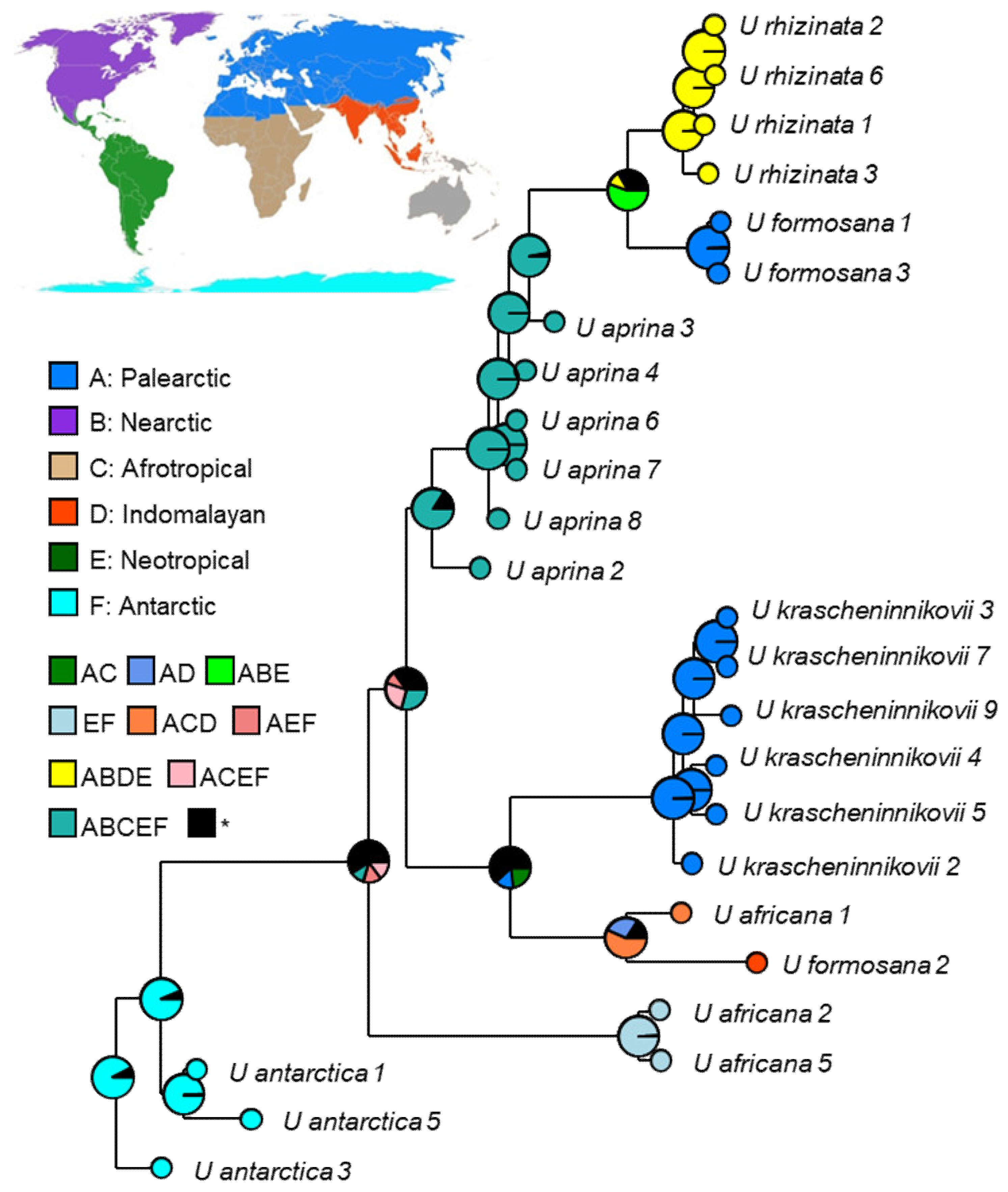
2.4. Reconstruction of the Biogeographic History in the Umbilicaria aprina Group
3. Discussion
3.1. Phylogeny of the Umbilicaria aprina Group
3.2. Phenotypic Traits and Endemism
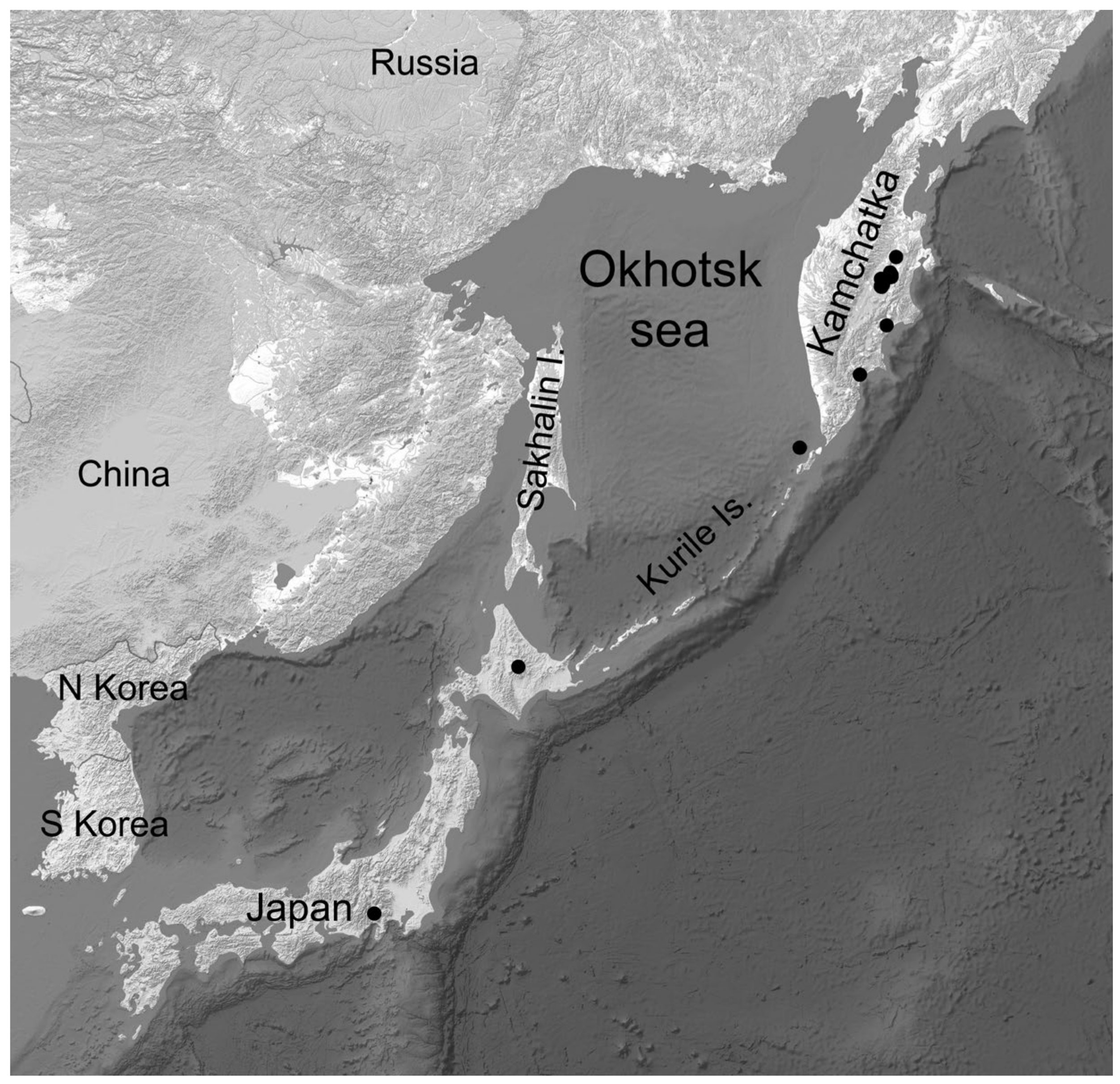
3.3. East Asian Distribution Pattern and Kamchatian Endemism
4. Materials and Methods
4.1. Sampling
4.2. Morphology and Anatomy
4.3. Chemical Analyses
4.4. DNA Extraction, Amplification, and Sequencing
4.5. Sequence Alignment and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | ITS1 | 5.8S rRNA | ITS2 | RPB2 | RPB2 Intron | mtLSU | mtSSU |
|---|---|---|---|---|---|---|---|
| Position | 1–175 | 176–333 | 334–486 | 487–1175 | 1176–1212 | 1213–1949 | 1950–2736 |
| PCR Settings | |||||||
| Primers | ITS 1F-5′/ITS 4-3′ ITS 1F-5′/LR3-3′ | RPB2-980F-5′/fRPB2-7cR-3′ | ML 3-A-5′/ ML 4-A-3′ | mrSSU1-5′/MSU7-3′ | |||
| References | [115,116,117] | [118,119] | [120] | [121,122] | |||
| Datasets | |||||||
| Alignment length (full) | 180 | 158 | 153 | 689 | 36 | 1374 | 803 |
| Alignment length (without ambiguous regions) | 175 | 158 | 153 | 689 | 36 | 736 | 787 |
| Polymorphic sites | 39 | 7 | 19 | 139 | 18 | 111 | 79 |
| Nucleotide diversity π | 0.03508 | 0.00327 | 0.04473 | 0.05463 | 0.12607 | 0.02256 | 0.01720 |
| Number of sequences (ingroup): | 47 | 47 | 47 | 13 | 13 | 41 | 35 |
| Substitution model (IQ-TREE) | TNe+G4 | K2P+I | TNe+G4 | pos.1: TN+F pos.2: F81+F pos.3: HKY+F+I | TNe | HKY+F+I | HKY+F+I |
| Substitution model (MrBayes) | K80+G | K80+I | K80+G | pos.1: K80+I pos.2: K80+I pos.3: HKY+I | HKY+I | HKY+G | HKY+G |
4.6. Reconstruction of the Character Evolution
4.7. Reconstruction of the Biogeographic History
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. (Eds.) The Lichens of Great Britain and Ireland; British Lichen Society: London, UK, 2009; 1046p. [Google Scholar]
- Feuerer, T.; Hawksworth, D.L. Biodiversity of lichens, including a world-wide analysis of checklist data based on Takhtajan’s floristic regions. Biodivers. Conserv. 2007, 16, 85–98. [Google Scholar] [CrossRef]
- Feuerer, T.; Höhne, H. Takhtajan’s definition of global floristic regions revisited. Graph. Scr. 2017, 29, 40–43. [Google Scholar]
- Galloway, D. Lichen biogeography. In Lichen Biology, 2nd ed.; Nash, T.H., III, Ed.; University Press Cambridge: Cambridge, UK, 2008; pp. 315–335. [Google Scholar] [CrossRef]
- Crespo, A.; Pérez-Ortega, S. Cryptic species and species pairs in lichens: A discussion on the relationship between molecularphylogenies and morphological characters. Anales del Jardín Botánico de Madrid 2009, 66, 71–81. [Google Scholar] [CrossRef]
- Lumbsch, H.T.; Leavitt, S.D. Goodbye morphology? A paradigm shift in the delimitation of species in lichenized fungi. Fungal Divers. 2011, 50, 59–72. [Google Scholar] [CrossRef]
- Leavitt, S.D.; Esslinger, T.L.; Divakar, P.K.; Crespo, A.; Lumbsch, H.T. Hidden diversity before our eyes: Delimiting and describingcryptic lichen-forming fungal species in camouflage lichens (Parmeliaceae, Ascomycota). Fungal Biol. 2016, 120, 1374–1391. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.H.; Lücking, R.; Liu, H.J.; Wei, X.L.; Xavier-Leite, A.B.; Portilla, C.V.; Ren, Q.; Wei, J.C. Twelve new species reveal cryptic diversification in foliicolous lichens of Strigula s.lat. (Strigulales, Ascomycota). J. Fungi 2022, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Magain, N.; Miadlikowska, J.; Goffinet, B.; Goward, T.; Pardo-De la Hoz, C.J.; Jüriado, I.; Simon, A.; Mercado-Díaz, J.A.; Barlow, T.; Moncada, B.; et al. High species richness in the lichen genus Peltigera (Ascomycota, Lecanoromycetes): 34 species in the dolichorhizoid and scabrosoid clades of section Polydactylon, including 24 new to science. Persoonia 2023, 51, 1–88. [Google Scholar] [CrossRef]
