
Quantitative Analysis of Seed Surface Tubercles in Silene Species
by
 , , and
, , and
José Luis Rodríguez-Lorenzo
1,
José Javier Martín-Gómez
2,
Ana Juan
3,
Ángel Tocino
4 and
Emilio Cervantes
2,*
1
Plant Developmental Genetics, Institute of Biophysics v.v.i, Academy of Sciences of the Czech Republic, Královopolská 135, 612 65 Brno, Czech Republic
2
Instituto de Recursos Naturales y Agrobiología, Consejo Superior de Investigaciones Científicas, Cordel de Merinas 40, 37008 Salamanca, Spain
3
Departamento de Ciencias Ambientales y Recursos Naturales, Universidad de Alicante, 03690 Alicante, Spain
4
Departamento de Matemáticas, Facultad de Ciencias, Universidad de Salamanca, Plaza de la Merced 1-4, 37008 Salamanca, Spain
*
Author to whom correspondence should be addressed.
Plants 2023, 12(19), 3444; https://doi.org/10.3390/plants12193444
Submission received: 21 August 2023
/
Revised: 14 September 2023
/
Accepted: 27 September 2023
/
Published: 29 September 2023
(This article belongs to the Special Issue Seed Biology at the System Level)
Abstract
:In the Caryophyllaceae, seed surfaces contain cell protrusions, of varying sizes and shapes, called tubercles. Tubercles have long been described in many species, but quantitative analyses with measurements of size and shape are lacking in the literature. Based on optical photography, the seeds of Silene were classified into four types: smooth, rugose, echinate and papillose. Seeds in each of these groups have characteristic geometrical properties: smooth seeds lack tubercles and have the highest values of circularity and solidity in their lateral views, while papillose seeds have the largest tubercles and lowest values of circularity and solidity both in lateral and dorsal views. Here, tubercle width, height and slope, maximum and mean curvature values and maximum to mean curvature ratio were obtained for representative seeds of a total of 31 species, 12 belonging to Silene subg. Behenantha and 19 to S. subg. Silene. The seeds of the rugose type had lower values of curvature. Additionally, lower values of curvature were found in species of S. subg. Silene in comparison with S. subg. Behenantha. The seeds of S. subg. Behenantha had higher values of tubercle height and slope and higher values of maximum and average curvature and maximum to mean curvature ratio.
1. Introduction
Caryophyllaceae Juss. contains about 100 genera and ca. 2000 species characterized by anatropous to campylotropous ovules [1] and a peripheral position of the embryo in the developing seed [2]. Although single-seeded fruits seem to be the primitive characteristic in the family [3,4], most of the genera in the Alsinoideae and Caryophylloideae subfamilies have dry-dehiscent multi-seed capsules as their fruits [3]. The seed surfaces present micromorphological features such as cell perimeter and tubercle shape. These attributes have been used for species description and taxonomic purposes in diverse genera [5,6,7,8,9,10,11,12,13,14,15].
Silene L., with ca. 800 species, is the largest genus in the Caryophyllaceae. Silene seeds have a characteristic shape in the lateral view that may be described and quantified by comparison with a cardioid and derived figures [16,17,18].
Surface characteristics of Silene seeds have been traditionally analyzed via Scanning Electron Microscopy (SEM). Protuberances of cells at the seed surface are called tubercles, and early work indicated that they could be absent or present, and, according to their properties, the protuberances provided the basis to classify seeds into five types [19]. Another work highlighted two important aspects in the analysis of surface morphology in Silene seeds. The first concerns the level of conservation of the type between populations of the same species. Infraspecific variation between different populations was found in S. italica (L.) Pers., S. spinescens Sm., S. nutans L. and S. coutinhoi Rothm. & P.Silva [19]. This aspect was also found in Geocarpon uniflorum (Walter) E.E. Schill. (referred to as Arenaria uniflora (Walter) Muhl. in [7]), and in Silene, it was addressed more recently by Tabaripur et al. [20], who reported infraspecific variation in S. odontopetala Fenzl.
A second aspect of importance involves the nomenclature of the tubercle shape diversity. The descriptors used by Ghazanfar for their description (round, cylindrical, conical, clavate, umbonate) had different success in later work by other groups based on SEM images. The group of Yildiz [21,22] retained the use of the terms, rounded, cylindrical and conical, replacing umbonated with mamillated and indicating that this characteristic is common in S. sect. Siphonomorpha as well. Ocaña et al. [23] distinguished between tubercles and papillae, the latter corresponding to umbonate tubercles of Ghazanfar and to mammillae of Dadandi and Yildiz. The group of Abbas Gholipour classified the seed coat as papillose, tuberculate or smooth [24], similar to the categories of Hoseini et al. [25]: flat, mammalian, echinate and tuberculate, where smooth means flat and papillose is equivalent to mammalian. Both the study of variation and the precise description of tubercles will benefit from the application of quantitative methods of tubercle analysis.
SEM photographs were the source with which to classify seeds by the shape of the tubercles at their surface in several species [26,27,28,29,30,31,32,33,34]. Nevertheless, optical photographs can be useful for seed surface description, and their results can be comparable with those of SEM. The observation of optical photographs of Silene seeds allowed us to differentiate between four groups, based on the tubercle structure: smooth, rugose, echinate and papillose [35,36]. Associated with these groups were differences in morphological measurements; thus, smooth seeds, without tubercles, had the highest values of circularity and solidity in their lateral views, while papillose seeds, with the largest tubercles, had the lowest values of circularity and solidity both in the lateral and dorsal views [35,36].
In the scientific literature reviewed here, the description of tubercles is based on qualitative features, but a quantitative description based on their measurements is lacking. A direct quantitative analysis of tubercle geometrics based on their width, height and curvature values is given for the first time in this work. Curvature represents the rate at which the unit tangent vector is changing with respect to arc length [37,38,39,40,41]. Curvature measurements are based on previous methods developed for roots analysis in Arabidopsis Heynh. in Holl & Heynh. (Brassicaceae) [37,38], as well as in wheat and grape seeds [39,40]. Curvature was measured in Silene seeds [41] and curvature values in the tubercles of S. chlorifolia Sm., a representative of rugose seeds, were compared with those in a population of S. latifolia Poir. Maximum and mean curvature values were higher in S. latifolia, suggesting that higher curvature values may be a property of echinate seeds in contrast with rugose seeds. In this work, we have analyzed the width and height of the tubercles, as well as curvature values in 31 representative species of Silene. The measurements of width, height and maximum and mean curvature revealed differences between rugose and echinate groups, as well as between the subgenera, Silene and Behenantha, and provide a basis for the quantitative analysis of tubercles and the relationship of tubercle type and taxonomic position.
2. Results
The results are organized into four subsections. The first one contains the comparison between rugose- and echinate-type seeds for all the tubercle measurements. These include width, height, slope, maximum curvature, mean curvature and the ratio of maximum and mean curvature for representative tubercles in 21 and 10 species of the rugose and echinate groups, respectively. The second and third subsections contain the comparison between species of each seed type, describing examples of the measurements in representative species. Finally, the fourth subsection contains the comparison of tubercle measurements between Silene species belonging to subg. Behenantha and Silene.
2.1. Comparison between Rugose and Echinate Types
On the basis of 21 species characterized by rugose seeds and 11 species with echinate seeds, the width at the base, height and slope, maximum and average curvature values and the ratio between maximum and average curvature values were calculated for a total of 393 tubercles. For each species, the number of seeds was between 1 and 3, and a total of 49 seeds were analyzed (see Figure A1 in Appendix A). ANOVA was carried out to compare all these measurements between the rugose and echinate types (Table 1).
Significant differences (p < 0.05) were found for all measurements with higher values for tubercle width at the base in the rugose seeds, and higher values for all the other measurements in the echinate seeds.
2.2. Comparison between Rugose Species
Table 2 contains the results of the ANOVA for the comparison of tubercle measurements between the 21 species belonging to the rugose seed type. Significant differences (p < 0.05) were found for all measurements. Tubercle width was between 45.0 (S. inaperta) and 138.7 μ (S. marizii), and tubercle height was between 9.5 (S. inaperta) and 28.0 μ (S. acutifolia). Tubercle slopes varied between 28.0 (S. marizii) and 63.4 (S. spinescens). Maximum curvature values were between 23.7 (S. integripetala) and 85.2 μ−1 (S. spinescens), with most of the values in the range of 20–50 μ−1. Mean curvature values were in the range of 12.7 (S. nocturna) to 48.1 μ−1 (S. spinescens). Maximum/mean curvature ratios were between 1.2 (S. caryophylloides) and 2.5 (S. squamigera subsp. vesiculifera), and only two species had values above 2: S. nocturna and S. squamigera subsp. vesiculifera. Few species had differences in curvature either for maximum or mean curvature values. Differences related to maximum curvature were detected between the group of lower values formed by S. bupleuroides, S. chlorantha, S. chlorifolia, S. foliosa, S. frivaldskiana, S. integripetala, S. koreana, S. marizii and S. nocturna and the group of species with higher values (S. otites, S. spinescens and S. squamigera subsp. vesiculifera). In relation to mean curvature, lower values were found in S. nocturna and higher values in S. otites and S. spinescens. Finally, the ratio between maximum and average curvatures showed lower values in S. caryophylloides, S. dinarica and S. integripetala, and higher values in S. squamigera subsp. vesiculifera.
The following sections describe the results obtained for six tubercles from representative species in this group. The data are grouped in three parts, corresponding to species with the lowest values of maximum curvature (S. bupleuroides, S. chlorantha, S. frivaldskiana, S. integripetala), species with the highest values of maximum curvature (S. spinescens and S. squamigera subsp. vesiculifera), and a separate section for S. gigantea, a species that presented two types of tubercles. The main aspects observed are indicated, as well as when these are shared by other species. The differences between the data presented in Table 2 and those in the subsections below are because the following subsections contain data for the first six tubercles measured for each species. The Mathematica®.nb files are provided as supplementary files (See Supplementary Materials).
2.2.1. Rugose Species with Lower Maximum Curvature Values
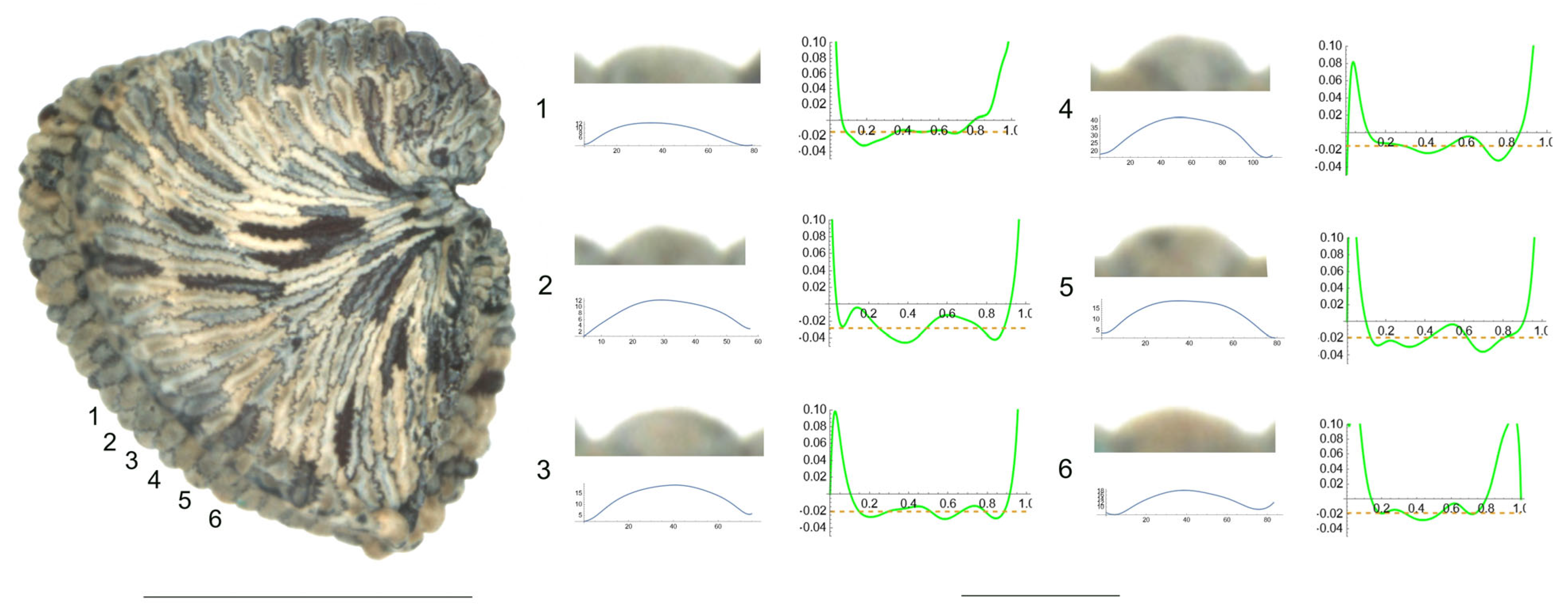
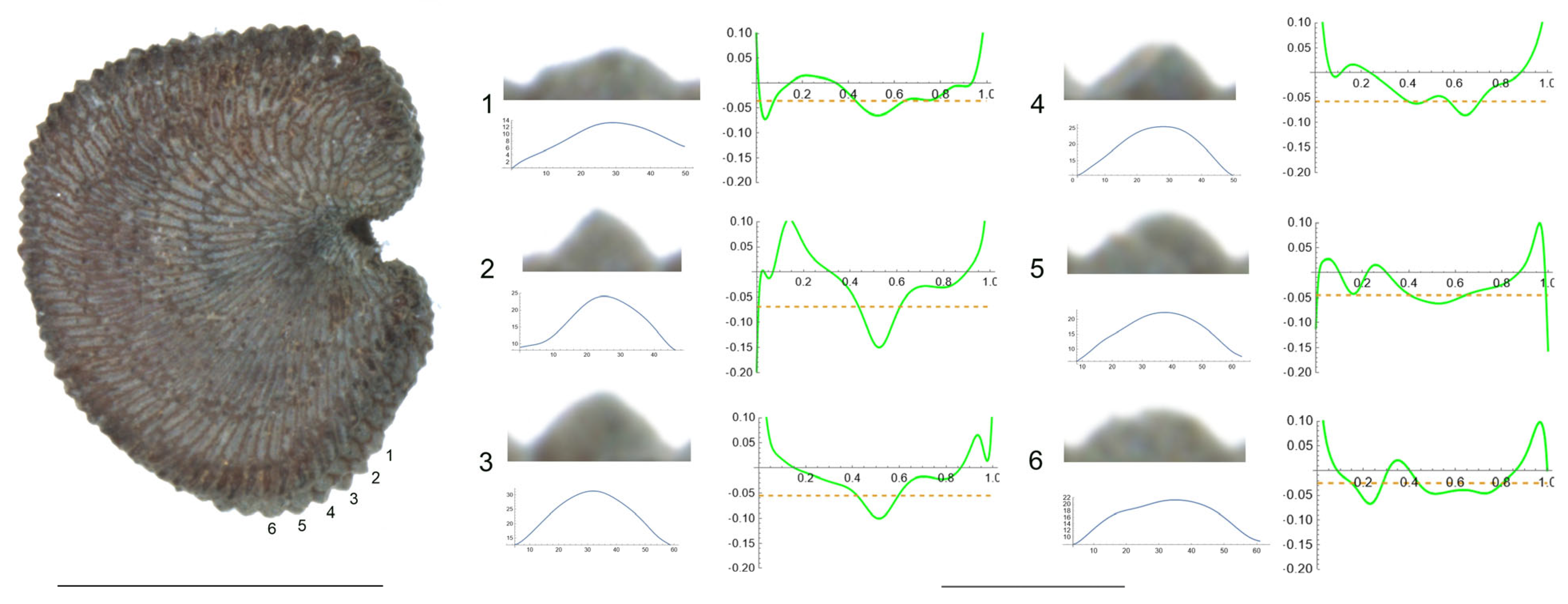
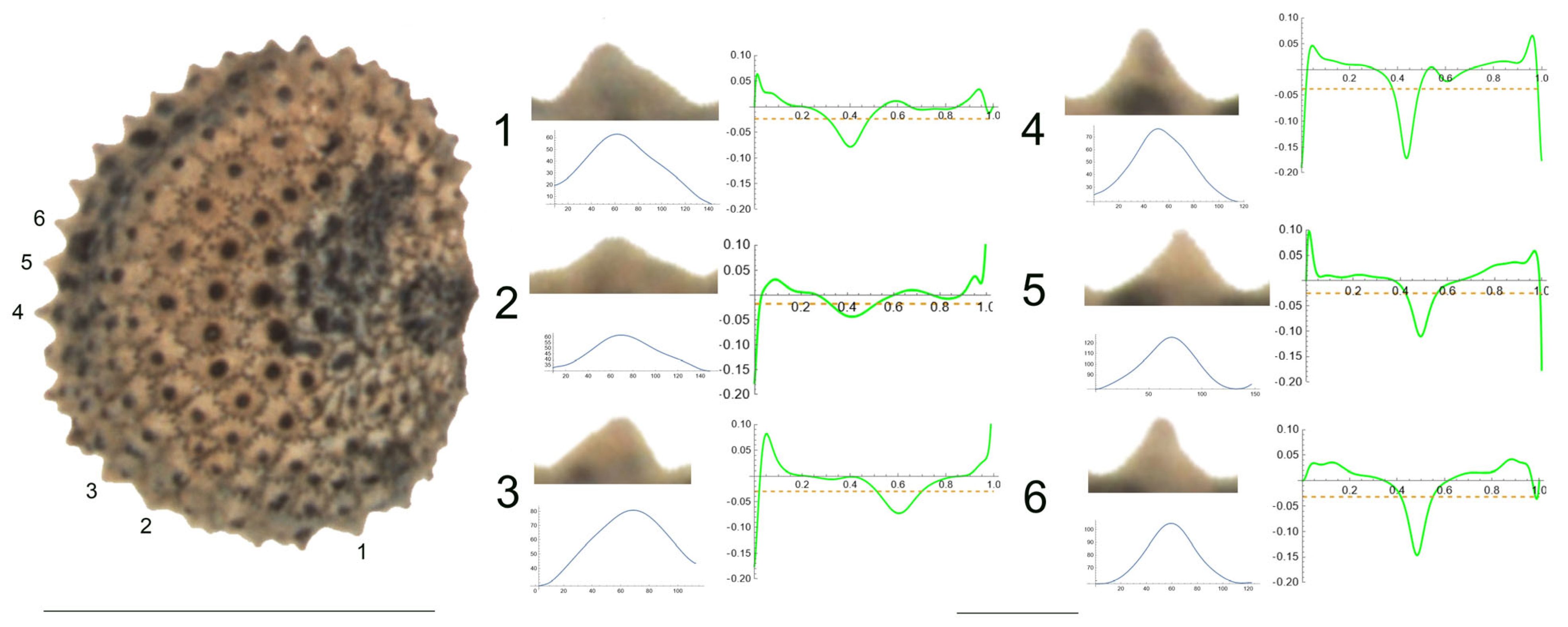
- S. bupleuroides Kom.
