
Generalist Predators Shape Biotic Resistance along a Tropical Island Chain

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. CAM Natural Enemy Surveys
2.2. FAW Predation Trials
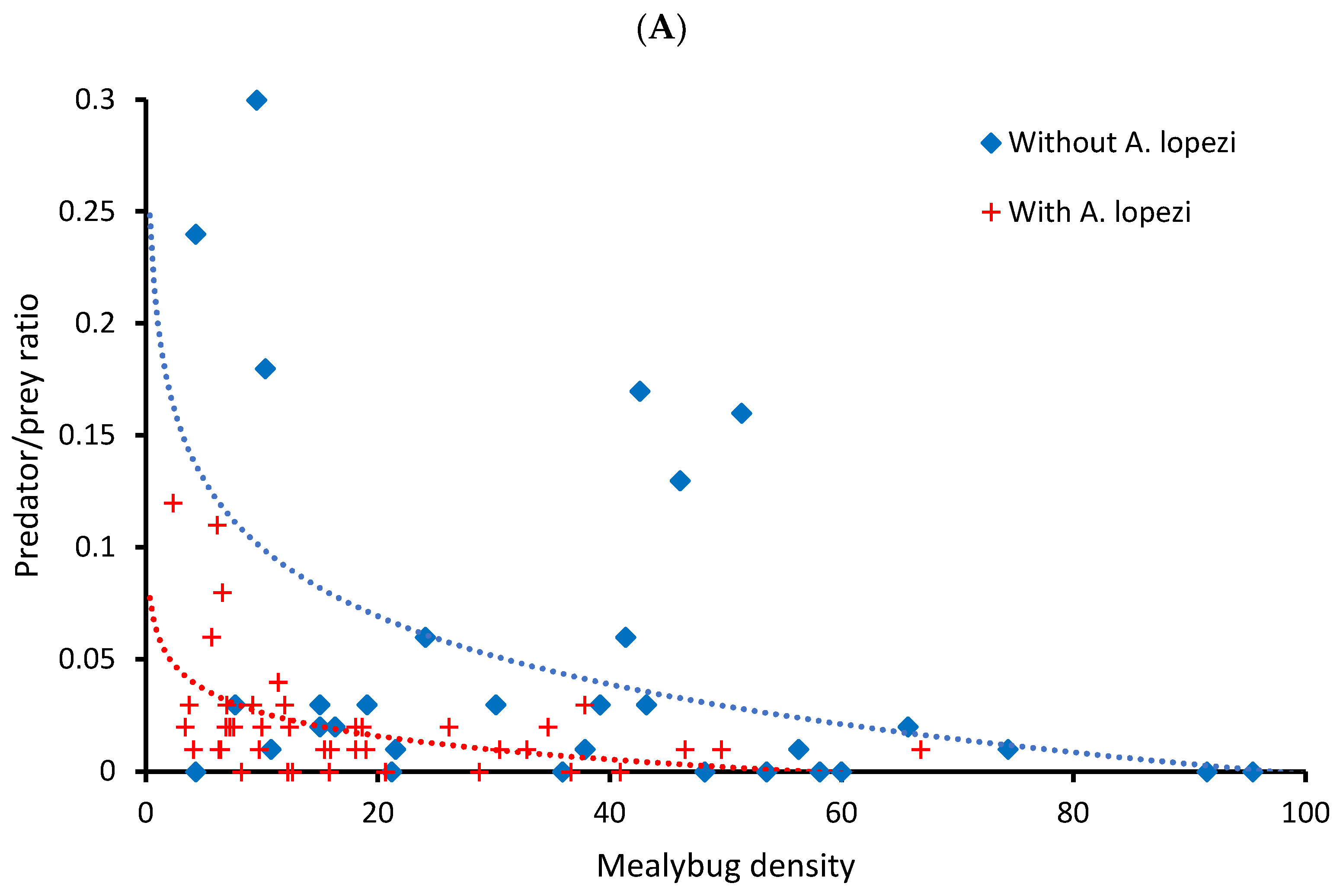
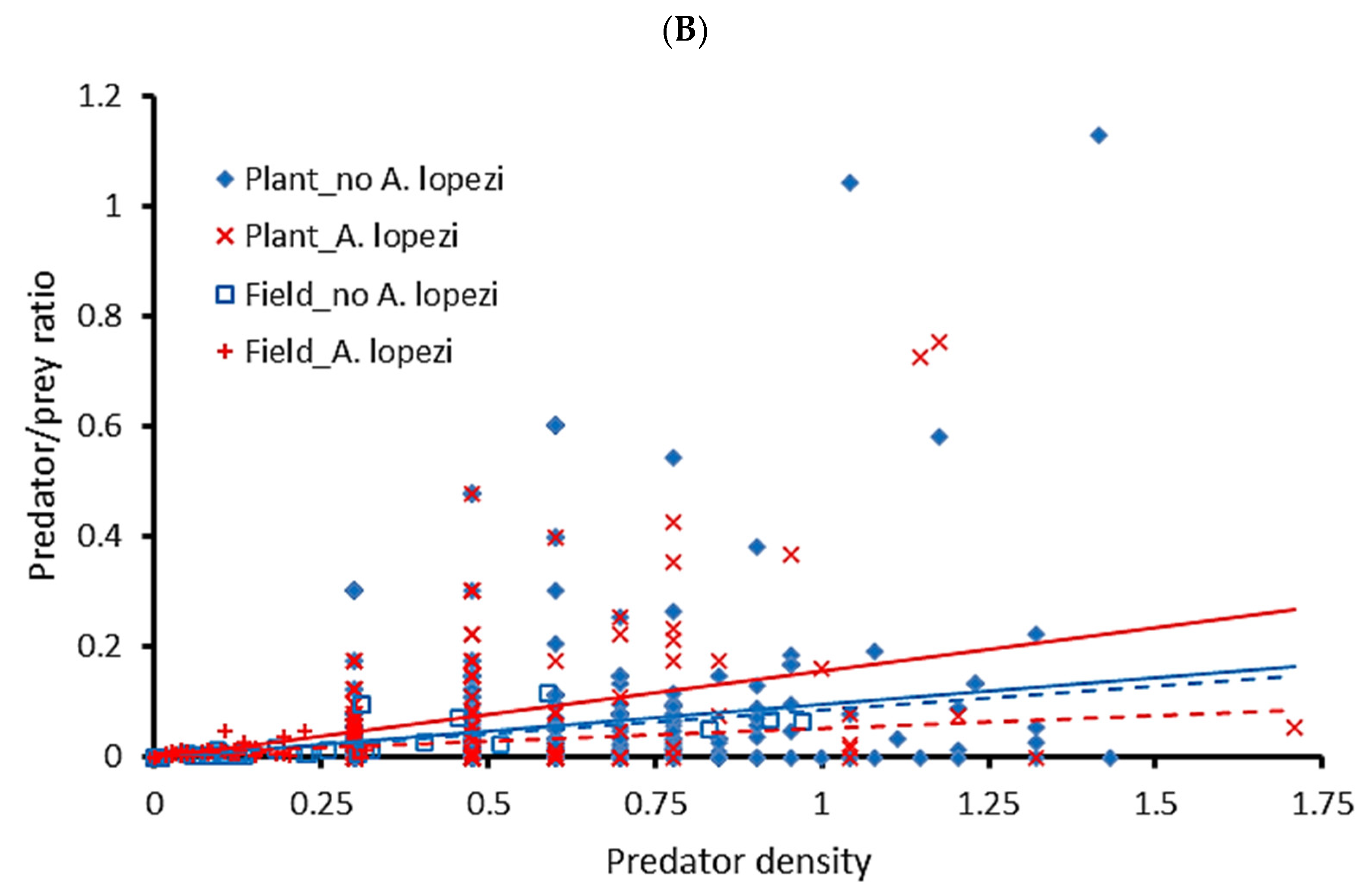
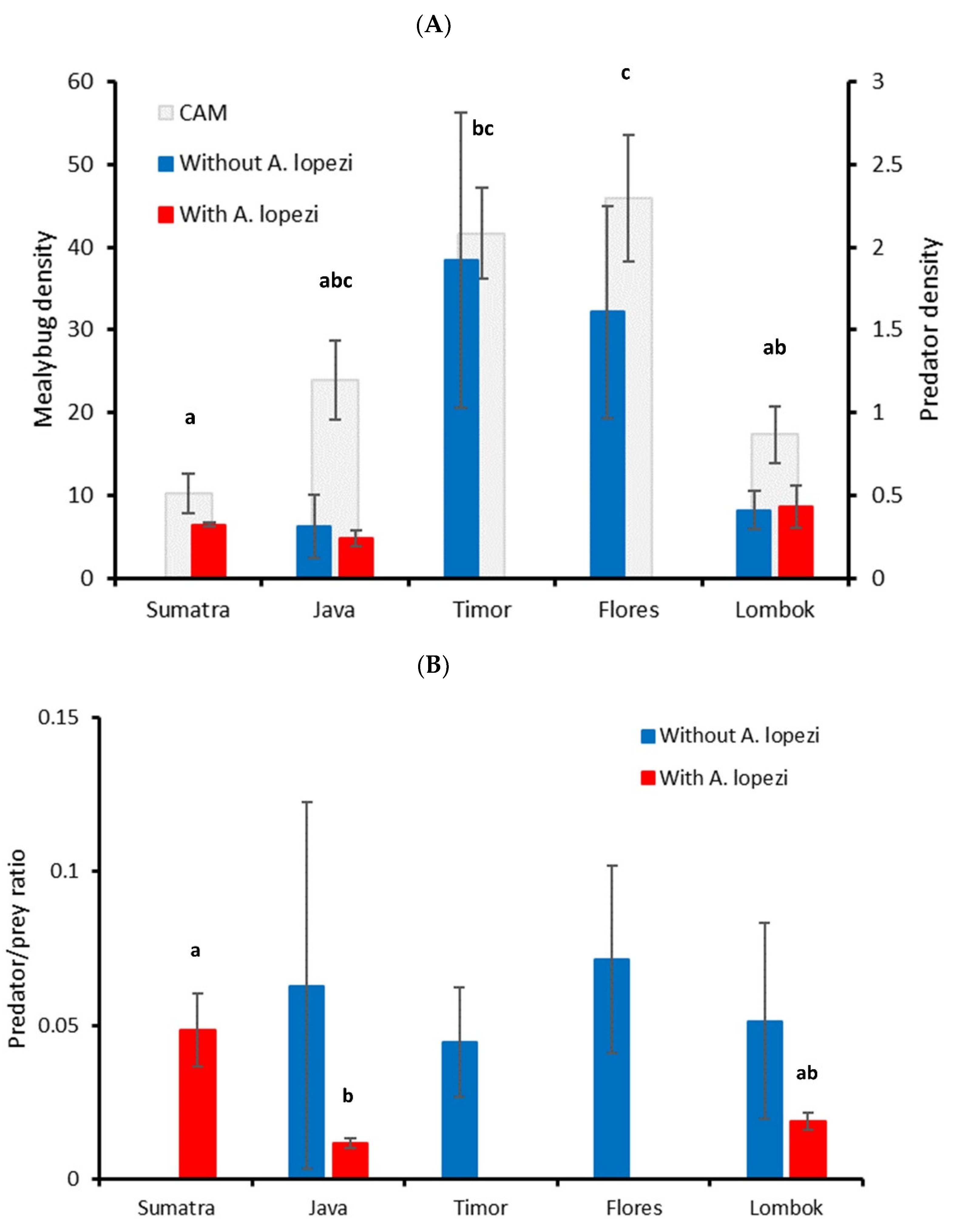
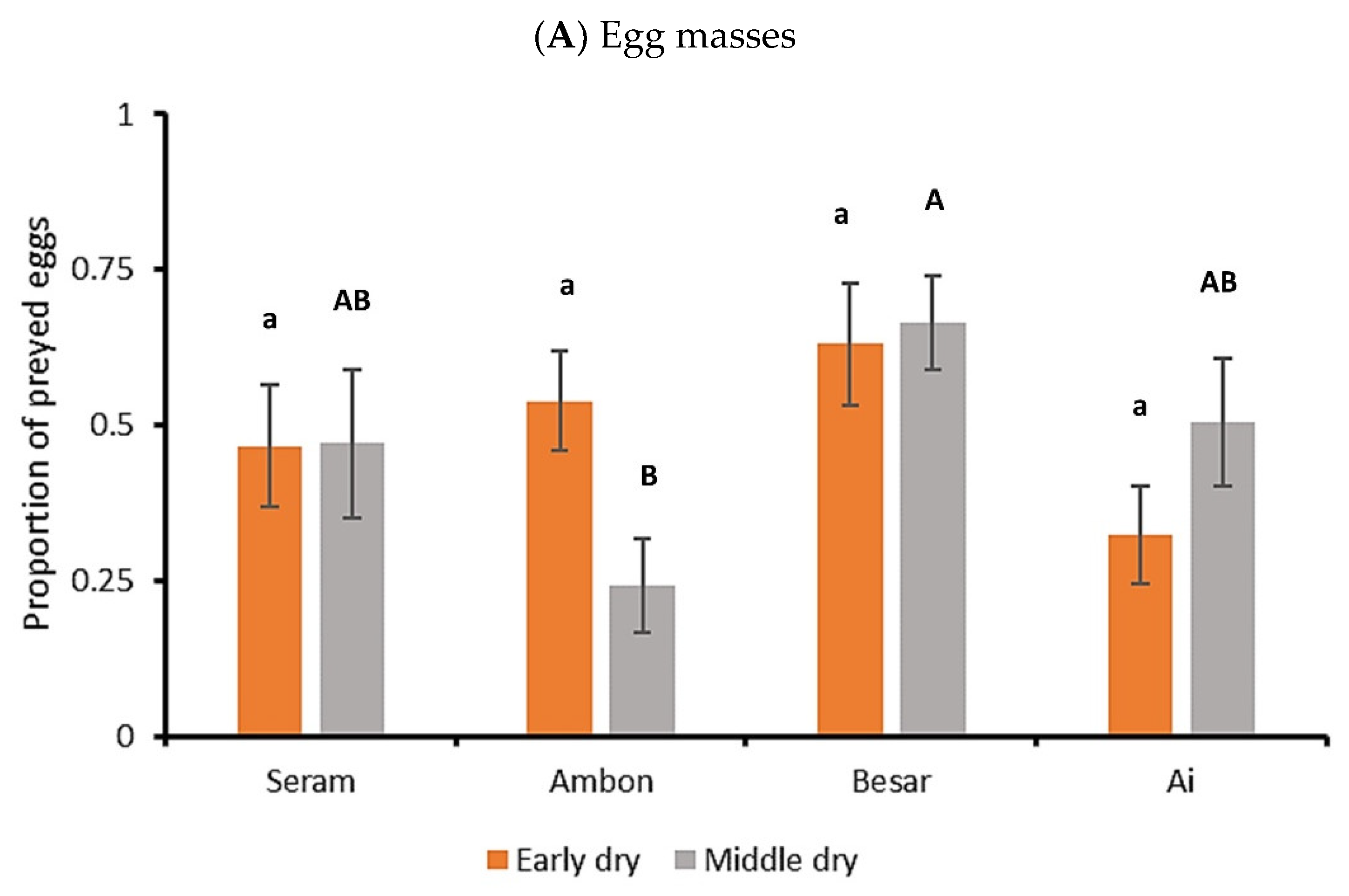
3. Results
3.1. CAM Natural Enemy Surveys
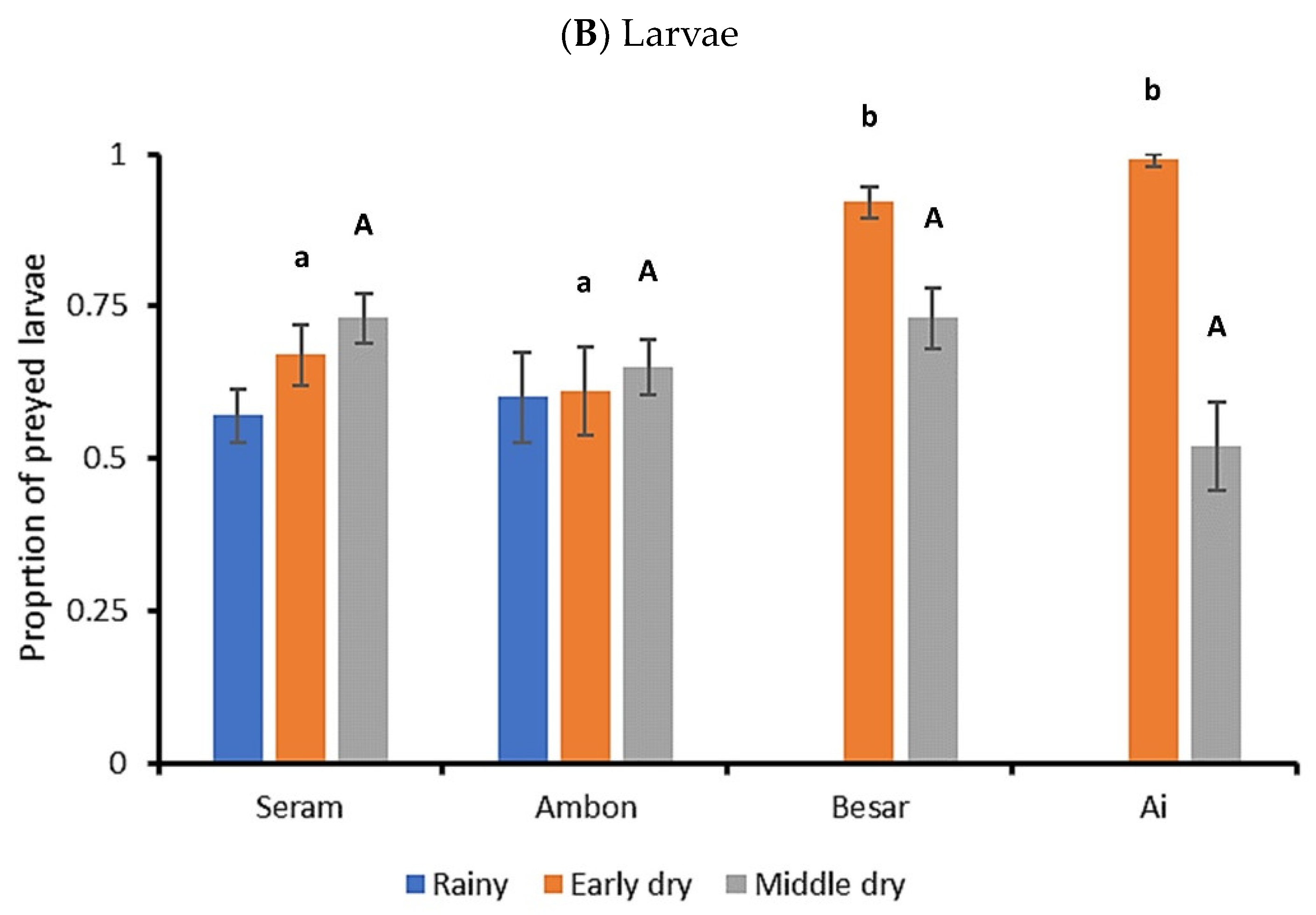
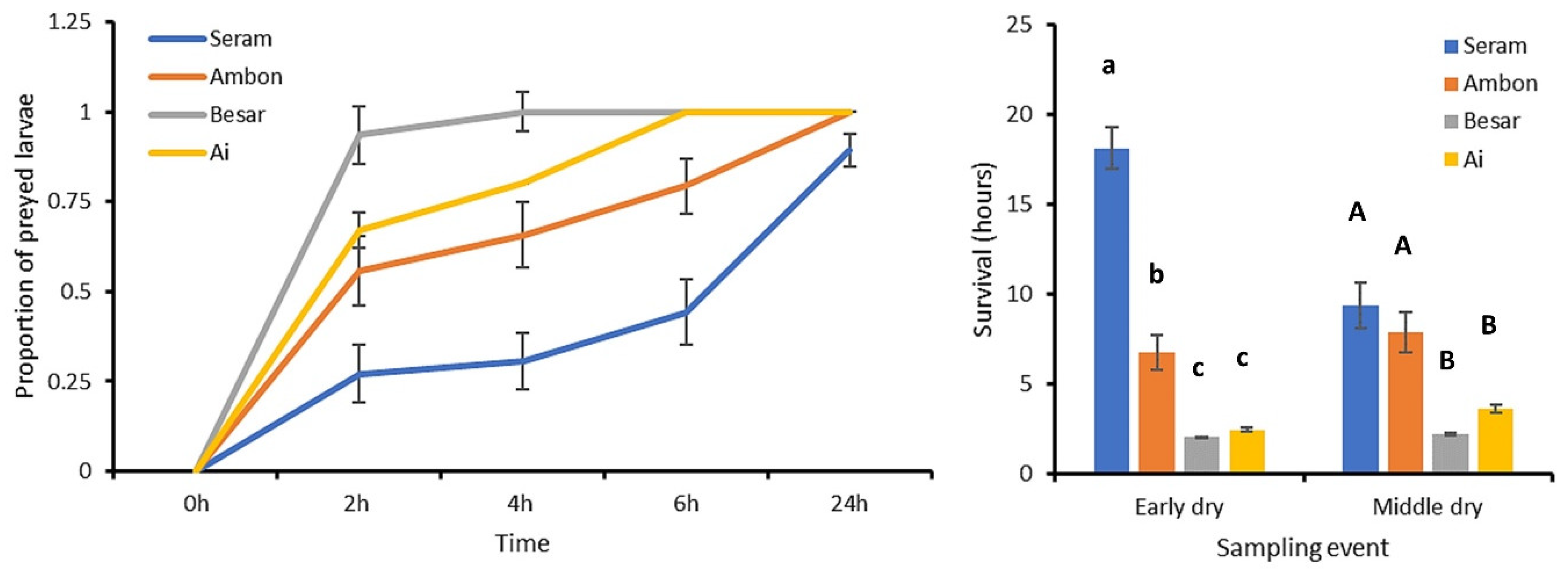
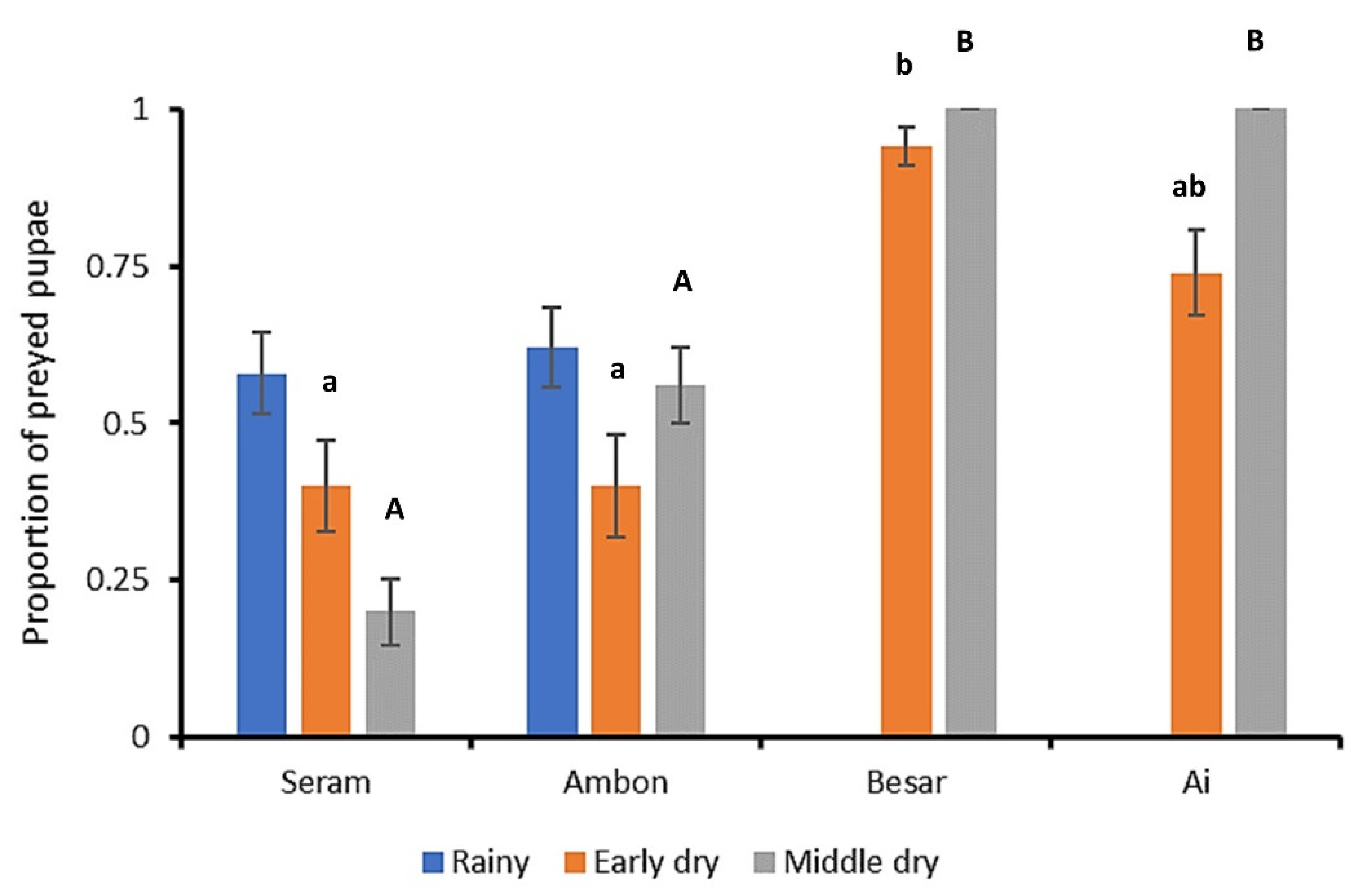
3.2. FAW Predation Trials
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gillespie, R.G.; Claridge, E.M.; Roderick, G.K. Biodiversity dynamics in isolated island communities: Interaction between natural and human-mediated processes. Mol. Ecol. 2008, 17, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilée, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J. Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef]
- Macarthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University: Princeton, NJ, USA, 1967; Volume 467. [Google Scholar]
- Wardle, D.A.; Zackrisson, O.; Hornberg, G.; Gallet, C. The influence of island area on ecosystem properties. Science 1997, 277, 1296–1299. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.C.; Kueffer, C. Island biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 2019, 44, 31–60. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef]
- Borges, P.A.; Rigal, F.; Ros-Prieto, A.; Cardoso, P. Increase of insular exotic arthropod diversity is a fundamental dimension of the current biodiversity crisis. Insect Conserv. Divers. 2020, 13, 508–518. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Hong, P.; Schmid, B.; De Laender, F.; Eisenhauer, N.; Zhang, X.; Chen, H.; Craven, D.; De Boeck, H.J.; Hautier, Y.; Petchey, O.L. Biodiversity promotes ecosystem functioning despite environmental change. Ecol. Lett. 2022, 25, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Valiente-Banuet, A.; Aizen, M.A.; Alcántara, J.M.; Arroyo, J.; Cocucci, A.; Galetti, M.; García, M.B.; García, D.; Gómez, J.M.; Jordano, P. Beyond species loss: The extinction of ecological interactions in a changing world. Funct. Ecol. 2015, 29, 299–307. [Google Scholar] [CrossRef]