- Santos, L.A.D.; Aptroot, A.; Lücking, R.; Cáceres, M.E.D.S. Lecanora s. lat. (Ascomycota, Lecanoraceae) in Brazil: DNA Barcoding Coupled with Phenotype Characters Reveals Numerous Novel Species. J. Fungi 2023, 9, 415. [Google Scholar] [CrossRef]
- Aptroot, A. Diversity and endemism in the pyrenocarpous lichen families Pyrenulaceae and Trypetheliaceae in the Malesian flora region. Blumea 2009, 54, 145–147. [Google Scholar] [CrossRef]
- Sérusiaux, E.; Villarreal, J.C.; Wheeler, A.T.; Goffinet, B. Recent origin, active speciation and dispersal for the lichen genus Nephroma (Peltigerales) in Macaronesia. J. Biogeogr. 2011, 38, 1138–1151. [Google Scholar] [CrossRef]
- Van den Boom, P.P.G.; Lücking, R.; Sipman, H.J.M. Notes on Graphidaceae in Macaronesia, with Descriptions of Four New Species. Diversity 2023, 15, 817. [Google Scholar] [CrossRef]
- Dal Forno, M.; Bungartz, F.; Yánez- Ayabaca, A.; Lücking, R.; Lawrey, J.D. High levels of endemism among Galapagos basidiolichens. Fungal Divers. 2017, 85, 45–73. [Google Scholar] [CrossRef]
- Bungartz, F.; Elix, J.A.; Printzen, C. Lecanoroid lichens in the Galapagos Islands: The genera Lecanora, Protoparmeliopsis, and Vainionora (Lecanoraceae, Lecanoromycetes). Phytotaxa 2020, 431, 1–85. [Google Scholar] [CrossRef]
- Lücking, R.; Moncada, B.; Smith, C.W. The genus Lobariella (Ascomycota: Lobariaceae) in Hawaii: Late colonization, high inferred endemism and three new species resulting from “micro–radiation”. Lichenologist 2017, 49, 673–691. [Google Scholar] [CrossRef]
- Mies, B.; Printzen, C. Notes on the lichens of Socotra (Yemen, Indian Ocean). In Progress and Problems in Lichenology in the Nineties. Bibliotheca Lichenologica; Türk, R., Zorer, R., Eds.; J. Cramer in der Gebrüder Borntraeger Verlagsbuchhandlung; Borntraeger Verlagsbuchhandlung: Berlin, Germany; Stuttgart, Germany, 1997; Volume 68, pp. 223–239. [Google Scholar]
- Mercado-Díaz, J.A.; Lücking, R.; Moncada, B.; Campbell, K.C.E.; Delnatte, C.; Familia, L.; Falcón-Hidalgo, B.; Motito-Marín, A.; Rivera-Queralta, Y.; Widhelm, T.J.; et al. Species assemblages of insular Caribbean Sticta (lichenized Ascomycota: Peltigerales) over ecological and evolutionary time scales. Mol. Phylogenet. Evol. 2023, 186, 107830. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Goffinet, B.; Magain, N.; Sérusiaux, E. High diversity, high insular endemism and recent origin in the lichen genus Sticta (lichenized Ascomycota, Peltigerales) in Madagascar and the Mascarenes. Mol. Phylogenet. Evol. 2018, 122, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S. Phytogeographical elements of the lichen flora of Japan. J. Hattori Bot. Lab. 2006, 100, 721–738. [Google Scholar] [CrossRef]
- Ohmura, Y.; Kashiwadani, H. Checklist of lichens and allied fungi of Japan. Natl. Mus. Nat. Sci. Monogr. 2018, 49, 1–140. [Google Scholar]
- Galloway, D.J. Flora of New Zealand Lichens, Including Lichen-Forming and Lichenicolous Fungi, 2nd ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2007; pp. 1–2397. [Google Scholar]
- Kantvilas, G. The genus Menegazzia (Lecanorales: Parmeliaceae) in Tasmania revisited. Lichenologist 2012, 44, 189–246. [Google Scholar] [CrossRef]
