
Foliar Application of Amino Acids and Nutrients as a Tool to Mitigate Water Stress and Stabilize Sugarcane Yield and Bioenergy Generation
 , and
, and
Abstract
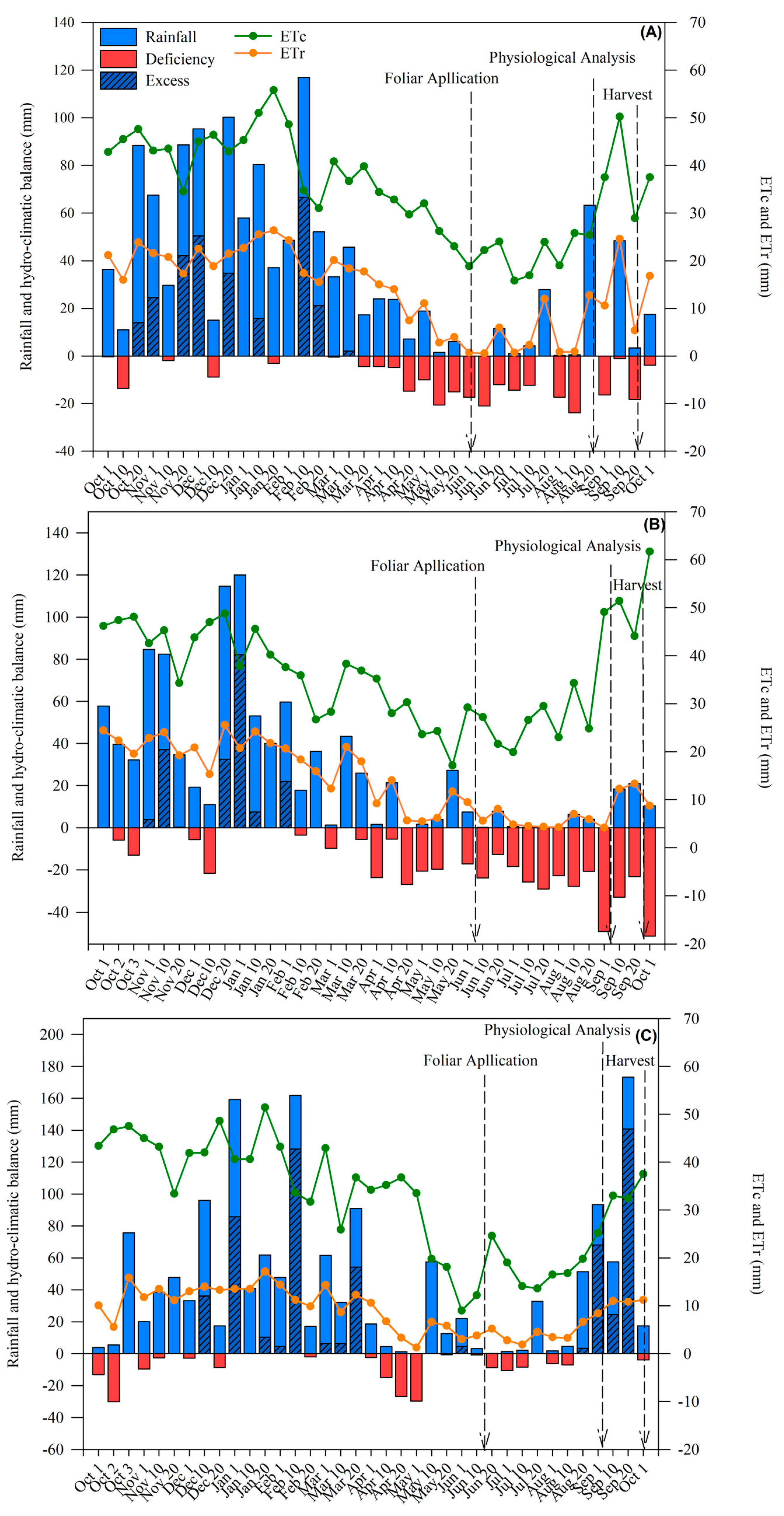
:1. Introduction
2. Results
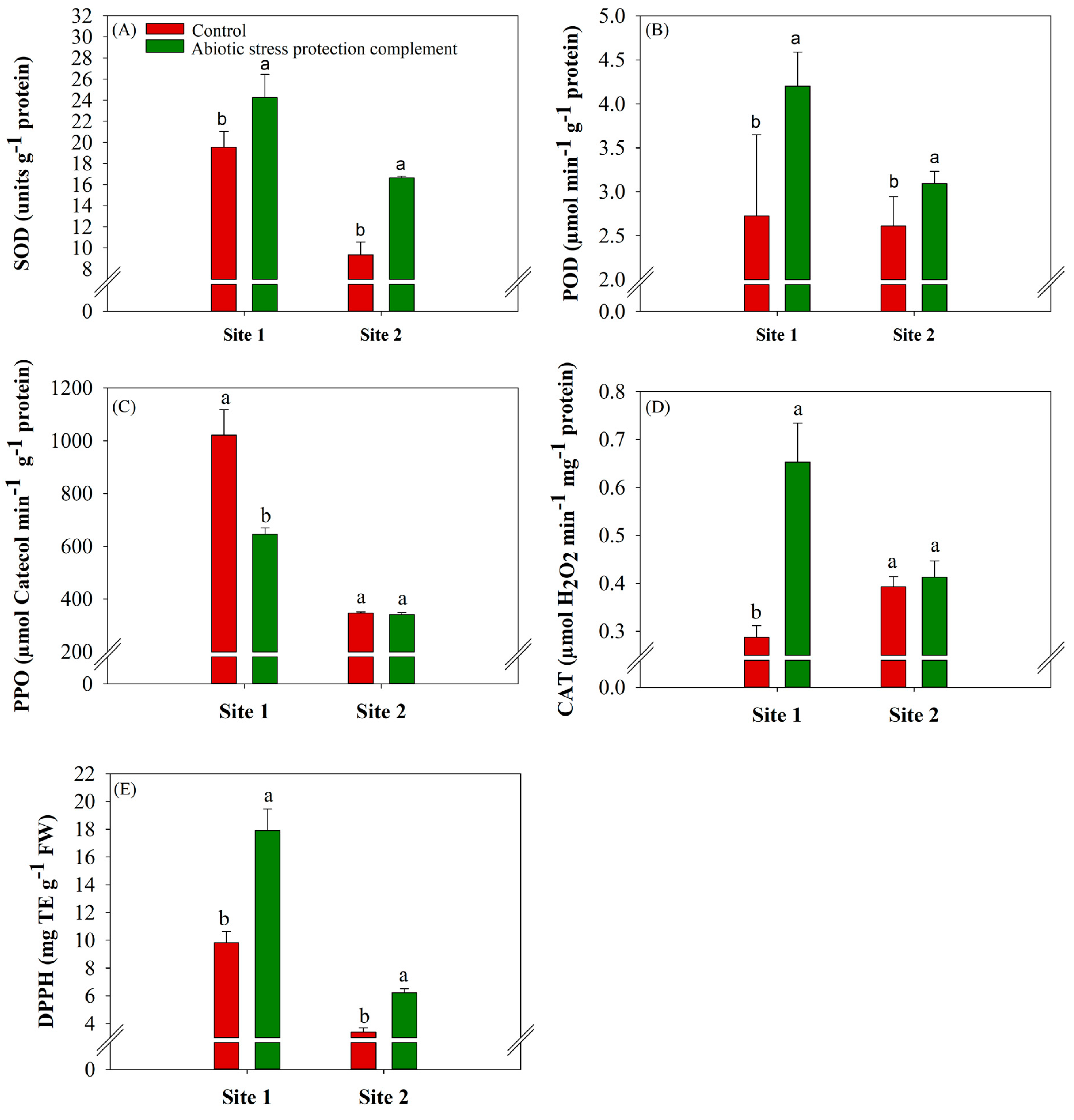
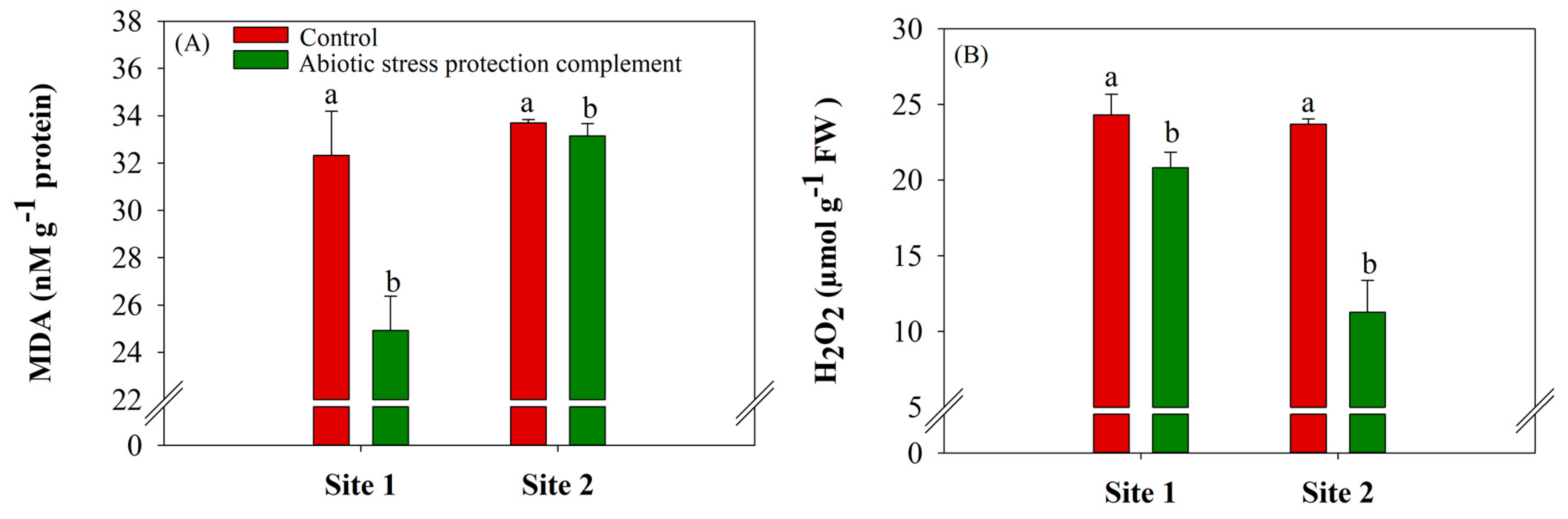
2.1. ASPC Application Increases Antioxidant Enzyme Activity and Reduces Oxidative Stress
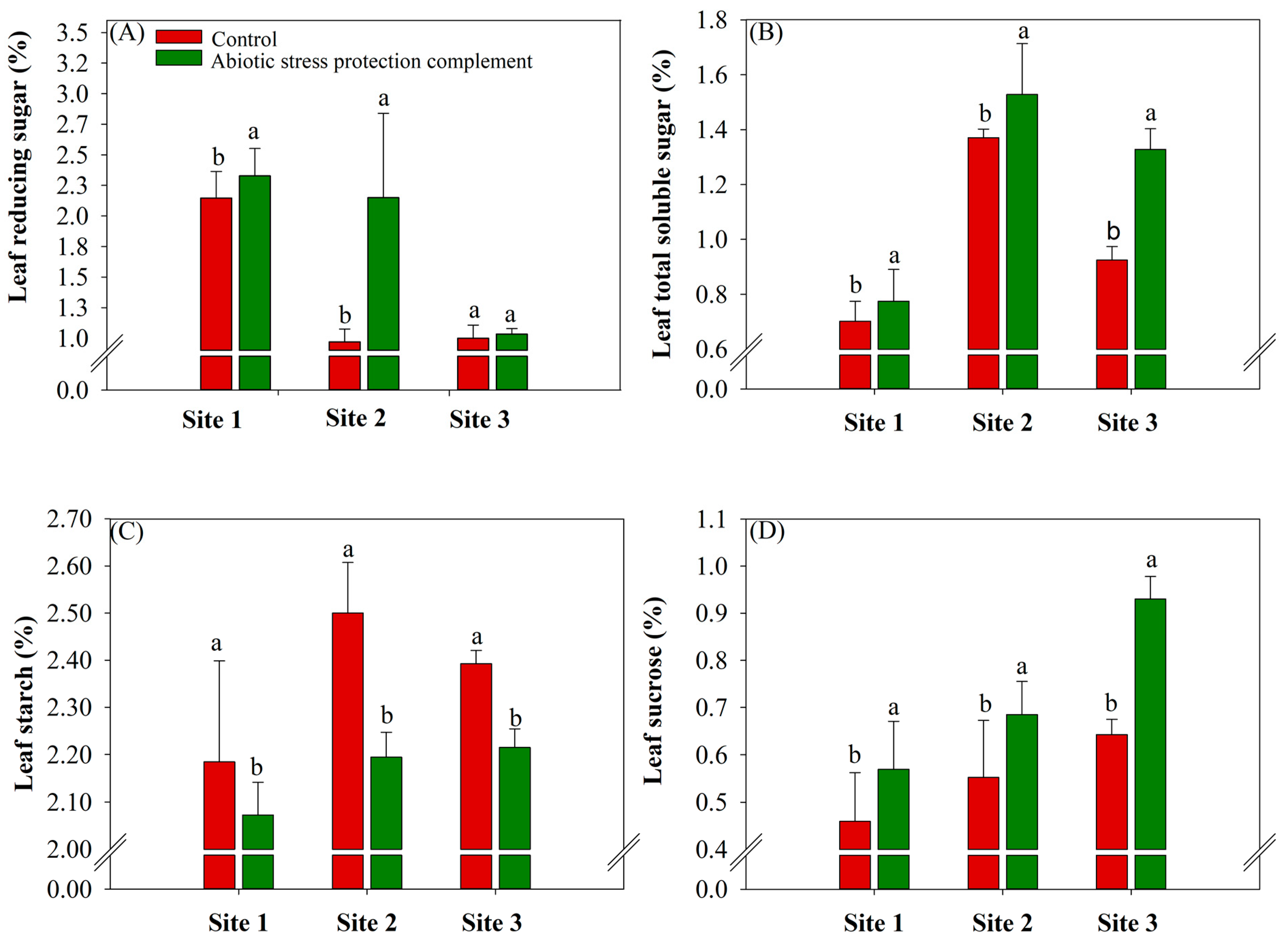
2.2. ASPC Application Increases Sugarcane Leaf Metabolites
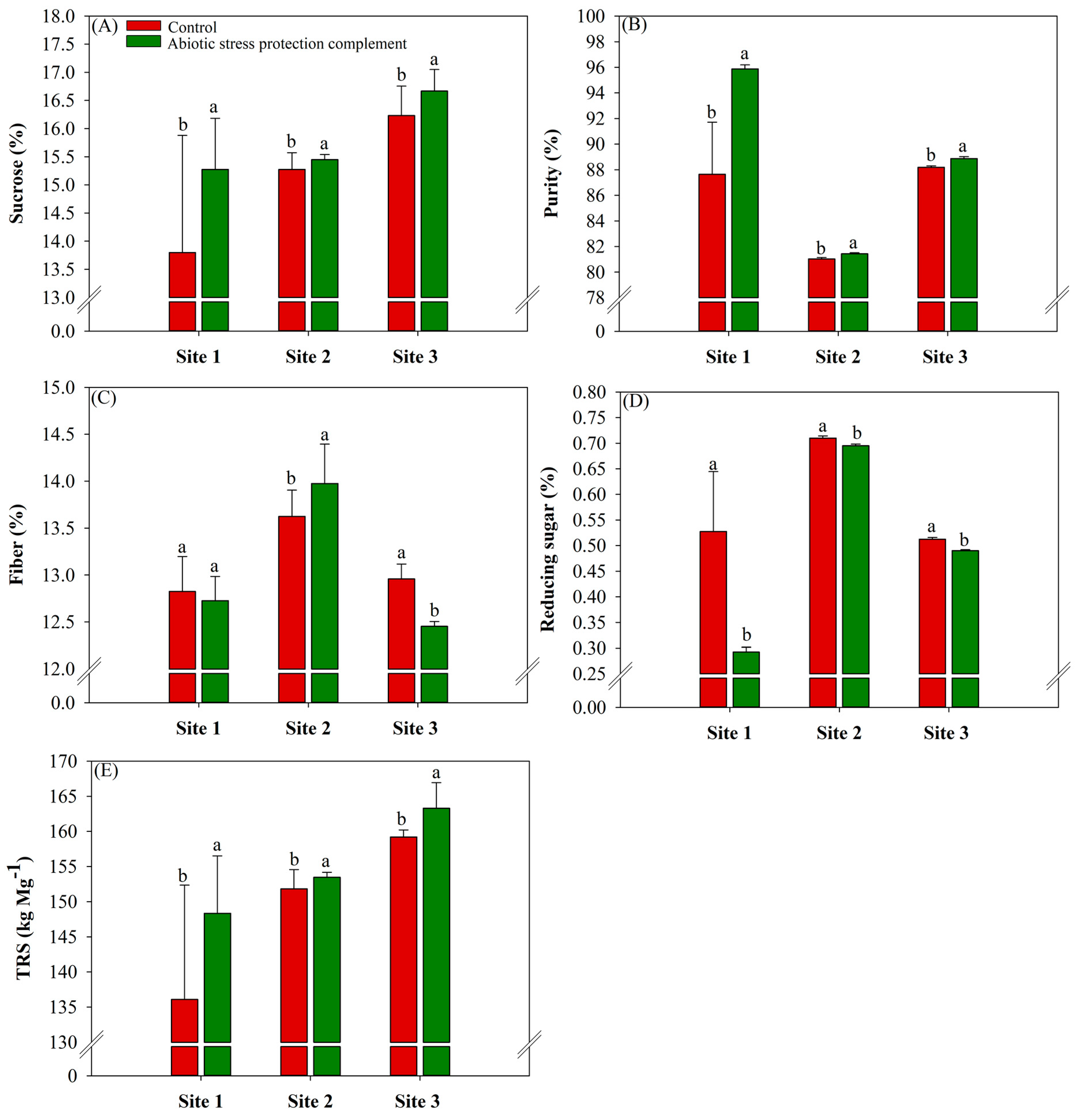
2.3. ASPC Application Improves Sugarcane Stalk Technological Parameters
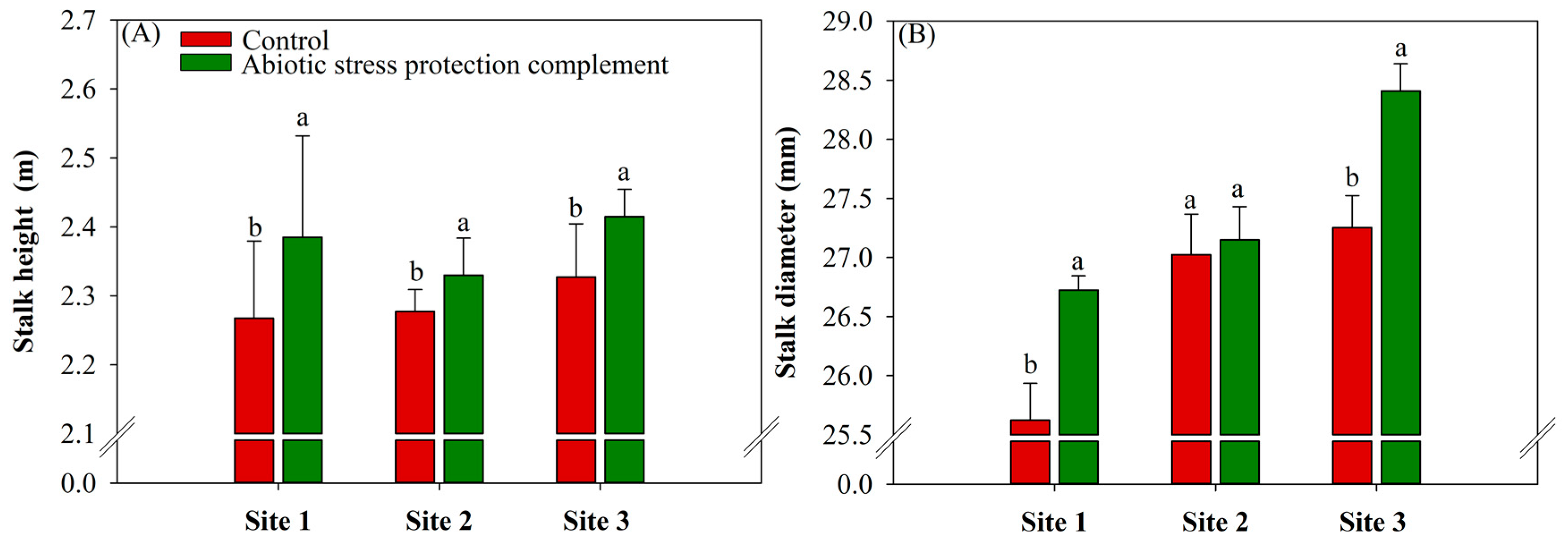
2.4. ASPC Application Increases Sugarcane Biometric Parameters
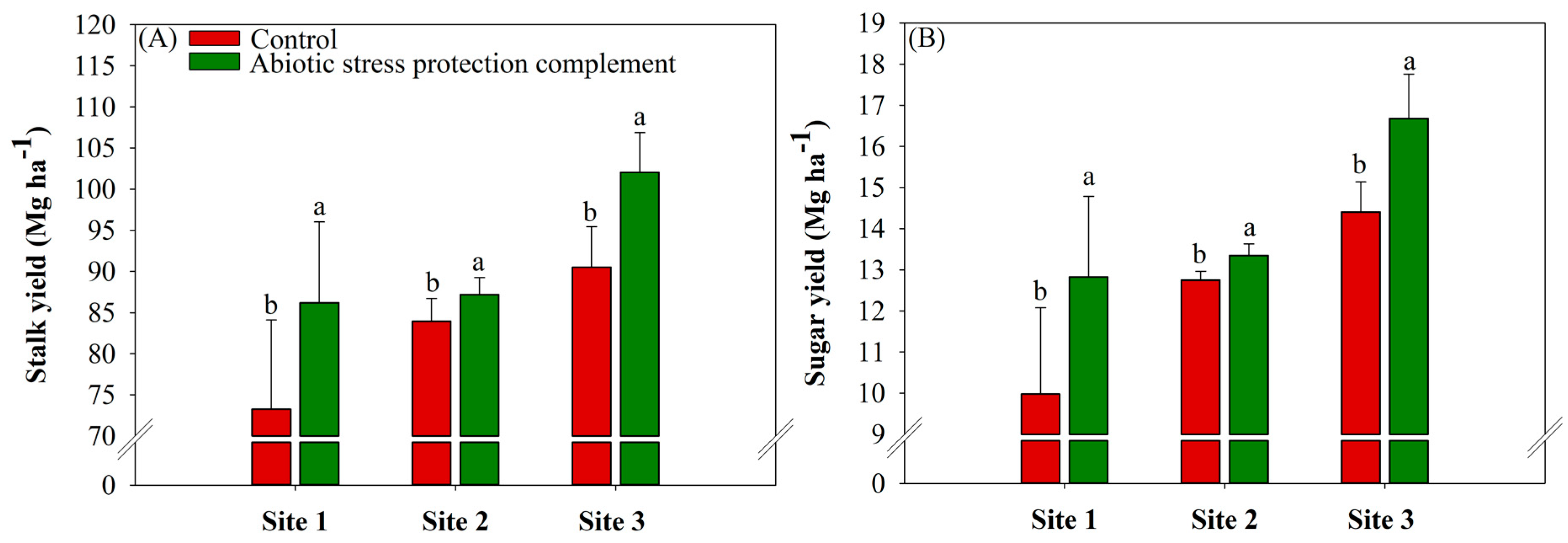
2.5. ASPC Application Increases Stalk and Sugar Yields
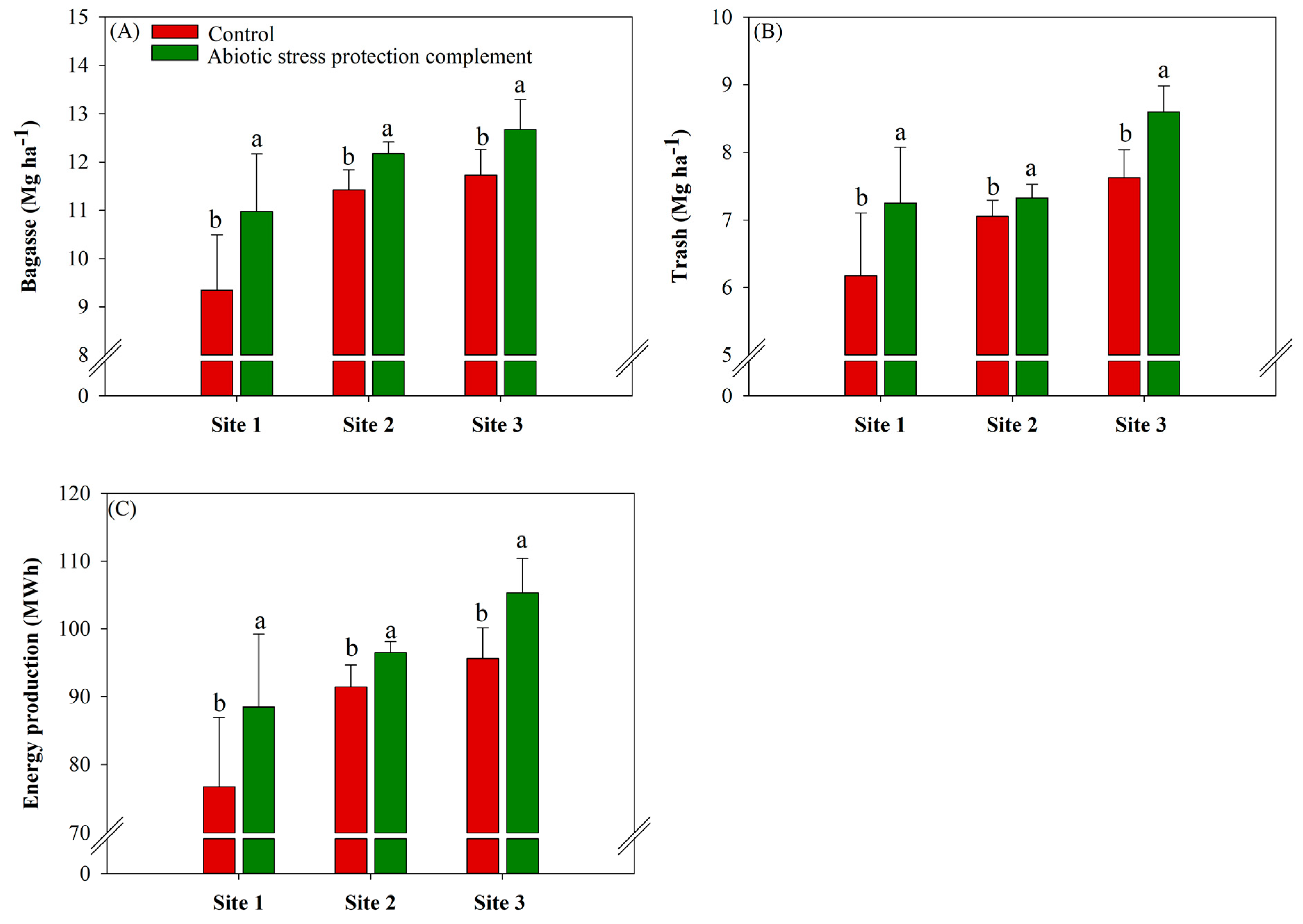
2.6. ASPC Application Increases Biomass and Energy Production
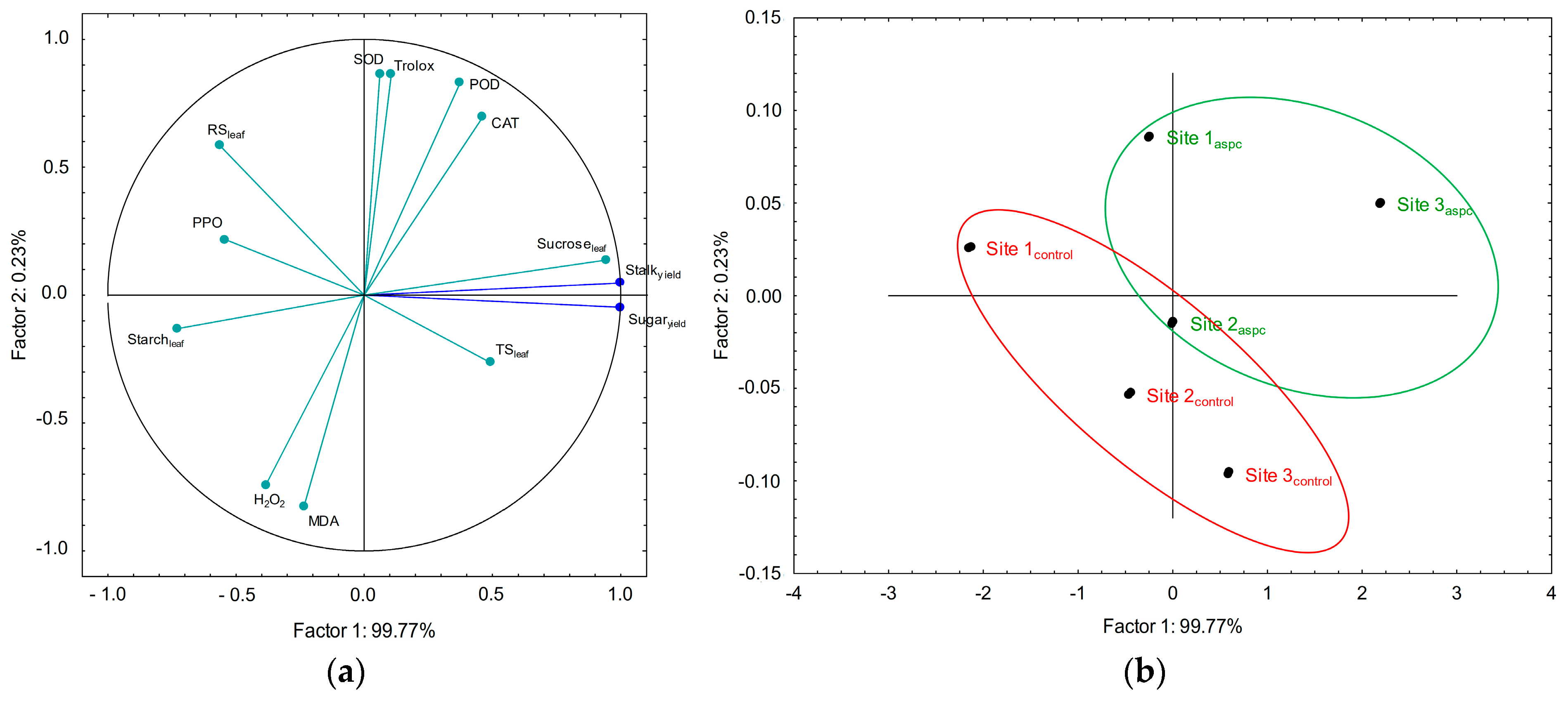
2.7. Principal Component Analysis (PCA) among Sugarcane Parameters
3. Discussion
4. Materials and Methods
4.1. Experimental Area
4.2. Experimental Design and Description of the Treatments
4.3. Leaf Collection for Enzyme Sampling
4.4. Metabolic Changes in Sugarcane Leaves
4.5. Oxidative Stress and Antioxidant Enzymes
4.6. Sugarcane Biometric Evaluations
4.7. Sugarcane Quality Evaluations
4.8. Stalk and Sugar Yield
4.9. Biomass and Energy Production
4.10. Principal Component Analysis (PCA) Data Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Dias, C.M.; Corsato, C.E.; dos Santos, V.M.; Santos, A.F.S. Indicadores Fito Técnicos de Produção Agroindustrial Em Cana-de-Açúcar Cultivada Sob Dois Regimes Hídricos. Rev. Caatinga 2012, 25, 58–65. [Google Scholar]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Núñez-Delgado, A.; Sultana, S.; Wang, J.; Munir, A.; Battaglia, M.L.; Sarker, T.; Seleiman, M.F.; Barmon, M.; Zhang, R. Additions of Optimum Water, Spent Mushroom Compost and Wood Biochar to Improve the Growth Performance of Althaea Rosea in Drought-Prone Coal-Mined Spoils. J. Environ. Manag. 2021, 295, 113076. [Google Scholar] [CrossRef] [PubMed]
- Bordonal, R.D.O.; Carvalho, J.L.N.; Lal, R.; de Figueiredo, E.B.; de Oliveira, B.G.; La Scala, N. Sustainability of Sugarcane Production in Brazil. A Review. Agron. Sustain. Dev. 2018, 38, 13. [Google Scholar] [CrossRef]
- Chiconato, D.A.; da Silveira Sousa Junior, G.; dos Santos, D.M.M.; Munns, R. Adaptation of Sugarcane Plants to Saline Soil. Environ. Exp. Bot. 2019, 162, 201–211. [Google Scholar] [CrossRef]
- Flack-Prain, S.; Shi, L.; Zhu, P.; da Rocha, H.R.; Cabral, O.; Hu, S.; Williams, M. The Impact of Climate Change and Climate Extremes on Sugarcane Production. GCB Bioenergy 2021, 13, 408–424. [Google Scholar] [CrossRef]
- dos Santos, C.M.; Endres, L.; da Silva, A.C.S.; Silva, J.V.; de Souza Barbosa, G.V.; Froehlich, A.; Teixeira, M.M. Water Relations and Osmolite Accumulation Related to Sugarcane Yield under Drought Stress in a Tropical Climate. Int. J. Plant Prod. 2019, 13, 227–239. [Google Scholar] [CrossRef]