- Bartley, T.J.; McCann, K.S.; Bieg, C.; Cazelles, K.; Granados, M.; Guzzo, M.M.; MacDougall, A.S.; Tunney, T.D.; McMeans, B.C. Food web rewiring in a changing world. Nat. Ecol. Evol. 2019, 3, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Hines, J.; van der Putten, W.H.; De Deyn, G.B.; Wagg, C.; Voigt, W.; Mulder, C.; Weisser, W.W.; Engel, J.; Melian, C.; Scheu, S. Towards an integration of biodiversity–ecosystem functioning and food web theory to evaluate relationships between multiple ecosystem services. Adv. Ecol. Res. 2015, 53, 161–199. [Google Scholar]
- Slobodkin, L.B.; Smith, F.E.; Hairston, N.G. Regulation in terrestrial ecosystems, and the implied balance of nature. Am. Nat. 1967, 101, 109–124. [Google Scholar] [CrossRef]
- Duffy, J.E. Biodiversity and ecosystem function: The consumer connection. Oikos 2002, 99, 201–219. [Google Scholar] [CrossRef]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A. A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef]
- Janssen, A.; van Rijn, P.C. Pesticides do not significantly reduce arthropod pest densities in the presence of natural enemies. Ecol. Lett. 2021, 24, 2010–2024. [Google Scholar] [CrossRef]
- Pimentel, D.; Wilson, C.; McCullum, C.; Huang, R.; Dwen, P.; Flack, J.; Tran, Q.; Saltman, T.; Cliff, B. Economic and environmental benefits of biodiversity. BioScience 1997, 47, 747–757. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, H.; Liu, D.; Khashaveh, A.; Li, Q.; Wyckhuys, K.A.; Wu, K. Long-term insect censuses capture progressive loss of ecosystem functioning in East Asia. Sci. Adv. 2023, 9, eade9341. [Google Scholar] [CrossRef]
- Gruner, D.S. Biotic resistance to an invasive spider conferred by generalist insectivorous birds on Hawai’i Island. Biol. Invasions 2005, 7, 541–546. [Google Scholar] [CrossRef]
- Ichihara, M.; Maruyama, K.; Yamashita, M.; Sawada, H.; Inagaki, H.; Asai, M. Quantifying the ecosystem service of non-native weed seed predation in traditional terraced paddy fields. Weed Biol. Manag. 2021, 21, 192–201. [Google Scholar] [CrossRef]
- Sunamura, E.; Tamura, S.; Urano, T.; Shoda-Kagaya, E. Predation of invasive red-necked longhorn beetle Aromia bungii (Coleoptera: Cerambycidae) eggs and hatchlings by native ants in Japan. Appl. Entomol. Zool. 2020, 55, 291–298. [Google Scholar] [CrossRef]
- Kamiyama, M.T.; Matsuura, K.; Yoshimura, T.; Yang, C.-C.S. Predation of the brown marmorated stink bug, Halyomorpha halys by the Japanese acrobat ants, Crematogaster matsumurai and Crematogaster osakensis. Biol. Control 2021, 157, 104570. [Google Scholar] [CrossRef]
