
Effects of Different Forms and Proportions of Nitrogen on the Growth, Photosynthetic Characteristics, and Carbon and Nitrogen Metabolism in Tomato

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Design
2.3. Determination of the Indexes and Methods
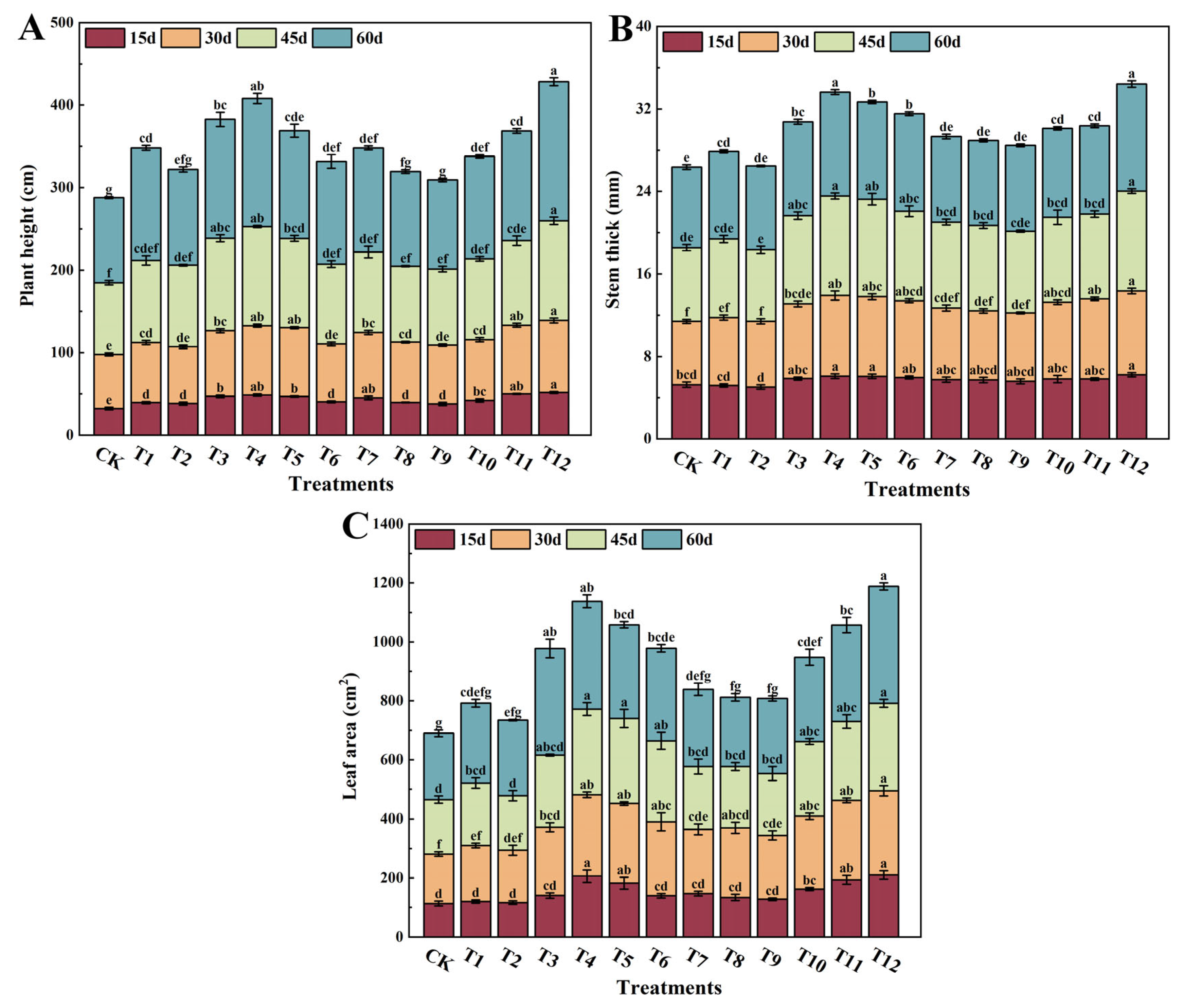
2.3.1. Determination of the Tomato Growth Index
- L: The distance from the petiole base to the leaf tip;
- W: The maximum width perpendicular to the main vein.
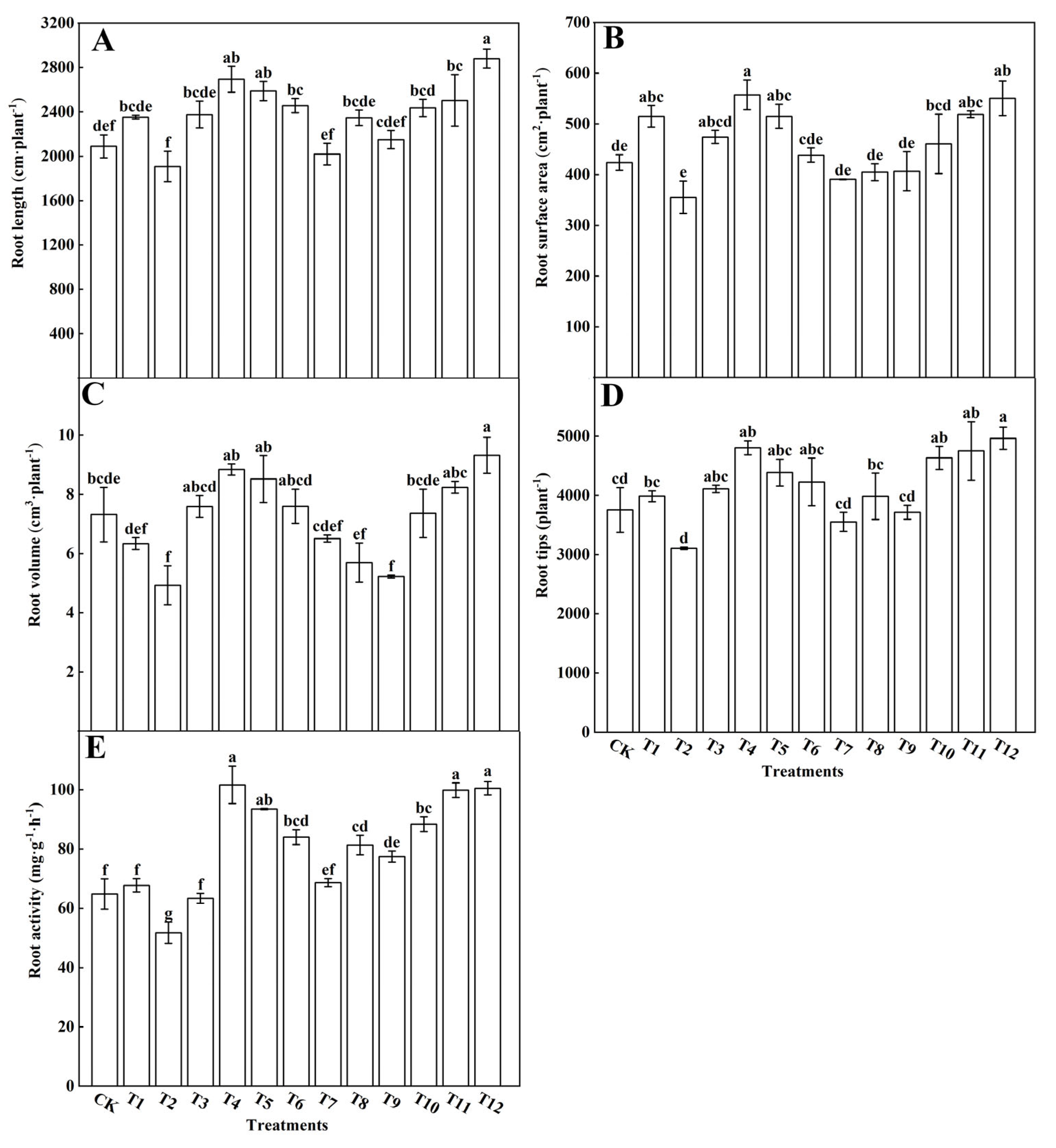
2.3.2. Determination of Root Morphological Parameters
2.3.3. Determination of Gas Exchange Parameters
2.3.4. Determination of Chlorophyll Fluorescence Parameters
2.3.5. Determination of Photosynthate Content
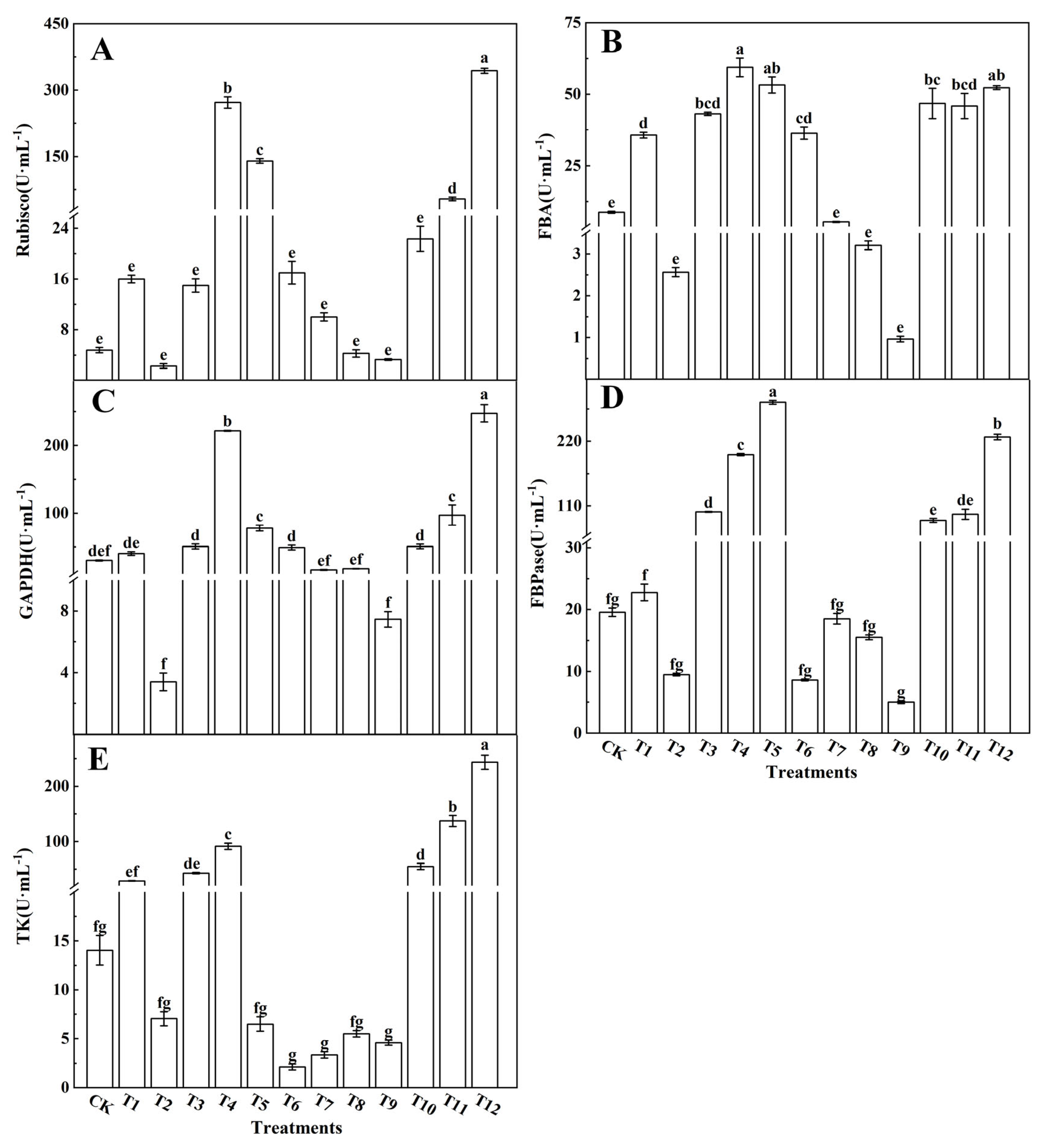
2.3.6. Determination of the Activities of Key Enzymes in the Calvin Cycle, Enzymes Related to Glucose Metabolism, and Enzymes Related to Nitrogen Metabolism
2.3.7. RNA Extraction and RT-qPCR Analysis
2.4. Data Analysis
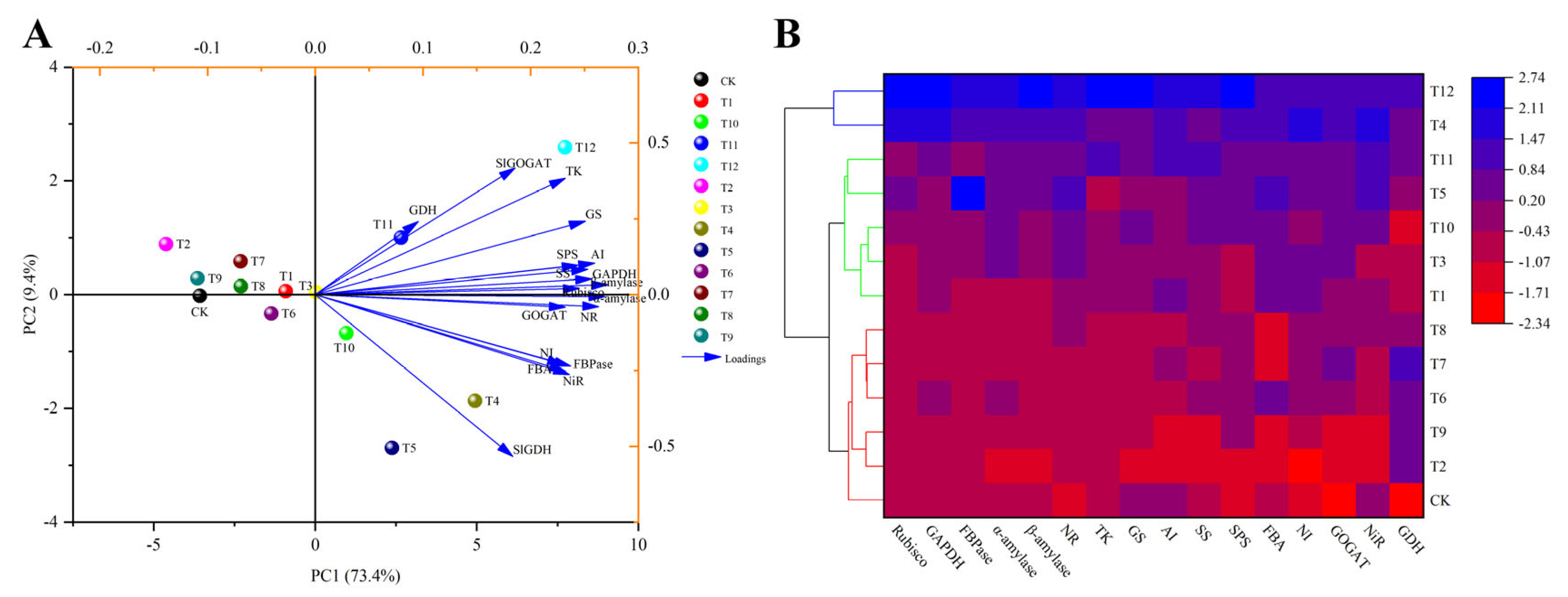
3. Results
3.1. Effects of Different Forms of Nitrogen on Tomato Plant Growth Parameters
3.2. Effects of the Different Forms of Nitrogen on Photosynthesis
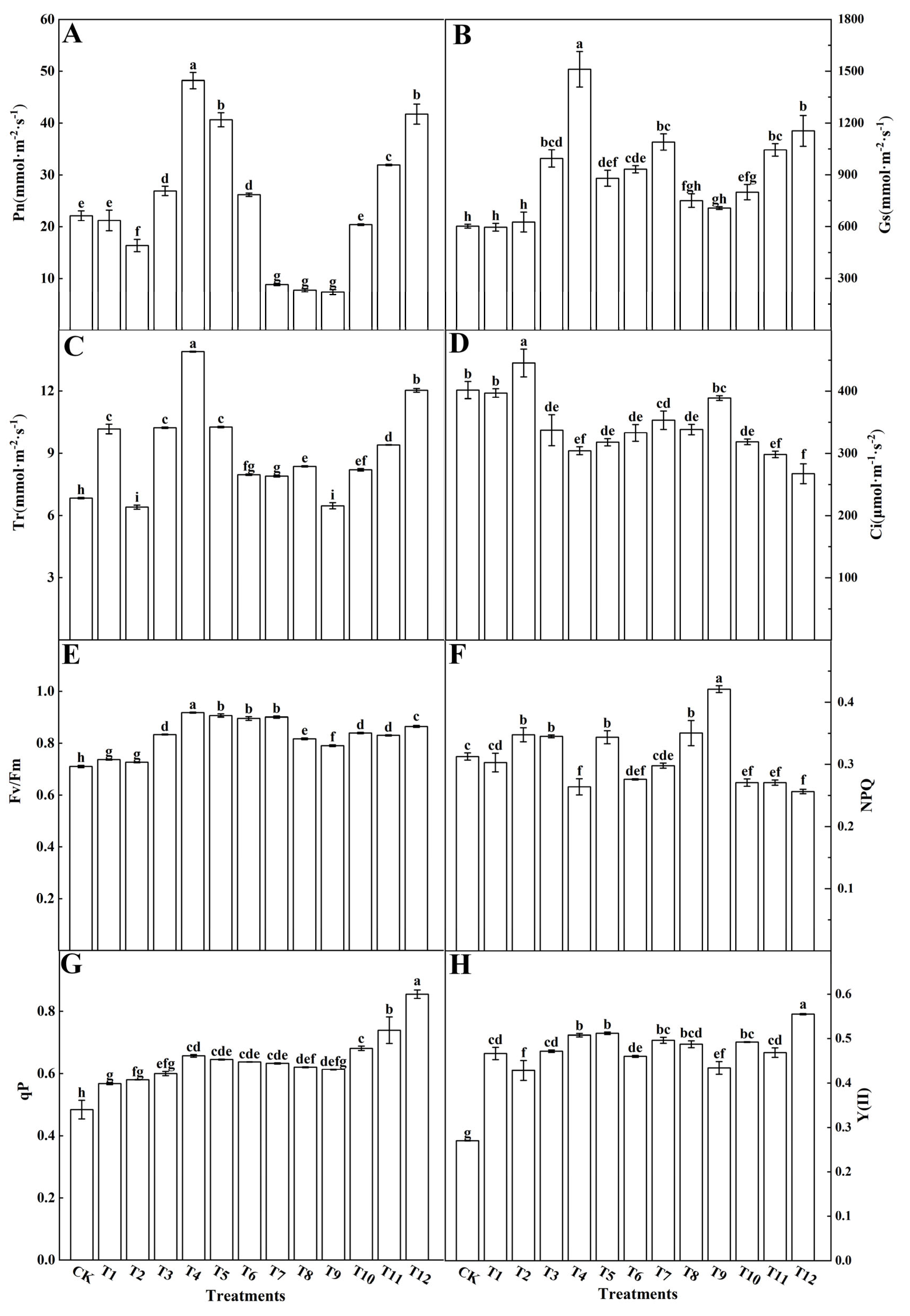
3.2.1. Gas Exchange and Chlorophyll Fluorescence Parameters
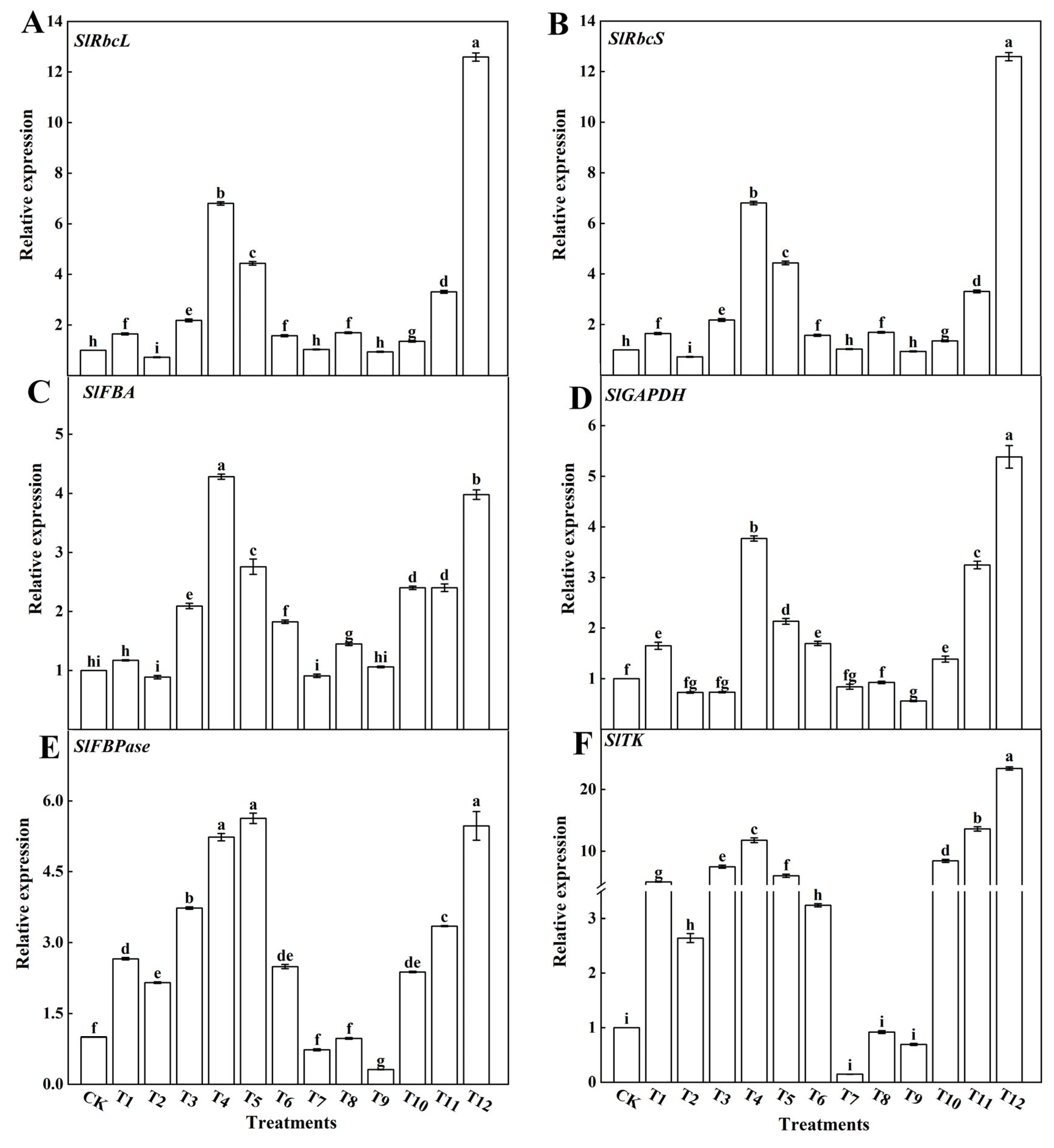
3.2.2. Expression of Key Enzyme Genes from the Calvin Cycle
3.3. Effects of Different Forms of Nitrogen on Sugar Metabolism
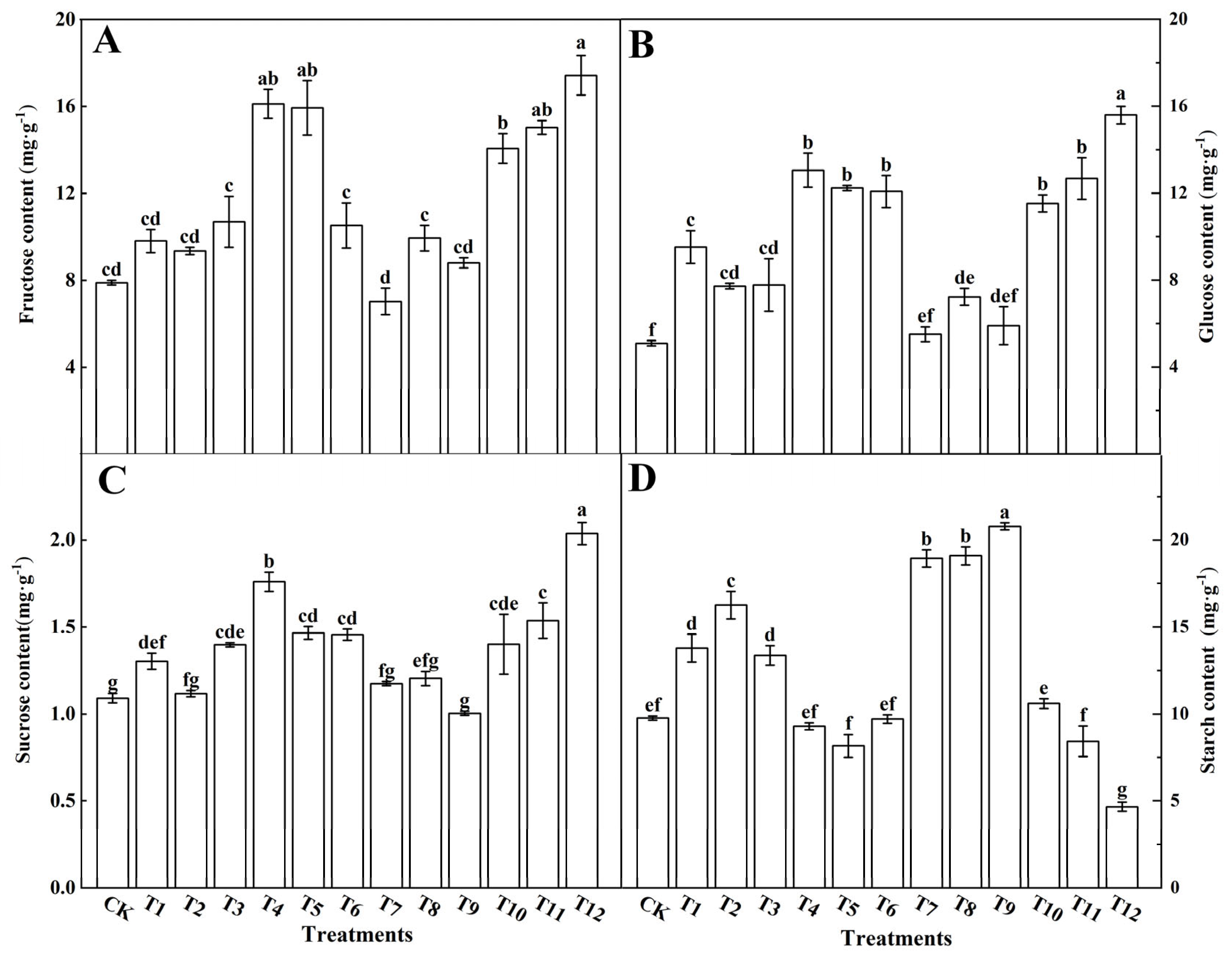
3.3.1. Effects on the Photosynthetic Products
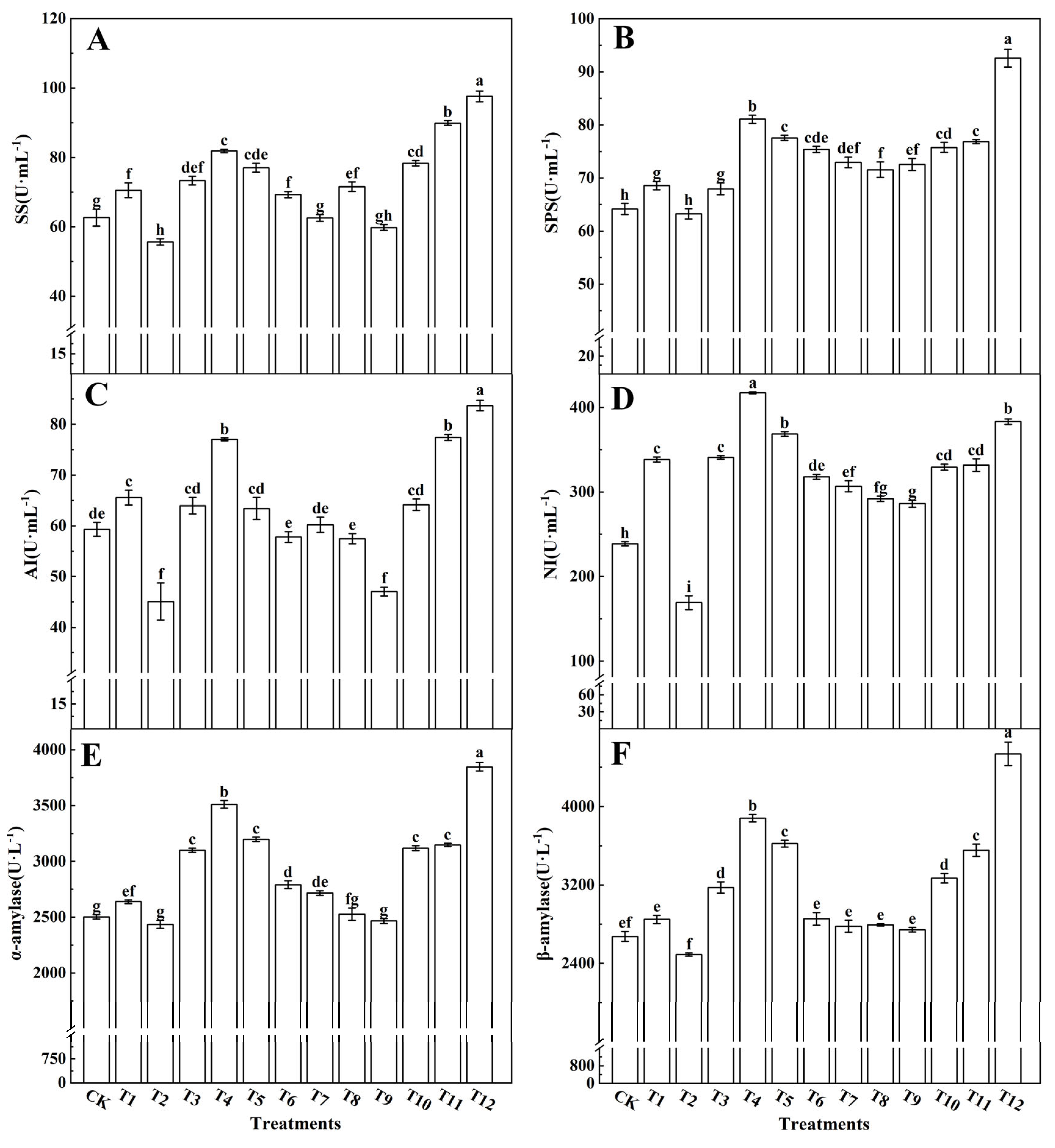
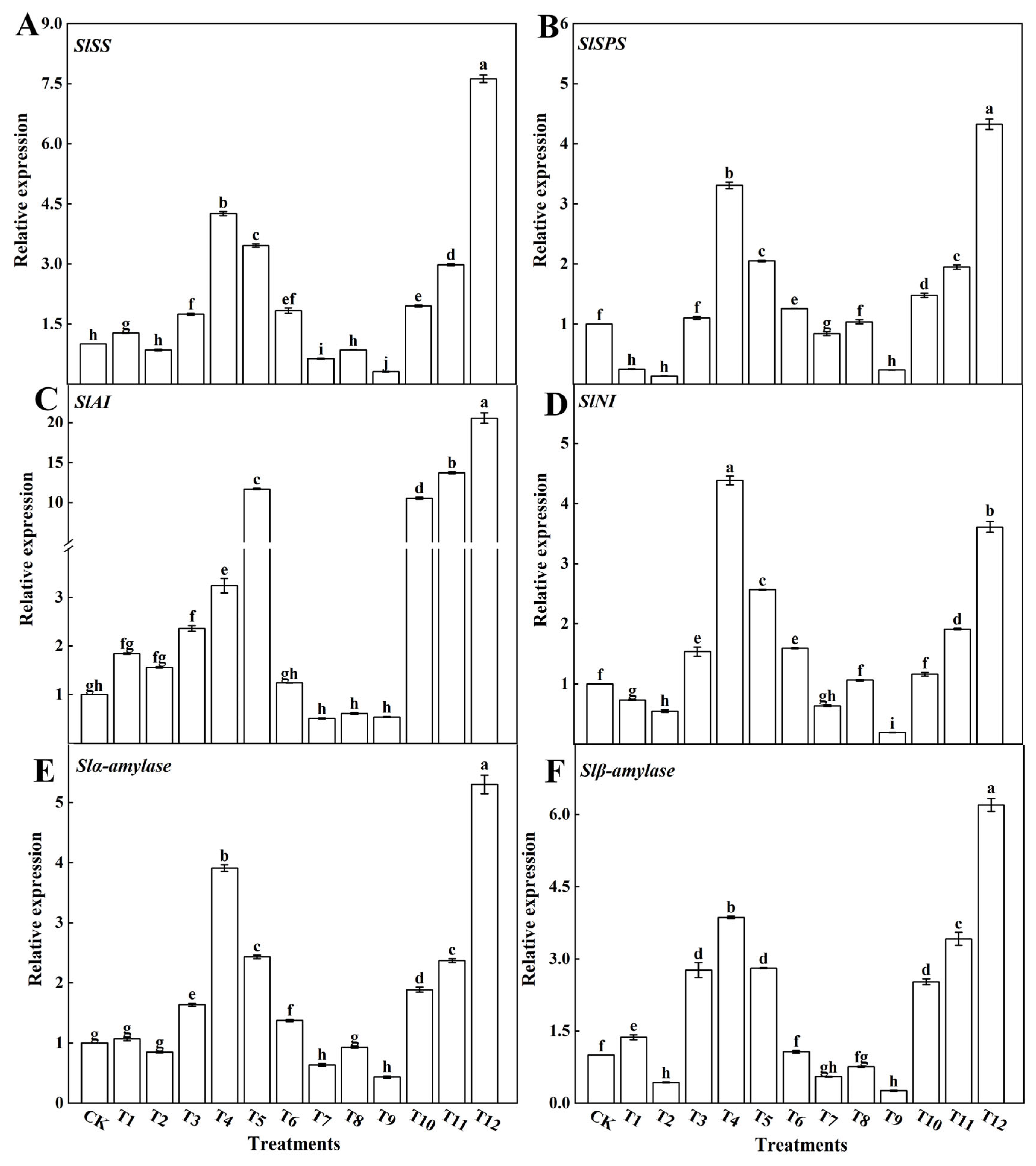
3.3.2. Sugar-Metabolism-Related Enzymes and Gene Expression
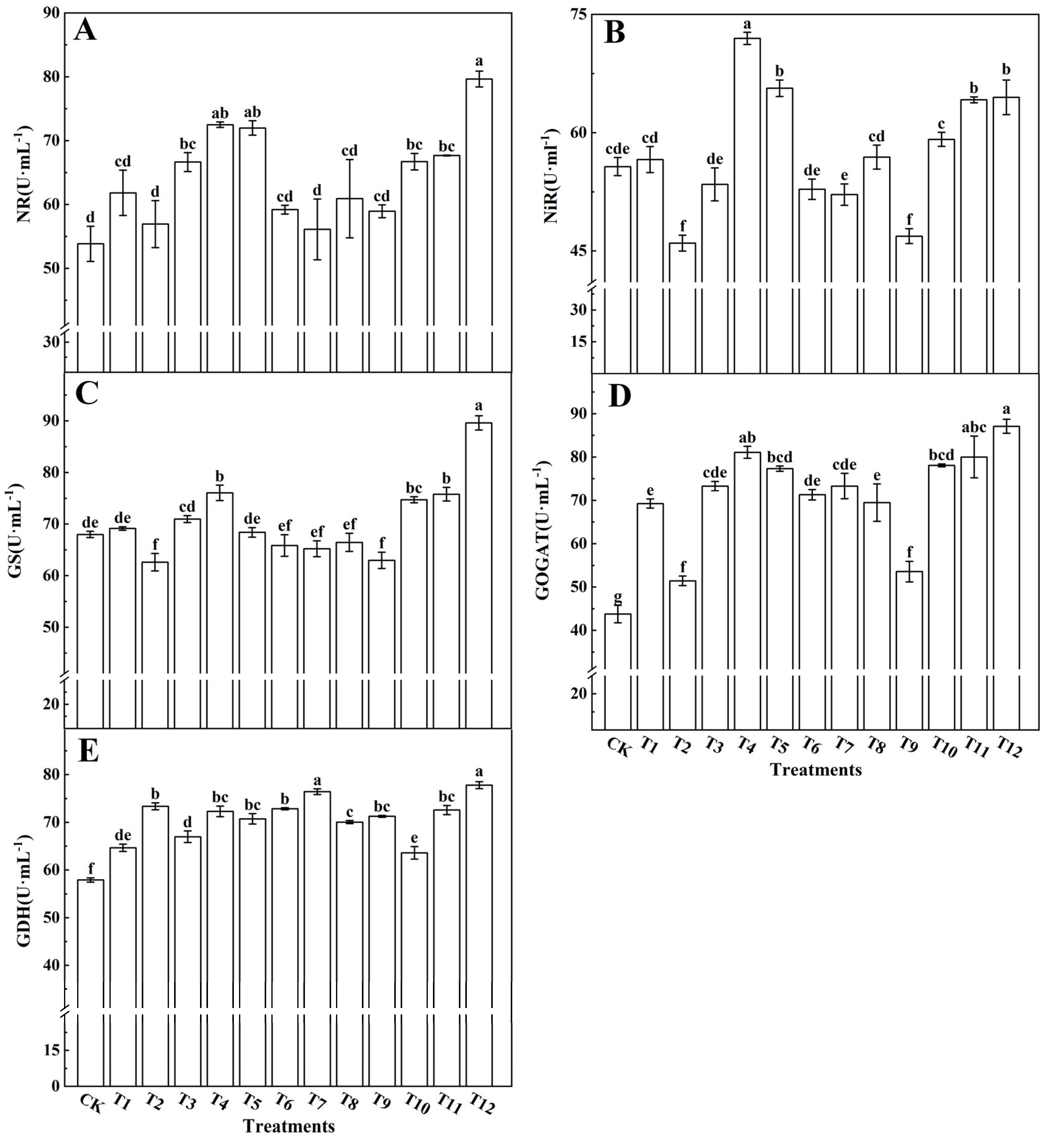
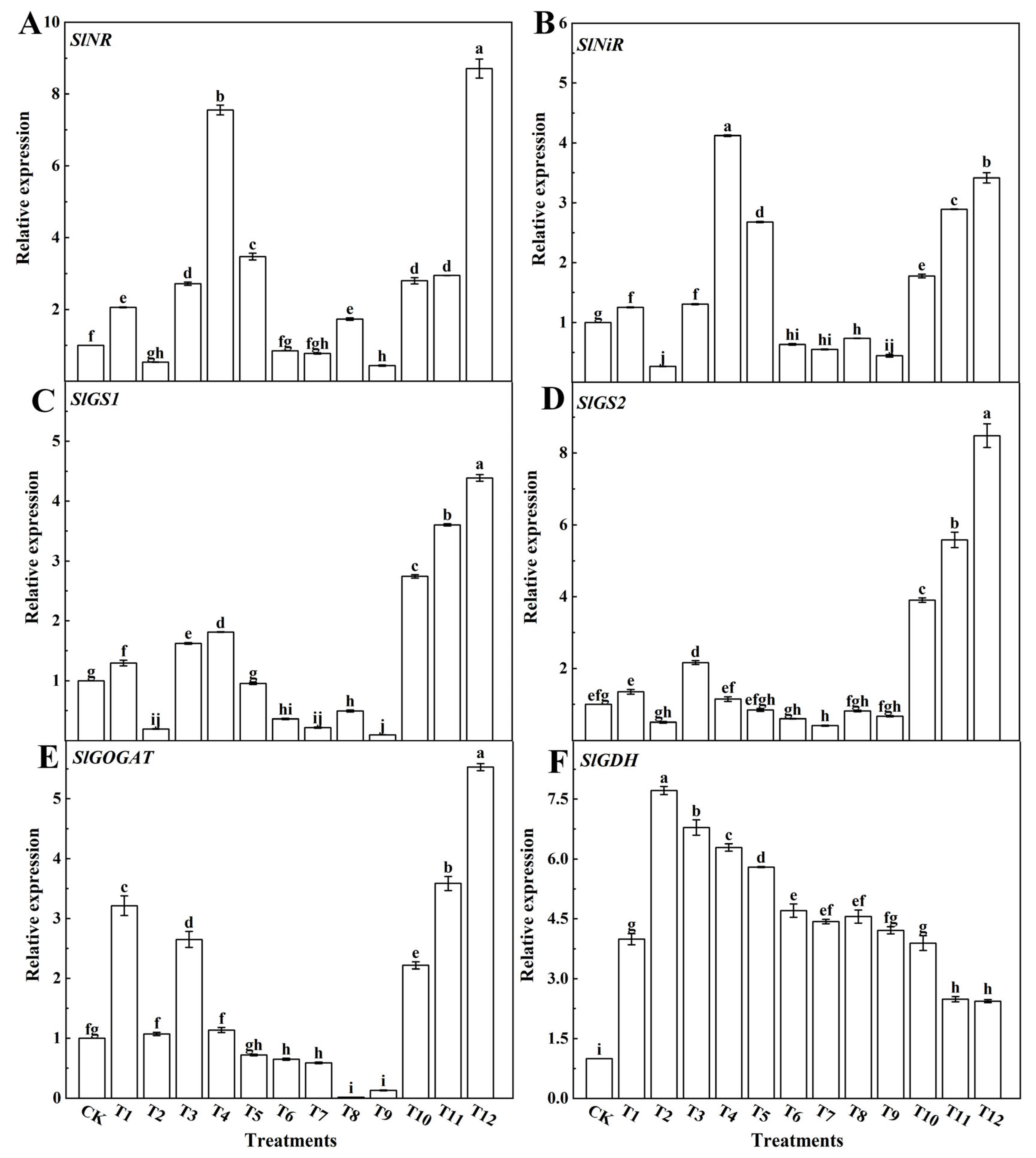
3.4. Effects of Different Forms of Nitrogen on the Metabolism of Nitrogen in Tomato Leaves
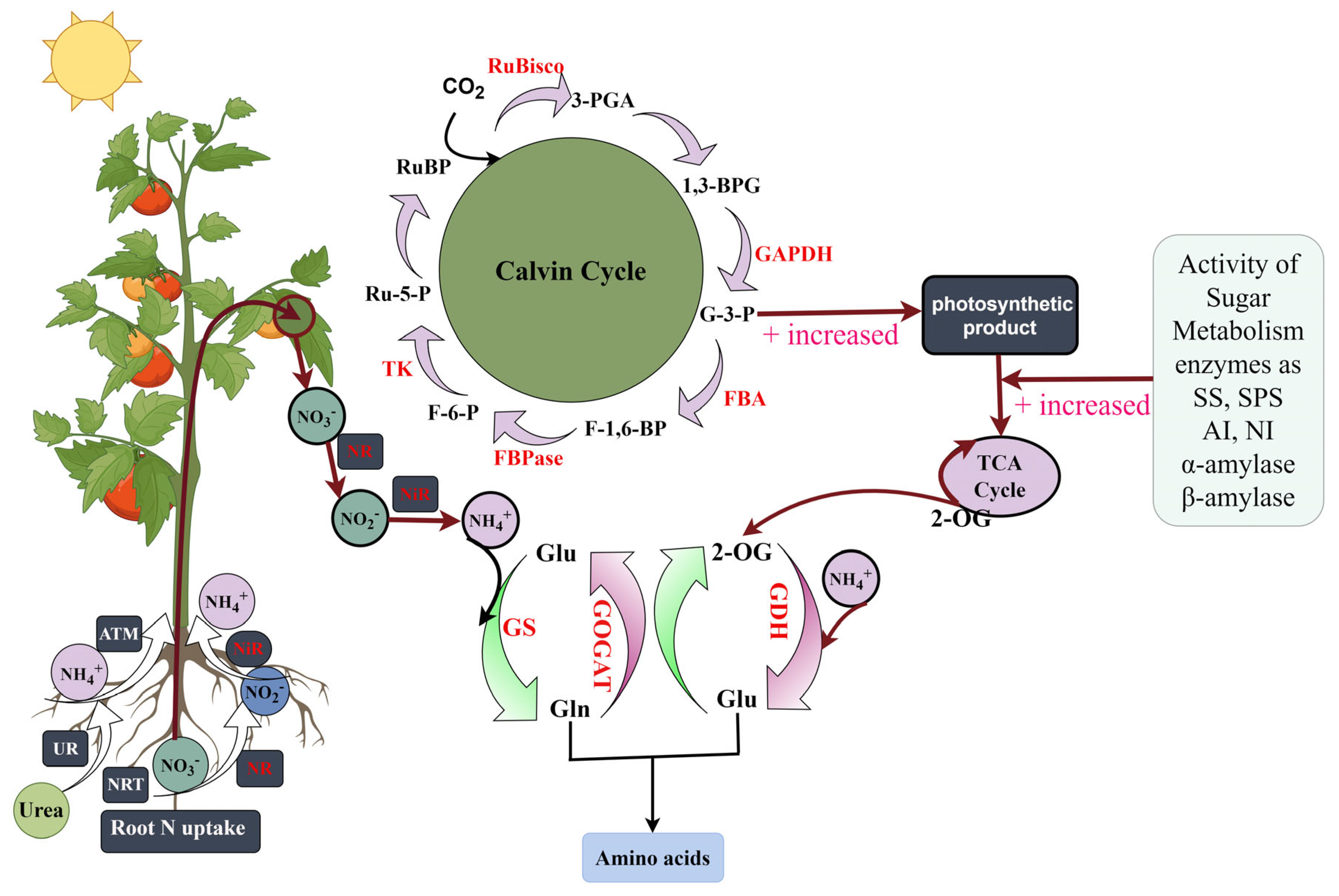
4. Discussion
4.1. Effects of Different Forms of Nitrogen on Tomato Growth and Root Morphological Parameters
4.2. Different Forms of Nitrogen Affected the Chlorophyll Fluorescence Parameters and Photosynthesis in the Tomato Leaves
4.3. Different Forms of Nitrogen Affect the Activity of Key Enzymes in the Calvin Cycle and the Relative Expression of Genes in Tomato Leaves
4.4. Effects of Different Forms of Nitrogen on Photosynthetic Products in Tomato Leaves
4.5. Effects of Different Forms of Nitrogen on the Metabolism of Nitrogen in Tomato Leaves
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glass, A. Homeostatic processes for the optimization of nutrient absorption: Physiology and molecular biology. In Nutrient Acquisition by Plants; Springer: Berlin, Germany, 2005; pp. 117–140. [Google Scholar]
- Piwpuan, N.; Zhai, X.; Brix, H. Nitrogen nutrition of Cyperus laevigatus and Phormium tenax: Effects of ammonium versus nitrate on growth, nitrate reductase activity and N uptake kinetics. Aquat. Bot. 2013, 106, 42–51. [Google Scholar] [CrossRef]
- Alt, D.S.; Doyle, J.W.; Malladi, A. Nitrogen-source preference in blueberry (Vaccinium sp.): Enhanced shoot nitrogen assimilation in response to direct supply of nitrate. J. Plant Physiol. 2017, 216, 79–87. [Google Scholar] [CrossRef]