- Jacomassi, L.M.; Viveiros, J.D.O.; Oliveira, M.P.; Momesso, L.; de Siqueira, G.F.; Crusciol, C.A.C. A Seaweed Extract-Based Biostimulant Mitigates Drought Stress in Sugarcane. Front. Plant Sci. 2022, 13, 865291. [Google Scholar] [CrossRef] [PubMed]
- Brunini, R.G.; Turco, J.E.P. Water Stress Index on Sugarcane in Different Developmental Phases. Ciênc. E Agrotecnol. 2018, 42, 204–215. [Google Scholar] [CrossRef]
- Vianna, M.D.S.; Sentelhas, P.C. Simulação Do Risco de Deficit Hídrico Em Regiões de Expansão Do Cultivo de Cana-de-Açúcar No Brasil. Pesqui. Agropecu. Bras. 2014, 49, 237–246. [Google Scholar] [CrossRef]
- FAOStat Land and Water: Sugarcane-Food and Agriculture Organization of the United Nations [WWW Document]. Available online: https://www.fao.org/land-water/databases-and-software/crop-information/sugarcane/en/ (accessed on 11 July 2022).
- Marcos, F.C.C.; Silveira, N.M.; Marchiori, P.E.R.; Machado, E.C.; Souza, G.M.; Landell, M.G.A.; Ribeiro, R.V. Drought Tolerance of Sugarcane Propagules Is Improved When Origin Material Faces Water Deficit. PLoS ONE 2018, 13, e0206716. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.A.; Jifon, J.L.; Sharma, V.; da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of Physiological Parameters in Screening Drought Tolerance in Sugarcane Genotypes. Sugar. Tech. 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Vieira, G.H.S.; Mantovani, E.C.; Sediyama, G.C.; Delazari, F.T. Indicadores Morfo-Fisiológicos Do Estresse Hídrico Para a Cultura Da Cana-de-Açúcar Em Função de Lâminas de Irrigação. Biosci. J. 2014, 30, 65–75. [Google Scholar]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.M.; Shami, A. Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber. Plants 2020, 9, 1783. [Google Scholar] [CrossRef]
- Foyer, C.H.; Baker, A.; Wright, M.; Sparkes, I.A.; Mhamdi, A.; Schippers, J.H.M.; Van Breusegem, F. On the Move: Redox-Dependent Protein Relocation in Plants. J. Exp. Bot. 2020, 71, 620–631. [Google Scholar] [CrossRef]
- Gratão, P.L.; Polle, A.; Lea, P.J.; Azevedo, R.A. Making the Life of Heavy Metal-Stressed Plants a Little Easier. Funct. Plant Biol. 2005, 32, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Dustgeer, Z.; Seleiman, M.F.; Khan, I.; Chattha, M.U.; Ali, E.F.; Alhammad, B.A.; Jalal, R.S.; Refay, Y.; Hassan, M.U. Glycine-Betaine Induced Salinity Tolerance in Maize by Regulating the Physiological Attributes, Antioxidant Defense System and Ionic Homeostasis. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12248. [Google Scholar] [CrossRef]
- Gavelienė, V.; Novickienė, L.; Pakalniškytė, L. Effect of Auxin Physiological Analogues on Rapeseed (Brassica napus) Cold Hardening, Seed Yield and Quality. J. Plant Res. 2013, 126, 283–292. [Google Scholar] [CrossRef]