The tubercles examined of S. bupleuroides are rather low and flat, with low values of slope, and their maximum curvature values are deviated from a central position (Figure 1). Table 3 contains the values for tubercle width, height, slope, maximum and average curvature for six tubercles. Tubercle 1 showed the lowest values of height and slope (15 and 34.5, respectively), whereas the highest values were found for tubercle 4 (23 and 51.7, respectively). The maximum values for curvature and mean curvature corresponded to tubercle 2 (45.7 and 28.5, respectively).
- 2.
- S. chlorantha (Willd.) Ehrh.
The tubercles of S. chlorantha are irregular in size and shape, resembling deformed arcs of circumference (Figure 2). Their curvature plots resemble horizontally placed curly brackets with multiple maximum values located in both central and lateral positions. Table 4 contains the values for tubercle width, height, slope, maximum and average curvature for each of these tubercles. Tubercles 4 and 2 showed the lowest and highest values for height and slope, respectively. Changes in the direction of the slope were observed in most of the tubercles.
- 3.
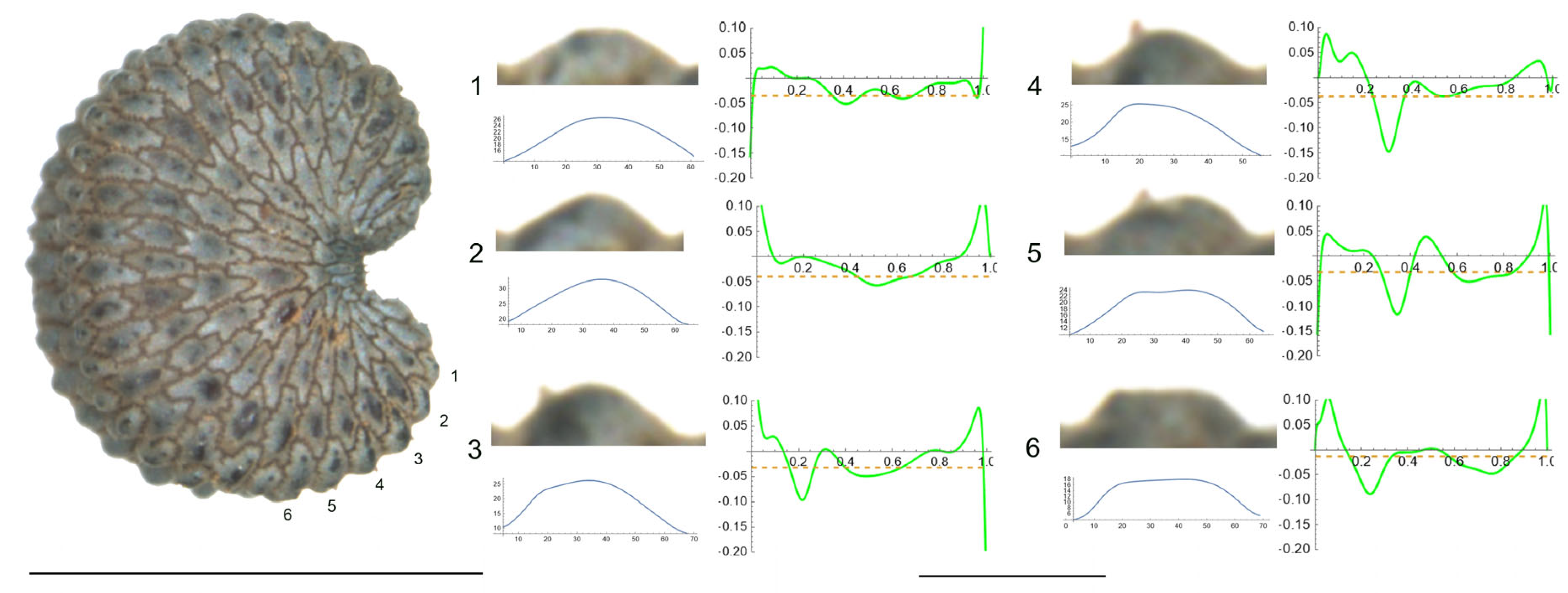
- S. frivaldskiana Hampe.
The tubercles studied of S. frivaldskiana are rather low and smooth, with low values of slope, and maximum curvature values maintained through most of their length in a central position (Figure 3 and Table 5). The measured features are, in general, quite similar among the tubercles, with the maximum differences in width found between tubercles 4 and 3 (63 and 83, respectively), in height and slope for tubercles 2 and 4. The ratio maximum/mean curvature is low (ranging from 1.2 to 1.6), indicating that both values are quite similar. Tubercles 4 and 5, with maximum and average curvature values close to 20, resemble the arc of a circumference of radium inverse to the curvature value, in this case, of 50 μ approximately.
- 4.
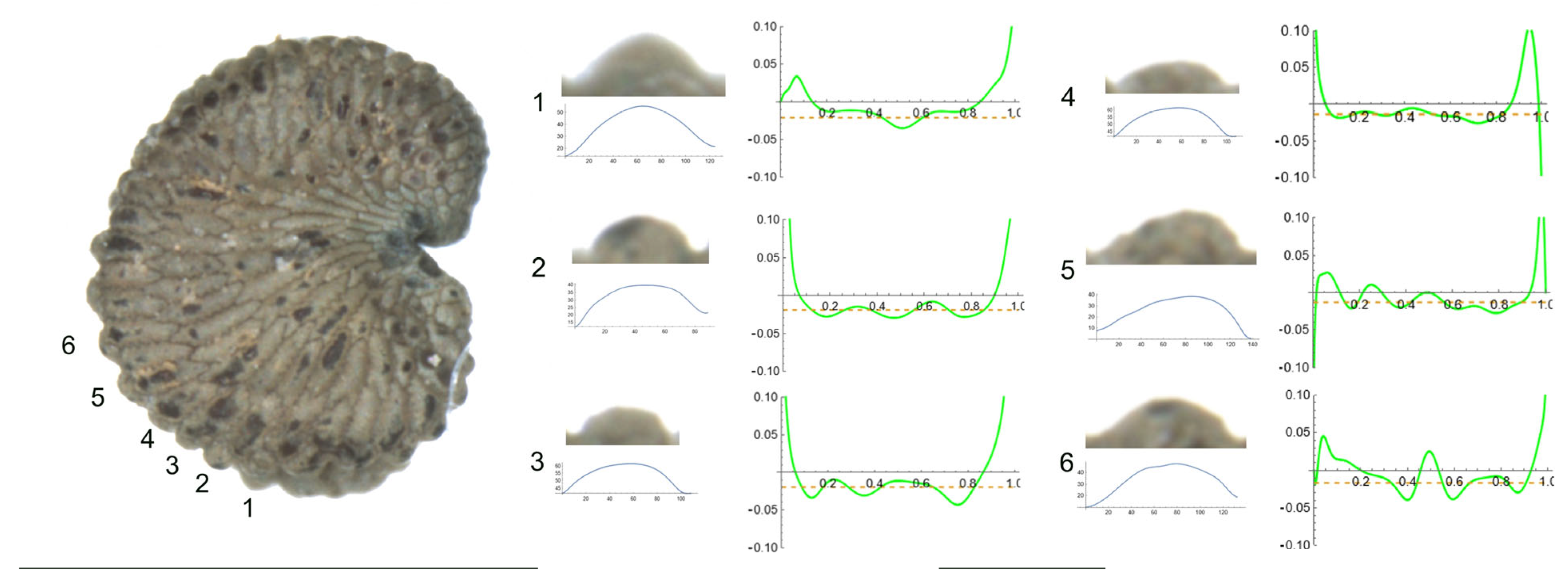
- S. integripetala Bory & Chaub.
Figure 4 contains the lateral view of a seed of S. integripetala with the indication of the tubercles analyzed in this section (numbered from 1 to 6), and the variation of curvature. The data in Table 6 reveal a gradation in their shape, from a central apical point with maximum curvature (in tubercle 1), to more plane-like (tubercles 2, 4, 5 and 6) and finally, a tubercle with two major curvature points at the sides (tubercle 3). Additionally, tubercle 1 is characterized by the lowest width and height, and tubercle 3 is the highest, with the maximum slope values.
As with most tubercles in this section, the ratio maximum to average curvature is low (1.4), indicating that both values are similar along the tubercle length and, thus, that the tubercle resembles the arc of a circumference of radium inverse to the mean curvature value, in this case, between 40 and 50 μ.
