
The Ectomycorrhizal Fungi and Soil Bacterial Communities of the Five Typical Tree Species in the Junzifeng National Nature Reserve, Southeast China


Abstract
:1. Introduction
2. Results
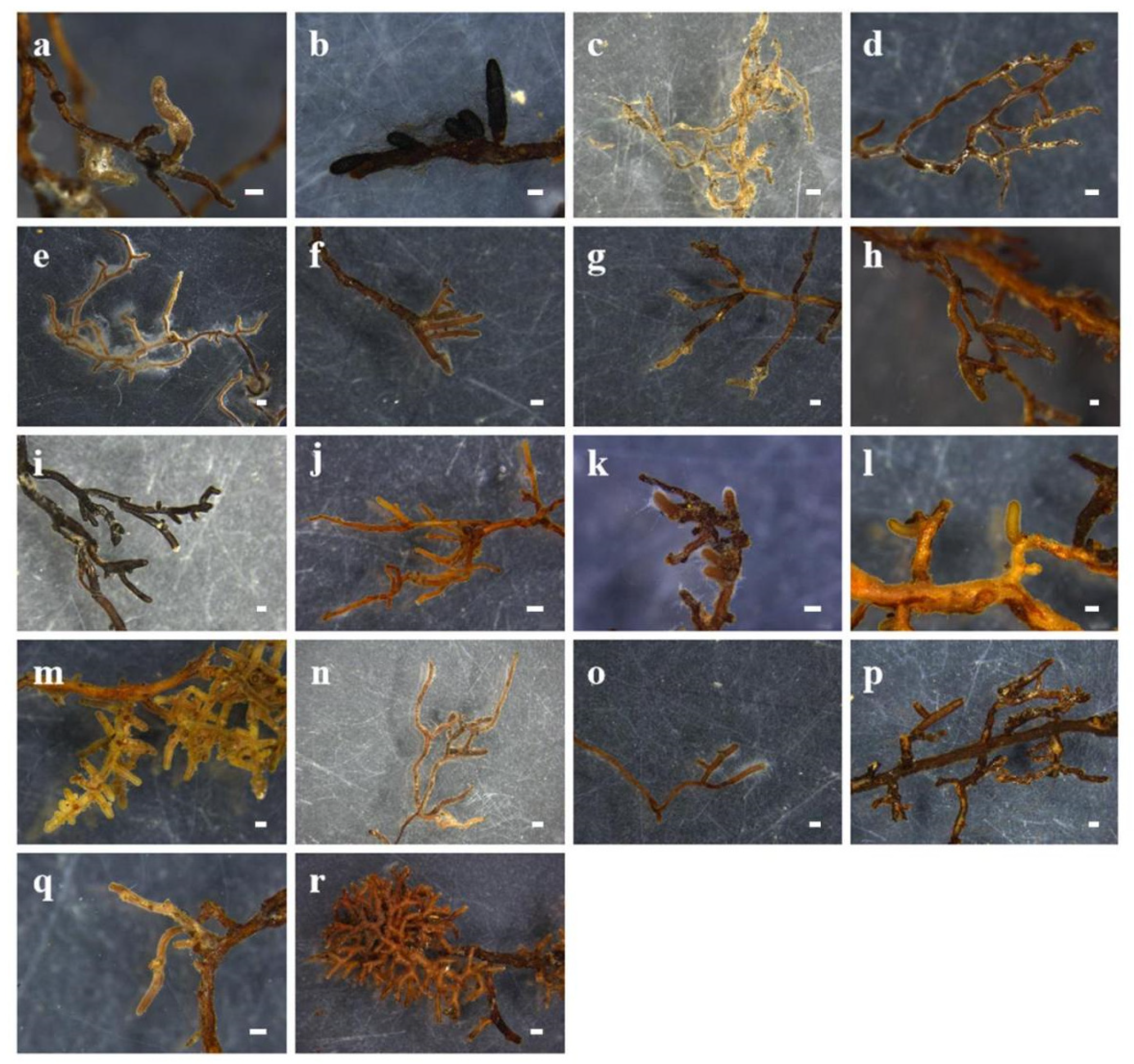
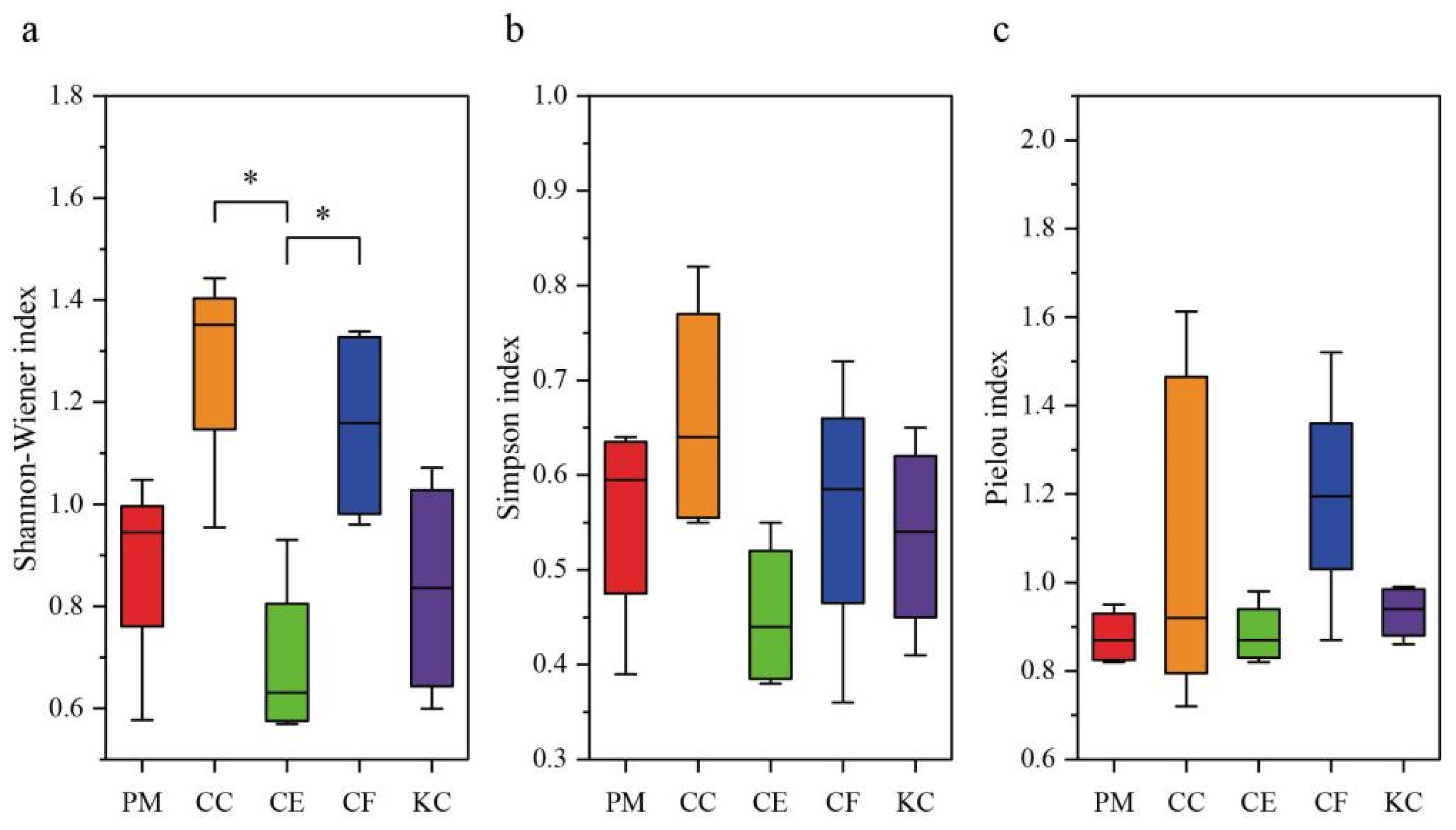
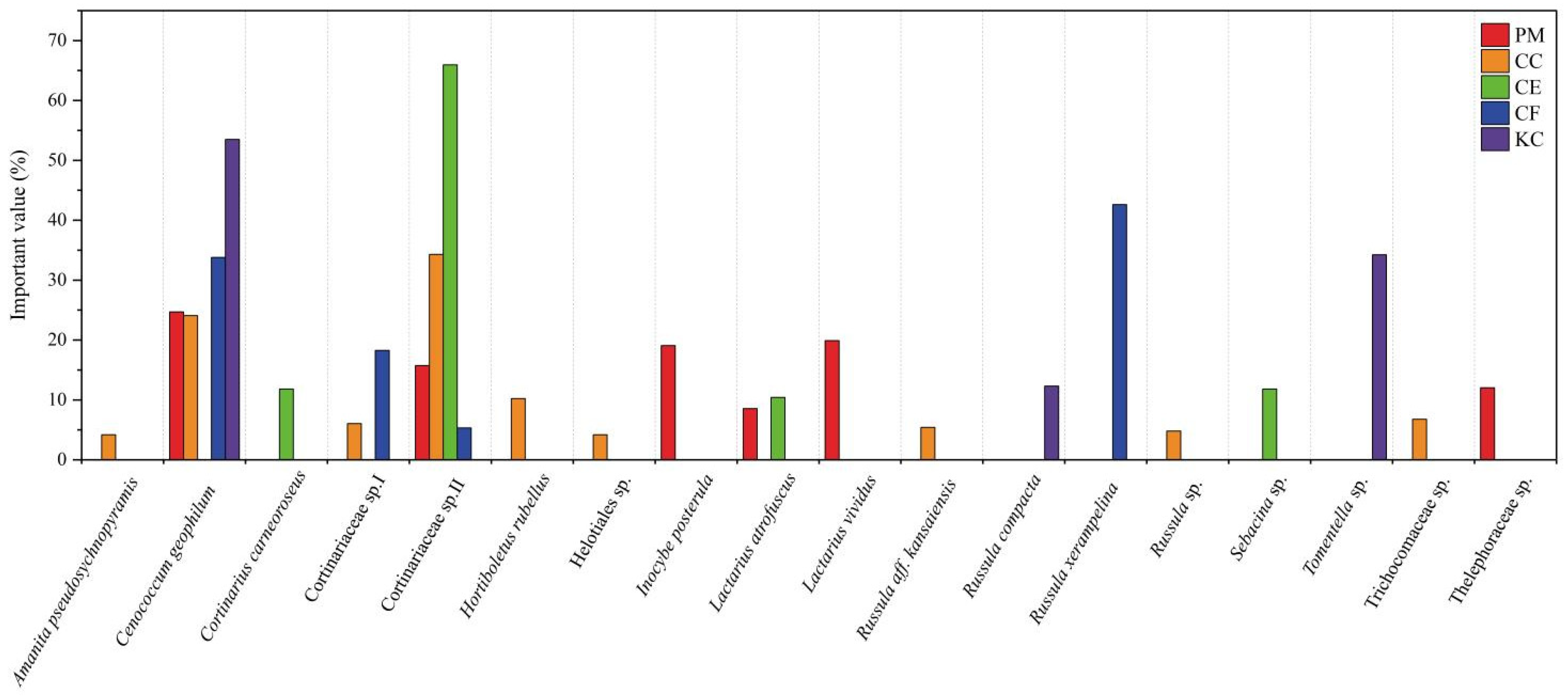
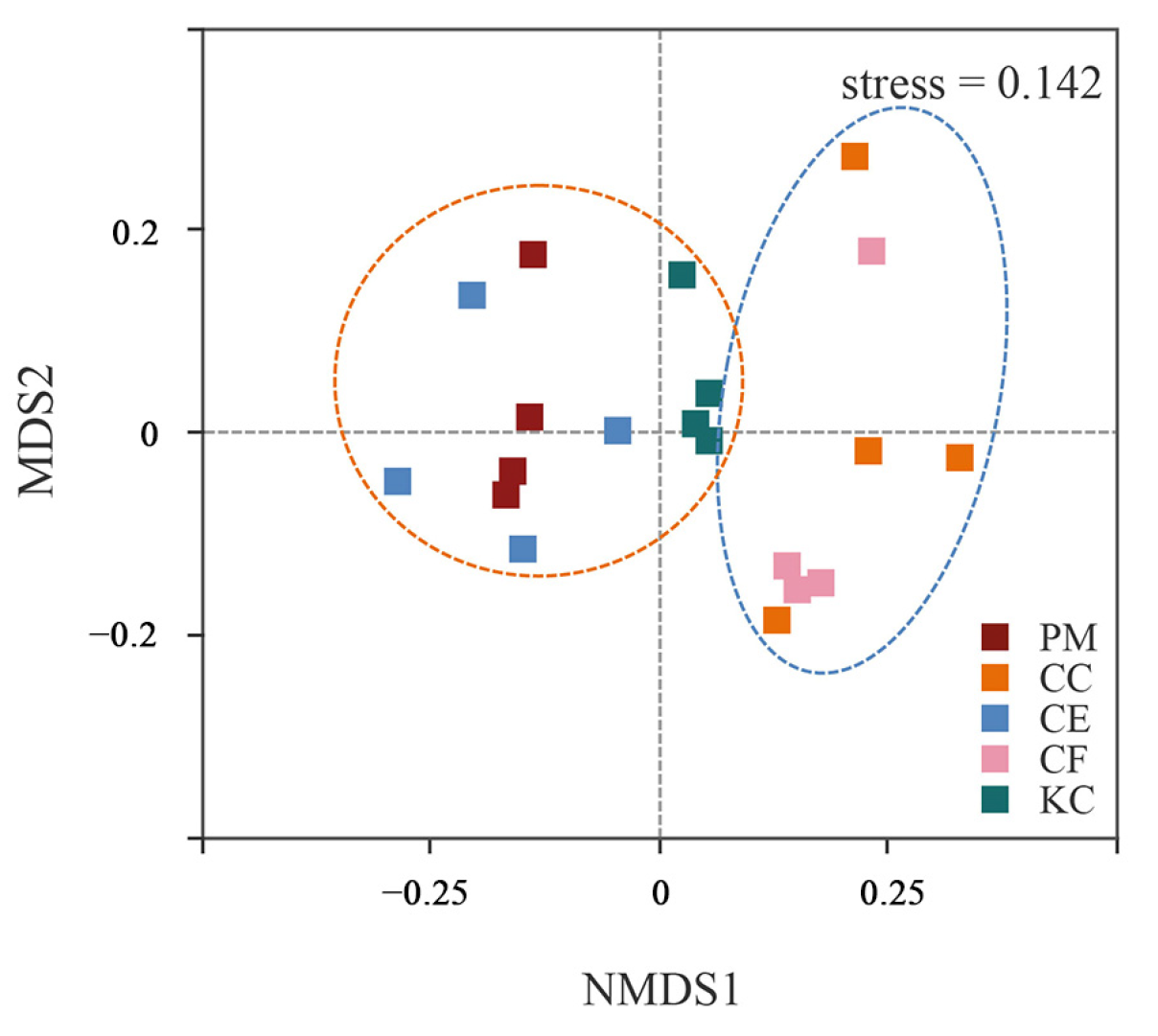
2.1. ECM Fungal Diversity and Community Composition in the Five Tree Species
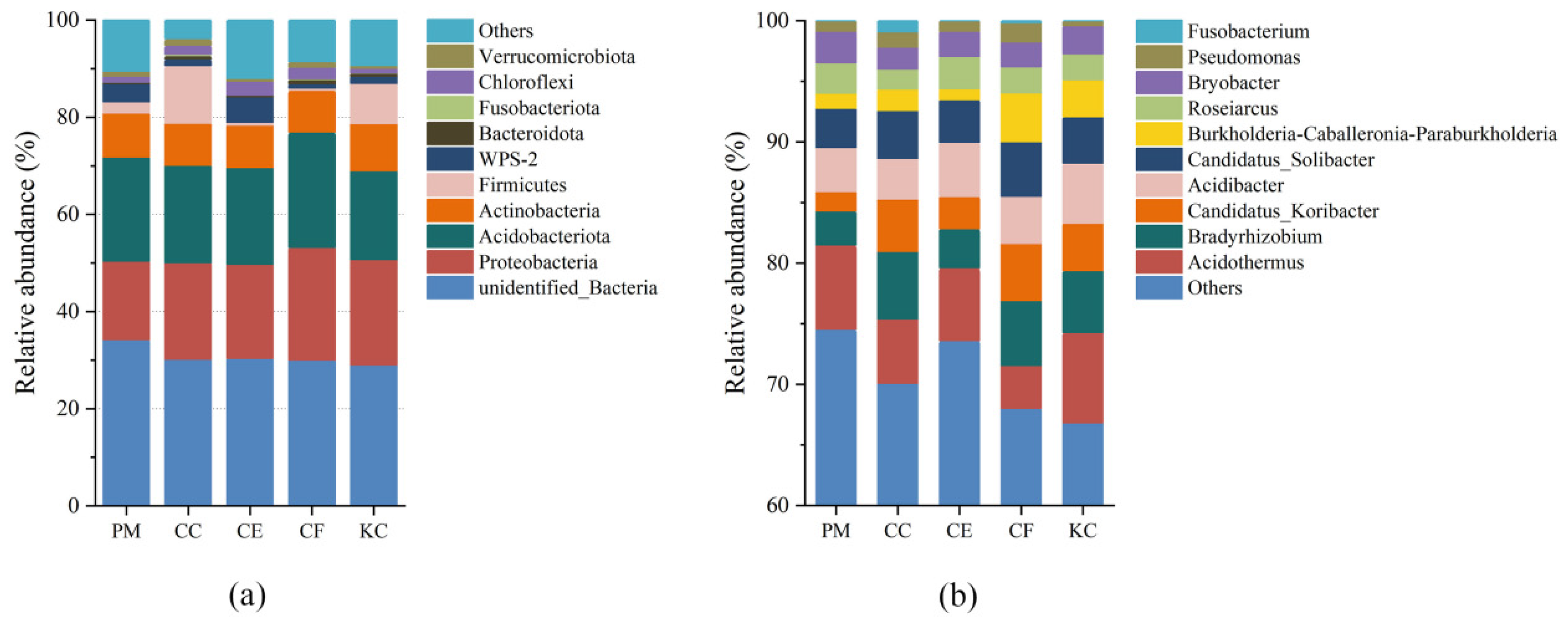
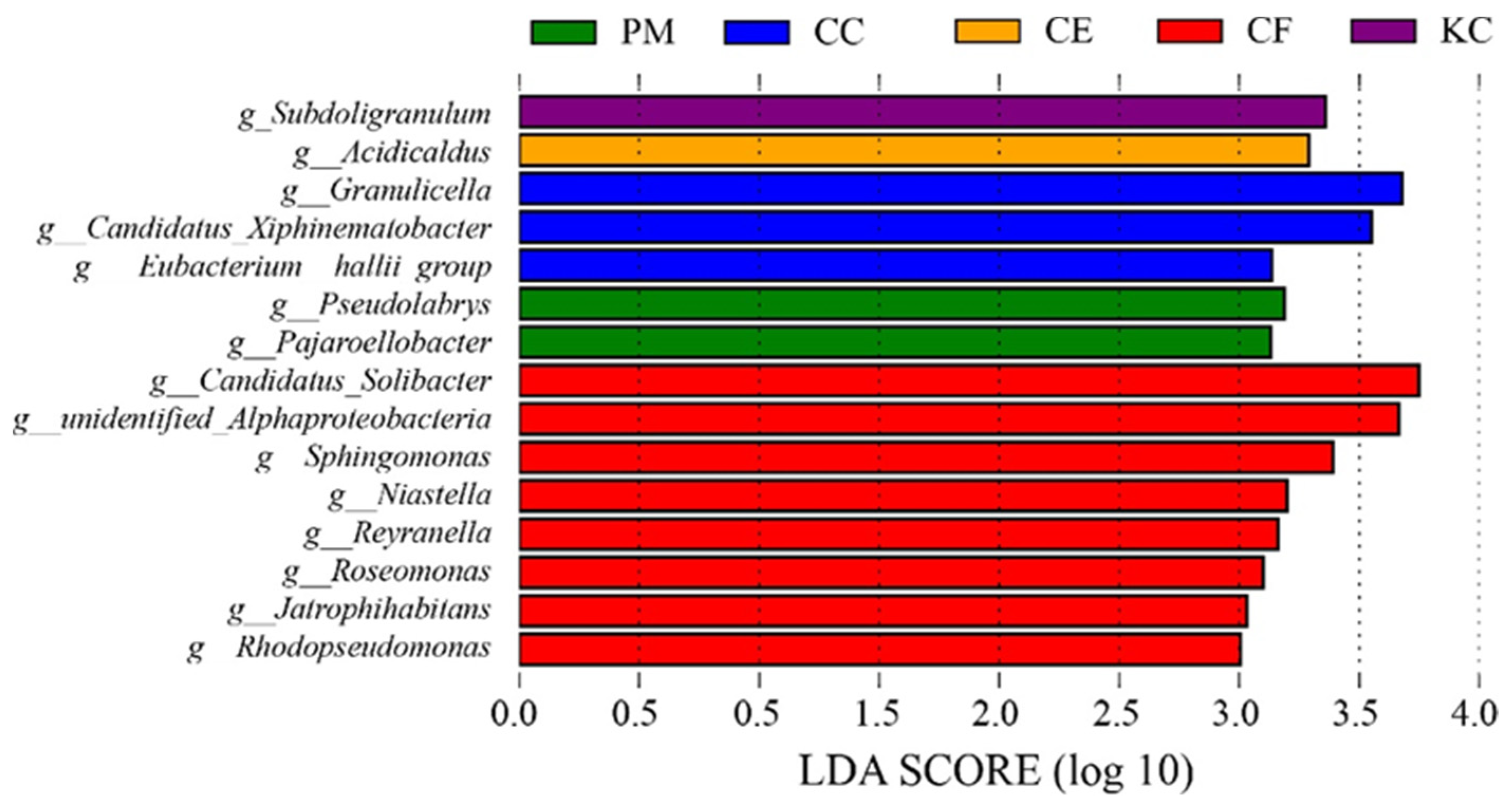
2.2. Bacterial Diversity and Community Composition in the Five Tree Species
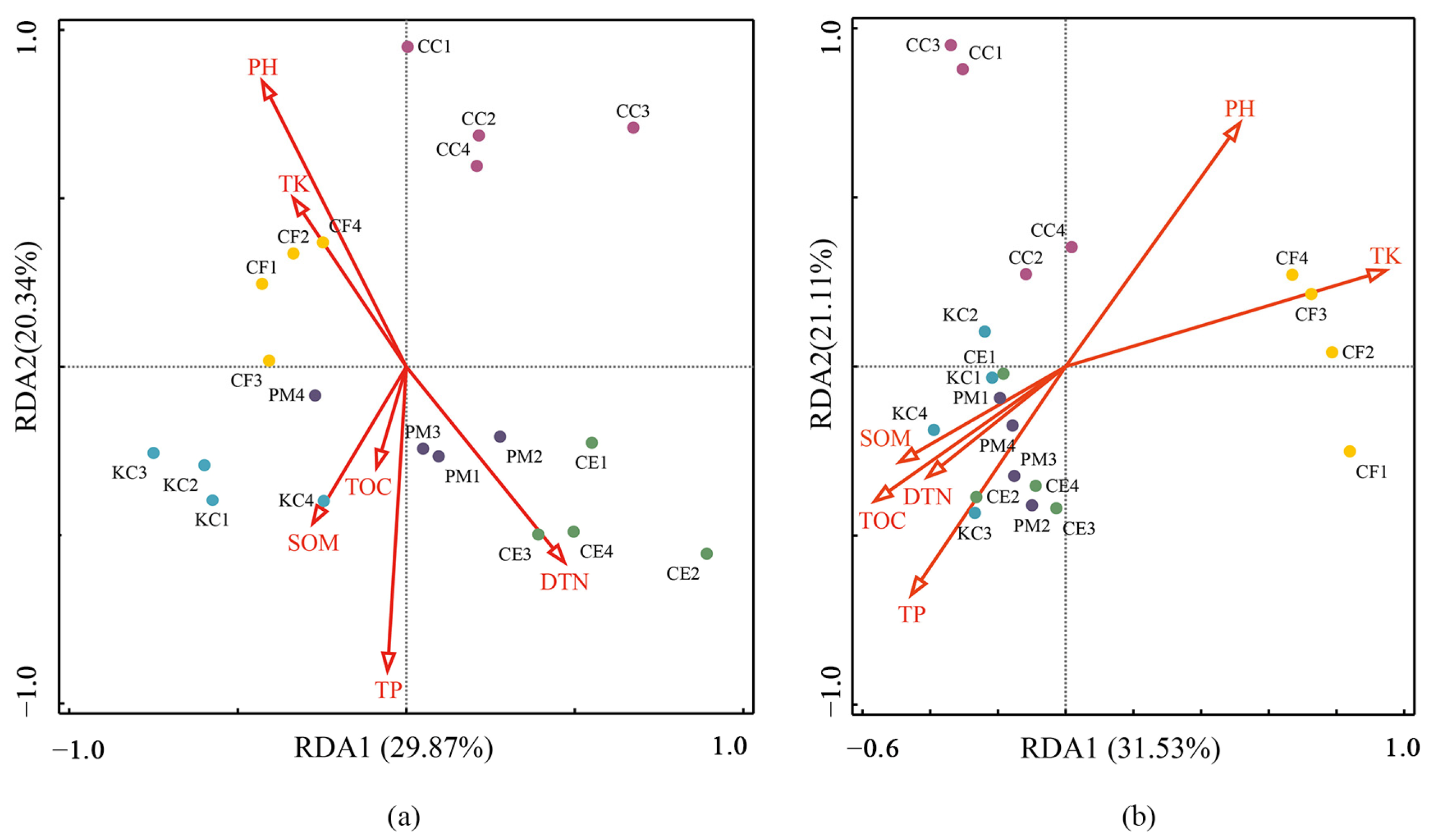
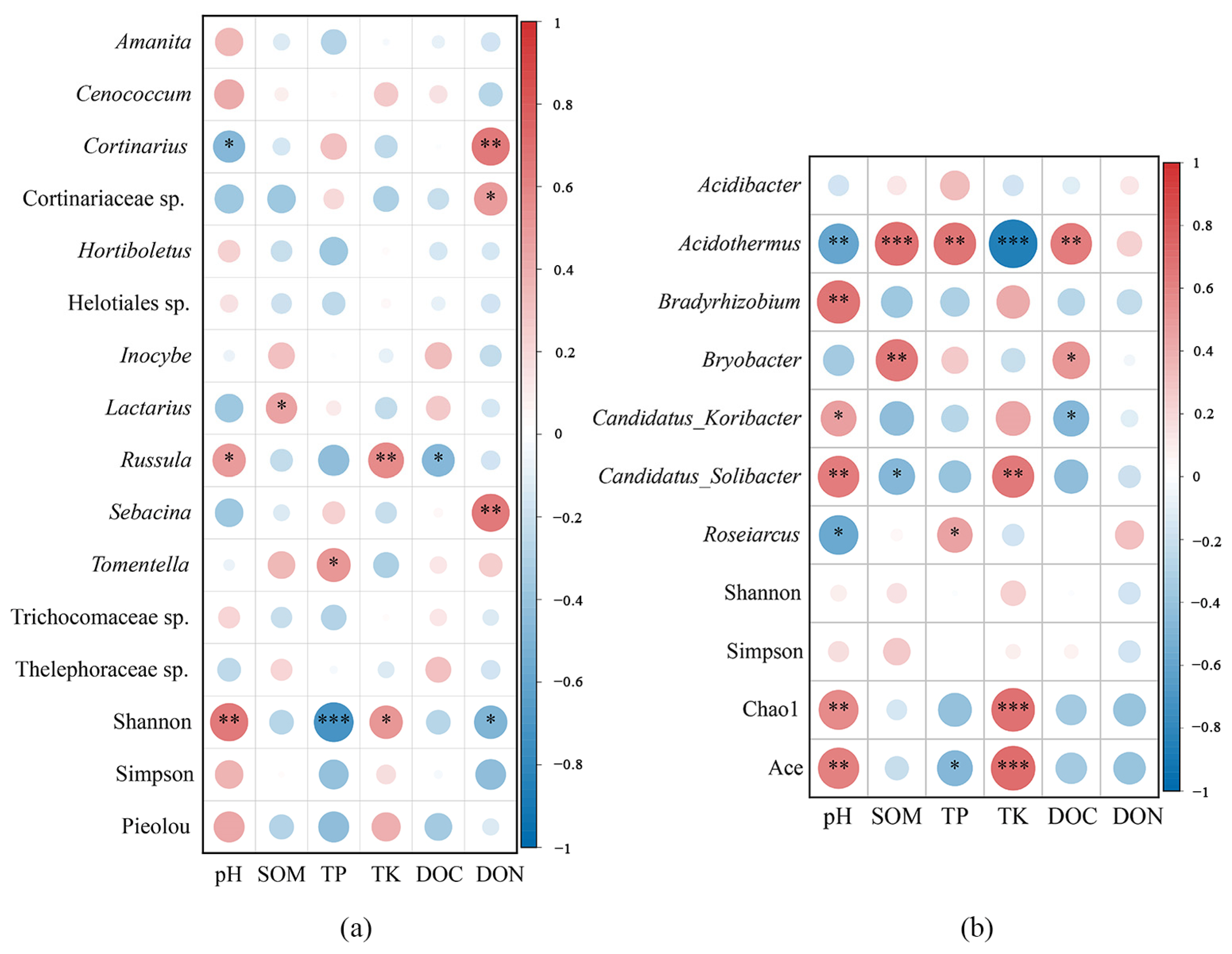
2.3. Effect of Soil Properties on ECM Fungal and Bacterial Communities
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Sample Collection and Processing
4.3. Soil Property Analysis
4.4. Identification and Data Analysis of ECM Fungi
4.5. DNA Extraction and Data Analysis of Soil Bacteria
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mohanram, S.; Kumar, P. Rhizosphere microbiome: Revisiting the synergy of plant-microbe interactions. Ann. Microbiol. 2019, 69, 307–320. [Google Scholar] [CrossRef]
- Berrios, L.; Yeam, J.; Holm, L.; Robinson, W.; Pellitier, P.T.; Chin, M.L.; Henkel, T.W.; Peay, K.G. Positive interactions between mycorrhizal fungi and bacteria are widespread and benefit plant growth. Curr. Biol. CB 2023, 33, 2878–2887.e4. [Google Scholar] [CrossRef] [PubMed]
- Futai, K.; Taniguchi, T.; Kataoka, R. Ectomycorrhizae and their importance in forest ecosystems. In Mycorrhizae: Sustainable Agriculture and Forestry; Siddiqui, Z.A., Akhtar, M.S., Futai, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 241–285. [Google Scholar]
- LLadó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef]
- Jiang, S.; Xing, Y.J.; Liu, G.C.; Hu, C.Y.; Wang, X.C.; Yan, G.Y.; Wang, Q.G. Changes in soil bacterial and fungal community composition and functional groups during the succession of boreal forests. Soil Biol. Biochem. 2021, 161, 108393. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Dang, P.; Gao, Y.; Zhu, H.L.; Zhu, H.N.; Zhao, F.; Zhao, Z. Effects of tree species and soil properties on the composition and diversity of the soil bacterial community following afforestation. For. Ecol. Manag. 2018, 427, 342–349. [Google Scholar] [CrossRef]