- Khan, I.; Seleiman, M.F.; Chattha, M.U.; Jalal, R.S.; Mahmood, F.; Hassan, F.A.S.; Izzet, W.; Alhammad, B.A.; Ali, E.F.; Roy, R.; et al. Enhancing Antioxidant Defense System of Mung Bean with a Salicylic Acid Exogenous Application to Mitigate Cadmium Toxicity. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12303. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Iwaniuk, P.; Łuniewski, S.; Kaczyński, P.; Łozowicka, B. The Influence of Humic Acids and Nitrophenols on Metabolic Compounds and Pesticide Behavior in Wheat under Biotic Stress. Agronomy 2023, 13, 1378. [Google Scholar] [CrossRef]
- Iwaniuk, P.; Kaczyński, P.; Pietkun, M.; Łozowicka, B. Evaluation of Titanium and Silicon Role in Mitigation of Fungicides Toxicity in Wheat Expressed at the Level of Biochemical and Antioxidant Profile. Chemosphere 2022, 308, 136284. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, F. Fisiologia e Desenvolvimento Vegetal; Artmed Editora: Porto Alegre, Brazil, 2017. [Google Scholar]
- Floss, E.L.; Floss, L.G. Fertilizantes Organominerais de Última Geração: Funções Fisiológicas e Uso Na Agricultura. Rev. Plantio. Direto. 2007, 100, 26–29. [Google Scholar] [CrossRef]
- Batista-Silva, W.; Heinemann, B.; Rugen, N.; Nunes-Nesi, A.; Araújo, W.L.; Braun, H.P.; Hildebrandt, T.M. The Role of Amino Acid Metabolism during Abiotic Stress Release. Plant. Cell Environ. 2019, 42, 1630–1644. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Saleem, M.F.; Wang, L.-C.; Bilal, M.F.; Saeed, A. Protective Role of Glycinebetaine in Maize against Drought-Induced Lipid Peroxidation by Enhancing Capacity of Antioxidative System. Aust. J. Crop Sci. 2012, 6, 576–583. [Google Scholar]
- Philippe, F.; Verdu, I.; Morère-Le Paven, M.C.; Limami, A.M.; Planchet, E. Involvement of Medicago Truncatula Glutamate Receptor-like Channels in Nitric Oxide Production under Short-Term Water Deficit Stress. J. Plant Physiol. 2019, 236, 1–6. [Google Scholar] [CrossRef]
- Russo, R.O.; Berlyn, G.P. The Use of Organic Biostimulants to Help Low Input Sustainable Agriculture. J. Sustain. Agric. 2008, 1, 19–42. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Fageria, N.K.; Filho, M.P.B.; Moreira, A.; Guimarães, C.M. Foliar Fertilization of Crop Plants. J. Plant Nutr. 2009, 32, 1044–1064. [Google Scholar] [CrossRef]
- Eid, M.A.M.; Abdel-Salam, A.A.; Salem, H.M.; Mahrous, S.E.; Seleiman, M.F.; Alsadon, A.A.; Solieman, T.H.I.; Ibrahim, A.A. Interaction Effects of Nitrogen Source and Irrigation Regime on Tuber Quality, Yield, and Water Use Efficiency of Solanum tuberosum L. Plants 2020, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F. Use of Plant Nutrients in Improving Abiotic Stress Tolerance in Wheat. In Wheat Production in Changing Environments; Springer: Singapore, 2019; pp. 481–495. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A.; Almutairi, K.F.; Al-Saif, A.M. Biochar and Its Broad Impacts in Soil Quality and Fertility, Nutrient Leaching and Crop Productivity: A Review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Mantovani, A.; Ribeiro, F.J.; Veiga, M.; Zilio, M.; Felicio, T.P. Métodos de Aplicação de Potássio Na Soja Em Nitossolo Vermelho|Unoesc & Ciência-ACBS. UnoescCiência 2017, 8, 169–176. [Google Scholar]