- M’rah Helali, S.; Nebli, H.; Kaddour, R.; Mahmoudi, H.; Lachaâl, M.; Ouerghi, Z. Influence of Nitrate-Ammonium ratio on growth and nutrition of Arabidopsis thaliana. Plant Soil 2010, 336, 65–74. [Google Scholar] [CrossRef]
- Mattson, N.; Leatherwood, R.; Peters, C. Nitrogen: All Forms Are Not Equal; Greenhouse Management and Production: New York, NY, USA, 2009; Volume 29, pp. 18–20, 22–23. [Google Scholar]
- Guo, L.; Meng, H.; Teng, K.; Fan, X.; Zhang, H.; Teng, W.; Yue, Y.; Wu, J. Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings. Plants 2022, 11, 2086. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Pang, Y.; Yuan, W.; Peng, J.; Zhao, Z. Growth and nitrogen metabolism in Sophora japonica (L.) as affected by salinity under different nitrogen forms. Plant Sci. 2022, 322, 111347. [Google Scholar] [CrossRef] [PubMed]
- Zanin, L.; Tomasi, N.; Wirdnam, C.; Meier, S.; Komarova, N.Y.; Mimmo, T.; Cesco, S.; Rentsch, D.; Pinton, R. Isolation and functional characterization of a high affinity urea transporter from roots of Zea mays. BMC Plant Biol. 2014, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, S.; Wang, Z.; Yin, X.; Chen, S. Response of lettuce to different nitrogen forms. J. Appl. Ecol. 2003, 14, 377–381. [Google Scholar]
- Cui, H.; Luo, Y.; Li, C. Effects of nitrogen forms on nitrogen utilization, yield, and quality of two wheat varieties with different gluten characteristics. Eur. J. Agron. 2023, 149, 126919. [Google Scholar] [CrossRef]
- Magalhaes, J.R.; Huber, D.M. Response of ammonium assimilation enzymes to nitrogen form treatments in different plant species. J. Plant Nutr. 1991, 14, 175–185. [Google Scholar] [CrossRef]
- Guo, S.; Chen, G.; Zhou, Y.; Shen, Q. Ammonium nutrition increases photosynthesis rate under water stress at early development stage of rice (Oryza sativa L.). Plant Soil 2007, 296, 115–124. [Google Scholar] [CrossRef]
- Fu, Y.; Zhong, X.; Lu, C.; Liang, K.; Pan, J.; Hu, X.; Hu, R.; Li, M.; Ye, Q.; Liu, Y. Growth, nutrient uptake and transcriptome profiling of rice seedlings in response to mixed provision of ammonium-and nitrate-nitrogen. J. Plant Physiol. 2023, 284, 153976. [Google Scholar] [CrossRef]
- Raab, T.K.; Terry, N. Nitrogen source regulation of growth and photosynthesis in Beta vulgaris L. Plant Physiol. 1994, 105, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, J.; Gan, Y.; Jeffrey, A.C.; Mohammed, M.D.; Yu, J.; Lv, J.; Li, J.; Zhang, X.; Tang, C.; et al. Corrigendum to: Promoting pepper (Capsicum annuum) photosynthesis via chloroplast ultrastructure and enzyme activities by optimising the ammonium to nitrate ratio. Funct. Plant Biol. 2020, 47, 473. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.X.; Liu, W.Q.; Shi, Y.C. Effects of different nitrogen forms on photosynthetic rate and the chlorophyll fluorescence induction kinetics of flue-cured tobacco. Photosynthetica 2006, 44, 140–142. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, S.; Liang, Y.; Li, B.; Ma, S.; Wang, Z.; Ma, B.; Li, M. Nitrogen levels regulate sugar metabolism and transport in the shoot tips of crabapple plants. Front. Plant Sci. 2021, 12, 626149. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lv, Y.; Lian, M.; Peng, F.; Xiao, Y. Effects of combined glycine and urea fertilizer application on the photosynthesis, sucrose metabolism, and fruit development of peach. Sci. Hortic. 2021, 289, 110504. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, L. Photosynthetic enzymes and carbohydrate metabolism of apple leaves in response to nitrogen limitation. J. Hortic. Sci. Biotechnol. 2004, 79, 923–929. [Google Scholar] [CrossRef]
- Paponov, I.A.; Engels, C. Effect of nitrogen supply on carbon and nitrogen partitioning after flowering in maize. J. Plant Nutr. Soil Sci. 2005, 168, 447–453. [Google Scholar] [CrossRef]
- Brajeul, E.; Vercambre, G.; Gautier, H. To Stop Nitrogen Overdose in Soilless Tomato Crop: A Way to Promote Fruit Quality without Affecting Fruit Yield. Agronomy 2019, 9, 80. [Google Scholar]
- Xue, Y.X.; Li, D.X.; Li, Y.L. Study on method to measure tomato leaf area. J. Northwest A F Univ. 2006, 34, 116–120. (In Chinese) [Google Scholar]
- Li, H. Principle and Technology of Plant Physiological and Biochemical Experiment. In Principle and Technology of Plant Physiological and Biochemical Experiment; Higher Education Research and Development: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Wang, S.; Meng, X.; Tang, Z.; Wu, Y.; Xiao, X.; Zhang, G.; Hu, L.; Liu, Z.; Lyu, J.; Yu, J. Red and Blue LED Light Supplementation in the Morning Pre-activates the Photosynthetic System of Tomato (Solanum lycopersicum L.) Leaves and Promotes Plant Growth. Agronomy 2022, 12, 897. [Google Scholar] [CrossRef]
- Aljarba, N.H.; Ali, H.; Alkahtani, S. Synergistic dose permutation of isolated alkaloid and sterol for anticancer effect on young Swiss albino mice. Drug Des. Dev. Ther. 2021, 15, 4043–4052. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Dixit, S.; Ali, D.; Alqahtani, S.M.; Alkahtani, S.; Alarifi, S. Isolation and evaluation of anticancer efficacy of stigmasterol in a mouse model of DMBA-induced skin carcinoma. Drug Des. Dev. Ther. 2015, 9, 2793–2800. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Jin, L.; Wang, S.; Wang, S.; Meng, X.; Ma, X.; He, X.; Zhang, G.; Luo, S.; Lyu, J.; et al. A comprehensive evaluation of effects on water-level deficits on tomato polyphenol composition, nutritional quality and antioxidant capacity. Antioxidants 2022, 11, 1585. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chen, W.; Chen, K.; Zhao, M.; Jin, G.; Wu, W. A Simple Method for Determining the Content of Starch-Iodine Colorimety. Biotechnology 1998, 8, 3. (In Chinese) [Google Scholar]
- Hu, L.; Gao, X.; Li, Y.; Lyu, J.; Xiao, X.; Zhang, G.; Yu, J. Nitric Oxide Induced by Ammonium/Nitrate Ratio Ameliorates Low-Light Stress in Brassica pekinesis: Regulation of Photosynthesis and Root Architecture. Int. J. Mol. Sci. 2023, 24, 7271. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Yang, H.; Yang, H.; Wei, Z.; Che, J.; Wu, W.; Lyu, L.; Li, W. Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms. Plants 2023, 12, 1480. [Google Scholar] [CrossRef]