- Chapman, R. Animal Ecology; University of Chicago Press: Chicago, IL, USA, 1931; 464p. [Google Scholar]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen and Co., Ltd.: London, UK, 1958. [Google Scholar]
- Levine, J.M.; D’Antonio, C.M. Elton revisited: A review of evidence linking diversity and invasibility. Oikos 1999, 87, 15–26. [Google Scholar] [CrossRef]
- Goldson, S.L.; Barker, G.M.; Chapman, H.M.; Popay, A.J.; Stewart, A.V.; Caradus, J.R.; Barratt, B.I. Severe insect pest impacts on New Zealand pasture: The plight of an ecological outlier. J. Insect Sci. 2020, 20, 17. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, J.M.; Debille, S.; Lortie, C.J. Biotic resistance and island susceptibility hypotheses. In Invasion Biology: Hypotheses and Evidence; CAB International: Wallingford, UK, 2018; pp. 60–70. [Google Scholar]
- Simberloff, D. Why do introduced species appear to devastate islands more than mainland areas? Pac. Sci. 1995, 49, 87–97. [Google Scholar]
- Graziosi, I.; Minato, N.; Alvarez, E.; Ngo, D.T.; Hoat, T.X.; Aye, T.M.; Pardo, J.M.; Wongtiem, P.; Wyckhuys, K.A. Emerging pests and diseases of South-east Asian cassava: A comprehensive evaluation of geographic priorities, management options and research needs. Pest Manag. Sci. 2016, 72, 1071–1089. [Google Scholar] [CrossRef]
- Thancharoen, A.; Lankaew, S.; Moonjuntha, P.; Wongphanuwat, T.; Sangtongpraow, B.; Ngoenklan, R.; Kittipadakul, P.; Wyckhuys, K.A. Effective biological control of an invasive mealybug pest enhances root yield in cassava. J. Pest Sci. 2018, 91, 1199–1211. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.; Hughes, A.; Buamas, C.; Johnson, A.; Vasseur, L.; Reymondin, L.; Deguine, J.-P.; Sheil, D. Biological control of an agricultural pest protects tropical forests. Commun. Biol. 2019, 2, 10. [Google Scholar] [CrossRef]
- Sartiami, D.; Dadang; Harahap, I.; Kusumah, Y.; Anwar, R. First Record of Fall Armyworm (Spodoptera frugiperda) in Indonesia and Its Occurence in Three Provinces. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; p. 012021. [Google Scholar]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Prot. 2021, 145, 105641. [Google Scholar] [CrossRef]