- Rodrigues, J.D.; Jadoski, C.J.; Fagan, E.B.; Ono, E.O.; Soares, L.H.; Dourado Neto, D. Fisiologia Da Produção de Cana-de-Açúcar; Andrei Editora: São Paulo, Brazil, 2018. [Google Scholar]
- Arsic, M.; Tougaard, S.L.; Persson, D.P.; Martens, H.J.; Doolette, C.L.; Lombi, E.; Schjoerring, J.K.; Husted, S. Bioimaging Techniques Reveal Foliar Phosphate Uptake Pathways and Leaf Phosphorus Status. Plant Physiol. 2020, 183, 11. [Google Scholar] [CrossRef]
- Arsic, M.; Persson, D.P.; Schjoerring, J.K.; Thygesen, L.G.; Lombi, E.; Doolette, C.L.; Husted, S. Foliar-Applied Manganese and Phosphorus in Deficient Barley: Linking Absorption Pathways and Leaf Nutrient Status. Physiol. Plant. 2022, 174, 15. [Google Scholar] [CrossRef]
- Ngoroyemoto, N.; Gupta, S.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Effect of Organic Biostimulants on the Growth and Biochemical Composition of Amaranthus hybridus L. S. Afr. J. Bot. 2019, 124, 87–93. [Google Scholar] [CrossRef]
- Silva, M.A.D.; Cavalcante, Í.H.L.; Mudo, L.E.D.; Paiva Neto, V.B.D.; E Amariz, R.A.; da Cunha, J.G. Bioestimulante Alivia o Estresse Abiótico Da Mangueira Cultivada Em Ambiente Semiárido. Rev. Bras. Eng. Agrícola Ambient. 2020, 24, 457–464. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 1st ed.; Sinauer: Sunderland, MA, USA, 2005; Volume 92, ISBN 0878931724. [Google Scholar]
- Marschner, P. Mineral Nutrition of Higher Plants: Third Edition; Academic Press/Elsevier Inc.: London, UK, 2012; ISBN 9780123849052. [Google Scholar]
- Kheir, A.M.S.; Alkharabsheh, H.M.; Seleiman, M.F.; Al-Saif, A.M.; Ammar, K.A.; Attia, A.; Zoghdan, M.G.; Shabana, M.M.A.; Aboelsoud, H.; Schillaci, C. Calibration and Validation of AQUACROP and APSIM Models to Optimize Wheat Yield and Water Saving in Arid Regions. Land 2021, 10, 1375. [Google Scholar] [CrossRef]
- Al-Selwey, W.A.; Alsadon, A.A.; Alenazi, M.M.; Tarroum, M.; Ibrahim, A.A.; Ahmad, A.; Osman, M.; Seleiman, M.F. Morphological and Biochemical Response of Potatoes to Exogenous Application of ZnO and SiO2 Nanoparticles in a Water Deficit Environment. Horticulturae 2023, 9, 883. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar Applications of ZnO and SiO2 Nanoparticles Mitigate Water Deficit and Enhance Potato Yield and Quality Traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdelhamid, M.T.; Schmidhalter, U. Efecto de La Aplicación Foliar de Aminoácidos Sobre El Rendimiento y Parámetros Fisiológicos En Plantas de Haba Irrigadas Con Agua de Mar. Acta Biol. Colomb. 2015, 20, 141–152. [Google Scholar] [CrossRef]
- Souri, M.K. Aminochelate Fertilizers: The New Approach to the Old Problem; a Review. Open Agric. 2016, 1, 118–123. [Google Scholar] [CrossRef]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M. Reactive Oxygen Species (ROS): Beneficial Companions of Plants’ Developmental Processes. Front. Plant Sci. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of Potassium and Magnesium in Photosynthesis, Photosynthate Translocation and Photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Kameritsch, P. Relative Association of Rubisco with Manganese and Magnesium as a Regulatory Mechanism in Plants. Physiol. Plant. 2017, 161, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Glaz, B.; Comstock, J.C. Sugarcane Leaf Photosynthesis and Growth Characters during Development of Water-Deficit Stress. Crop Sci. 2013, 53, 1066–1075. [Google Scholar] [CrossRef]