- Boschiero, B.N.; Mariano, E.; Azevedo, R.A. Influence of nitrate-ammonium ratio on the growth, nutrition, and metabolism of sugarcane. Plant Physiol. Biochem. 2019, 139, 246–255. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Scalschi, L.; García-Agustín, P.; Camañes, G. Tomato root development and N assimilation depend on C and ABA content under different N sources. Plant Physiol. Biochem. 2020, 148, 368–378. [Google Scholar] [CrossRef]
- Criddle, R.S.; Ward, M.R.; Huffaker, R.C. Nitrogen uptake by wheat seedlings, interactive effects of four nitrogen sources: NO3−, NO2−, NH4+, and urea. Plant Physiol. 1988, 86, 166–175. [Google Scholar] [CrossRef]
- Breteler, H.; Siegerist, M. Effect of ammonium on nitrate utilization by roots of dwarf bean. Plant Physiol. 1984, 75, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Kronzucker, N.; Siddiqi, N.; Glass, N. Nitrate-ammonium synergism in rice. A subcellular flux analysis. Plant Physiol. 1999, 119, 1041–1046. [Google Scholar] [PubMed]
- Rufty, T.W., Jr.; Raper, C.D., Jr.; Jackson, W.A. Growth and nitrogen assimilation of soybeans in response to ammonium and nitrate nutrition. Bot. Gaz. 1983, 144, 466–470. [Google Scholar] [CrossRef]
- Yan, F.; Schubert, S.; Mengel, K. Effect of Low Root Medium pH on Net Proton Release, Root Respiration, and Root Growth of Corn (Zea mays L.) and Broad Bean (Vicia faba L.). Plant Physiol. 1992, 99, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Forde, B.G. Nitrate signalling mediated by the NRT1.1 nitrate transporter antagonises L-glutamate-induced changes in root architecture. Plant J. 2008, 54, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, E.A.; Mengel, K. Ionic balance in different tissues of the tomato plant in relation to nitrate, urea, or ammonium nutrition. Plant Physiol. 1967, 42, 6–14. [Google Scholar] [CrossRef]
- Garnica, M.; Houdusse, F.; Zamarreño, A.M. Nitrate modifies the assimilation pattern of ammonium and urea in wheat seedlings. J. Sci. Food Agric. 2009, 90, 357–369. [Google Scholar] [CrossRef]
- Garnica, M.; Houdusse, F.; Yvin, J.C. Nitrate modifies urea root uptake and assimilation in wheat seedlings. J. Sci. Food Agric. 2009, 89, 55–62. [Google Scholar] [CrossRef]
- Bradley, D.P.; Morgan, M.A.; O’Toole, P. Uptake and apparent utilization of urea and ammonium nitrate in wheat seedlings. Fertil. Res. 1989, 20, 41–49. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants—Growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef]
- Zou, N.; Qiang, X.; Shi, W. Effects of different ammonium levels on root growth of tomato. Soil 2012, 44, 827–833. (In Chinese) [Google Scholar]
- Golvano, M.P.; Felipe, M.R.; Cintas, A.M. Influence of nitrogen sources on chloroplast development in wheat seedlings. Physiol. Plant. 1982, 56, 353–360. [Google Scholar] [CrossRef]