2.2.2. Rugose Species with High Curvature Values
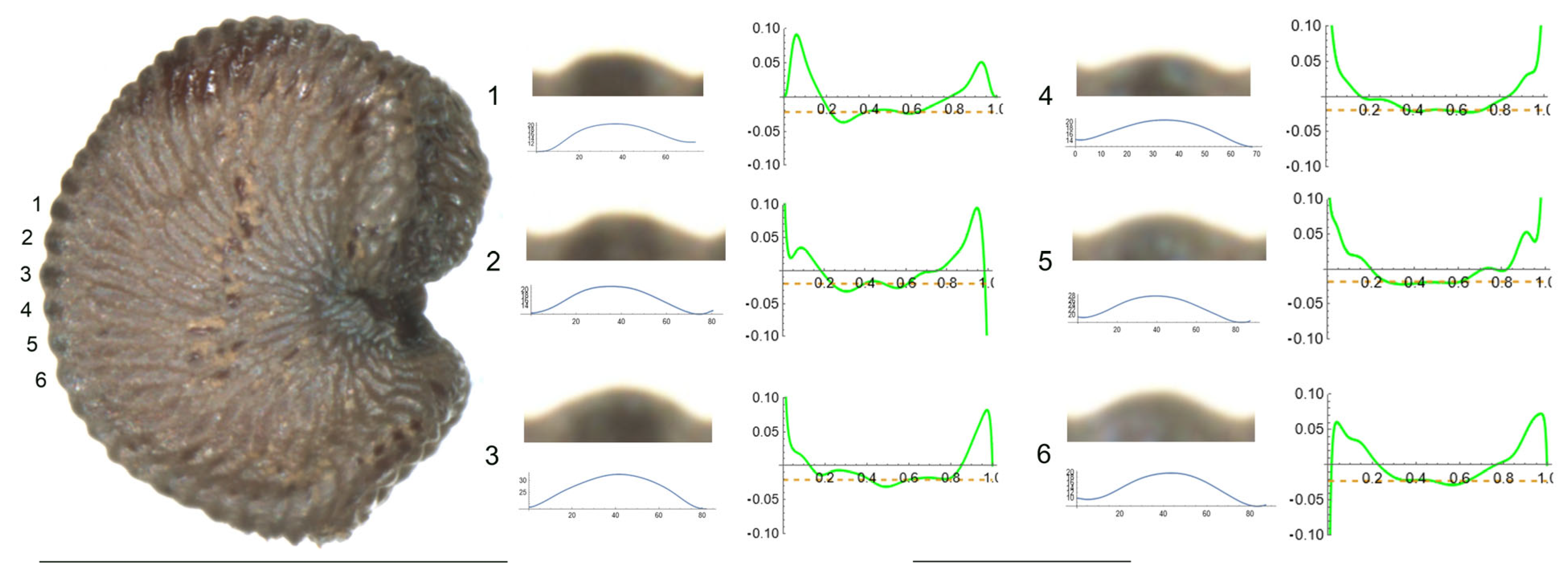
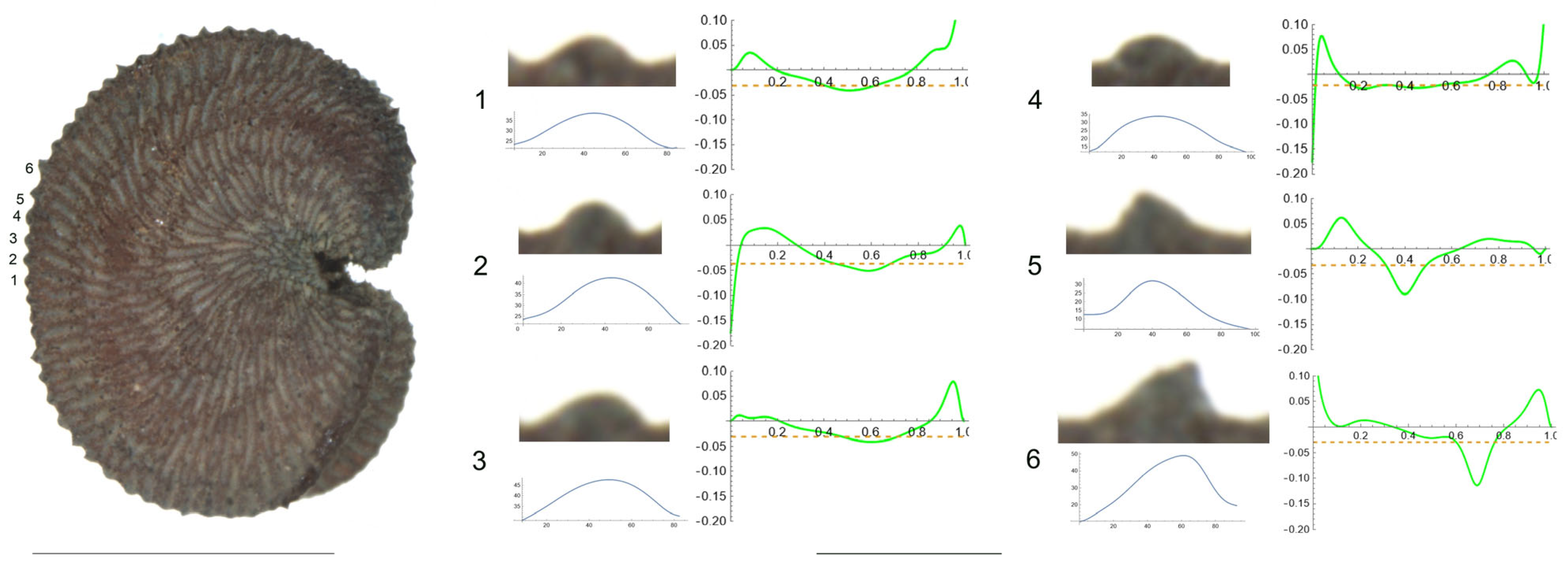
- S. spinescens Sm.
Figure 5 presents a seed of S. spinescens in the lateral view. Tubercles are numbered from 1 to 6. Table 7 contains the values for tubercle length, height, slope, maximum and average curvature for each of these tubercles. This seed provides an example of two types of tubercles: rounded, typically of the rugose type (tubercle numbers 1, 4, 5 and 6) and more pointed, with higher curvature values, tending towards echinate (2 and 3). The measured values also support this differentiation since the maximum curvature values were observed for tubercles 2 and 3.
- 2.
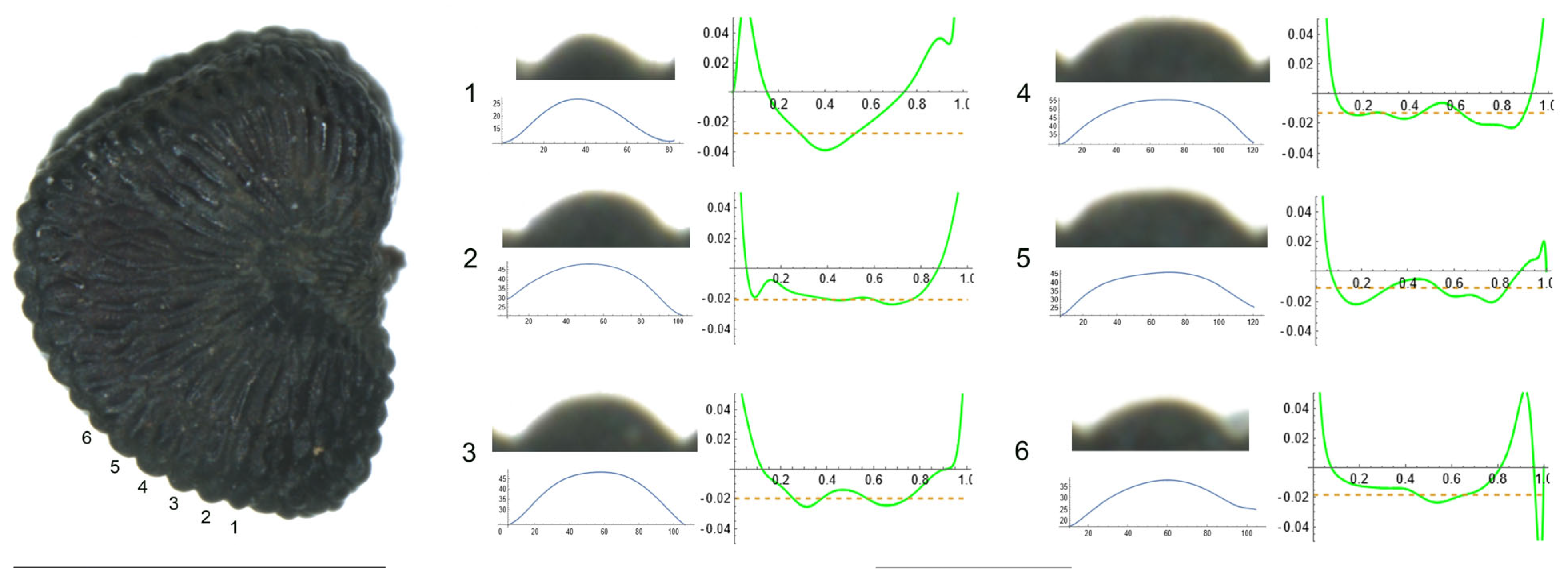
- S. squamigera Boiss. subsp. vesiculifera (J.Gay ex Boiss.) Coode and Cullen
Figure 6 presents a seed of S. squamigera subsp. vesiculifera in the lateral view. Tubercles are numbered from 1 to 6. No clear acute tubercles were observed, and the curvature shape is mostly rounded. However, tubercles 3, 4, 5 and 6 were characterized by the presence of a certain type of secondary excrescence, the presence of which appears to affect to the maximum curvature variable (Table 8). The remaining tubercles have lower curvature values.
2.2.3. Rugose Species with Two Types of Tubercles
- S. gigantea (L.) L.
Figure 7 contains a seed of S. gigantea in the lateral view. Tubercles are numbered from 1 to 6. Table 9 shows the values for tubercle length, height, slope and maximum and average curvature for each of these tubercles. The analysis of the curvature, based on Bézier curves, revealed two morphological types of tubercles: (i) rounded, with lower curvature values (tubercles 1 to 4) and (ii) acute, with a peak at the point of maximum curvature (tubercles 5 and 6). These latter two tubercles resemble those typical of the echinate type. The highest values for most the measured variables (H, S, maximum curvature and ratio maximum curvature/mean curvature) were found for tubercles 5 and 6, but for width, tubercle 2 (Table 9).
2.3. Comparison between Echinate Species
Table 10 contains the results of the ANOVA for the comparison of tubercle measurements among 10 species belonging to the echinate seed type. The differences between species were significant for all measurements. Tubercle width was between 42.2 (S. yunnanensis) and 91.2 microns (S. samojedorum), and tubercle height was in the range of 23.7 (S. longicilia) to 83.3 (S. petersonii). Tubercle slope varied between 67.3 (S. longicilia) and 278.5 (S. yunnanensis). Maximum curvature values were between 62.6 (S. dichotoma) and 219.2 (S. samojedorum). Mean curvature values were in the range of 20.5 (S. caroliniana) to 91.5 (S. samojedorum). The maximum/mean curvature ratio was between 1.6 (S. dichotoma) and 6.1 (S. caroliniana). There were large differences between species for all measurements.
According to the maximum curvature values, two groups were distinguishable, corresponding, respectively, to species with lower and higher maximum curvature values. The group of higher maximum curvature values comprised S. behen, S. samojedorum and S. yunnanensis, while the group of lower maximum curvature values comprised S. ciliata, S. dichotoma and S. longicilia.
According to the mean curvature values, two groups were distinguishable, corresponding, respectively, to species with lower and higher mean curvature values. The group of higher mean curvature values comprised S. fabaria, S. petersonii, S. samojedorum and S. yunnanensis, while the group of lower mean curvature values comprised S. aprica subsp. oldhamiana, S. caroliniana, S. ciliata, S. dichotoma and S. longicilia.
Finally, the ratio between maximum and average curvatures revealed two groups comprising S. caroliniana (higher value) and the group formed by S. ciliata, S. dichotoma, S. fabaria, S. longicilia and S. samojedorum.
The following sections describe the results of representative species in the above groups: S. aprica subsp. oldhamiana, S. caroliniana S. ciliata (lower maximum curvature) and S. yunnanensis (higher maximum curvature values).
2.3.1. Echinate Species with Low Maximum Curvature Values
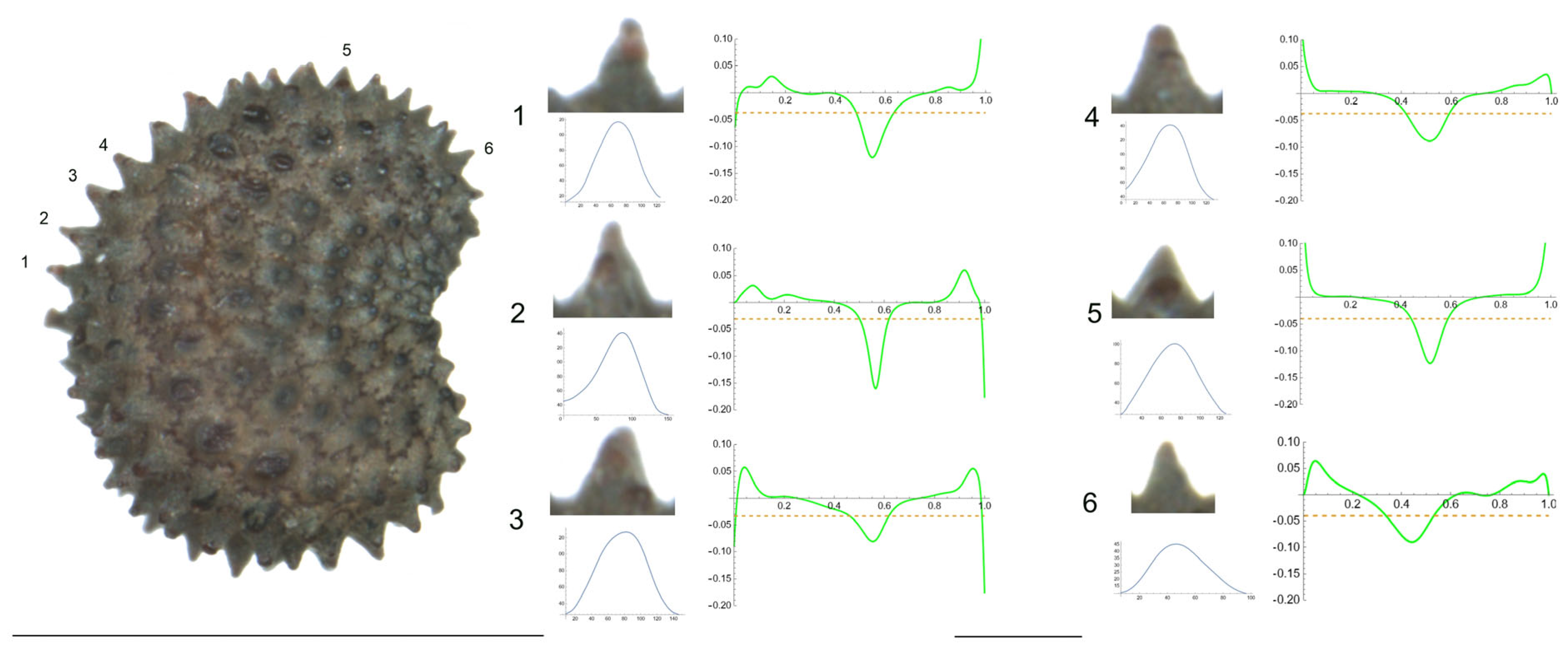
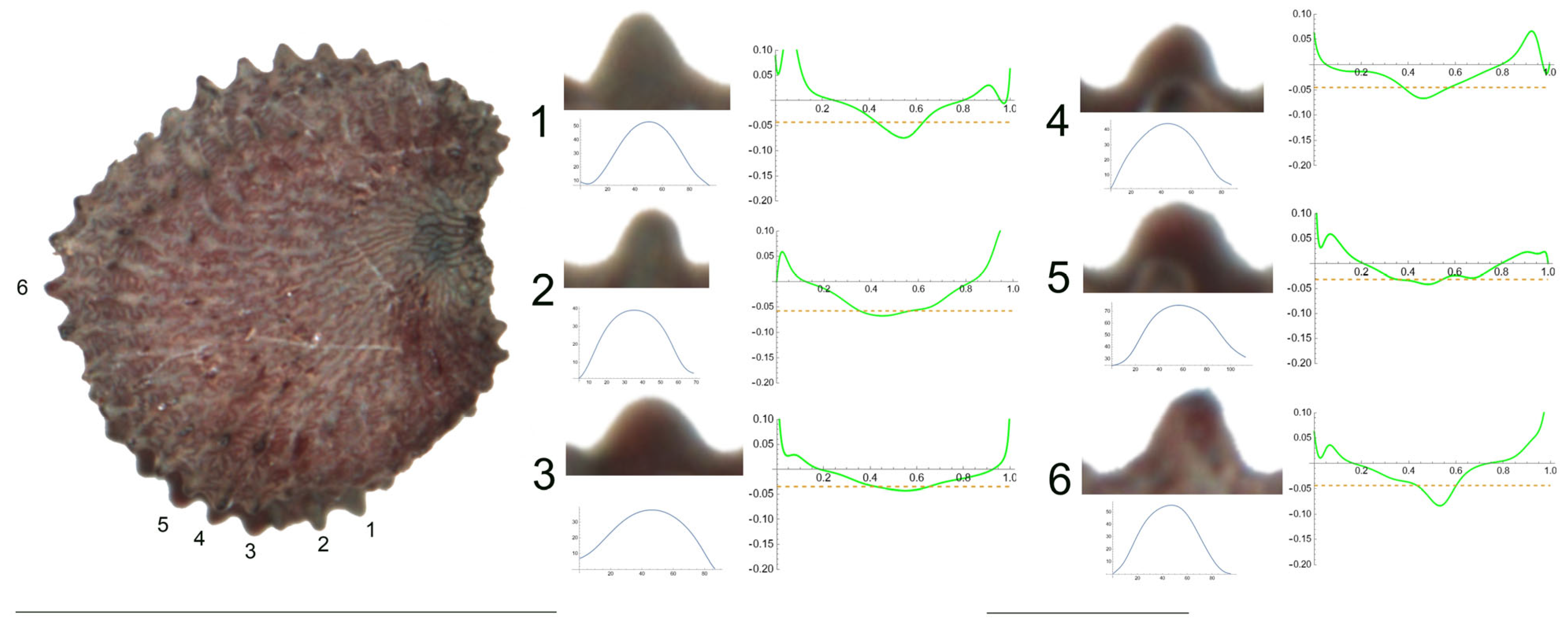
- S. aprica subsp. oldhamiana (Miq.) C.Y.Wu ex C.L.Tang.