- Maranon-Jimenez, S.; Radujkovic, D.; Verbruggen, E.; Grau, O.; Cuntz, M.; Penuelas, J.; Richter, A.; Schrumpf, M.; Rebmann, C. Shifts in the abundances of saprotrophic and ectomycorrhizal fungi with altered leaf litter inputs. Front. Plant Sci. 2021, 12, 682142. [Google Scholar] [CrossRef]
- Okubo, A.; Matsusaka, M.; Sugiyama, S. Impacts of root symbiotic associations on interspecific variation in sugar exudation rates and rhizosphere microbial communities: A comparison among four plant families. Plant Soil 2016, 399, 345–356. [Google Scholar] [CrossRef]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Jones, M.D.; Kranabetter, J.M. Ectomycorrhizae and tree seedling nitrogen nutrition in forest restoration. New For. 2015, 46, 747–771. [Google Scholar] [CrossRef]
- Moyersoen, B.; Fitter, A.H. Presence of arbuscular mycorrhizas in typically ectomycorrhizal host species from Cameroon and New Zealand. Mycorrhiza 1999, 8, 247–253. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.; Tisserand, E.; Cebron, A.; Turpault, M.P.; Buee, M.; De Boer, W.; Leveau, J.H.J.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef]
- Florio, A.; Marechal, M.; Legout, A.; Creuse des Chatelliers, C.; Gervaix, J.; Didier, S.; Zeller, B.; Le Roux, X. Influence of biological nitrification inhibition by forest tree species on soil denitrifiers and N2O emissions. Soil Biol. Biochem. 2021, 155, 108164. [Google Scholar] [CrossRef]
- Bonfante, P.; Anca, I.-A. Plants, mycorrhizal fungi, and bacteria: A network of interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Toljander, J. Interactions between Soil Bacteria and Arbuscular Mycorrhizal Fungi; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2006. [Google Scholar]
- Nurmiaho-Lassila, E.L.; Timonen, S.; Haahtela, K.; Sen, R. Bacterial colonization patterns of intact Pinus sylvestris mycorrhizospheres in dry pine forest soil: An electron microscopy study. Can. J. Microbiol. 1997, 43, 1017–1035. [Google Scholar] [CrossRef]