- Claussen, W.; Lenz, F. Effect of ammonium or nitrate nutrition on net photosynthesis, growth, and activity of the enzymes nitrate reductase and glutamine synthetase in blueberry, raspberry and strawberry. Plant Soil 1999, 208, 95–102. [Google Scholar] [CrossRef]
- Puritch, G.S.; Barker, A.V. Structure and function of tomato leaf chloroplasts during ammonium toxicity. Plant Physiol. 1967, 42, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Frantz, T.A.; Peterson, D.M.; Durbin, R.D. Sources of ammonium in oat leaves treated with tabtoxin or methionine sulfoximine. Plant Physiol. 1982, 69, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Givan, C.V. Metabolic detoxification of ammonia in tissues of higher plants. Phytochemistry 1979, 18, 375–382. [Google Scholar] [CrossRef]
- Yin, H.; Li, B.; Wang, X.; Yin, H.; Li, B.; Wang, X.; Xi, Z. Effect of Ammonium and Nitrate Supplies on Nitrogen and Sucrose Metabolism of Cabernet Sauvignon (Vitis vinifera cv.). J. Sci. Food Agric. 2020, 100, 5239–5250. [Google Scholar] [CrossRef]
- Tabatabaei, S.J.; Yusefi, M.; Hajiloo, J. Effects of shading and NO3−: NH4+ ratio on the yield, quality and N metabolism in strawberry. Sci. Hortic. 2008, 116, 264–272. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. A chlorophyll fluorescence analysis of photosynthetic efficiency, quantum yield and photon energy dissipation in PSII antennae of Lactuca sativa L. leaves exposed to cinnamic acid. Plant Physiol. Biochem. 2011, 49, 1290–1298. [Google Scholar] [CrossRef]
- Rimando, A.M.; Dayan, F.E.; Czarnota, M.A.; Weston, L.A.; Duke, S.O. A new photosystem II electron transfer inhibitor from Sorghum bicolor. J. Nat. Prod. 1998, 61, 927–930. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Csintalan, Z. Chlorophyll Fluorescence during Drying and Rehydration in the Mosses Rhytidiadelphus loreus (Hedw.) Warnst., Anomodon viticulosus (Hedw.) Hook. & Tayl. and Grimmia pulvinata (Hedw.) Sm. Ann. Bot. 1999, 84, 235–244. [Google Scholar]
- Ehlert, B.; Hincha, D.K. Chlorophyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in Arabidopsis leaves. Plant Methods 2008, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence as a tool in plant physiology: II. Interpretation of fluorescence signals. Photosynth. Res. 1984, 5, 139–157. [Google Scholar] [CrossRef]
- Li, Y.; Shi, S.; Zhang, Y.; Zhang, A.; Wang, Z.; Yang, Y. Copper stress-induced phytotoxicity associated with photosynthetic characteristics and lignin metabolism in wheat seedlings. Ecotoxicol. Environ. Saf. 2023, 254, 114739. [Google Scholar] [CrossRef]
- Hu, L.; Yu, J.; Liao, W.; Zhang, G.; Xie, J.; Lyu, J.; Xiao, X.; Yang, B.; Zhou, R.; Bu, R. Moderate ammonium: Nitrate alleviates low light intensity stress in mini Chinese cabbage seedling by regulating root architecture and photosynthesis. Sci. Hortic. 2015, 186, 143–153. [Google Scholar] [CrossRef]
- Rehab, A.E.; Reem, H.B.; Marwa, H.Z. Effect of chilling and salinity stress on photosynthetic performance and ultrastructure of chloroplast in faba beans (Vicia faba L.) leaves. Rend. Lincei. Sci. Fis. E Nat. 2023, 34, 447–456. [Google Scholar]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Krause, A.G.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Badger, M.R.; von Caemmerer, S.; Ruuska, S.; Nakano, H. Electron flow to oxygen in higher plants and algae: Rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 1433–1446. [Google Scholar] [CrossRef] [PubMed]