Figure 8 presents a seed of S. aprica subsp. oldhamiana in the lateral view. The tubercles analyzed are numbered from 1 to 6. The six studied tubercles are notably triangular, with a well-defined central peak and a maximum curvature between 81 (tubercle 3) and 168 (tubercle 2) (Table 11). Although the minimum values for the studied variables were observed for different tubercles, the maximum data mostly corresponded to tubercles 2 and 5. The observed values of the ratio of maximum/mean curvature, up to 5.2, illustrated the notable difference between the maximum curvature and the mean curvature of the tubercles.
- 2.
- S. caroliniana Walter
Figure 9 shows a seed of S. caroliniana in the lateral view. The tubercles analyzed in this section are numbered from 1 to 6. For this species, the studied tubercles showed a well-visible central peak but they were remarkably different, with a gradation of their shape, from a flattened (2), to clearly acute (4,5,6) tubercles. These differences were supported by the measured variables (Table 12), since the minimum values mostly corresponded to tubercle 2, but the highest value to tubercle 4. The one exception was related to the width of the tubercles, with tubercle 1 having the maximum values (79) and tubercle 3 the minimum ones (59).
- 3.
- S. ciliata Pourr.
Figure 10 shows a seed of S. ciliata in the lateral view. The tubercles analyzed in this section are numbered from 1 to 6. The studied tubercles had a similar shape with a central peak, with the maximum curvature values between 41 (tubercle 5) and 83 (tubercle 6) (Table 13). For most of the studied variables, tubercles 3 and 5 showed the lowest values, whereas the highest ones corresponded to tubercles 2 and 6.
2.3.2. Echinate Species with Higher Maximum Curvature Values
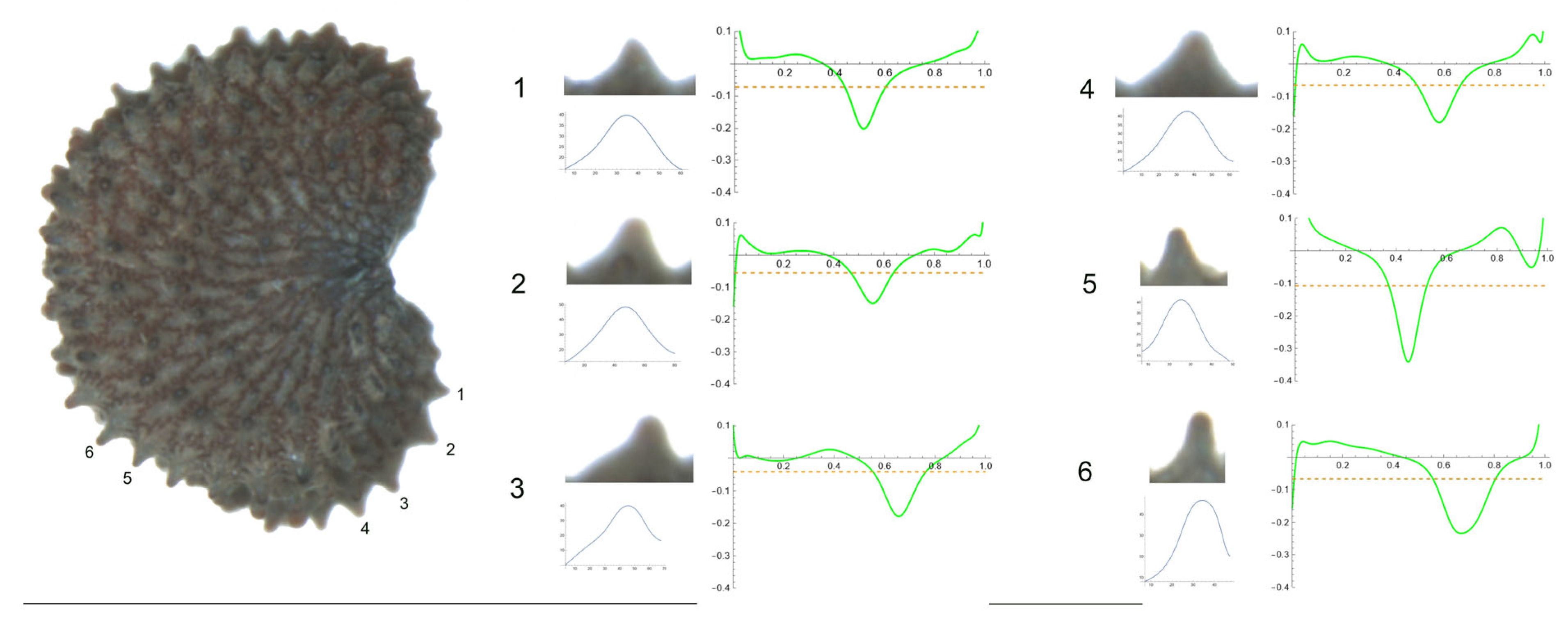
- S. yunnanensis Franch
Figure 11 contains a seed of S. yunnanensis in the lateral view. Tubercles are numbered from 1 to 6. The studied tubercles were quite similar—clearly acute with a central peak, with no noticeable differences in their shape. Tubercle 1 showed the lowest values of height and slope, and the maximum values corresponded to tubercles 2 and 3 (Table 14). The maximum curvature data were found for tubercles 5 and 6 (341.2 and 234.2, respectively), with values notably different from the remaining tubercles.
2.4. Relationship between Tubercle Type and Subgenera
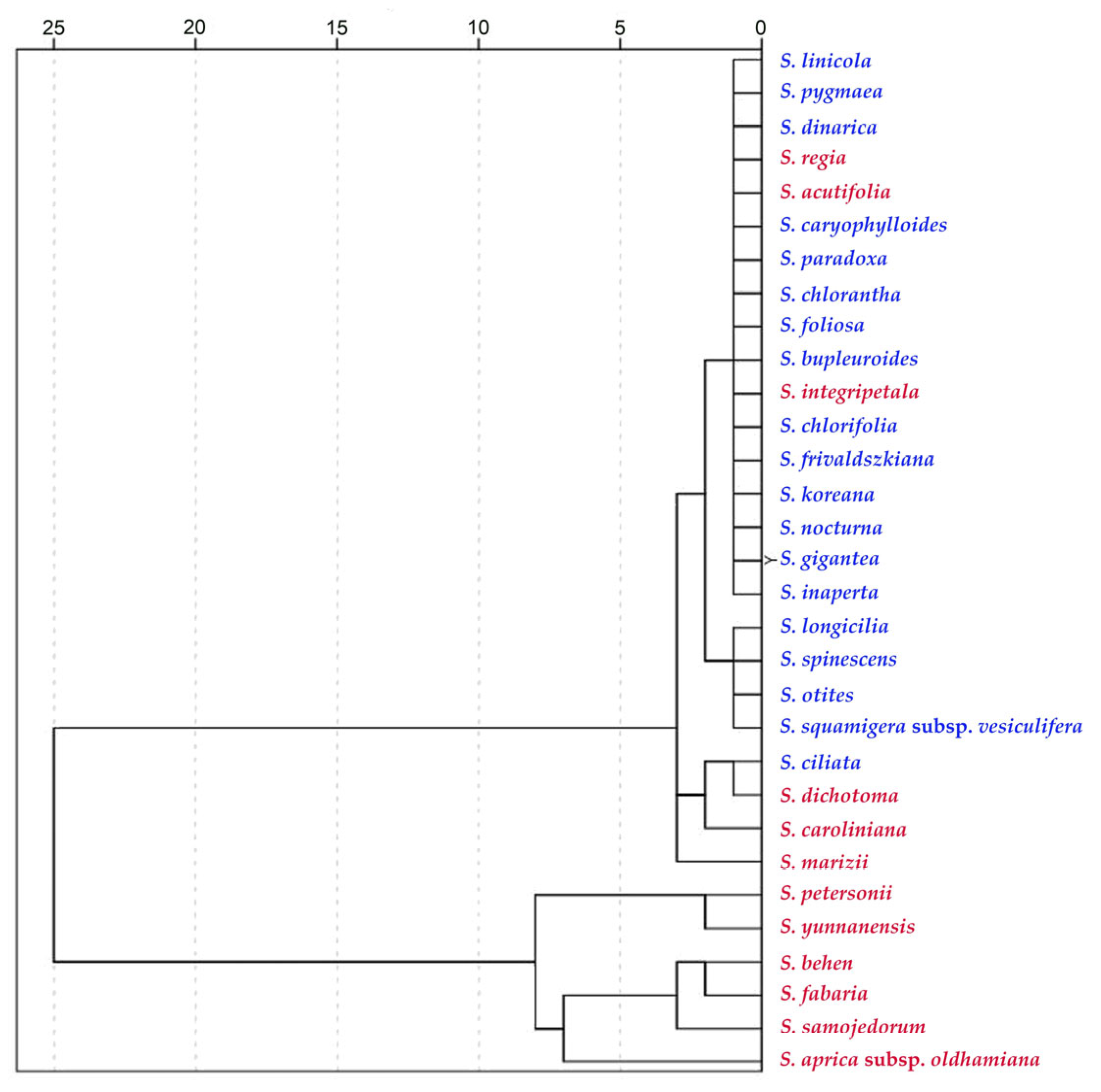
The results show a relationship between the measurements of the tubercles and the subgenera. Table 15 contains the comparison of the mean values of width, height, slope, maximum curvature, mean curvature and the ratio of maximum to mean curvature between seeds of Silene species belonging to S. subg. Behenantha and S. subg. Silene. The values were higher in subg. Behenantha for all the measurements, except tubercle width.
3. Discussion
3.1. Seed Surface Morphology in Silene Species
The analysis of Silene seed surface morphology has been traditionally based on Scanning Electron Microscopy (SEM) because this technique gives a clear definition of cell boundaries, tubercle shape, their distribution and, hence, aspects of the seed surface. Greuter [34] stated that cell shape, cell surface granularity and the presence/lack of mamilla could be used as diagnostic features for his proposed infrageneric classification of Silene. As an example, and in relation to the morphology of the tubercles, Ghazanfar [19] described them as rounded, cylindrical, clavate, umbonate or conical; Dadandi and Yildiz [21] as rounded, cylindrical, conical and mamillated. Runyeon and Prentice [33] defined the seed surface as tubercled or smooth for diverse populations of Silene vulgaris and S. uniflora, and more recently, Hosseini et al. [25] expanded seed surface morphological characterization using terms such as mammalian, echinate, tuberculate and flat. Although there are some differences in the uses of specific nomenclatures, the results may be comparable between them and with those obtained via optical imaging.
Recent studies based on optical imaging classified Silene seeds into four morphological types named smooth, rugose, echinate and papillose [35,36]. Out of 100 species analyzed, the majority were classified as rugose and echinate, with 16 species being classified as smooth and only 4 species as papillose. Most of the smooth seeds belonged to S. subg. Silene sect. Silene, and the four species with papillose seeds belonged to S. subg. Behenantha sect. Physolychnis (S. laciniata Cav. and S. magellanica (Desr.) Bocquet), to S. subg. Behenantha sect. Behenantha (S. holzmannii Heldr. ex Boiss.) and to S. subg. Silene sect. Silene (S. perlmanii W.L.Wagner, D.R.Herbst & Sohmer). The geometric characteristics of smooth and papillose seeds allow them to be distinguished from among each other and from other seed groups due to (i) the lack of tubercles in the case of smooth seeds and (ii) for papillose seeds, the tubercles or papillae showing a varied shape and uneven size, characterized by lower values of circularity and solidity [35,36].
According to previous studies based on optical seed images [35,36], rugose tubercles are typically rounded and echinates are more acute or triangular. This difference suggests that these two types of tubercles could be also characterized by quantitative geometric properties. A series of measurements presented in this work allow them to be differentiated from each other. Both types of seed tubercle may be thoroughly differentiated via curvature analysis. Curvature is the magnitude that indicates the rate of change in the slope through the points of a curve [37,38,39,40,41]. This has a constant value in a circumference, which equals the inverse of its radius with units of length−1. The maximum to mean curvature ratio gives a measure of the separation from the figure of an arc of circumference. Both maximum and mean curvature values and maximum to mean curvature ratios were significant higher for echinate tubercles than for rugose ones. Tubercle curvature, as with any other tubercle measurement, is a geometric feature of tubercles, not of seeds. These curvature values could adequately characterize the geometric differences between them. In addition, the combination of a smaller width, with a higher height and notably higher values of slope might also support the definition of echinate tubercles in contrast to rugose.
Silene cell surface projections may vary infraspecifically (e.g., S. viscosa Pers., see [25]). In addition, it may be possible to find diverse types of tubercles within the same seed. Our data demonstrated that the presence of two types of tubercles was relatively frequent, as it was observed for S. spinescens and S. gigantea. Rugose tubercles were well characterized by the highest values of width and the lowest values for the remaining measurements in comparison to echinate tubercles. Therefore, the geometric variables would be very useful in distinguishing and characterizing the morphology of tubercles from quantitative data, as opposed to only via qualitative description. The presence of additional excrescences on the top of rugose tubercles, as it was observed for S. squamigera subsp. vesiculifera, can change the typical values of the curvature variables of a rugose tubercle.