- Kollist, H.; Nuhkat, M.; Roelfsema, M.R.G. Closing Gaps: Linking Elements That Control Stomatal Movement. New Phytol. 2014, 203, 44–62. [Google Scholar] [CrossRef] [PubMed]
- Lawson, T.; Flexas, J. Fuelling Life: Recent Advances in Photosynthesis Research. Plant J. 2020, 101, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Vavasseur, A.; Raghavendra, A.S. Guard Cell Metabolism and CO2 Sensing. New Phytol. 2005, 165, 665–682. [Google Scholar] [CrossRef]
- Austin, J.K.; Dunn, D.W.; Caffrey, H.M.; Perkins, S.M.; Harezlak, J.; Rose, D.F. Recurrent Seizures and Behavior Problems in Children with First Recognized Seizures: A Prospective Study. Epilepsia 2002, 43, 1564–1573. [Google Scholar] [CrossRef]
- Nissen, P.; Hansen, J.; Ban, N.; Moore, P.B.; Steitz, T.A. The Structural Basis of Ribosome Activity in Peptide Bond Synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of Temperature Stress by Nutrient Management in Crop Plants: A Review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Yaseen Ashraf, M.; Saifullah, S.; Ahmad, M. Improving Agricultural Water Use Efficiency by Nutrient Management in Crop Plants. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Tóth, B.; Moloi, M.J.; Mousavi, S.M.N.; Illés, Á.; Bojtor, C.; Szőke, L.; Nagy, J. The Evaluation of the Effects of Zn, and Amino Acid-Containing Foliar Fertilizers on the Physiological and Biochemical Responses of a Hungarian Fodder Corn Hybrid. Agron. 2022, 12, 1523. [Google Scholar] [CrossRef]
- Bhattacharjee, S. Sites of Generation and Physicochemical Basis of Formation of Reactive Oxygen Species in Plant Cell. In Reactive Oxygen Species and Antioxidants in Higher Plants; Science Publishers: New York, NY, USA, 2010; pp. 1–30. [Google Scholar] [CrossRef]
- Dinakar, C.; Djilianov, D.; Bartels, D. Photosynthesis in Desiccation Tolerant Plants: Energy Metabolism and Antioxidative Stress Defense. Plant Sci. 2012, 182, 29–41. [Google Scholar] [CrossRef]
- Ken, C.F.; Hsiung, T.M.; Huang, Z.X.; Juang, R.H.; Lin, C.T. Characterization of Fe/Mn-Superoxide Dismutase from Diatom Thallassiosira Weissflogii: Cloning, Expression, and Property. J. Agric. Food Chem. 2005, 53, 1470–1474. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Valero, E.; Sanchez-Ferrer, A.; Varon, R.; García-Carmona, F. Evolution of Grape Polyphenol Oxidase Activity and Phenolic Content during Maturation and Vinification-CORE Reader. Vitis 1989, 28, 85–95. [Google Scholar]
- Hammer, E.F. Oxidoreductores. In Enzymes in Food Processing; Springer: Berlin/Heidelberg, Germany, 1993; pp. 221–247. [Google Scholar]