- Osório, J.; Osório, M.L.; Correia, P.J.; Varennes, A.; Pestana, M. Chlorophyll fluorescence imaging as a tool to understand the impact of iron deficiency and resupply on photosynthetic performance of strawberry plants. Sci. Hortic. 2014, 165, 148–155. [Google Scholar] [CrossRef]
- Nasraoui-Hajaji, A.; Gouia, H. Photosynthesis sensitivity to NH4+-N change with nitrogen fertilizer type. Plant Soil Environ. 2014, 60, 274–279. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.V.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Raab, T.K.; Terry, N. Carbon, nitrogen, and nutrient interactions in Beta vulgaris L. as influenced by nitrogen source, NO3− versus NH4+. Plant Physiol. 1995, 107, 575–585. [Google Scholar] [CrossRef]
- Kasukabe, Y.; He, L.; Nada, K.; Misawa, S.; Ihara, L.; Tachibana, S. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.X.; Cao, B.H.; Feng, X.P.; He, Y.; Jiang, D. Photosynthetic response of salt-tolerant and sensitive soybean varieties. Photosynthetica 2009, 47, 381–387. [Google Scholar] [CrossRef]
- Spreitzer, R.J. Role of the small subunit in ribulose-1, 5-bisphosphate carboxylase/oxygenase. Arch. Biochem. Biophys. 2003, 414, 141–149. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Tamoi, M.; Shigeoka, S. Overexpression of a cyanobacterial fructose-1, 6-/sedoheptulose-1, 7-bisphosphatase in tobacco enhances photosynthesis and growth. Nat. Biotechnol. 2001, 19, 965–969. [Google Scholar] [CrossRef]
- Xia, X.; Huang, L.; Zhou, Y.; Mao, W.; Shi, K.; Wu, J.; Asami, T.; Chen, Z.; Yu, J. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185–1196. [Google Scholar] [CrossRef]
- Chen, G.; Wang, L.; Fabrice, M.R.; Tian, Y.; Qi, K.; Chen, Q.; Cao, P.; Wang, P.; Zhang, S.; Wu, J.; et al. Physiological and Nutritional Responses of Pear Seedlings to Nitrate Concentrations. Front. Plant Sci. 2018, 9, 1679. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.W.; Jiang, M.M.; Lu, D.K. Nitrogen forms affect the root characteristic, photosynthesis, grain yield, and nitrogen use efficiency of rice under different irrigation regimes. Crop Sci. 2020, 60, 2594–2610. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, K.; Cheng, M. Effects of nitrogen form on growth and quality of Chrysanthemums morifolium. China J. Chin. Mater. Med. 2014, 39, 3263–3268. [Google Scholar]
- Moreau, D.; Milard, G.; Munier-Jolain, N. A plant nitrophily index based on plant leaf area response to soil nitrogen availability. Agron. Sustain. Dev. 2013, 33, 809–815. [Google Scholar] [CrossRef]
- Sagi, M.; Dovrat, A.; Kipnis, T.; Lips, H. Nitrate reductase, phosphoenolpyruvate carboxylase, and glutamine synthetase in annual ryegrass as affected by salinity and nitrogen. J. Plant Nutr. 1998, 21, 707–723. [Google Scholar] [CrossRef]