- Eschen, R.; Beale, T.; Bonnin, J.M.; Constantine, K.L.; Duah, S.; Finch, E.A.; Makale, F.; Nunda, W.; Ogunmodede, A.; Pratt, C.F. Towards estimating the economic cost of invasive alien species to African crop and livestock production. CABI Agric. Biosci. 2021, 2, 18. [Google Scholar]
- Neuenschwander, P.; Hennessey, R.; Herren, H. Food web of insects associated with the cassava mealybug, Phenacoccus manihoti Matile-Ferrero (Hemiptera: Pseudococcidae), and its introduced parasitoid, Epidinocarsis lopezi (De Santis) (Hymenoptera: Encyrtidae), in Africa. Bull. Entomol. Res. 1987, 77, 177–189. [Google Scholar] [CrossRef]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.-A.; Day, R.; Desneux, N.; Harrison, R.D.; Kriticos, D.; Rwomushana, I.; van den Berg, J. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2022, 43, 187–241. [Google Scholar] [CrossRef]
- Hopper, J.V.; Mills, N.J. Novel multitrophic interactions among an exotic, generalist herbivore, its host plants and resident enemies in California. Oecologia 2016, 182, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.W.; Sloan, R.J. Natural maintenance and decline of gypsy moth outbreaks. Environ. Entomol. 1978, 7, 389–395. [Google Scholar] [CrossRef]
- Broadley, H.J.; Boettner, G.H.; Schneider, B.; Elkinton, J.S. Native generalist natural enemies and an introduced specialist parasitoid together control an invasive forest insect. Ecol. Appl. 2022, 32, e2697. [Google Scholar] [CrossRef]
- Schoener, T.W.; Spiller, D.A. Effect of predators and area on invasion: An experiment with island spiders. Science 1995, 267, 1811–1813. [Google Scholar] [CrossRef]
- Schulz, A.N.; Lucardi, R.D.; Marsico, T.D. Strengthening the ties that bind: An evaluation of cross-disciplinary communication between invasion ecologists and biological control researchers in entomology. Ann. Entomol. Soc. Am. 2021, 114, 163–174. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.; Sanchez Garcia, F.J.; Santos, A.M.; Canal, N.A.; Furlong, M.J.; Melo, M.C.; Gc, Y.D.; Pozsgai, G. Island and mountain ecosystems as testbeds for biological control in the anthropocene. Front. Ecol. Evol. 2022, 10, 912628. [Google Scholar] [CrossRef]
- De Bach, P. An analysis of successes in biological control of insects in the Pacific Area. In Proceedings of the Hawaiian Society of Entomology, Honolulu, HI, USA, 21 August–6 September 1962; Volume XVIII, pp. 69–79. [Google Scholar]