- Dubey, D.; Pandey, A. Effect of Nickel (Ni) on Chlorophyll, Lipid Peroxidation and Antioxidant Enzymes Activities in Black Gram (Vigna Mungo) Leaves. Int. J. Sci. Nat. 2011, 2, 395–401. [Google Scholar]
- Elansary, H.O.; Skalicka-Woźniak, K.; King, I.W. Enhancing Stress Growth Traits as Well as Phytochemical and Antioxidant Contents of Spiraea and Pittosporum under Seaweed Extract Treatments. Plant Physiol. Biochem. 2016, 105, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on Health Promoting Phytochemicals Following Seaweed Application, in Potato and Onion Crops Grown under a Low Input Agricultural System. Sci. Hortic. 2014, 170, 224–227. [Google Scholar] [CrossRef]
- Spann, T.M.; Little, H.A. Applications of a Commercial Extract of the Brown Seaweed Ascophyllum Nodosum Increases Drought Tolerance in Container-Grown ‘Hamlin’ Sweet Orange Nursery Trees. HortScience 2011, 46, 577–582. [Google Scholar] [CrossRef]
- Carmody, N.; Goñi, O.; Łangowski, Ł.; O’Connell, S. Ascophyllum Nodosum Extract Biostimulant Processing and Its Impact on Enhancing Heat Stress Tolerance during Tomato Fruit Set. Front. Plant Sci. 2020, 11, 807. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Jaiswal, V.P.; Sharma, L.; Pathak, A.D.; Singh, A.K.; Gupta, R.; Awasthi, S.K.; Gaur, A.; Zubair, A.; Tiwari, R. Subsoiling Affecting Soil Quality Parameters and Sugarcane Yield in Multiratooning System in Subtropical India. Commun. Soil Sci. Plant Anal. 2021, 52, 2125–2144. [Google Scholar] [CrossRef]
- Mano, J. Reactive Carbonyl Species: Their Production from Lipid Peroxides, Action in Environmental Stress, and the Detoxification Mechanism. Plant Physiol. Biochem. PPB 2012, 59, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Beckles, D.M. Dynamic Changes in the Starch-Sugar Interconversion within Plant Source and Sink Tissues Promote a Better Abiotic Stress Response. J. Plant Physiol. 2019, 234–235, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, M.; Santelia, D. Starch as a Determinant of Plant Fitness under Abiotic Stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef]
- MacNeill, G.J.; Mehrpouyan, S.; Minow, M.A.A.; Patterson, J.A.; Tetlow, I.J.; Emes, M.J. Starch as a Source, Starch as a Sink: The Bifunctional Role of Starch in Carbon Allocation. J. Exp. Bot. 2017, 68, 4433–4453. [Google Scholar] [CrossRef]
- Zanella, M.; Borghi, G.L.; Pirone, C.; Thalmann, M.; Pazmino, D.; Costa, A.; Santelia, D.; Trost, P.; Sparla, F. β-Amylase 1 (BAM1) Degrades Transitory Starch to Sustain Proline Biosynthesis during Drought Stress. J. Exp. Bot. 2016, 67, 1819–1826. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its Metabolism, Evolution, and Biotechnological Modification in Plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef]
- Merwad, A.R.M.A.; Desoky, E.S.M.; Rady, M.M. Response of Water Deficit-Stressed Vigna Unguiculata Performances to Silicon, Proline or Methionine Foliar Application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Souza, J.L.; Barbosa, R.; Nunes, V.V.; Torres, F.O.; Calazans, C.C.; Fernando, L.; Oliveira, G.; Silva-mann, R. Déficit Hídrico No Desenvolvimento de Cultivares de Cana-de-Açúcar. Glob. Sci. Technol. 2020, 13, 196–210. [Google Scholar]