- Frey, P.; Frey-Klett, P.; Garbaye, J.; Berge, O.; Heulin, T. Metabolic and genotypic fingerprinting of fluorescent pseudomonads associated with the Douglas Fir-Laccaria bicolor mycorrhizosphere. Appl. Environ. Microbiol. 1997, 63, 1852–1860. [Google Scholar] [CrossRef]
- Warmink, J.A.; Nazir, R.; van Elsas, J.D. Universal and species-specific bacterial ‘fungiphiles’ in the mycospheres of different basidiomycetous fungi. Environ. Microbiol. 2009, 11, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Deveau, A.; Palin, B.; Delaruelle, C.; Peter, M.; Kohler, A.; Pierrat, J.C.; Sarniguet, A.; Garbaye, J.; Martin, F.; Frey-Klett, P. The mycorrhiza helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytol. 2007, 175, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Nazir, R.; Tazetdinova, D.I.; van Elsas, J.D. Burkholderia terrae BS001 migrates proficiently with diverse fungal hosts through soil and provides protection from antifungal agents. Front. Microbiol. 2014, 5, 598. [Google Scholar] [CrossRef]
- Guennoc, C.M.; Rose, C.; Labbé, J.; Deveau, A. Bacterial biofilm formation on the hyphae of ectomycorrhizal fungi: A wide-spreadability under controls? FEMS Microbiol. Ecol. 2018, 94, fiy093. [Google Scholar] [CrossRef]
- Shirakawa, M.; Uehara, I.; Tanaka, M. Mycorrhizosphere bacterial communities and their sensitivity to antibacterial activity of ectomycorrhizal fungi. Microbes Environ. 2019, 34, 191–198. [Google Scholar] [CrossRef]
- Shuai, L.Y.; Xiao, S.P.; Xie, Y.P.; Chen, X.M.; Song, X.R.; Fan, T.Q.; Xie, Y.H.; Liu, W. Ecological drivers of avian diversity in a subtropical landscape: Effects of habitat diversity, primary productivity and anthropogenic disturbance. Ecol. Evol. 2022, 12, e9166. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Li, Z.; Zhang, J. Comprehensive Observations Report on Junzifeng Nature Reserves of Fujian; Xiamen University Press: Xiamen, China, 2005. [Google Scholar]
- Huang, T. Research on the development potential and realization path of forest health service in nature reserve. Issues For. Econ. 2021, 41, 658–664. [Google Scholar] [CrossRef]