- Cudjoe, A.; Neuenschwander, P.; Copland, M. Interference by ants in biological control of the cassava mealybug Phenacoccus manihoti (Hemiptera: Pseudococcidae) in Ghana. Bull. Entomol. Res. 1993, 83, 15–22. [Google Scholar] [CrossRef]
- Mace, K.C.; Mills, N.J. Connecting natural enemy metrics to biological control activity for aphids in California walnuts. Biol. Control 2017, 106, 16–26. [Google Scholar] [CrossRef]
- Ritchie, S.; Hanway, J.; Benson, G. How a Corn Plant Develops; Special Report No. 48; Iowa State University Extension: Ames, IA, USA, 1986. [Google Scholar]
- Bellows, T.S., Jr.; Van Driesche, R.; Elkinton, J. Life-table construction and analysis in the evaluation of natural enemies. Annu. Rev. Entomol. 1992, 37, 587–612. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Staab, M.; Gossner, M.M.; Simons, N.K.; Achury, R.; Ambarlı, D.; Bae, S.; Schall, P.; Weisser, W.W.; Blüthgen, N. Insect decline in forests depends on species’ traits and may be mitigated by management. Commun. Biol. 2023, 6, 338. [Google Scholar] [CrossRef] [PubMed]
- D’antonio, C.; Dudley, T. Biological Invasions as Agents of Change on Islands versus Mainlands. In Islands: Biological Diversity and Ecosystem Function; Springer: Berlin/Heidelberg, Germany, 1995; pp. 103–121. [Google Scholar]
- Crawley, M.J. The population biology of invaders. Philos. Trans. R. Soc. Lond. Ser. B 1986, 314, 711–731. [Google Scholar]
- Beard, K.H.; Durham, S.L.; Willig, M.R.; Zimmerman, J.K. Lizard and frog removal increases spider abundance but does not cascade to increase herbivory. Biotropica 2021, 53, 681–692. [Google Scholar] [CrossRef]
- Rosenheim, J.A. Source–sink dynamics for a generalist insect predator in habitats with strong higher-order predation. Ecol. Monogr. 2001, 71, 93–116. [Google Scholar]
- Bennett, A.; Gratton, C. Measuring natural pest suppression at different spatial scales affects the importance of local variables. Environ. Entomol. 2012, 41, 1077–1085. [Google Scholar] [CrossRef]
- De Heij, S.E.; Willenborg, C.J. Connected carabids: Network interactions and their impact on biocontrol by carabid beetles. Bioscience 2020, 70, 490–500. [Google Scholar] [CrossRef]
- Skein, L.; Alexander, M.E.; Robinson, T.B. Co-occurring predators increase biotic resistance against an invasive prey. Mar. Environ. Res. 2020, 157, 104929. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.D. Toward a Trophic Island Biogeography; The Theory of Island Biogeography Revisited; Princeton University Press: Princeton, NJ, USA, 2009; pp. 143–185. [Google Scholar]