- Diwen, C.; Wenling, Z.; Junhua, A.; Zhenrui, H.; Shixiang, B.; Xiaoxiao, Z.; Hong, S. Alleviation Effect of Foliar Application with Seaweed Extract on Sugarcane under Drought Stress. Chin. J. Trop. Crop 2021, 42, 1348. [Google Scholar] [CrossRef]
- Chen, D.; Zhou, W.; Yang, J.; Ao, J.; Huang, Y.; Shen, D.; Jiang, Y.; Huang, Z.; Shen, H. Effects of Seaweed Extracts on the Growth, Physiological Activity, Cane Yield and Sucrose Content of Sugarcane in China. Front. Plant Sci. 2021, 12, 865. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.C. Cálculos Na Agroindústria Da Cana-de-Açúcar, 3rd ed.; STAB: Piracicaba, Brazil, 2011; p. 416. [Google Scholar]
- Silva, D.P.; Oliveira, M.P.; Oliveira, J.A.V.; Jacomassi, L.M.; Momesso, L.; Garcia, A.; Ferraz de Siqueira, G.; Foltran, R.; Soratto, R.P.; Dinardo-Miranda, L.L.; et al. Phytotonic Effects of Thiamethoxam on Sugarcane Managed with Glyphosate as a Ripener. Pest Manag. Sci. 2022, 78, 4006–4017. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, N.P.; Sentelhas, P.C. Climatic Effects on Sugarcane Ripening under the Infl Uence of Cultivars and Crop Age. Sci. Agric. 2013, 70, 449–456. [Google Scholar] [CrossRef]
- Sadak, M.S.; Bakry, B.A.; Abdel-Razik, T.M.; Hanafy, R.S. Amino acids foliar application for maximizing growth, productivity and quality of peanut grown under sandy soil. Braz. J. Biol. 2023, 83, 1–13. [Google Scholar] [CrossRef] [PubMed]
- El-Ghareib, E.A.; El-Sayed, M.A.A.; Mesbah, E.A.E.; Azzam, K.A.A. Effect of Foliar Spraying with Dolfan and Zinc on Yield and Yield Components of Maize (Zea mays L.) Under Different Nitrogen Fertilizer Rates. Middle East J. Agric. Res. 2014, 3, 465–471. [Google Scholar]
- Jamro, G.H.; Kazi, B.R.; Oad, F.C.; Jamali, N.M.; Oad, N.L. Effect of Foliar Application of Micro Nutrients on the Growth Traits of Sugarcane Variety Cp-65/357 (Ratoon Crop). Asian J. Plant Sci. 2002, 1, 462–463. [Google Scholar] [CrossRef]
- Wijma, M.; Lembke, C.G.; Diniz, A.L.; Santini, L.; Zambotti-Villela, L.; Colepicolo, P.; Carneiro, M.S.; Souza, G.M. Planting Season Impacts Sugarcane Stem Development, Secondary Metabolite Levels, and Natural Antisense Transcription. Cells 2021, 10, 3451. [Google Scholar] [CrossRef] [PubMed]
- Pammenter, N.W.; Allison, J.C.S. Effects of Treatments Potentially Influencing the Supply of Assimilate on Its Partitioning in Sugarcane. J. Exp. Bot. 2002, 53, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Hassuani, S. Biomass Power Generation: Sugar Cane Bagasse and Trash; CTC: Piracicaba, Brazil, 2005. [Google Scholar]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Leonardo, J.; Gonçalves, M.; Sparovek, G. Köppen’s Climate Classification Map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Ditzler, C.; Scheffe, K.; Monger, H.C.; Soil Survey Staff. Keys to Soil Taxonomy; Soil Conservation Service Government Printing Office: Washington, DC, USA, 2014; p. 603.