- Huang, L.; Zhou, M.; Lv, J.; Chen, K. Trends in global research in forest carbon sequestration: A bibliometric analysis. J. Clean. Prod. 2020, 252, 119908. [Google Scholar] [CrossRef]
- Liu, Y.P.; Ye, X.Z.; Ye, L.Q.; Chen, X.H.; Zheng, S.Q.; Chen, S.P.; Zhang, G.F.; Liu, B. Niche and interspecific association of dominant tree species in Michelia odora community. Chin. J. Appl. Ecol. 2022, 33, 2670–2678. [Google Scholar] [CrossRef]
- Singh, B.K.; Quince, C.; Macdonald, C.A.; Khachane, A.; Thomas, N.; Al-Soud, W.A.; Sørensen, S.J.; He, Z.; White, D.; Sinclair, A.; et al. Loss of microbial diversity in soils is coincident with reductions in some specialized functions. Env. Microbiol. 2014, 16, 2408–2420. [Google Scholar] [CrossRef]
- Vitali, F.; Mastromei, G.; Senatore, G.; Caroppo, C.; Casalone, E. Long lasting effects of the conversion from natural forest to poplar plantation on soil microbial communities. Microbiol. Res. 2016, 182, 89–98. [Google Scholar] [CrossRef]
- Dong, X.; Lv, L.; Wang, W.; Liu, Y.; Yin, C.; Xu, Q.; Yan, H.; Fu, J.; Liu, X. Differences in distribution of potassium-solubilizing bacteria in forest and plantation soils in myanmar. Int. J. Environ. Res. Public Health 2019, 16, 700. [Google Scholar] [CrossRef]
- Lofgren, L.; Nguyen, N.H.; Kennedy, P.G. Ectomycorrhizal host specificity in a changing world: Can legacy effects explain anomalous current associations? New Phytol. 2018, 220, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Dawud, S.M.; Raulund-Rasmussen, K.; Ratcliffe, S.; Domisch, T.; Finér, L.; Joly, F.X.; Hättenschwiler, S.; Vesterdal, L.; Ostertag, R. Tree species functional group is a more important driver of soil properties than tree species diversity across major European forest types. Funct. Ecol. 2017, 31, 1153–1162. [Google Scholar] [CrossRef]
- Hüblová, L.; Frouz, J. Contrasting effect of coniferous and broadleaf trees on soil carbon storage during reforestation of forest soils and afforestation of agricultural and post-mining soils. J. Environ. Manag. 2021, 290, 112567. [Google Scholar] [CrossRef]