- Santos, A.M.; Fontaine, C.; Quicke, D.L.; Borges, P.A.; Hortal, J. Are island and mainland biotas different? Richness and level of generalism in parasitoids of a microlepidopteran in Macaronesia. Oikos 2011, 120, 1256–1262. [Google Scholar] [CrossRef]
- Massol, F.; Dubart, M.; Calcagno, V.; Cazelles, K.; Jacquet, C.; Kefi, S.; Gravel, D. Island biogeography of food webs. Adv. Ecol. Res. 2017, 56, 183–262. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M.; Matthews, T.J.; Borregaard, M.K.; Triantis, K.A. Island biogeography: Taking the long view of nature’s laboratories. Science 2017, 357, eaam8326. [Google Scholar] [CrossRef] [PubMed]
- Hertzog, L.R.; Ebeling, A.; Weisser, W.W.; Meyer, S.T. Plant diversity increases predation by ground-dwelling invertebrate predators. Ecosphere 2017, 8, e01990. [Google Scholar] [CrossRef]
- Perfecto, I. Indirect and direct effects in a tropical agroecosystem: The maize-pest-ant system in Nicaragua. Ecology 1990, 71, 2125–2134. [Google Scholar] [CrossRef]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin III, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Matthews, T.J.; Triantis, K.A.; Whittaker, R.J. (Eds.) The Species-Area Relationship: Theory and Application; Cambridge University Press: Cambridge, UK, 2021; pp. 289–318. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wyckhuys, K.A.; O’Neil, R.J. Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize. Crop Prot. 2006, 25, 1180–1190. [Google Scholar] [CrossRef]
- Kindlmann, P.; Yasuda, H.; Kajita, Y.; Sato, S.; Dixon, A.F. Predator efficiency reconsidered for a ladybird-aphid system. Front. Ecol. Evol. 2015, 3, 27. [Google Scholar] [CrossRef]
- Cuff, J.P.; Tercel, M.P.; Drake, L.E.; Vaughan, I.P.; Bell, J.R.; Orozco-terWengel, P.; Müller, C.T.; Symondson, W.O. Density-independent prey choice, taxonomy, life history, and web characteristics determine the diet and biocontrol potential of spiders (Linyphiidae and Lycosidae) in cereal crops. Environ. DNA 2022, 4, 549–564. [Google Scholar] [CrossRef]
- Kinney, K.A.; Pintor, L.M.; Mell, A.C.; Byers, J.E. Density-dependent predation and predator preference for native prey may facilitate an invasive crab’s escape from natural enemies. Biol. Invasions 2023, 25, 2967–2976. [Google Scholar] [CrossRef]