- Sipman, H.J.M. Lichens from Mount Kinabalu. Trop. Bryol. 1993, 8, 281–314. [Google Scholar] [CrossRef]
- Hestmark, G. The lichen genus Umbilicaria in Ecuador. Nord. J. Bot. 2016, 34, 257–268. [Google Scholar] [CrossRef]
- Hestmark, G.; Miadlikowska, J.; Kauff, F.; Fraker, E.; Molnar, K.; Lutzoni, F. Single origin and subsequent diversification of central Andean endemic Umbilicaria species. Mycologia 2011, 103, 45–56. [Google Scholar] [CrossRef]
- Tibell, L.B. Distribution patterns and dispersal strategies of Caliciales. Bot. J. Linn. Soc. 1994, 116, 159–202. [Google Scholar] [CrossRef]
- Muñoz, J.; Felicísimo, Á.M.; Cabezas, F.; Burgaz, A.R.; Martínez, I. Wind as a Long-Distance Dispersal Vehicle in the Southern Hemisphere. Science 2004, 304, 1144–1147. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.M.; Koumoutsaris, S.; Mitchell, E.A.D.; Bey, I. Modelling the effect of size on the aerial dispersal of microorganisms. J. Biogeogr. 2012, 39, 89–97. [Google Scholar] [CrossRef]
- Bendiksby, M.; Timdal, E. Molecular phylogenetics and taxonomy of Hypocenomyce sensu lato (Ascomycota: Lecanoromycetes): Extreme polyphyly and morphological/ecological convergence. Taxon 2013, 62, 940–956. [Google Scholar] [CrossRef]
- Davydov, E.A.; Peršoh, D.; Rambold, G. Umbilicariaceae (lichenized Ascomycota)—Trait evolution and a new generic concept. Taxon 2017, 66, 1282–1303. [Google Scholar] [CrossRef]
- Prieto, M.; Wedin, M. Dating the diversification of the major lineages of Ascomycoa (fungi). PLoS ONE 2013, 8, e65576. [Google Scholar] [CrossRef]
- Frey, E. Cladoniaceae (unter Ausschluß d. Gatt. Cladonia) Umbilicariaceae. In Dr. L. Rabenhorst’s Kryptogamenflora von Deutschland, Österreich und der Schweiz; Zahlbruckner, A., von Keissler, K., Eds.; Akademische Verlagsgeselschaft M.B.H.: Leipzig, Germany, 1933; Volume 9, pp. 1–424. [Google Scholar]
- Frey, E. Neue Beitrage zu einer Monographie des Genus Umbilicaria. (Hoffm.) Nyl. Ber. Schweiz. Bot. Ges. 1949, 59, 427–470. [Google Scholar]
- Llano, G.A. A Monograph of the Lichen Family Umbilicariaceae in the Western Hemisphere; Navexos P-831, Office of Naval Research: Washington, DC, USA, 1950; pp. 1–281. [Google Scholar]
- Wei, J.C.; Jiang, Y.M. The Asian Umbilicariaceae (Ascomycota). Mycosystema monographicum Ser. No. 1; International Academic Publishers: Beijing, China, 1993; pp. 1–217. [Google Scholar]
- Galloway, D.J.; Sancho, L.G. Umbilicaria murihikuana and U. robusta (Umbilicariaceae: Ascomycota), two new taxa from Aotearoa New Zealand. Australas. Lichenol. 2005, 56, 16–19. [Google Scholar]
- Davydov, E.A.; Yakovchenko, L.S.; Urbanavichene, I.; Konoreva, L.; Chesnokov, S.; Kharpukhaeva, T.; Obermayer, W. Umbilicaria orientalis—A new species of Umbilicaria subg. Papillophora with an East Asian distribution: Morphological delimitation and molecular evidence. Lichenologist 2020, 52, 353–364. [Google Scholar] [CrossRef]
- Davydov, E.A. Umbilicaria platyrhiza—A new Mediterranean endemic species of the subgenus Papillophora (Umbilicariaceae, lichenized Ascomycota). Phytotaxa 2022, 533, 143–148. [Google Scholar] [CrossRef]
- Davydov, E.A.; Masson, D. Umbilicaria meizospora comb. nov.—A southwestern European endemic species of the subgenus Papillophora. Lichenologist 2022, 54, 1–12. [Google Scholar] [CrossRef]
- Savicz, V.P. Neue Flechten aus Kamtschatka. Bull. Jard. Imp. Bot. Pierre Gd. 1914, 14, 111–128. [Google Scholar]