- van der Linde, S.; Suz, L.M.; Orme, C.D.L.; Cox, F.; Andreae, H.; Asi, E.; Atkinson, B.; Benham, S.; Carroll, C.; Cools, N.; et al. Environment and host as large-scale controls of ectomycorrhizal fungi. Nature 2018, 558, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Rosinger, C.; Sandén, H.; Matthews, B.; Mayer, M.; Godbold, D.L. Patterns in ectomycorrhizal diversity, community composition, and exploration types in european beech, pine, and spruce forests. Forests 2018, 9, 445. [Google Scholar] [CrossRef]
- Wu, D.; Li, T.; Wan, S. Time and litter species composition affect litter-mixing effects on decomposition rates. Plant Soil 2013, 371, 355–366. [Google Scholar] [CrossRef]
- Lu, R. Analytical Methods of Soil and Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Giltrap, N.J.; Lewis, D.H. Inhibition of growth of ectomycorrhizal fungi in culture by phosphate. New Phytol. 1981, 87, 669–675. [Google Scholar] [CrossRef]
- Dickie, I.A.; Martínez-García, L.B.; Koele, N.; Grelet, G.A.; Tylianakis, J.M.; Peltzer, D.A.; Richardson, S.J. Mycorrhizas and mycorrhizal fungal communities throughout ecosystem development. Plant Soil 2013, 367, 11–39. [Google Scholar] [CrossRef]
- Obase, K.; Douhan, G.W.; Matsuda, Y.; Smith, M.E. Progress and Challenges in Understanding the Biology, Diversity, and Biogeography of Cenococcum geophilum. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 299–317. [Google Scholar]
- Shi, Y.; Yan, T.; Yuan, C.; Li, C.; Rensing, C.; Chen, Y.; Xie, R.; Zhang, T.; Lian, C. Comparative physiological and transcriptome analysis provide insights into the response of Cenococcum geophilum, an ectomycorrhizal fungus to cadmium stress. J. Fungi 2022, 8, 724. [Google Scholar] [CrossRef]
- Yan, T.; Zhang, P.; Pang, W.; Zhang, X.; Lian, C.; Zhang, T. Effects of high temperature-triggered transcriptomics on the physiological adaptability of Cenococcum geophilum, an ectomycorrhizal fungus. Microorganisms 2022, 10, 2039. [Google Scholar] [CrossRef]
- Li, M.; Yuan, C.; Zhang, X.; Pang, W.; Zhang, P.; Xie, R.; Lian, C.; Zhang, T. The transcriptional responses of ectomycorrhizal fungus, Cenococcum geophilum, to drought stress. J. Fungi 2023, 9, 15. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, P.; Pang, W.; Zhang, Y.; Alwathnani, H.A.; Rensing, C.; Yang, W. Increased tolerance of massion’s pine to multiple-toxic-metal stress mediated by ectomycorrhizal fungi. Plants 2023, 12, 3179. [Google Scholar] [CrossRef]