- Jampeetong, A.; Brix, H. Nitrogen nutrition of Salvinia natans: Effects of inorganic nitrogen form on growth, morphology, nitrate reductase activity and uptake kinetics of ammonium and nitrate. Aquat. Bot. 2009, 90, 67–73. [Google Scholar] [CrossRef]
- Konnerup, D.; Brix, H. Nitrogen nutrition of Canna indica: Effects of ammonium versus nitrate on growth, biomass allocation, photosynthesis, nitrate reductase activity and N uptake rates. Aquat. Bot. 2010, 92, 142–148. [Google Scholar] [CrossRef]
- Glass, A.D.; Britto, D.T.; Kaiser, B.N.; Kinghorn, J.R.; Kronzucker, J.H.; Kumar, A.; Okamoto, M.; Rawat, S.; Siddiqi, M.Y.; Unkles, S.E.; et al. The regulation of nitrate and ammonium transport systems in plants. J. Exp. Bot. 2002, 53, 855–864. [Google Scholar] [CrossRef]
- Mihaljević, S.; Radić, S.; Bauer, N.; Garić, R.; Mihaljević, B.; Horvat, G.; Leljak-Levanić, D.; Jelaska, S. Ammonium-related metabolic changes affect somatic embryogenesis in pumpkin (Cucurbita pepo L.). J. Plant Physiol. 2011, 168, 1943–1951. [Google Scholar] [CrossRef]
- Ma, C.; Ban, T.; Yu, H.; Li, X.; Jiang, W.; Xie, J. Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber. Agronomy 2019, 9, 262. [Google Scholar] [CrossRef]
- Choi, S.; Park, D.; Hong, K. Status of nitrogenous and carbohydrate compounds as affected by nitrogen fertigation rates in young persimmon trees. Sci. Hortic. 2011, 130, 354–356. [Google Scholar] [CrossRef]
- Alexander, K.G.; Miller, M.H.; Beauchamp, E.G. The effect of an NH4+-enhanced nitrogen source on the growth and yield of hydroponically grown maize (Zea mays L.). J. Plant Nutr. 1991, 14, 31–44. [Google Scholar] [CrossRef]
- Takács, E.; Técsi, L. Effects of NO3−/NH4+ ratio on photosynthetic rate, nitrate reductase activity and chloroplast ultrastructure in three cultivars of red pepper (Capsicum annuum L.). J. Plant Physiol. 1992, 140, 298–305. [Google Scholar] [CrossRef]
- Shi, J.L.; Jiang, Y.L.; Song, Y.H. Effects of different ratios of nitrogen forms on enzyme activities related to nitrogen metabolism and growth of cucumber seedlings. Chin. Agric. Sci. Bull. 2009, 25, 225–227. [Google Scholar]
- Zhang, S.Y.; Chu, G.X.; Liang, Y.C. Effects of enhancing ammonium nutrition on the nitrogenous metabolisms of cotton seedlings grown hydroponically under low-temperature stress. J. Plant Nutr. Fertil. 2017, 23, 983–990. [Google Scholar]
- Horchani, F.; Hajri, R.; Aschi-Smiti, S. Effect of ammonium or nitrate nutrition on photosynthesis, growth, and nitrogen assimilation in tomato plants. J. Plant Nutr. Soil Sci. 2010, 173, 610–617. [Google Scholar] [CrossRef]
- Bubner, B.; Baldwin, I.T. Use of real-time PCR for determining copy number and zygosity in transgenic plants. Plant Cell Rep. 2004, 23, 263–271. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Nitrogen Form and Ratio | Elements Concentration (mmol·L−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| NO3−-N | NH4+-N | (CO(NH2)2) | P | K | Ca | Mg | ||
| CK | - | 0 | 0 | 0 | 1 | 6 | 5 | 2 |
| T1 | NO3−-N (NN) | 15 | 0 | 0 | 1 | 6 | 5 | 2 |
| T2 | NH4+-N (AN) | 0 | 15 | 0 | 1 | 6 | 5 | 2 |