The physiological function of the tubercles and their adaptive meaning is still a matter of debate, and an association between tubercle types and particular environmental factors has not been conclusively proven. The tubercles may be involved in energy homeostasis and as mechanical factors facilitating seed transport [42]. Increased tubercle size means a higher surface area to volume ratio with the consequent increase in energy and gas exchange [43]. Additionally, tubercles may have a role for anchoring seeds in the rocky soil of cliffs [44,45,46].
There is no clear adaptive benefit of tubercle shape in relation to geographic localization or climatic conditions. Based on the observation of 167 collections of wild seeds and 97 sets of mature greenhouse-grown progeny, Prentice analyzed the variation in tubercle shape for S. latifolia and S. dioica, concluding that seeds with conical tubercles have a more eastern localization than those with rounded tubercles [47]. In another study with herbarium material, the analysis of 216 seeds from diverse origins [48] allowed the researchers to conclude that while seeds from cold-winter regions have tall tubercles, seeds from warm-winter regions possess, in general, shorter tubercles.
Concerning the relationship between tubercle type and taxonomic subgenera, our results show an association of the rugose tubercle type with subg. Silene and the echinate type with Behenantha. Although in many instances, morphological characteristics appear independently in separate lineages over the course of evolution (homoplasy) [19,21,25,27,36], the quantitative analyses presented here support the predominance of the rugose tubercle type, characterized by lower curvature values, in the species of S. subg. Silene and the echinate type, of higher curvature values, in S. subg. Behenantha. The analysis may be applied to other species and populations to confirm whether this possibility has a general application.
3.2. Seed Surface Morphology in the Caryophyllaceae
Several articles have been dedicated to seed micromorphology in species other than Silene in the Caryophyllaceae [5,6,7,8,9,10,11,12,13,14,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64]. In some instances, tubercle shape is associated with a given phylogenetic group or a particular geographic location [6,10,49,50,51]. The seeds of Moheringia L. species with long tubercles (papillate) have been described in warm-winter regions in the Eastern Balkans and the Iberian Peninsula [10]. In Schiedea Cham. & Schltdl., papillate margins are found in species occurring in ecologically extreme environments, such as wet forests of the higher elevations (S. trinervis) or dry slopes (S. adamantis) [50].
Finally, micromorphological characters have been recently considered as possible taxonomic criteria in other families of the order, Caryophyllales, such as the Aizoaceae and the Molluginaceae [65,66]. The seeds of Portulacca, in the Portulacaceae, have tubercles resembling those of Silene [67,68], which can be accurately measured using the methods described here.
There is a long-standing controversy about the applicability of tubercles in taxonomy that may benefit from the quantitative methods described here.
4. Materials and Methods
4.1. Silene Seeds
The populations of seeds analyzed in this work are listed in Table 16. Representative seed images from populations labelled JBUV can be observed and downloaded at https://zenodo.org/record/7330942#.Y3 Y8 Hn3 MJD8 (accessed on 26 September 2023). Seed images of the populations of S. acutifolia and S. otites are available at https://zenodo.org/record/4035649 (accessed on 26 September 2023). Seed images for the remaining populations (S. behen, S. caroliniana, S. inaperta and S. nocturna) are available under request.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 16.
List of seed populations analyzed in this work. The populations labelled JBUV were obtained from the carpoespermateca at the Botanical Garden of the University of Valencia and originate from an exchange protocol between seed collections through the world. The name of the subgenus and the corresponding section (between brackets) are indicated. “u.” means unknown place of origin. The ascription of each species to subgenera and sections is taken from [69].
Table 16.
List of seed populations analyzed in this work. The populations labelled JBUV were obtained from the carpoespermateca at the Botanical Garden of the University of Valencia and originate from an exchange protocol between seed collections through the world. The name of the subgenus and the corresponding section (between brackets) are indicated. “u.” means unknown place of origin. The ascription of each species to subgenera and sections is taken from [69].
| Species | Type (Rugose or Echinate) | Received from | Registered by (Place of Origin) | Subgenus (Section) |
|---|---|---|---|---|
| Silene acutifolia (L.) Link ex Rohrb. | R | IBP collection Acad Sci CzR, Brno | IBP collection (u.) | Behenantha (Acutifoliae) |
| Silene aprica subsp. oldhamiana (Miq.) C.Y.Wu ex C.L.Tang | E | JBUV333 | Chollipo Arboretum BG, South Korea (u.) | Behenantha(Physolychnis) |
| S. behen L. | E | IBP collection Acad Sci CzR, Brno | IBP collection (u.) | Behenantha(Behenantha) |
| S. bupleuroides L. | R | JBUVC2787 | u. Hortus Botanicus Vacratot, Hungary (u.) | Silene(Sclerocalycinae) |
| S. caroliniana Walter | E | IBP collection Acad Sci CzR, Brno | IBP collection (u.) | Behenantha (Physolychnis) |
| S. caryophylloides (Poir.) Otth | R | JBUV950 | Botanischer Garten der Universität Tübingen (Holubec, Turkey, Ulu Dag) | Silene (Auriculatae) |
| S. chlorantha (Willd.) Ehrh. | R | JBUV1028 | Botanischer Garten der Universität Potsdam Germany. (Brandenburg, Odergebiet, an der Bahn, SW Bahnhof Podelzig, Germany) | Silene (Siphonomorpha) |
| S. chlorifolia Sm. | R | JBUV1000 | BG der Martin-Luther-Univ. Halle-Wittenberg, Germany (u.) | Silene (Sclerocalycinae) |
| S. ciliata Pourr. | E | JBUV219 | Botany Hung. Acad. of Sciences (Botanical Garden) | Silene (Silene) |
| S. dichotoma Ehrh. | E | JBUV668 | Botanic Garden of the University of Copenhagen, Denmark (u.) | Behenantha (Dichotomae) |
| S. dinarica Spreng. | R | JBUV1600 | Botanicka Zahrada Teplice, Czech Republic (u.) | Silene (Siphonomorpha) |
| S. fabaria (L.) Coyte | E | JBUV3188 | Botanischer Garten der Universität Bonn, Germany (Chakidiki, south of Ouranopolis, towards the border of Athos, Greece) | Behenantha (Behenantha) |
| S. foliosa Maxim. | R | JBUV2177 | Vladivostok Botanical Garden, Russia (Gamow Peninsula) | Silene (Siphonomorpha) |
| S. frivaldszkyana Hampe | R | JBUV2881 | Siberian Botanical Garden of Tomsk State University, Russia (u.) | Silene (Siphonomorpha) |
| S. gigantea (L.) L. | R | JBUV1494 | Julia & Alexander N. Diomides Botanic Garden, Athens, Greece (Cult./ATHD) | Silene (Siphonomorpha) |
| S. inaperta L. | R | AJ270 | Collection of Ana Juan (Rambla Bateig, Elda, Alicante, Spain) | Silene (Muscipula) |
| S. inaperta L. | R | AJ335 | Collection of Ana Juan (Rambla de Caprala, Petrer, Alicante, Spain) | Silene (Muscipula) |
| S. integripetala Bory and Chaub. | R | JBUV1001 | BG der Martin-Luther-Univ. Halle-Wittenberg, Germany | Behenantha (Sedoides) |
| S. koreana Kom. | R | JBUV1917 | Botanic Garden of Perm State University, Russia | Silene (Siphonomorpha) |
| S. linicola C.C. Gmel. | R | JBUV1056 | Jardin Botanique de la ville de Lyon Aube (10), entre Auxerre et Troyes, France. | Silene (Lasiocalycinae) |
| S. longicilia (Brot.) Otth | E | JBUV712 | BG Universidade de Coimbra (Serra da Boa Viagem—Figueira da Foz, Portugal) | Silene (Siphonomorpha) |
| S. marizii Samp. | R | JBUV713 | BG Universidade de Coimbra (Nespereira—Celorico da Beira, Portugal) | Behenantha (Melandrium) |
| S. nocturna L. | R | AJ316 | Collection of Ana Juan (Sierra de Irta, Peñíscola, Castellón, Spain) | Silene (Silene) |
| S. otites (L.) Wibel | R | (IBP collection, Brno) | IBP collection (u.) | Silene (Siphonomorpha) |
| S. paradoxa L. | R | JBUV1142 | Station Alpine du Lautaret. Univ. Joseph Fourier Défilé d’Inzecca (Corse, France, 250 m) | Silene (Siphonomorpha) |
| S. petersonii Maguire | E | JBUV1002 | BG der Martin-Luther-univ. Halle-Wittenberg, Germany (Mount Brocken Garden) | Behenantha (Physolychnis) |
| S. pygmaea Adams | R | JBUV964 | St Andrews Botanical Garden, Scotland (u.) | Silene (Auriculatae) |
| S. regia Sims | R | JBUV2630 | Botanischer Garten Universität Hamburg Nachzucht BG Hamburg, Germany (Winona, MN/US; Prairie Moon Nursery, USA) | Behenantha (Physolychnis) |
| S. samojedorum (Sambuk) Oxelman | E | JBUV1069 | Hortus Botanicus Patavinus, Italy | Behenantha (Physolychnis) |
| S. spinescens Sm. | R | JBUV1496 | Julia & Alexander N. Diomides Botanic Garden (Spont./Sounion National Park-Attiki, Athens, Greece) | Silene (Siphonomorpha) |
| S. squamigera Boiss. subsp. vesiculifera (J.Gay ex Boiss.) Coode and Cullen | R | JBUV671 | Botanic Garden of the University of Copenhagen, Danmark | Silene (Lasiocalycinae) |
| S. yunnanensis Franch. | E | JBUV2050 | Botanischer Garten der Universität Zürich, Germany | Behenantha (Cucubaloides) |
4.2. Seed Images
For the analysis of individual tubercles, photographs were taken with a Nikon Stereomicroscope Model SMZ1500 (Nikon, Tokio, Japan), equipped with a camera Nikon DS-Fi1 of 5.24 megapixels (Nikon, Tokio, Japan). Photographs were stored as JPG images of 2560 × 1920 with 300 ppp.
4.3. Tubercle Measurements
Tubercle measurements and curvature analyses were applied to seeds of 31 species of Silene. Most of them were obtained from the carpoespermateca at the Botanical Garden of the University of Valencia by an exchange protocol between seed collections through the world. Measurements of width, height, slope and curvature (maximum and average curvature values) were taken for a total of 396 tubercles (268 corresponding to rugose seeds and 128 to echinate seeds). The number of seeds was between 1 and 3 for each species, as indicated in Table 2 and Table 10 and Figure A1 in Appendix A.
4.3.1. Width, Height and Slope Measurements in the Tubercles
Seed images containing a ruler of 1 mm were opened in Image J. Two direct measurements were made for each of the tubercles indicated in Figure 4: tubercle width at the base (W) and tubercle height (H). The slope of each tubercle (S) was calculated as
4.3.2. Curvature Measurements
Maximum absolute values and average curvatures were determined for individual tubercles of each species according to established procedures [37,38,39,40,41] (See Supplementary Materials). In the measurements of curvature for individual tubercles, the points were taken automatically with the function, Analyze line graph, of Image J. Between 6 and 22 tubercles of representative seeds (see Figure A1 in Appendix A) were analyzed for each species. Individual images (JPEG) were kept of each tubercle and vertically oriented. Images were opened in Image J and converted to 8-bit, threshold values were adjusted, and the image was analyzed. The curve corresponding to the tubercle was selected from the outlines, and a new threshold was defined before the corresponding line graph was analyzed, to obtain the x,y coordinates. The coordinates were the basis of obtaining the Bézier curve and the corresponding curvature values according to published protocols [37,38,39,40,41]. Curvature was given in micron−1 × (1000); thus, a curvature of 20 corresponds to a circumference of 50 microns (1/50 × 1000) and a curvature of 100 to a to a circumference of 10 microns.
4.4. Statistical Analysis
ANOVA was used to show significant differences between populations for the measured variables. In the case of the comparison of morphological characteristics involving more than two populations, ANOVA was followed by a Tukey test to obtain specific information on which means were significantly different from one another. Statistical analyses (ANOVA) were conducted with IBM SPSS statistics v28 (SPSS 2021).
5. Conclusions
A quantitative analysis of tubercles has been made here of seeds of Silene, belonging to 31 species that had previously been classified as rugose (21 species) or echinate (10 species). Of these, 19 belonged to S. subg. Silene and 12 to Behenantha. The analysis was based on tubercle dimensions and curvature. Width at the base, height and slope, together with maximum and average curvature values were obtained for 396 tubercles, in a total of 49 seeds corresponding to 21 species. Rugose tubercles had higher values of tubercle width and lower values for the other measurements, in comparison to echinate-type tubercles. The seeds of the rugose type had lower values of curvature. Additionally, lower values of curvature were found in species of S. subg. Silene in comparison with S. subg. Behenantha. The protocols applied in this article may be helpful for detailed morphological analyses, determining not only differences of tubercle type (rounded or rugose versus conical or echinate) but also more subtle quantitative changes in tubercle morphology.
Supplementary Materials
The protocol to measure curvature in seed tubercles is available at https://zenodo.org/record/8162247 (accessed on 26 September 2023). The Mathematica codes for the curvature analysis presented are available at https://zenodo.org/record/8269393 (accessed on 26 September 2023).
Author Contributions
Conceptualization, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; methodology, J.L.R.-L., Á.T. and E.C.; software, J.L.R.-L., J.J.M.-G., Á.T. and E.C.; validation, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; formal analysis, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; investigation, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; resources, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; data curation, J.L.R.-L., J.J.M.-G. and E.C.; writing—original draft preparation, E.C.; writing—review and editing J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; visualization, J.L.R.-L., J.J.M.-G., A.J., Á.T. and E.C.; supervision, E.C., J.L.R.-L. and Á.T.; project administration, E.C., J.L.R.-L. and Á.T.; funding acquisition, E.C., J.L.R.-L. and Á.T. All authors have read and agreed to the published version of the manuscript.
Funding
Project “CLU-2019-05-IRNASA/CSIC Unit of Excellence”, funded by the Junta de Castilla y León and co-financed by the European Union (ERDF “Europe drives our growth”).
Data Availability Statement
Data are contained within the article or Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Tubercles analyzed in this work. Measurements of width, height, slope and curvature (maximum and average curvature values) were made for a total of 396 tubercles in a total of 49 seeds corresponding to 21 species. From the total of tubercles measured, 268 corresponded to rugose seeds and 128 to echinate seeds.
Figure A1.
Tubercles analyzed in this work. Measurements of width, height, slope and curvature (maximum and average curvature values) were made for a total of 396 tubercles in a total of 49 seeds corresponding to 21 species. From the total of tubercles measured, 268 corresponded to rugose seeds and 128 to echinate seeds.