- Wurentaogesi; Han, S.; Yan, W. The relationship between natural infection rate of Cenococcum geophilum and vegetation, rhizosphere soil factors. Chin. Agric. Sci. Bull. 2012, 28, 47–51. [Google Scholar] [CrossRef]
- Truong, C.; Gabbarini, L.A.; Corrales, A.; Mujic, A.B.; Escobar, J.M.; Moretto, A.; Smith, M.E. Ectomycorrhizal fungi and soil enzymes exhibit contrasting patterns along elevation gradients in southern Patagonia. New Phytol. 2019, 222, 1936–1950. [Google Scholar] [CrossRef]
- Horton, B.M.; Glen, M.; Davidson, N.J.; Ratkowsky, D.; Close, D.C.; Wardlaw, T.J.; Mohammed, C. Temperate eucalypt forest decline is linked to altered ectomycorrhizal communities mediated by soil chemistry. For. Ecol. Manag. 2013, 302, 329–337. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, L.; Liang, T.; Man, J.; Wang, Y.; Li, Y.; Chen, H.; Zhang, T. Deciphering rhizosphere microbiome assembly of castanea henryi in plantation and natural forest. Microorganisms 2021, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kang, W.; Liu, S.; Yin, H.; Lyu, Q.; Su, Y.; Liu, J.; Liu, J.; Fan, C.; Chen, G.; et al. Diversity of ectomycorrhizal fungal communities in four types of stands in Pinus massoniana plantation in the west of China. Forests 2021, 12, 719. [Google Scholar] [CrossRef]
- Yang, X.; Xu, M.; Zhang, J.; Wen, C.-Y.; Zhang, J. Effects of resin tapping on ectomycorrhizal fungal community composition and structure of Pinus massoniana in subtropical mountain forest ecosystems in southwestern China. For. Ecol. Manag. 2023, 540, 121030. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Jose, P.A.; Maharshi, A.; Jha, B. Actinobacteria in natural products research: Progress and prospects. Microbiol. Res. 2021, 246, 126708. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cui, Y.; Ma, D.; Li, X.; Liu, L. Spatial variation of microbial community structure and its driving environmental factors in two forest types in permafrost region of Greater Xing′an Mountains. Sustainability 2022, 14, 9284. [Google Scholar] [CrossRef]