| T3 | Urea (UN) | 0 | 0 | 15 | 1 | 6 | 5 | 2 |
| T4 | 75%NN:25%AN | 11.25 | 3.75 | 0 | 1 | 6 | 5 | 2 |
| T5 | 50%NN:50%AN | 7.5 | 7.5 | 0 | 1 | 6 | 5 | 2 |
| T6 | 25%NN:75%AN | 3.75 | 11.25 | 0 | 1 | 6 | 5 | 2 |
| T7 | 25%AN:75%UN | 0 | 3.75 | 11.25 | 1 | 6 | 5 | 2 |
| T8 | 50%AN:50%UN | 0 | 7.5 | 7.5 | 1 | 6 | 5 | 2 |
| T9 | 75%AN:25%UN | 0 | 11.25 | 3.75 | 1 | 6 | 5 | 2 |
| T10 | 75%NN:25%UN | 11.25 | 0 | 3.75 | 1 | 6 | 5 | 2 |
| T11 | 50%NN:50%UN | 7.5 | 0 | 7.5 | 1 | 6 | 5 | 2 |
| T12 | 25%NN:75%UN | 3.75 | 0 | 11.25 | 1 | 6 | 5 | 2 |
| Gene Name | Accession Number | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| SlRbcL | NC_007898.3 | CTTTCCAAGGTCCGCCTCAT | AAGTCCACCGCGAAGACATT |
| SlRbcS3 | NM_001347911.1 | GCTTCTTCAGTAATGTCCTCAGC | TCCAAGCAAGGAACCCATCC |
| SlFBPase | NM_001328673.1 | GGTCCAGATCAGCAATGCCT | CTCCCTGGCTGACAAACACT |
| SlFBA | NM_001321372.1 | GAAGAGGAAGCCACCGTCAA | GAAGAGCACGTCCGAAGGAA |
| SlTK | XM_004248512.4 | CTGTCAAGGCTGCTGAGGAA | CCCGTCAACCCCAATAGCTT |
| SlGAPDH | NM_001247874.2 | AGCCACTCAGAAGACCGTTG | AGGTCAACCACGGACACATC |
| SlAI | NM_001246913.2 | AACCCGCTATCTACCCGTCT | TCGGGCTTGATCCACTTACG |
| SlNI | XM_004249939.3 | GCGTATGGGAAGTCCTCTGG | TACGGCGGTCTATCATGCAC |
| SlSPS | NM_001246991.2 | AAAACGCCGTCAAGAACGTG | GCAATCGGCCTCTGGTACTT |
| Slα-Amylase | XM_004238109.4 | AGGCGGATGGTACAACTCTC | GCAACCGATTTGATCCCGTG |
| Slβ-Amylase | NM_001247627.2 | GAAGGAAGGTGGTGGATGGG | TTGGGCGATGGGAAGGTAAC |
| SlSS | NM_001247875.1 | GGTACGCCAAGAATCCACGACTAAG | CTTCTTCATCTCTGCCTGCTCTTCC |
| SlNR | Solyc11g013810.3.1 | GCAACTTCCCTCCTTCATCCAAC | TCGTCATCGTCATCCTCGTCTTC |
| SlNiR | Solyc10g050890.2.1 | CCGCAGAAACAGGAAGGATACAG | TGAACCATACTCATCAGCCAAACG |
| SlGS1 | Solyc11g011380.2.1 | GCGTCGTCTCACTGGAAAGC | TGCCTGCCTTCTCTGTGTCTC |
| SlGS2 | Solyc01g080280.3.1 | TACTGGACAAGCACCTGGAGAAG | AGATGTTGTTACCACCACGGAAAG |
| SlGOGAT | Solyc03g063560.3.1 | GTTATGCCGCCACTAATAGGAGAAG | ATGTCATCCAAGTCAGCAACCTTAG |
| SlGDH | Solyc10g078550.4.1 | GAAGACAGCGGTCGCCAATATAC | TCCAACTCAGAGATACTCAGGTCAC |
| Actin | Solyc11g005330 | TGTCCCTATTTACGAGGGTTATGC | CAGTTAAATCACGACCAGCAAGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Jin, L.; Li, R.; Meng, X.; Jin, N.; Wang, S.; Xu, Z.; Liu, Z.; Lyu, J.; Yu, J. Effects of Different Forms and Proportions of Nitrogen on the Growth, Photosynthetic Characteristics, and Carbon and Nitrogen Metabolism in Tomato. Plants 2023, 12, 4175. https://doi.org/10.3390/plants12244175
Sun J, Jin L, Li R, Meng X, Jin N, Wang S, Xu Z, Liu Z, Lyu J, Yu J. Effects of Different Forms and Proportions of Nitrogen on the Growth, Photosynthetic Characteristics, and Carbon and Nitrogen Metabolism in Tomato. Plants. 2023; 12(24):4175. https://doi.org/10.3390/plants12244175
Chicago/Turabian StyleSun, Jianhong, Li Jin, Ruirui Li, Xin Meng, Ning Jin, Shuya Wang, Zhiqi Xu, Zitong Liu, Jian Lyu, and Jinhua Yu. 2023. "Effects of Different Forms and Proportions of Nitrogen on the Growth, Photosynthetic Characteristics, and Carbon and Nitrogen Metabolism in Tomato" Plants 12, no. 24: 4175. https://doi.org/10.3390/plants12244175