References
- Johri, B.M.; Ambegaokar, K.B.; Srivastava, P.S. Comparative Embryology of Angiosperms; Springer: Heidelberg, Germany, 1992; Volume 1. [Google Scholar]
- Martin, A.C. The comparative internal morphology of seeds. Am. Midl. Nat. 1946, 36, 513–660. [Google Scholar] [CrossRef]
- Greenberg, A.K.; Donoghue, M.J. Molecular systematics and character evolution in Caryophyllaceae. Taxon 2011, 60, 1637–1652. [Google Scholar] [CrossRef]
- Sukhorukov, A.P.; Mavrodiev, E.V.; Struwig, M.; Nilova, M.V.; Dzhalilova, K.K.; Balandin, S.A.; Erst, A.; Krinitsyna, A.A. One-Seeded fruits in the core Caryophyllales: Their origin and structural diversity. PLoS ONE 2015, 10, e0117974. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Papini, A.; Shah, S.N.; Zaman, W.; Sohail, A.; Iqbal, M. Seed micromorphology and its taxonomic evidence in subfamily Alsinoideae (Caryophyllaceae). Microsc. Res. Tech. 2019, 82, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Wofford, B.E. External seed morphology of Arenaria (Caryophyllaceae) of the Southeastern United States. Syst. Bot. 1981, 6, 126. [Google Scholar] [CrossRef]
- Wyatt, R. Intraspecific variation in seed morphology of Arenaria uniflora (Caryophyllaceae). Syst. Bot. 1984, 9, 423. [Google Scholar] [CrossRef]
- Amini, E.; Zarre, S.; Assadi, M. Seed micro-morphology and its systematic significance in Gypsophila (Caryophyllaceae) and allied genera. Nord. J. Bot. 2011, 29, 660–669. [Google Scholar] [CrossRef]
- Kovtonyuk, N.K. The structure of seed surface and the systematics of the Siberian Gypsophila species (Caryophyllaceae). Bot Zhurn 1994, 79, 48–51. [Google Scholar]
- Minuto, L.; Fior, S.; Roccotiello, E.; Casazza, G. Seed morphology in Moehringia L. and its taxonomic significance in comparative studies within the Caryophyllaceae. Plant Syst. Evol. 2006, 262, 189–208. [Google Scholar] [CrossRef]
- Kaplan, A.; Çölgeçen, H.; Büyükkartal, H.N. Seed morphology and histology of some Paronychia taxa (Caryophyllaceae) from Turkey. Bangladesh J. Bot. 2009, 38, 171–176. [Google Scholar] [CrossRef]
- Crow, G.E. The systematic significance of seed morphology in Sagina (Caryophyllaceae) Under Scanning Electron Microscopy. Brittonia 1979, 31, 52–63. [Google Scholar] [CrossRef]
- Volponi, C.R. Stellaria cuspidata (Caryophyllaceae) and some related species in the Andes. Willdenowia 1993, 23, 193–209. [Google Scholar]
- Poyraz, I.E.; Ataşlar, E. Pollen and seed morphology of Velezia L. (Caryophyllaceae) genus in Turkey. Turk. J. Bot. 2010, 34, 179–190. [Google Scholar] [CrossRef]
- Hong, S.-P.; Han, M.-J.; Kim, K.-J. Systematic significance of seed coat morphology in Silene L. s. str. (Sileneae-Caryophyllaceae) from Korea. J. Plant Biol. 1999, 42, 146–150. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rewicz, A.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. Seed morphology in Silene based on geometric models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef] [PubMed]
- Juan, A.; Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. New techniques for seed shape description in Silene species. Taxonomy 2022, 2, 1–19. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Juan, A.; Cervantes, E. Comparison of seed images with geometric models, an approach to the morphology of Silene (Caryophyllaceae). Taxonomy 2023, 3, 109–132. [Google Scholar] [CrossRef]
- Ghazanfar, S.A. Seed characters as diagnostic in the perennial sections of the genus Silene (Family Caryophyllaceae). Pak. J. Bot. 1983, 15, 7–12. [Google Scholar]
- Tabaripur, R.; Koohdar, F.; Sheidai, M.; Gholipour, A. Intra-specific variations in Silene: Morphometry and micromorphometry analysis. Afr. J. Biotech. 2013, 12, 5208–5217. [Google Scholar]
- Dadandi, M.Y.; Yildiz, K. Seed morphology of some Silene L. (Caryophyllaceae) species collected from Turkey. Turk. J. Bot. 2015, 39, 280–297. [Google Scholar]
- Kuh, M.; Yildiz, K.; Minareci, E. A taxonomic study of the Silene sections Behenantha and Dichotomae (Caryophyllaceae) in Turkey based on the micromorphology of their seed and pollen. Turk. J. Bot. 2017, 41, 493–504. [Google Scholar] [CrossRef]
- Ocaña, M.A.; Juan, R.; Fernàndez, I.; Pastor, J. Estudio morfológico de semillas de Silene (Caryophyllaceae) del Suroeste de España. Lagascalia 2011, 31, 21–45. [Google Scholar]
- Arman, M.; Gholipour, A. Seed morphology diversity in some Iranian endemic Silene (Caryophyllaceae) species and their taxonomic significance. Acta Biol. Szeged. 2013, 57, 31–37. [Google Scholar]
- Hoseini, E.; Ghahremaninejad, F.; Assadi, M.; Edalatiyan, M.N. Seed micromorphology and its implication in subgeneric classification of Silene (Caryophyllaceae, Sileneae). Flora 2017, 228, 31–38. [Google Scholar] [CrossRef]
- Keshavarzi, M.; Mahdavinejad, M.; Sheidai, M.; Gholipour, A. Seed and pollen morphology of some Silene species (Caryophyllaceae) in Iran. Phytol. Balc. 2015, 21, 7–12. [Google Scholar]
- Fawzi, N.M.; Fawzi, A.M.; Mohamed, A.A.-H.A. Seed morphological studies on some species of Silene L. (Caryophyllaceae). Int. J. Bot. 2010, 6, 287–292. [Google Scholar] [CrossRef]
- El-Oqlah, A.A.; Karim, F.M. Morphological and anatomical studies of seed coat in Silene species (Caryophyllaceae) from Jordan. Arab. Gulf. J. Sci. Res. 1990, 8, 121–139. [Google Scholar]
- Perveen, A. Seed morphology of the genus Silene: Caryophyllaceae from Pakistan and Kashmir. Int. J. Biol. Biotechnol. 2009, 6, 219–227. [Google Scholar]
- Özbek, M.U.; Uzunhisarcikli, M.E. A new species of Silene (Caryophyllaceae) from Turkey. Phytotaxa 2019, 397, 74–82. [Google Scholar] [CrossRef]
- Atazadeh, N.; Keshavarzi, M.; Sheidai, M.; Gholipour, A. Seed morphology of Silene commelinifolia Boiss. Complex (Caryophyllaceae Juss.). Mod. Phytomorphol. 2017, 11, 5–13. [Google Scholar]
- Firat, M.; Yildiz, K. Silene konuralpii (Sect. Spergulifoliae, Caryophyllaceae), a new species from eastern Anatolia. Phytotaxa 2016, 288, 214–226. [Google Scholar]
- Runyeon, H.; Prentice, H.C. Patterns of seed polymorphism and allozyme variation in the bladder campions, Silene vulgaris and Silene uniflora (Caryophyllaceae). Can. J. Bot. 1977, 75, 1868–1886. [Google Scholar] [CrossRef]
- Greuter, W. Silene (Caryophyllaceae) in Greece: A subgeneric and sectional classification. Taxon 1995, 44, 543–581. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Tocino, Á.; Janoušek, B.; Juan, A.; Cervantes, E. The outline of seed silhouettes: A morphological approach to Silene (Caryophyllaceae). Plants 2022, 11, 3383. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Juan, A.; Tocino, Á.; Janousek, B.; Cervantes, E. Seed morphological properties related to taxonomy in Silene L. Species. Taxonomy 2022, 2, 298–323. [Google Scholar] [CrossRef]
- Cervantes, E.; Tocino, A. Geometric analysis of Arabidopsis root apex reveals a new aspect of the ethylene signal transduction pathway in development. J. Plant Physiol. 2005, 162, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Noriega, A.; Tocino, A.; Cervantes, E. Hydrogen peroxide treatment results in reduced curvature values in the Arabidopsis root apex. J. Plant Physiol. 2009, 166, 554–558. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rewicz, A.; Goriewa-Duba, K.; Wiwart, M.; Tocino, Á.; Cervantes, E. Morphological description and classification of wheat kernels based on geometric models. Agronomy 2019, 9, 399. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello Sáenz de Santamaría, F. Seed apex curvature in key Spanish grapevine cultivars. Vitic. Data J. 2021, 3, e66478. [Google Scholar] [CrossRef]
- Cervantes, E.; Rodríguez-Lorenzo, J.L.; Martín-Gómez, J.J.; Tocino, Á. Curvature analysis of seed silhouettes in Silene L. Plants 2023, 12, 2439. [Google Scholar] [CrossRef]
- Bittrich, V. “Caryophyllaceae” Flowering Plants Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families; Springer: Berlin/Heidelberg, Germany, 1993; pp. 206–236. [Google Scholar]
- Harper, J.L.; Lovell, P.H.; Moore, K.G. The shapes and sizes of seeds. Annu. Rev. Ecol. Evol. Syst. 1970, 1, 327–356. [Google Scholar] [CrossRef]
- Bochet, E.; García-Fayos, P. Factors controlling vegetation establishment and water erosion on motorway slopes in Valencia, Spain. Restor Ecol. 2004, 12, 166–174. [Google Scholar] [CrossRef]
- Cerdà, A.; Garcıa-Fayos, P. The influence of seed size and shape on their removal by water erosion. Catena 2002, 48, 293–301. [Google Scholar] [CrossRef]
- Thompson, K.B.; Band, S.R.; Hodgson, J.G. Seed size and shape predict persistence in soil. Funct. Ecol. 1993, 48, 236–241. [Google Scholar] [CrossRef]
- Prentice, H. Numerical analysis of infraspecific variation in European Silene alba and S. dioica (Caryophyllaceae). Bot. J. Linn. Soc. 1979, 78, 181–212. [Google Scholar] [CrossRef]
- Prentice, H. Climate and clinal variation in seed morphology of the white campion, Silene latifolia (Caryophyllaceae). Bot. J. Linn. Soc. 1986, 27, 179–189. [Google Scholar] [CrossRef]
- Wagner, L.W.; Weller, S.G.; Sakai, A.K. Monograph of Schiedea (Caryophyllaceae-Alsinoideae). Syst. Bot. Monogr. 2005, 72, 1–169. [Google Scholar]
- Frajman, B.; Oxelman, B. Reticulate phylogenetics and phytogeographical structure of Heliosperma (Sileneae, Caryophyllaceae) inferred from chloroplast and nuclear DNA sequences. Mol. Phylogenet. Evol. 2007, 43, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Sadeghian, S.; Zarre, S.; Heubl, G. Systematic implication of seed micromorphology in Arenaria (Caryophyllaceae) and allied genera. Flora 2014, 209, 513–529. [Google Scholar] [CrossRef]
- Arabi, Z.; Ghahremaninejad, F.; Rabeler, R.K.; Heubl, G.; Zarre, S. Seed micromorphology and its systematic significance in tribe Alsineae (Caryophyllaceae). Flora 2017, 234, 41–59. [Google Scholar] [CrossRef]
- Mostafavi, G.; Assadi, M.; Nejadsattari, T.; Sharifnia, F.; Mehregan, I. Seed micromorphological survey of the Minuartia species (Caryophyllaceae) in Iran. Turk. J. Bot. 2013, 37, 446–454. [Google Scholar] [CrossRef]
- Abdel-Maksoud, H.S.; Fawzi, N.M. Seed Morphology of Some Taxa of Caryophyllaceae. Bull. Fac. Agric. Cairo Univ. 2016, 67, 213–227. [Google Scholar] [CrossRef]
- Minuto, L.; Roccotiello, E.; Casazza, G. New seed morphological features in Moehringia L. (Caryophyllaceae) and their taxonomic and ecological significance. Plant Biosyst. 2011, 145, 60–67. [Google Scholar] [CrossRef]
- Mahdavi, M.; Asadi, M.; Fallahian, F.; Nejadsattari, T. The systematic significance of seed micro-morphology in Stellaria L. (Caryophyllaceae) and its closest relatives in Iran. Iran. J. Bot. 2012, 18, 302–310. [Google Scholar]
- Volponi, C.R. Stellaria yungasensis (Caryophyllaceae), una nueva cita para Argentina. An. Soc. Cient. Argent. 1992, 222, 69–72. [Google Scholar]
- Volponi, C.R. Contribución a la espermatología de especies argentinas de Stellaria (Caryophyllaceae). Bol. Soc. Argent. Bot. 1986, 24, 283–294. [Google Scholar]
- Pirani, A.; Zarre, S.; Rabeler, R.; Assadi, M.; Reza, M.; Joharchi, M.R.; Oxelman, B. Systematic significance of seed morphology in Acanthophyllum (Caryophyllaceae: Tribe Caryophylleae) in Iran. Phytotaxa 2019, 387, 105–118. [Google Scholar] [CrossRef]
- Bülbül, A.S.; Armaðan, M.; Varlik, K. Seed micromorphology of Acanthophyllum C.A.Mey. (Caryophyllaceae) genus in Turkey. Kastamonu Univ. J. For. Fac. 2017, 17, 215–224. [Google Scholar]
- Ataşlar, E.; Ocak, A. Gypsophila osmangaziensis (Caryophyllaceae), a new species from Central Anatolia, Turkey. Ann. Bot. Fenn. 2005, 331, 84–92. [Google Scholar]
- Deniz, I.G.; Aykurt, C.; Genç, I.; Aksoy, A. A new species of Dianthus (Caryophyllaceae) from Antalya, South Anatolia, Turkey. PhytoKeys 2016, 63, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ocaña Amante, M.E.; Fernández González, I.; Pastor Díaz, J.E. Fruit and seed morphology in Paronychia Miller from South-west Spain. Lascalia 1997, 19, 521–528. [Google Scholar]
- Hosny, A.I.; Zareh, M.M. Taxonomic studies of Silenoideae (Caryophyllaceae) in Egypt. 2. Taxonomic significance of seed characters in genus Silene L. with special reference to the seed coat sculpture. Taeckholmia 1993, 14, 37–44. [Google Scholar]
- Sukhorukov, A.P.; Nilova, M.V.; Kushunina, M.; Mazei, Y.; Klak, C. Evolution of seed characters and of dispersal modes in Aizoaceae. Front. Plant Sci. 2023, 14, 1140069. [Google Scholar] [CrossRef] [PubMed]
- Sukhorukov, A.P.; Zhang, M.-L.; Kushunina, M.; Nilova, M.V.; Krinitsina, A.; Zaika, M.A.; Mazei, Y. Seed characters in Molluginaceae (Caryophyllales): Implications for taxonomy and evolution. Bot. J. Linn. Soc. 2018, 187, 167–208. [Google Scholar] [CrossRef]
- Danin, A.; Domina, G.; Raimondo, F.M. Microspecies of the Portulaca oleracea aggregate found on major Mediterranean islands (Sicily, Cyprus, Crete, Rhodes). Fl. Medit. 2008, 18, 89–107. [Google Scholar]
- Ocampo, G.; Travis Columbus, J. Molecular phylogenetics, historical biogeography, and chromosome number evolution of Portulaca (Portulacaceae). Mol. Phylogenet. Evol. 2012, 63, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabeler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
Figure 1.