- Van Dlllewijn, C. Botany of Sugarcane; Waltham, M., Sons, W.D., Eds.; Chronica Botanica: London, UK, 1952. [Google Scholar]
- Nelson, N. A Photometric Adaptation of the Somogyi Method for the Determination of Glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Somogyi, M. A New Reagent for the Determination of Sugars. J. Biol. Chem. 1945, 160, 61–68. [Google Scholar] [CrossRef]
- Rodrigues Ferreira, R.; Francisco Fornazier, R.; Pierre Vitória, A.; Lea, P.J.; Antunes Azevedo, R. Changes in antioxidant enzyme activities in soybean under cadmium stress. J. Plant Nutr. 2002, 25, 327–342. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The Effect of Drought and Ultraviolet Radiation on Growth and Stress Markers in Pea and Wheat. Plant, Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: II. Purification and Quantitative Relationship with Water-Soluble Protein in Seedlings 1,2. Plant Physiol. 1977, 59, 315. [Google Scholar] [CrossRef] [PubMed]
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Allain, C.C.; Poon, L.S.; Chan, C.S.G.; Richmond, W.; Fu, P.C. Enzymatic Determination of Total Serum Cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef]
- Ozgen, M.; Reese, R.N.; Tulio, A.Z.; Scheerens, J.C.; Miller, A.R. Modified 2,2-Azino-Bis-3-Ethylbenzothiazoline-6-Sulfonic Acid (ABTS) Method to Measure Antioxidant Capacity of Selected Small Fruits and Comparison to Ferric Reducing Antioxidant Power (FRAP) and 2,2′-Diphenyl-1-Picrylhydrazyl (DPPH) Methods. J. Agric. Food Chem. 2006, 54, 1151–1157. [Google Scholar] [CrossRef]
- Marafon, A.C. Análise Quantitativa de Crescimento Em Cana-de-Açúcar: Uma Introdução Ao Procedimento Prático; Embrapa Tabuleiros Costeiros: Aracajú, Brazil, 2012. [Google Scholar]
- STATSOFT, Inc. STATISTICA (Data Analysis Software System), Version 7. 2004. Available online: www.statsoft.com (accessed on 19 October 2023).
- Kaiser, H.F. The Varimax Criterion for Analytic Rotation in Factor Analysis. Psychometrika 1958, 23, 187–200. [Google Scholar] [CrossRef]
- Manly, B.F.J.; Navarro Alberto, J.A. Multivariate Statistical Methods: A Primer, Fourth Edition. In Multivariate Statistical Methods; Taylor & Francis Ltd: London, UK, 2016. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eigenvalue | Total Variance % | Cumulative Eigenvalues | Cumulative % | |
|---|---|---|---|---|
| 1 | 1.995 | 99.773 | 1.995 | 99.773 |
| 2 | 0.004 | 0.226 | 2.000 | 100.000 |
| Eigenvectors | Correlations | |||
| Active 2 | Factor 1 | Factor 2 | Factor 1 | Factor 2 |
| Stalk yield | 0.707 | 0.707 | 0.998 | 0.047 |
| Sugar yield | 0.707 | −0.707 | 0.998 | −0.047 |
| Correlations 1 | ||||
| Average of three sites | ||||
| Supplementary 3 | Factor 1 | Factor 2 | ||
| RSleaf | −0.56 | 0.58 | ||
| Starchleaf | −0.72 | −0.13 | ||
| Sucroseleaf | 0.94 | 0.13 | ||
| TSleaf | 0.49 | −0.26 | ||
| SOD | 0.06 | 0.86 | ||
| POD | 0.37 | 0.83 | ||
| PPO | −0.54 | 0.21 | ||
| CAT | 0.46 | 0.70 | ||
| MDA | −0.23 | −0.82 | ||
| Trolox | 0.10 | 0.86 | ||
| H2O2 | −0.38 | −0.74 | ||
| Location | Site | Classification | Depth | pH | SOM | P(resina) | S | Al+3 | H+Al+3 | K | Ca | Mg | CEC | BS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CaCl2 | g dm−3 | mg dm−3 | mmolc dm−3 | % | ||||||||||
| Ponta Porã (MS) 2018 | 1 | Rhodic Eutrodaf | 0.00–0.20 | 5.1 | 34 | 13 | 18 | 1 | 36 | 0.77 | 42 | 11 | 90 | 58 |
| 0.20–0.40 | 4.9 | 29 | 10 | 30 | 5 | 44 | 0.82 | 25 | 7 | 77 | 39 | |||
| Pradópolis (SP) 2019 | 2 | Rhodic Eutrudox | 0.00–0.20 | 5.1 | 26 | 19 | 9 | 0 | 40 | 2.6 | 42 | 15 | 100 | 60 |
| 0.20–0.40 | 5.1 | 23 | 16 | 29 | 0 | 42 | 1.7 | 33 | 15 | 91 | 53 | |||
| Motuca (SP) 2020 | 3 | Hapludox Rhodic | 0.00–0.20 | 5.3 | 17 | 55 | 27 | 0 | 21 | 2.6 | 27 | 11 | 61 | 67 |
| 0.20–0.40 | 5.0 | 14 | 34 | 33 | 1 | 25 | 0.8 | 17 | 6 | 49 | 49 | |||
| Macronutrients | ||||||
|---|---|---|---|---|---|---|
| Mg | K2O | P2O5 | N | S | TOC | |
| Dose 6 L ha−1 (g) | 153.6 | 460.8 | 460.8 | 76.8 | 245.76 | 122.8 |
| Concentration (%) | 2.0 | 6.0 | 6.0 | 1.0 | 3.2 | 1.6 |
| Amino Acids | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gly | Glu | His | Ala | Tau | Pro | Asp | Lys | Arg | Leu | Ser | Thr | Tyr | Val | Phe | Met | Ile | Cys | |
| Dose 6 L ha−1 (g) | 180.5 | 139.8 | 135.2 | 113.7 | 104.5 | 87.6 | 86.8 | 69.9 | 68.4 | 57.6 | 44.5 | 40.7 | 37.6 | 37.6 | 29.9 | 26.9 | 23.8 | 10.8 |
| Concentration (%) | 2.35 | 1.82 | 1.76 | 1.48 | 1.36 | 1.14 | 1.13 | 0.91 | 0.89 | 0.75 | 0.58 | 0.53 | 0.49 | 0.49 | 0.39 | 0.35 | 0.31 | 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacomassi, L.M.; Pacola, M.; Momesso, L.; Viveiros, J.; Júnior, O.A.; Siqueira, G.F.d.; Campos, M.d.; Crusciol, C.A.C. Foliar Application of Amino Acids and Nutrients as a Tool to Mitigate Water Stress and Stabilize Sugarcane Yield and Bioenergy Generation. Plants 2024, 13, 461. https://doi.org/10.3390/plants13030461
Jacomassi LM, Pacola M, Momesso L, Viveiros J, Júnior OA, Siqueira GFd, Campos Md, Crusciol CAC. Foliar Application of Amino Acids and Nutrients as a Tool to Mitigate Water Stress and Stabilize Sugarcane Yield and Bioenergy Generation. Plants. 2024; 13(3):461. https://doi.org/10.3390/plants13030461
Chicago/Turabian StyleJacomassi, Lucas Moraes, Marcela Pacola, Letusa Momesso, Josiane Viveiros, Osvaldo Araújo Júnior, Gabriela Ferraz de Siqueira, Murilo de Campos, and Carlos Alexandre Costa Crusciol. 2024. "Foliar Application of Amino Acids and Nutrients as a Tool to Mitigate Water Stress and Stabilize Sugarcane Yield and Bioenergy Generation" Plants 13, no. 3: 461. https://doi.org/10.3390/plants13030461