- Izumi, H.; Anderson, I.C.; Alexander, I.J.; Killham, K.; Moore, E.R. Endobacteria in some ectomycorrhiza of Scots pine (Pinus sylvestris). FEMS Microbiol. Ecol. 2006, 56, 34–43. [Google Scholar] [CrossRef]
- Lehr, N.A.; Schrey, S.D.; Bauer, R.; Hampp, R.; Tarkka, M.T. Suppression of plant defence response by a mycorrhiza helper bacterium. New Phytol. 2007, 174, 892–903. [Google Scholar] [CrossRef]
- Sarathambal, C.; Dinesh, R.; Srinivasan, V.; Sheeja, T.E.; Jeeva, V.; Manzoor, M. Changes in bacterial diversity and composition in response to co-inoculation of arbuscular mycorrhizae and zinc-solubilizing bacteria in turmeric rhizosphere. Curr. Microbiol. 2021, 79, 4. [Google Scholar] [CrossRef]
- Li, Y.-C.; Liu, B.-R.; Li, S.-H.; Qin, H.; Fu, W.-J.; Xu, Q.-F. Shift in abundance and structure of soil ammonia-oxidizing bacteria and archaea communities associated with four typical forest vegetations in subtropical region. J. Soils Sediments 2014, 14, 1577–1586. [Google Scholar] [CrossRef]
- Gui, H.; Purahong, W.; Hyde, K.D.; Xu, J.; Mortimer, P.E. The arbuscular mycorrhizal fungus Funneliformis mosseae alters bacterial communities in subtropical forest soils during litter decomposition. Front. Microbiol. 2017, 8, 1120. [Google Scholar] [CrossRef]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef]
- Zhang, R.; Rong, L.; Zhang, L. Soil nutrient variability mediates the effects of erosion on soil microbial communities: Results from a modified topsoil removal method in an agricultural field in Yunnan plateau, China. Environ. Sci. Pollut. Res. 2022, 29, 3659–3671. [Google Scholar] [CrossRef]
- Boersma, F.G.H.; Warmink, J.A.; Andreote, F.A.; van Elsas, J.D. Selection of Sphingomonadaceae at the base of Laccaria proxima and Russula exalbicans fruiting bodies. Appl. Environ. Microbiol. 2009, 75, 1979–1989. [Google Scholar] [CrossRef]
- Agerer, R. Colour Atlas of Ectomycorrhizae; Einhorn-Verlag Eduard Dietenberger: Schwabisch Gmund, Germany, 1987. [Google Scholar]