- Calatayud, P.-A. Influence of linamarin and rutin on biological performances of Phenacoccus manihoti in artificial diets. Entomol. Exp. Appl. 2000, 96, 81–86. [Google Scholar] [CrossRef]
- Weber, D.C.; Lundgren, J.G. Assessing the trophic ecology of the Coccinellidae: Their roles as predators and as prey. Biol. Control 2009, 51, 199–214. [Google Scholar] [CrossRef]
- Roubinet, E.; Straub, C.; Jonsson, T.; Staudacher, K.; Traugott, M.; Ekbom, B.; Jonsson, M. Additive effects of predator diversity on pest control caused by few interactions among predator species. Ecol. Entomol. 2015, 40, 362–371. [Google Scholar] [CrossRef]
- Gravel, D.; Massol, F.; Canard, E.; Mouillot, D.; Mouquet, N. Trophic theory of island biogeography. Ecol. Lett. 2011, 14, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, R.N.; Callaghan, A.; Dick, J.T. A novel metric reveals biotic resistance potential and informs predictions of invasion success. Sci. Rep. 2019, 9, 15314. [Google Scholar] [CrossRef]
- Bryan, K.M.; Wratten, S.D. The responses of polyphagous predators to prey spatial heterogeneity: Aggregation by carabid and staphylinid beetles to their cereal aphid prey. Ecol. Entomol. 1984, 9, 251–259. [Google Scholar] [CrossRef]
- Freier, B.; Triltsch, H.; Möwes, M.; Moll, E. The potential of predators in natural control of aphids in wheat: Results of a ten-year field study in two German landscapes. BioControl 2007, 52, 775–788. [Google Scholar] [CrossRef]
- Lin, W.T.; Pennings, S.C. Predator–prey interactions in a ladybeetle–aphid system depend on spatial scale. Ecol. Evol. 2018, 8, 6537–6546. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Cornell, H.V. Maximum parasitism rates and successful biological control. Science 1994, 266, 1886. [Google Scholar] [CrossRef] [PubMed]
- Kaser, J.M.; Heimpel, G.E. Impact of the parasitoid Aphelinus certus on soybean aphid populations. Biol. Control 2018, 127, 17–24. [Google Scholar] [CrossRef]
- Beaury, E.M.; Fusco, E.J.; Jackson, M.R.; Laginhas, B.B.; Morelli, T.L.; Allen, J.M.; Pasquarella, V.J.; Bradley, B.A. Incorporating climate change into invasive species management: Insights from managers. Biol. Invasions 2020, 22, 233–252. [Google Scholar] [CrossRef]
- Van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef] [PubMed]