Left: lateral view of a seed of S. bupleuroides with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side, below the tubercle images, represents 100 μ.
Figure 1.
Left: lateral view of a seed of S. bupleuroides with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side, below the tubercle images, represents 100 μ.

Figure 2.
Left: lateral view of a seed of S. chlorantha with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. Bar on the right side, below the tubercle images, represents 100 μ.
Figure 2.
Left: lateral view of a seed of S. chlorantha with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. Bar on the right side, below the tubercle images, represents 100 μ.

Figure 3.
Left: lateral view of a seed of S. frivaldskiana with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. Bar on the right side, below the tubercle and curvature images, represents 100 μ.
Figure 3.
Left: lateral view of a seed of S. frivaldskiana with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. Bar on the right side, below the tubercle and curvature images, represents 100 μ.

Figure 4.
Left: lateral view of a seed of S. integripetala with the tubercles analyzed numbered 1 to 6. Bar represents 1 mm. Right: individual view of the analyzed tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. The bar below the tubercle and curvature images equals 100 μ.
Figure 4.
Left: lateral view of a seed of S. integripetala with the tubercles analyzed numbered 1 to 6. Bar represents 1 mm. Right: individual view of the analyzed tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents mean curvature. The bar below the tubercle and curvature images equals 100 μ.

Figure 5.
Left: Lateral view of a seed of S. spinescens with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the analyzed tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 5.
Left: Lateral view of a seed of S. spinescens with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the analyzed tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 6.
Left: lateral view of a seed of S. squamigera subsp. vesiculifera with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images, represents 100 μ.
Figure 6.
Left: lateral view of a seed of S. squamigera subsp. vesiculifera with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images, represents 100 μ.

Figure 7.
Left: lateral view of a seed of S. gigantea with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 7.
Left: lateral view of a seed of S. gigantea with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 8.
Left: lateral view of a seed of S. aprica with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 8.
Left: lateral view of a seed of S. aprica with the tubercles object of analysis numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 9.
Left: lateral view of a seed of S. caroliniana with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 9.
Left: lateral view of a seed of S. caroliniana with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 10.
Left: lateral view of a seed of S. ciliata with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 10.
Left: lateral view of a seed of S. ciliata with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 11.
Left: lateral view of a seed of S. yunnanensis with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.
Figure 11.
Left: lateral view of a seed of S. yunnanensis with the tubercles analyzed numbered 1 to 6. The bar on the left represents 1 mm. Right: individual view of the tubercles, Bézier curves (below each tubercle) and curvature plots representing the variation in curvature along the Bézier curve. The discontinuous line in the curvature plot represents the value of mean curvature. Bar on the right side below the tubercle images represents 100 μ.

Figure 12.
Dendrogram based on hierarchical clustering with the values of six measurements indicated in Table 15. Species of subg. Behenantha are concentrated in the lower part of the diagram, corresponding with the higher values of curvature, while species of subg. Silene are grouped in the upper branches. Results are shown for the 31 species within Table 16 (12 belonging to S. subg. Behenantha in red and 19 belonging to S. subg. Silene in blue).
Figure 12.
Dendrogram based on hierarchical clustering with the values of six measurements indicated in Table 15. Species of subg. Behenantha are concentrated in the lower part of the diagram, corresponding with the higher values of curvature, while species of subg. Silene are grouped in the upper branches. Results are shown for the 31 species within Table 16 (12 belonging to S. subg. Behenantha in red and 19 belonging to S. subg. Silene in blue).

Table 1.
Results of the comparison (ANOVA) between rugose and echinate seeds for tubercle width (W), height (H), slope (S), maximum curvature, mean curvature and ratio between maximum and mean curvatures. Mean values and coefficients of variation (given in parentheses) are indicated in the upper row, and the minimum and maximum values are shown in the lower row. Different superscript letters indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed.
Table 1.
Results of the comparison (ANOVA) between rugose and echinate seeds for tubercle width (W), height (H), slope (S), maximum curvature, mean curvature and ratio between maximum and mean curvatures. Mean values and coefficients of variation (given in parentheses) are indicated in the upper row, and the minimum and maximum values are shown in the lower row. Different superscript letters indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed.
| N | W | H | Slope | Max Curvature | Mean Curvature | Max C/Mean C | |
|---|---|---|---|---|---|---|---|
| Rugose | 268 | 83.3 a (24.8) 31.1/174.0 | 20.7 b (35.5) 6.8/42.0 | 49.8 b (25.8) 22.5/89.7 | 41.4 b (47.1) 14.5/150.4 | 26.9 b (41.3) 9.4/69.0 | 1.6 b (30.8) 1.0/3.9 |
| Echinate | 128 | 72.0 b (23.1) 31.0/127.0 | 43.6 a (49.7) 13.0/100.0 | 124.3 a (52.2) 40.0/394.3 | 118.7 a (63.8) 17.3/477.0 | 44.2 a (48.3) 6.7/126.7 | 3.1 a (77.0) 0.27/18.4 |
Table 2.
Results of the comparison (ANOVA) between 21 species with rugose seeds for tubercle width (W), height (H), slope (S), maximum curvature, mean curvature and ratio between maximum and mean curvatures. Mean values and coefficients of variation (given in parentheses) are indicated in the upper row, and the minimum and maximum values are shown in the lower row. Mean values marked with different superscript letters indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed (the number of seeds is indicated between parenthesis). A total of 268 analyzed tubercles are presented in this section.
Table 2.
Results of the comparison (ANOVA) between 21 species with rugose seeds for tubercle width (W), height (H), slope (S), maximum curvature, mean curvature and ratio between maximum and mean curvatures. Mean values and coefficients of variation (given in parentheses) are indicated in the upper row, and the minimum and maximum values are shown in the lower row. Mean values marked with different superscript letters indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed (the number of seeds is indicated between parenthesis). A total of 268 analyzed tubercles are presented in this section.
| Species | N | W | H | Slope | Max Curvature | Mean Curvature | Max C/Mean Curv |
|---|---|---|---|---|---|---|---|
| S. acutifolia (L.) Link ex Rohrb. | 22 (2) | 92.3 c (17.4) 68.6/127.0 | 28.0 d (21.1) 18.9/41.8 | 61.5 c (20.1) 41.9/89.7 | 51.0 abcd (10.9) 43.7/64.8 | 33.2 cdef (15.5) 17.9/42.7 | 1.6 abc (23.8) 1.1/3.0 |
| S. bupleuroides L. | 18 (2) | 88.6 c (16.8) 53.2/115.5 | 20.8 abcd (27.9) 10.9/34.9 | 46.6 abc (15.6) 34.5/60.4 | 30.7 a (29.2) 17.0/49.2 | 18.5 abcd (27.5) 9.8/28.5 | 1.7 abc (17.9) 1.3/2.5 |
| S. caryophylloides (Poir.) Otth | 20 (2) | 91.8 c (21.8) 60.4/129.0 | 26.4 cd (24.3) 12.7/40.0 | 57.4 bc (12.3) 42.0/71.1 | 40.3 abc (16.3) 23.7/50.0 | 32.9 bcdef (18.2) 19.9/42.0 | 1.2 a (10.8) 1.1/1.5 |
| S. chlorantha (Willd.) Ehrh. | 20 (2) | 76.1 abc (19.5) 45.0/95.8 | 23.2 bcd (30.2) 12.0/36.9 | 60.3 c (19.2) 37.9/77.8 | 31.4 a (18.4) 22.2/43.0 | 18.9 abcd (19.7) 12.6/24.6 | 1.7 abc (23.7) 1.2/2.8 |
| S. chlorifolia Sm. | 10 (1) | 98.5 c (10.7) 79.0/117.3 | 26.4 cd (18.5) 14.5/31.7 | 53.7 bc (18.3) 32.5/67.6 | 29.6 a (36.7) 17.8/57.2 | 15.1 ab (14.0) 11.2/17.9 | 1.9 abc (29.6) 1.2/3.2 |
| S. dinarica Spreng. | 18 (3) | 86.1 bc (13.7) 64.8/108.4 | 20.4 abcd (18.2) 13.9/26.2 | 47.5 abc (16.4) 35.0/66.1 | 42.5 abc (34.2) 28.0/73.0 | 32.8 bcdef (26.9) 23.1/56.0 | 1.3 a (11.0) 1.1/1.6 |
| S. foliosa Maxim. | 16 (2) | 85.2 bc (18.0) 60.8/108.0 | 26.2 cd (34.9) 13.6/42.0 | 60.7 c (23.6) 33.3/86.6 | 32.2 a (27.7) 20.6/55.2 | 21.8 abcde (23.4) 11.4/34.1 | 1.5 abc (20.5) 1.2/2.3 |
| S. frivaldskiana Hampe | 18 (2) | 71.8 abc (12.5) 55.4/87.7 | 12.2 ab (21.9) 9.3/17.4 | 34.3 ab (23.5) 24.9/50.7 | 30.4 a (30.4) 19.5/56.2 | 18.8 abcd (22.0) 9.6/29.8 | 1.6 abc (21.4) 1.2/2.7 |
| S. gigantea (L.) L | 10 (1) | 66.0 abc (8.9) 61.0/81.0 | 16.6 abcd (22.8) 14.0/27.0 | 50.5 abc (23.0) 39.5/80.6 | 51.3 abcd (59.1) 18.0/114.0 | 28.6 abcde (29.3) 14.0/37.0 | 1.8 abc (50.3) 1.1/3.9 |
| S. inaperta L. | 19 (2) | 45.0 a (16.8) 31.1/63.0 | 9.5 a (18.2) 6.8/12.9 | 42.5 abc (15.4) 32.6/53.9 | 54.1 abcd (37.1) 18.0/112.0 | 39.6 ef (28.2) 14.0/58.0 | 1.4 abc (31.6) 1.1/2.8 |
| S. integripetala Bory & Chaub. | 13 (1) | 86.4 bc (12.8) 61.0/104.9 | 22.5 abcd (12.5) 16.0/27.7 | 52.4 bc (11.1) 43.9/66.3 | 23.7 a (26.3) 14.5/39.4 | 16.7 abc (30.6) 10.0/28.0 | 1.4 a (11.7) 1.1/1.7 |
| S. koreana Kom. | 12 (1) | 73.1 abc (19.3) 50.0/94.0 | 17.8 abcd (41.2) 8.0/31.0 | 46.9 abc (26.4) 24.2/66.7 | 32.9 a (31.9) 17.0/53.9 | 22.7 abcde (32.4) 9.4/34.5 | 1.4 abc (46.6) 1.0/3.7 |
| S. linicola C.C. Gmel. | 20 (2) | 95.4 c (11.9) 80.0/115.0 | 22.9 abcd (17.3) 16.0/29.0 | 48.2 abc (17.5) 33.7/62.7 | 40.0 ab (27.2) 21.9/58.3 | 25.5 abcde (26.2) 16.7/41.1 | 1.6 abc (23.0) 1.2/2.6 |
| S. marizii Samp. | 6 (1) | 138.7 d (15.0) 118.0/174.0 | 19.5 abcd (21.0) 15.0/26.0 | 28.0 a (8.9) 24.2/30.0 | 33.3 a (18.9) 26.0/43.0 | 20.2 abcd (37.0) 14.3/33.6 | 1.7 abc (15.9) 1.3/2.0 |
| S. nocturna L. | 6 (1) | 54.7 ab (14.5) 45.0/64.0 | 13.5 abc (16.7) 10.0/16.0 | 49.3 abc (5.7) 44.4/53.1 | 30.3 a (17.4) 24.9/39.1 | 12.7 a (9.9) 10.6/14.0 | 2.4 bc (17.7) 2.0/3.0 |
| S. otites (L.) Wibel | 6 (1) | 93.2 c (10.7) 78.0/109.0 | 16.2 abcd (32.8) 11.0/24.0 | 34.2 ab (24.3) 24.4/44.0 | 75.6 bcd (30.4) 46.5/103.9 | 47.7 f (33.0) 24.8/62.7 | 1.7 abc (29.2) 1.3/2.6 |
| S. paradoxa L. | 10 (1) | 88.7 c (9.6) 78.0/101.0 | 23.2 bcd (14.6) 20.0/29.0 | 52.3 abc (12.1) 45.5/63.0 | 54.7 abcd (25.2) 45.0/80.0 | 35.3 def (4.4) 33.1/36.8 | 1.5 abc (22.3) 1.3/2.2 |
| S. pygmaea Adams. | 6 (1) | 97.3 c (10.3) 82.0/108.0 | 20.2 abcd (29.3) 11.0/26.0 | 41.2 abc (24.7) 22.5/48.5 | 43.4 abc (19.6) 34.8/59.0 | 29.6 abcde (12.5) 23.8/34.3 | 1.5 abc (19.1) 1.2/1.9 |
| S. regia Sims. | 6 (1) | 91.7 c (4.7) 86.0/96.0 | 16.2 abcd (23.9) 12.0/21.0 | 35.1 ab (20.9) 25.5/43.8 | 46.5 abc (9.5) 40.2/53.9 | 36.3 def (8.4) 32.3/40.5 | 1.3 ab (10.2) 1.2/1.4 |
| S. spinescens Sm. | 6 (1) | 75.2 abc (4.4) 71.0/80.0 | 23.7 bcd (19.8) 18.0/30.0 | 63.4 c (22.5) 46.2/80.0 | 85.2 d (43.5) 47.3/150.4 | 48.1 f (33.0) 25.3/69.0 | 1.8 abc (16.7) 1.4/2.2 |
| S. squamigera Boiss. subsp. vesiculifera (J.Gay ex Boiss.) Coode & Cullen | 6 (1) | 88.0 c (5.0) 81.0/92.0 | 18.8 abcd (14.0) 15.0/23.0 | 42.9 abc (13.6) 32.6/50.6 | 77.4 cd (56.6) 40.4/147.0 | 32.0 bcdef (30.2) 13.3/40.3 | 2.5 c (45.7) 1.4/3.9 |
Table 3.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature of the six tubercles of Silene bupleuroides numbered 1 to 6 in Figure 1.