- Davydov, E.A.; Blum, O.B.; Kashevarov, G.P.; Grakhov, V.P. Umbilicaria subpolyphylla Oxner the correct name for U. iberica Sancho et Krzewicka and its bipolar distribution pattern. Lichenologist 2019, 51, 205–220. [Google Scholar] [CrossRef]
- Himelbrant, D.E.; Stepanchikova, I.S.; Kuznetsova, E.S. Lichens. In Vegetation Cover of Volcanic Plateaus of Central Kamchatka; Neshataeva, V.Y., Ed.; KMK: Moscow, Russia, 2014; pp. 121–164. [Google Scholar]
- Himelbrant, D.E.; Stepanchikova, I.S.; Ahti, T.; Neshataeva, V.Y. The first lichenological survey in Koryakia (Northern Kamchatka, Russia)—The last unexplored part of Beringia. Nov. Syst. Nizshikh Rastenii 2019, 53, 107–142. [Google Scholar] [CrossRef]
- Himelbrant, D.E.; Stepanchikova, I.S.; Ahti, T.; Neshataeva, V.Y. New exploration in Koryakia—The lichens of the Cape Goven, Bering Sea coast (Northern Kamchatka, Russia). Nov. Syst. Nizshikh Rastenii 2021, 55, 121–162. [Google Scholar] [CrossRef]
- Himelbrant, D.E.; Stepanchikova, I.S.; Neshataeva, V.Y.; Kirichenko, V.E.; Skvortsov, K.I.; Timofeeva, E.A. Further knowledge on the lichen biota of Koryakia (Northern Kamchatka, Russia). Nov. Syst. Nizshikh Rastenii 2023, 57, L29–L37. [Google Scholar] [CrossRef]
- Scholander, P.F. On the apothecia in the lichen family Umbilicariaceae. Nytt Mag. Naturvidenskaberne 1934, 75, 1–132. [Google Scholar]
- Du Rietz, G.E. The lichens of the Swedish Kamtchatka expedition. Ark. För Bot. 1929, 22, 1–25. [Google Scholar]
- Lynge, B.; Scholander, P.F. Lichens from North East Greenland collected on the Norwegian Scientific Expeditions in 1929 and 1930. Skr. Svalbard Ishavet 1932, 41, 1–116. [Google Scholar]
- Imshaug, H.A. Alpine lichens of western United States and adjacent Canada. I. The Macrolichens. Bryologist 1957, 60, 177–272. [Google Scholar] [CrossRef]
- Poelt, J. Die Gattung Umbilicaria (Umbilicariaceae) (Flechten des Himalaya 14). Khumbu Himal Ergeb. Forsch. Nepal Himalaya 1977, 6, 397–435. [Google Scholar]
- Galloway, D.J. Additional lichen records from New Zealand 34. Caloplaca tornoensis H. Magn., Umbilicaria grisea Hoffm., and Umbilicaria krascheninnikovii (Savicz) Zahlbr., three bipolar lichens. Australas. Lichenol. 2001, 49, 21–27. [Google Scholar]
- Øvstedal, D.O.; Lewis Smith, R.I. Lichens of Antarctica and South Georgia: A Guide to Their Identification and Ecology; Cambridge University Press: Cambridge, UK, 2001; pp. 1–424. [Google Scholar]
- Krzewicka, B.; Smykla, J. The lichen genus Umbilicaria from the neighbourhood of Admiralty Bay (King George Island, maritime Antarctic), with a proposed new key to all Antarctic taxa. Polar Biol. 2004, 28, 15–25. [Google Scholar] [CrossRef]
- Kristinsson, H.; Zhurbenko, M.; Hansen, E.S. Panarctic Checklist of Lichens and Lichenicolous Fungi, CAFF Technical Report No. 20; CAFF International Secretariat: Akureyri, Iceland, 2010; pp. 1–120. [Google Scholar]
- Spribille, T.; Fryday, A.M.; Hampton-Miller, C.J.; Ahti, T.; Dillman, K.; Thor, G.; Tonsberg, T.; Schirokauer, D. Compendium of the lichens and associated fungi of Alaska. Bibl. Lichenol. 2023, 112, 1–522. [Google Scholar] [CrossRef]
- Hasenhüttl, G.; Poelt, J. Über die Brutkorner bei der Flechtengattung Umbilicaria. Berichte Dtsch. Bot. Geselschaft 1978, 91, 275–296. [Google Scholar] [CrossRef]
- Hestmark, G. Teleomorph-anamorph relationships in Umbilicaria II. Patterns in propagative morph production. Lichenologist 1991, 23, 361–380. [Google Scholar] [CrossRef]