- Lian, C.; Hogetsu, T.; Matsushita, N.; Guerin-Laguette, A.; Suzuki, K.; Yamada, A. Development of microsatellite markers from an ectomycorrhizal fungus, Tricholoma matsutake, by an ISSR-suppression-PCR method. Mycorrhiza 2003, 13, 27–31. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Calaça, F.J.S.; Bustamante, M.M. Richness of arbuscular mycorrhizal fungi (Glomeromycota) along a vegetation gradient of Brazilian Cerrado: Responses to seasonality, soil types, and plant communities. Mycol. Prog. 2022, 21, 27. [Google Scholar] [CrossRef]
- López, C.L.; Mayta, C.; Naoki, K.; Quezada, J.A.; Hensen, I.; Gallegos, S.C. Bracken fern does not diminish arbuscular mycorrhizal fungus inoculum potential in tropical deforested areas. Mycorrhiza 2022, 32, 123–131. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Chao1 Index | ACE Index | Simpson Index | Shannon Index |
|---|---|---|---|---|
| PM | 1637.22 ± 155.21 ab | 1647.12 ± 120.8 b | 0.99 ± 0 a | 7.96 ± 0.2 a |
| CC | 1654.06 ± 127.23 ab | 1695.17 ± 150.16 ab | 0.99 ± 0 a | 7.73 ± 0.29 a |
| CE | 1552.47 ± 136.08 b | 1587.25 ± 141.19 b | 0.99 ± 0.01 a | 7.86 ± 0.19 a |
| CF | 1898.41 ± 118.04 a | 1915.39 ± 68.05 a | 0.99 ± 0 a | 8.02 ± 0.12 a |
| KC | 1646.99 ± 69.74 ab | 1665.88 ± 50.9 b | 0.99 ± 0 a | 7.85 ± 0.23 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, W.; Zhang, P.; Zhang, Y.; Zhang, X.; Huang, Y.; Zhang, T.; Liu, B. The Ectomycorrhizal Fungi and Soil Bacterial Communities of the Five Typical Tree Species in the Junzifeng National Nature Reserve, Southeast China. Plants 2023, 12, 3853. https://doi.org/10.3390/plants12223853
Pang W, Zhang P, Zhang Y, Zhang X, Huang Y, Zhang T, Liu B. The Ectomycorrhizal Fungi and Soil Bacterial Communities of the Five Typical Tree Species in the Junzifeng National Nature Reserve, Southeast China. Plants. 2023; 12(22):3853. https://doi.org/10.3390/plants12223853
Chicago/Turabian StylePang, Wenbo, Panpan Zhang, Yuhu Zhang, Xiao Zhang, Yanbin Huang, Taoxiang Zhang, and Bao Liu. 2023. "The Ectomycorrhizal Fungi and Soil Bacterial Communities of the Five Typical Tree Species in the Junzifeng National Nature Reserve, Southeast China" Plants 12, no. 22: 3853. https://doi.org/10.3390/plants12223853