Table 3.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature of the six tubercles of Silene bupleuroides numbered 1 to 6 in Figure 1.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 87 | 68 | 91 | 89 | 91 | 90 |
| H | 15 | 17 | 21 | 23 | 20 | 16 |
| S | 34.5 | 50.0 | 46.2 | 51.7 | 44.0 | 35.6 |
| Max curvature | 32.1 | 45.7 | 29.8 | 24.2 | 36.1 | 28.4 |
| Mean curvature | 14.9 | 28.5 | 20.7 | 15.7 | 19.0 | 18.8 |
| Ratio Max/Mean | 2.2 | 1.6 | 1.4 | 1.5 | 1.9 | 1.5 |
Table 4.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature of the six tubercles of Silene chlorantha numbered 1 to 6 in Figure 2.
Table 4.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature of the six tubercles of Silene chlorantha numbered 1 to 6 in Figure 2.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 64 | 54 | 45 | 57 | 76 | 73 |
| H | 18 | 20 | 12 | 12 | 19 | 18 |
| S | 56.3 | 74.1 | 53.3 | 42.1 | 50.0 | 49.3 |
| Max curvature | 35.2 | 29.9 | 43.6 | 26.1 | 27.4 | 39.9 |
| Mean curvature | 20.9 | 19.0 | 19.6 | 13.6 | 12.6 | 16.8 |
| Ratio Max/Mean | 1.7 | 1.6 | 1.6 | 1.8 | 1.7 | 2.4 |
Table 5.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. frivaldskiana numbered 1 to 6 in Figure 3.
Table 5.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. frivaldskiana numbered 1 to 6 in Figure 3.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 65 | 73 | 83 | 63 | 77 | 74 |
| H | 11 | 9 | 14 | 15 | 10 | 12 |
| S | 32.9 | 25.5 | 34.3 | 47.0 | 26.7 | 31.5 |
| Max curvature | 35.7 | 32.1 | 31.5 | 22.6 | 22.0 | 28.7 |
| Mean curvature | 22.0 | 20.2 | 21.0 | 19.4 | 18.1 | 22.7 |
| Ratio Max/Mean | 1.6 | 1.6 | 1.5 | 1.2 | 1.2 | 1.3 |
Table 6.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. integripetala numbered 1 to 6 in Figure 4.
Table 6.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. integripetala numbered 1 to 6 in Figure 4.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 61 | 81 | 88 | 96 | 98 | 79 |
| H | 16 | 22 | 25 | 23 | 22 | 22 |
| S | 52.5 | 54.3 | 56.8 | 47.9 | 44.9 | 55.7 |
| Max curvature | 39.4 | 23.9 | 25.4 | 23.2 | 20.9 | 23.9 |
| Mean curvature | 28.0 | 20.7 | 19.9 | 12.9 | 10.9 | 18.6 |
| Ratio Max/Mean | 1.4 | 1.2 | 1.3 | 1.8 | 1.9 | 1.3 |
Table 7.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. spinescens numbered 1 to 6 in Figure 5.
Table 7.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. spinescens numbered 1 to 6 in Figure 5.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 78 | 71 | 75 | 73 | 74 | 80 |
| H | 18 | 27 | 30 | 26 | 21 | 20 |
| S | 46.2 | 76.1 | 80.0 | 71.2 | 56.8 | 50.0 |
| Max curvature | 65.4 | 150.4 | 100.9 | 85.7 | 61.7 | 47.3 |
| Mean curvature | 35.9 | 69.0 | 55.1 | 57.8 | 45.2 | 25.3 |
| Ratio Max/Mean | 1.8 | 2.2 | 1.8 | 1.5 | 1.4 | 1.9 |
Table 8.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. squamigera subsp. vesiculifera numbered 1 to 6 in Figure 6.
Table 8.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. squamigera subsp. vesiculifera numbered 1 to 6 in Figure 6.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 92 | 81 | 91 | 86 | 86 | 92 |
| H | 20 | 18 | 23 | 18 | 19 | 15 |
| S | 43.5 | 44.4 | 50.5 | 41.9 | 44.2 | 32.6 |
| Max curvature | 52.1 | 57.6 | 97.3 | 147.0 | 117.4 | 89.7 |
| Mean curvature | 33.7 | 43.3 | 52.1 | 46.2 | 33.6 | 35.4 |
| Ratio Max/Mean | 1.5 | 1.3 | 1.9 | 3.2 | 3.5 | 2.5 |
Table 9.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. gigantea numbered 1 to 6 in Figure 7.
Table 9.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio maximum curvature/mean curvature in six tubercles of S. gigantea numbered 1 to 6 in Figure 7.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 65 | 61 | 62 | 67 | 81 | 67 |
| H | 14 | 17 | 15 | 14 | 16 | 27 |
| S | 43.1 | 55.7 | 48.4 | 41.8 | 39.5 | 80.6 |
| Max curvature | 41.0 | 51.5 | 41.6 | 27.2 | 90.2 | 114.3 |
| Mean curvature | 31.1 | 37.2 | 31.4 | 21.7 | 32.5 | 29.5 |
| Ratio Max/Mean | 1.3 | 1.4 | 1.3 | 1.3 | 2.8 | 3.9 |
Table 10.
Results of the comparison (ANOVA) between 11 species of echinate seeds for tubercle width (W), height (H) and slope (S), maximum curvature, mean curvature and ratio of maximum and mean curvatures. Different letters in the superindex indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed (the number of seeds is indicated between parentheses). A total of 128 tubercles were analyzed in this section.
Table 10.
Results of the comparison (ANOVA) between 11 species of echinate seeds for tubercle width (W), height (H) and slope (S), maximum curvature, mean curvature and ratio of maximum and mean curvatures. Different letters in the superindex indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed (the number of seeds is indicated between parentheses). A total of 128 tubercles were analyzed in this section.
| Species | N | W | H | S | Max Curvature | Mean Curvature | Max C/Mean Curv |
|---|---|---|---|---|---|---|---|
| S. aprica subsp. oldhamiana (Miq.) C.Y.Wu ex C.L.Tang | 12 (2) | 61.0 ab (20.5) 42.2/77.0 | 49.3 bcd (15.5) 40.8/64.0 | 164.4 c (12.3) 135.5/206.6 | 104.2 abc (25.1) 77.0/167.7 | 38.4 ab (22.1) 27.0/58.0 | 2.9 ab (39.0) 1.7/5.4 |
| S. behen L. | 14 (2) | 84.5 bc (15.7) 61.5/101.7 | 77.3 f (23.4) 42.0/100.0 | 182.8 c (18.8) 133.3/229.9 | 199.9 cd (67.5) 86.1/447.0 | 49.1 bc (15.0) 39.0/66.0 | 4.4 ab (79.5) 1.7/10.9 |
| S. caroliniana Walter | 22 (2) | 72.5 bc (22.9) 53.0/108.1 | 32.8 ab (31.9) 15.0/56.0 | 90.6 ab (20.6) 40.5/123.1 | 108.5 abcd (24.3) 62.5/172.0 | 20.5 a (42.4) 6.7/41.3 | 6.1 b (52.1) 3.1/18.4 |
| S. ciliata Pourr. | 20 (2) | 64.9 ab (15.7) 48.0/83.9 | 32.8 ab (21.6) 20.3/45.6 | 101.1 ab (15.4) 62.3/128.1 | 78.5 a (28.9) 42.0/126.0 | 38.7 ab (19.2) 28.7/57.7 | 2.1 a (27.4) 1.2/3.3 |
| S. dichotoma Ehrh. | 13 (2) | 81.2 bc (14.3) 60.9/94.3 | 35.1 abc (18.8) 23.2/46.9 | 86.8 ab (15.3) 64.9/108.2 | 62.6 a (18.1) 49.2/84.1 | 40.6 ab (17.5) 32.7/53.8 | 1.6 a (14.7) 1.2/2.0 |
| S. fabaria (L.) Coyte | 6 (2) | 84.5 bc (10.8) 74.0/97.0 | 71.2 ef (13.2) 59.0/83.0 | 168.3 c (6.8) 156.5/185.2 | 161.2 abcd (21.4) 110.0/202.0 | 78.5 de (13.1) 68.0/92.0 | 2.1 a (20.4) 1.6/2.8 |
| S. longicilia (Brot.) Otth | 25 (1) | 70.9 bc (15.7) 42.0/87.0 | 23.7 a (30.9) 13.0/36.0 | 67.3 a (29.4) 40.0/120.0 | 81.5 ab (50.7) 17.3/167.0 | 40.5 ab (29.0) 20.2/64.1 | 2.0 a (33.6) 0.3/3.3 |
| S. petersonii Maguire | 4 (2) | 68.0 abc (10.5) 61.0/78.0 | 83.3 f (13.4) 71.0/98.0 | 247.8 d (20.5) 212.8/321.3 | 191.5 bcd (11.7) 178.0/225.0 | 79.6 de (17.2) 60.3/89.6 | 2.4 ab (16.1) 2.1/3.0 |
| S. samojedorum (Sambuk) Oxelman | 6 (2) | 91.2 c (25.5) 72.0/127.0 | 65.2 def (21.2) 52.0/87.0 | 146.6 bc (19.2) 100.0/175.0 | 219.2 d (37.6) 143.0/337.0 | 91.5 e (28.0) 71.0/126.7 | 2.4 a (25.1) 1.8/3.4 |
| S. yunnanensis Franch | 6 (1) | 42.2 a (33.8) 31.0/66.0 | 54.2 cde (30.8) 27.0/71.0 | 278.5 d (38.9) 101.9/394.3 | 214.3 cd (31.8) 149.8/341.2 | 68.0 cd (32.4) 42.2/107.2 | 3.2 ab (18.3) 2.8/4.2 |
Table 11.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. aprica numbered 1 to 6 in Figure 8.
Table 11.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. aprica numbered 1 to 6 in Figure 8.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 75 | 76 | 77 | 73 | 62 | 54 |
| H | 59 | 64 | 55 | 53 | 42 | 43 |
| S | 157.3 | 168.4 | 142.9 | 145.2 | 135.5 | 159.3 |
| Max curvature | 120.5 | 160.5 | 80.9 | 88.6 | 123.5 | 90.2 |
| Mean curvature | 37.1 | 30.8 | 33.3 | 37.4 | 40.2 | 39.9 |
| Ratio Max/Mean | 3.2 | 5.2 | 2.4 | 2.4 | 3.1 | 2.3 |
Table 12.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. caroliniana numbered 1 to 6 in Figure 9.
Table 12.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. caroliniana numbered 1 to 6 in Figure 9.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 79 | 74 | 59 | 64 | 69 | 54 |
| H | 32 | 15 | 26 | 35 | 29 | 29 |
| S | 81.0 | 40.5 | 88.1 | 109.4 | 84.1 | 107.4 |
| Max curvature | 78.3 | 44.2 | 72.6 | 172.3 | 110.5 | 146.7 |
| Mean curvature | 23.6 | 17.7 | 29.9 | 37.7 | 25.2 | 31.9 |
| Ratio Max/Mean | 3.3 | 2.5 | 2.4 | 4.6 | 4.4 | 4.6 |
Table 13.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. ciliata numbered 1 to 6 in Figure 10.
Table 13.
Values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. ciliata numbered 1 to 6 in Figure 10.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 69 | 49 | 48 | 61 | 73 | 73 |
| H | 34 | 28 | 23 | 30 | 33 | 42 |
| S | 99.9 | 116.3 | 95.8 | 98.0 | 91.5 | 115.5 |
| Max curvature | 74.4 | 67.7 | 42.9 | 67.2 | 41.8 | 83.7 |
| Mean curvature | 43 | 58 | 34 | 45 | 32 | 44 |
| Ratio Max/Mean | 1.7 | 1.8 | 1.3 | 1.5 | 1.3 | 1.9 |
Table 14.
Mean values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. yunnanensis numbered 1 to 6 in Figure 11.
Table 14.
Mean values of width (W), height (H), slope (S), maximum curvature, mean curvature and ratio of maximum curvature/mean curvature in six tubercles of S. yunnanensis numbered 1 to 6 in Figure 11.
| Tubercle Number | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| W | 53 | 66 | 35 | 31 | 31 | 37 |
| H | 27 | 71 | 69 | 58 | 43 | 57 |
| S | 101.9 | 215.2 | 394.3 | 374.2 | 277.4 | 308.1 |
| Max curvature | 202.5 | 149.8 | 178.6 | 179.6 | 341.2 | 234.2 |
| Mean curvature | 72.5 | 54.5 | 42.2 | 65.0 | 107.2 | 66.5 |
| Ratio Max/Mean | 2.8 | 2.7 | 4.2 | 2.8 | 3.2 | 3.5 |
Table 15.
Results of the comparison (ANOVA) between 12 species of Silene subg. Behenantha and 19 species of S. subg. Silene for tubercle width (W), height (H) and slope (S), maximum curvature, mean curvature, and ratio of maximum and mean curvature. Different letters in the superindex indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed.
Table 15.
Results of the comparison (ANOVA) between 12 species of Silene subg. Behenantha and 19 species of S. subg. Silene for tubercle width (W), height (H) and slope (S), maximum curvature, mean curvature, and ratio of maximum and mean curvature. Different letters in the superindex indicate significant differences (p < 0.05). N indicates the number of tubercles analyzed.
| N | W | H | Slope | Max Curvature | Mean Curvature | Max/Mean Curvature | |
|---|---|---|---|---|---|---|---|
| Behenantha | 130 | 82.2 a (27.4) 31.0/174.0 | 41.9 a (53.4) 12.0/100.0 | 112.4 a (64.2) 24.2/394.3 | 103.8 a (79.4) 14.5/447.0 | 39.7 a (57.6) 6.7/126.7 | 2.9 a (84.2) 1.1/18.4 |
| Silene | 263 | 78.4 a (23.9) 31.1/129.0 | 21.4 b (37.9) 6.8/45.6 | 55.1 b (35.4) 22.4/128.1 | 48.2 b (57.1) 17.0/167.0 | 29.0 b (42.0) 9.4/69.0 | 1.7 b (33.3) 0.3/3.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodríguez-Lorenzo, J.L.; Martín-Gómez, J.J.; Juan, A.; Tocino, Á.; Cervantes, E. Quantitative Analysis of Seed Surface Tubercles in Silene Species. Plants 2023, 12, 3444. https://doi.org/10.3390/plants12193444
AMA Style
Rodríguez-Lorenzo JL, Martín-Gómez JJ, Juan A, Tocino Á, Cervantes E. Quantitative Analysis of Seed Surface Tubercles in Silene Species. Plants. 2023; 12(19):3444. https://doi.org/10.3390/plants12193444
Chicago/Turabian StyleRodríguez-Lorenzo, José Luis, José Javier Martín-Gómez, Ana Juan, Ángel Tocino, and Emilio Cervantes. 2023. "Quantitative Analysis of Seed Surface Tubercles in Silene Species" Plants 12, no. 19: 3444. https://doi.org/10.3390/plants12193444
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.