- Girina, O.; Manevich, A.; Loupian, E.; Uvarov, I.; Korolev, S.; Sorokin, A.; Romanova, I.; Kramareva, L.; Burtsev, M. Monitoring the thermal activity of Kamchatkan volcanoes during 2015–2022 Using Remote Sensing. Remote Sens. 2023, 15, 4775. [Google Scholar] [CrossRef]
- Solomina, O.; Wiles, G.; Shiraiwa, T.; d’Arrigo, R. Multiproxy records of climate variability for Kamchatka for the past 400 years. Clim. Past 2007, 3, 119–128. [Google Scholar] [CrossRef]
- Cain, S.A. Some observations on the concept of species senescence. Ecology 1940, 21, 213–215. [Google Scholar] [CrossRef]
- Stebbins, G.L.; Major, J. Endemism and speciation in the California flora. Ecol. Monogr. 1965, 35, 2–35. [Google Scholar] [CrossRef]
- Lücking, R.; Moncada, B.; McCune, B.; Farkas, E.; Goffinet, B.; Parker, D.; Chaves, J.L.; Lőkös, L.; Nelson, P.R.; Spribille, T.; et al. Pseudocyphellaria crocata (Ascomycota: Lobariaceae) in the Americas is revealed to be thirteen species, and none of them is P. crocata. Bryologist 2017, 120, 441–500. [Google Scholar] [CrossRef]
- Poelt, J.; Nash, T.H. Studies in the Umbilicaria vellea group (Umbilicariaceae) in North America. Bryologist 1993, 96, 422–430. [Google Scholar] [CrossRef]
- Frey, E.; Lamb, I.M. A new species of Umbilicaria from the Antarctic. Trans. Br. Mycol. Soc. 1939, 22, 270–273. [Google Scholar] [CrossRef]
- Romeike, J.; Friedl, T.; Helms, G.; Ott, S. Genetic diversity of algal and fungal partners in four species of Umbilicaria (Lichenized Ascomycetes) along a transect of the Antarctic Peninsula. Mol. Biol. Evol. 2002, 19, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.; Brinkmann, M.; Wirtz, N.; Lumbsch, H.T. Mitochondrial and nuclear ribosomal DNA data do not support the separation of the Antarctic lichens Umbilicaria kappeni and Umbilicaria antarctica as distinct species. Lichenologist 2004, 36, 227–234. [Google Scholar] [CrossRef]
- Grebelnyi, S.D. Natural clonal races and species: Aristocrats or degenerates? In Cloning in Nature. A Part Playing by Gene Recombination Stoppage in Fauna and Flora Formation; Grebelnyi, S.D., Ed.; Zoological Institute RAS: St. Petersburg, Russia, 2008; pp. 1–288. [Google Scholar]
- Drenth, A.; McTaggart, A.R.; Wingfield, B.D. Fungal clones win the battle, but recombination wins the war. IMA Fungus 2019, 10, 18. [Google Scholar] [CrossRef]
- Nieuwenhuis, B.P.S.; James, T.Y. The frequency of sex in fungi. Philos. Trans. R. Soc. B 2016, 371, 20150540. [Google Scholar] [CrossRef]
- Hestmark, G. To sex, or not to sex… Structures and strategies of reproduction in the family Umbilicariaceae (Lecanorales, Ascomycetes). Sommerfeltia 1991, 3 (Suppl. 3), 1–47. [Google Scholar] [CrossRef]
- Seymour, F.; Crittenden, P.; Dyer, P. Sex in the extremes: Lichen-forming fungi. Mycologist 2005, 19, 51–58. [Google Scholar] [CrossRef]
- Buschbom, J.; Mueller, G.M. Testing “species pair” hypotheses: Evolutionary processes in the lichen-forming species complex Porpidia flavocoerulescens and Porpidia melinodes. Mol. Biol. Evol. 2006, 23, 574–586. [Google Scholar] [CrossRef]
- Hestmark, G. Conidiogenesis in five species of Umbilicaria. Mycol. Res. 1992, 96, 1033–1043. [Google Scholar] [CrossRef]
- Dobzhansky, T. Genetics and the Origin of Species, 3rd ed.; Columbia University Press: New York, NY, USA, 1951; pp. 1–364. [Google Scholar]
- Davydov, E.A.; Chesnokov, S.V.; Konoreva, L.A.; Andreev, M.P. Umbilicariaceae (lichenized Ascomycota) from the Stanovoye Nagor’e Highlands (South Siberia, Russia). Herzogia 2019, 32, 472–484. [Google Scholar] [CrossRef]