- Ricci, B.; Lavigne, C.; Alignier, A.; Aviron, S.; Biju-Duval, L.; Bouvier, J.; Choisis, J.-P.; Franck, P.; Joannon, A.; Ladet, S. Local pesticide use intensity conditions landscape effects on biological pest control. Proc. R. Soc. B 2019, 286, 20182898. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Grass, I.; Wanger, T.C.; Westphal, C.; Batáry, P. Restoring biodiversity needs more than reducing pesticides. Trends Ecol. Evol. 2022, 37, 115–116. [Google Scholar] [CrossRef]
- Willcock, S.; Cooper, G.S.; Addy, J.; Dearing, J.A. Earlier collapse of Anthropocene ecosystems driven by multiple faster and noisier drivers. Nat. Sustain. 2023, 1–12. [Google Scholar] [CrossRef]
- Finn, C.; Grattarola, F.; Pincheira-Donoso, D. More losers than winners: Investigating Anthropocene defaunation through the diversity of population trends. Biol. Rev. 2023, 98, 1732–1748. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Island | Development Stage | ||||
|---|---|---|---|---|---|---|
| Egg | Early Instar Larva | Late-Instar Larva | Pupa | Adult | ||
| Cumulative mortality | Seram | - | 0.818 | 0.867 | 0.919 | - |
| Ambon | - | 0.768 | 0.897 | 0.951 | - | |
| Besar | - | 0.948 | 0.997 | 0.999 | - | |
| Ai | - | 0.856 | 0.953 | 0.994 | - | |
| Number of individuals entering stage | Seram | 10,000 | 5320 | 1825 | 1330 | 807 |
| Ambon | 10,000 | 6100 | 2318 | 1027 | 486 | |
| Besar | 10,000 | 3530 | 522 | 33 | 1 | |
| Ai | 10,000 | 5860 | 1435 | 472 | 61 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyckhuys, K.A.G.; Leatemia, J.A.; Fanani, M.Z.; Furlong, M.J.; Gu, B.; Hadi, B.A.R.; Hasinu, J.V.; Melo, M.C.; Noya, S.H.; Rauf, A.; et al. Generalist Predators Shape Biotic Resistance along a Tropical Island Chain. Plants 2023, 12, 3304. https://doi.org/10.3390/plants12183304
Wyckhuys KAG, Leatemia JA, Fanani MZ, Furlong MJ, Gu B, Hadi BAR, Hasinu JV, Melo MC, Noya SH, Rauf A, et al. Generalist Predators Shape Biotic Resistance along a Tropical Island Chain. Plants. 2023; 12(18):3304. https://doi.org/10.3390/plants12183304
Chicago/Turabian StyleWyckhuys, Kris A. G., Johanna Audrey Leatemia, Muhammad Zainal Fanani, Michael J. Furlong, Baogen Gu, Buyung Asmara Ratna Hadi, Jeffij Virgowat Hasinu, Maria C. Melo, Saartje Helena Noya, Aunu Rauf, and et al. 2023. "Generalist Predators Shape Biotic Resistance along a Tropical Island Chain" Plants 12, no. 18: 3304. https://doi.org/10.3390/plants12183304