- Takhtajan, A. Floristic Regions of the World, 1st ed.; University of California Press: Berkeley, CA, USA, 1986; pp. 1–522. (In English) [Google Scholar]
- Kamelin, R.V. Flora of the Earth: Floristic Regions of the Land; Piat’ Plus: St. Petersburg, Russia; Barnaul, Russia, 2017; pp. 1–128. [Google Scholar]
- Wang, S.-Q.; Dong, X.-Y.; Ye, L.; Wang, H.-F.; Ma, K.-P. Flora of Northeast Asia. Plants 2023, 12, 2240. [Google Scholar] [CrossRef]
- Urbanavichus, G.P. Specific features of lichen diversity of Russia. Izv. Ross. Akad. Nauk Ser. Geogr. 2011, 1, 66–78. [Google Scholar]
- Wei, J.C. The Enumeration of Lichenized Fungi in China; China Forestry Publihsing House: Beijing, China, 2020; pp. 1–606. [Google Scholar]
- Davydov, E.A.; Yakovchenko, L.; Ohmura, Y. The new lectotypification of Umbilicaria kisovana (Umbilicariaceae, lichenized Ascomycota). Mycotaxon 2014, 129, 415–419. [Google Scholar] [CrossRef]
- Wei, J.C.; Jiang, Y.M.; Guo, S.Y. A new species of Umbilicaria. Mycosystema 1996, 8–9, 65–70. [Google Scholar]
- Davydov, E.A.; Ohmura, Y. Lectotypification of the name Umbilicaria esculenta. Mycotaxon 2016, 131, 407–412. [Google Scholar] [CrossRef]
- Davydov, E.A.; Himelbrant, D.E.; Stepanchikova, I.S. Contribution to the study of Umbilicariaceae (lichenized Ascomycota) in Russia. II. Kamchatka Peninsula. Herzogia 2011, 24, 229–241. [Google Scholar] [CrossRef]
- Davydov, E.A.; Tchabanenko, S.I.; Makryi, T.V.; Khanin, V.A. The second confirmed record of Umbilicaria pulvinaria (lichenized Ascomycota) since its original description in 1914. Turczaninowia 2011, 14, 119–122. [Google Scholar]
- Trass, H.H. On the lichen flora of Kamchatka. In Investigationes Naturae Orientis Extremis; Parmasto, E.H., Ryke, A.A., Trass, H.H., Eds.; Academia Scientiarum, R.S.S.: Tallin, Estonia, 1963; pp. 170–220. [Google Scholar]
- Mikulin, A.G. A contribution to the lichen flora of the Kronotsky State Reserve (Kamchatskaya oblast). In Flora and Systematics of Non-Vascular Plants of Far East; Vasilieva, L.N., Ed.; Far Eastern Scientific Center of the Academy of Sciences of USSR: Vladivostok, Russia, 1986; pp. 137–150. [Google Scholar]
- Mikulin, A.G. Lichenes pro Paeninsula Kamczatka novi. Nov. Syst. Nizshikh Rastenii 1987, 24, 163–165, (In Russian with Latin Abstract). [Google Scholar]
- Mikulin, A.G. High-mountain lichens of the Kronotsky State Reserve (Kamchatka). In The Vegetation of High-Mountain Ecosystems of USSR; Barkalov, V.Y., Ivanina, L.I., Krasnoborov, I.M., Eds.; Far Eastern Scientific Center of the Academy of Sciences of USSR: Vladivostok, Russia, 1988; pp. 149–158. [Google Scholar]
- Yakubov, V.V. Endemics of the Kamchatka flora. In Conservation of Biodiversity of Kamchatka and Adjacent Seas: Materials of V International Scientific Conference; Balykin, P.A., Lysenko, V.N., Moiseev, R.S., Tokranov, A.M., Chernyagina, O.A., Eds.; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2004; pp. 112–115. [Google Scholar]
- Koval, M.V.; Esin, E.V.; Bugaev, A.V.; Karas, V.A.; Gorin, S.L.; Shatilo, I.V.; Pogdaev, E.G.; Shubkin, S.V.; Zavarina, L.O.; Frolov, O.V.; et al. Freshwater ichtyofauna of the rivers Penzhina and Talovka (North-West Kamchatka). Stud. Water Biol. Resour. Kamchatka Northwest. Part Pac. Ocean 2015, 37, 53–145. [Google Scholar] [CrossRef]
- Esin, E.V.; Markevich, G.N. Charrs of Genus Salvelinus of Asian North Pacific: Origin, Evolution and Modern Diversity; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2017; pp. 1–187. [Google Scholar]
- Kharkevich, S.S.; Buch, T.G. Flora of the Russian Far East: Flora Exsiccata; Dalnauka: Vladivostok, Russia, 1999; pp. 1–250. [Google Scholar]
- Mutin, V.A. Scandinavia and Kamchatka: Keys to research of fauna genesis in Palaearctic. Chteniya Pamyati Aleksandra Ivanovicha Kurentsova 2014, 25, 14–24. [Google Scholar]
- Ahti, T.; Stenroos, S. Nuevos datos sobre nomenclatura, taxonomía y distribución de algunas especies del género “Cladonia”. Bot. Complut. 2012, 36, 31–34. [Google Scholar] [CrossRef]
- Huovinen, K.; Ahti, T.; Stenroos, S. The composition and contents of aromatic lichen substances in Cladonia, cection Cocciferae. Ann. Bot. Fenn. 1989, 26, 133–148. [Google Scholar]
- Ickert-Bond, S.M.; Murray, D.F.; Dechaine, E. Contrasting patterns of plant distribution in Beringia. Park Sci. 2009, 8, 26–32. [Google Scholar]
- Novichkova, A.A.; Chertoprud, E.S. The species structure, biogeographical status, and the relation to the Beringian fauna of microcrustaceans (Cladocera, Copepoda) of the Magadan Area (Far East, Russia). Arthropoda Sel. 2022, 31, 283–292. [Google Scholar] [CrossRef]
- Oke, T.A.; Stralberg, D.; Reid, D.G.; Bennett, B.A.; Cannings, S.; Willier, C.; Fulkerson, J.R.; Cooke, H.A.; Mantyka-Pringle, C.S. Warming drives poleward range contractions of Beringian endemic plant species at high latitudes. Divers. Distrib. 2023, 29, 509–523. [Google Scholar] [CrossRef]
- Spribille, T.; Fryday, A.; Pérez-Ortega, S.; Svensson, M.; Tønsberg, T.; Ekman, S.; Holien, H.; Resl, P.; Schneider, K.; Stabentheiner, E.; et al. Lichens and associated fungi from Glacier Bay National Park, Alaska. Lichenologist 2020, 52, 61–181. [Google Scholar] [CrossRef]
- McCune, B.; Arup, U.; Breuss, O.; Di Meglio, E.; Di Meglio, J.; Esslinger, T.L.; Miadlikowska, J.; Miller, A.E.; Rosentreter, R.; Schultz, M.; et al. Biodiversity and ecology of lichens of Kenai Fjords National Park, Alaska. Plant Fungal Syst. 2020, 65, 586–619. [Google Scholar] [CrossRef]
- Frolov, I.; Himelbrant, D.; Stepanchikova, I.; Prokopiev, I. Polycauliona comandorica, a new fruticulose species in the family Teloschistaceae from the Commander Islands, Russia. Lichenologist 2021, 53, 299–306. [Google Scholar] [CrossRef]
- Thomson, J.W. American Arctic Lichens 1. The Macrolichens; Columbia University Press: New York, NY, USA, 1984; pp. 1–504. [Google Scholar]
- Culberson, C.F.; Kristinsson, H.A. A standardized method for the identification of lichen product. J. Chromatogr. 1970, 46, 85–93. [Google Scholar] [CrossRef]
- Wedin, M.; Wiklund, E.; Crewe, A.; Döring, H.; Ekman, S.; Nyberg, Å.; Schmitt, I.; Lumbsch, H.T. Phylogenetic relationships of Lecanoromycetes (Ascomycota) as revealed by analyses of mtSSU and nLSU rDNA sequence data. Mycol. Resour. 2005, 109, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Larget, B.; Simon, D. Markov chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- Ronquist, R.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Ekman, S.; Blaalid, R. The Devil in the Details: Interactions between the Branch-Length Prior and Likelihood Model Affect Node Support and Branch Lengths in the Phylogeny of the Psoraceae. Syst. Biol. 2011, 60, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the Pezizomycotina (Euascomycetes, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef] [PubMed]
- Printzen, C. Fungal specific primers for PCR-amplification of mitochondrial LSU in lichens. Mol. Ecol. Notes 2002, 2, 130–132. [Google Scholar] [CrossRef]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. [Google Scholar] [CrossRef]
- Zhou, S.; Stanosz, G.R. Primers for amplification of mtSSU rDNA, and a phylogenetic study of Botryosphaeria and associated anamorphic fungi. Mycol. Res. 2001, 105, 1033–1044. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.81. 2023. Available online: http://www.mesquiteproject.org (accessed on 12 December 2023).
- Yu, Y.; Blair, C.; He, X.J. RASP 4: Ancestral State Reconstruction Tool for Multiple Genes and Characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef]
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar] [CrossRef]
- Matzke, N.J. Model selection in historical biogeography reveals that founder-event speciation is a crucial process in Island clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef]
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef]
- Ronquist, F. Dispersal-vicariance analysis: A new approach to the quantification of historical biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian analysis of biogeography when the number of areas is large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef] [PubMed]











| U. krascheninnikovii | U. rhizinata | U. aprina | U. formosana | U. polaris | |
|---|---|---|---|---|---|
| Thallus size | Small | Small | Medium | Medium | Medium |
| Upper surface center | Elevated, with rough “crystals” of necral layer, newer reticulate | Elevated, pruinose to areolate, newer reticulate | Elevated or depressed or weakly ridded, pruinose | Elevated or depressed, with reticulate pattern, pruinose | With reticulate pattern and rough “crystals” of necral layer |
| Apothecia | Common and abundant | Unknown | Rare | Common | Common and abundant |
| Thalloconidia | Eventually present, light brown to brown, non-septate | Common, brown, (1–2)3–4 cellular | Common, brown, non-septate | Common, brown, non-septate | Absent |
| Rhizinomorphs | Scarce or absent | Absent or scarce | Numerous | Absent, often with marginal bristles | Absent or scarce |
| Log Likelihood | Number of Parameters | AICc | AICc Model Weight | |
|---|---|---|---|---|
| DEC | −105.8 | 2 | 216.1 | 1.2 × 10−30 |
| DEC+J | −88.54 | 3 | 184.2 | 1.1 × 10−23 |
| DIVALIKE | −113.4 | 2 | 231.3 | 6.3 × 10−34 |
| DIVALIKE+J | −91.56 | 3 | 190.3 | 5.1 × 10−25 |
| BAYAREALIKE | −37.28 | 2 | 79.12 | 0.70 |
| BAYAREALIKE+J | −36.85 | 3 | 80.85 | 0.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davydov, E.A.; Himelbrant, D.E.; Kuznetsova, E.S.; Stepanchikova, I.S.; Yakovchenko, L.S. Multilocus Molecular Phylogeny of the Umbilicaria aprina Group (Umbilicariaceae, Lichenized Ascomycota) Supports Species Level and Neo-Endemic Status of Umbilicaria krascheninnikovii. Plants 2024, 13, 729. https://doi.org/10.3390/plants13050729
Davydov EA, Himelbrant DE, Kuznetsova ES, Stepanchikova IS, Yakovchenko LS. Multilocus Molecular Phylogeny of the Umbilicaria aprina Group (Umbilicariaceae, Lichenized Ascomycota) Supports Species Level and Neo-Endemic Status of Umbilicaria krascheninnikovii. Plants. 2024; 13(5):729. https://doi.org/10.3390/plants13050729
Chicago/Turabian StyleDavydov, Evgeny A., Dmitry E. Himelbrant, Ekaterina S. Kuznetsova, Irina S. Stepanchikova, and Lidia S. Yakovchenko. 2024. "Multilocus Molecular Phylogeny of the Umbilicaria aprina Group (Umbilicariaceae, Lichenized Ascomycota) Supports Species Level and Neo-Endemic Status of Umbilicaria krascheninnikovii" Plants 13, no. 5: 729. https://doi.org/10.3390/plants13050729