
Silicon Supply Improves Nodulation and Dinitrogen Fixation and Promotes Growth in Trifolium incarnatum Subjected to a Long-Term Sulfur Deprivation
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
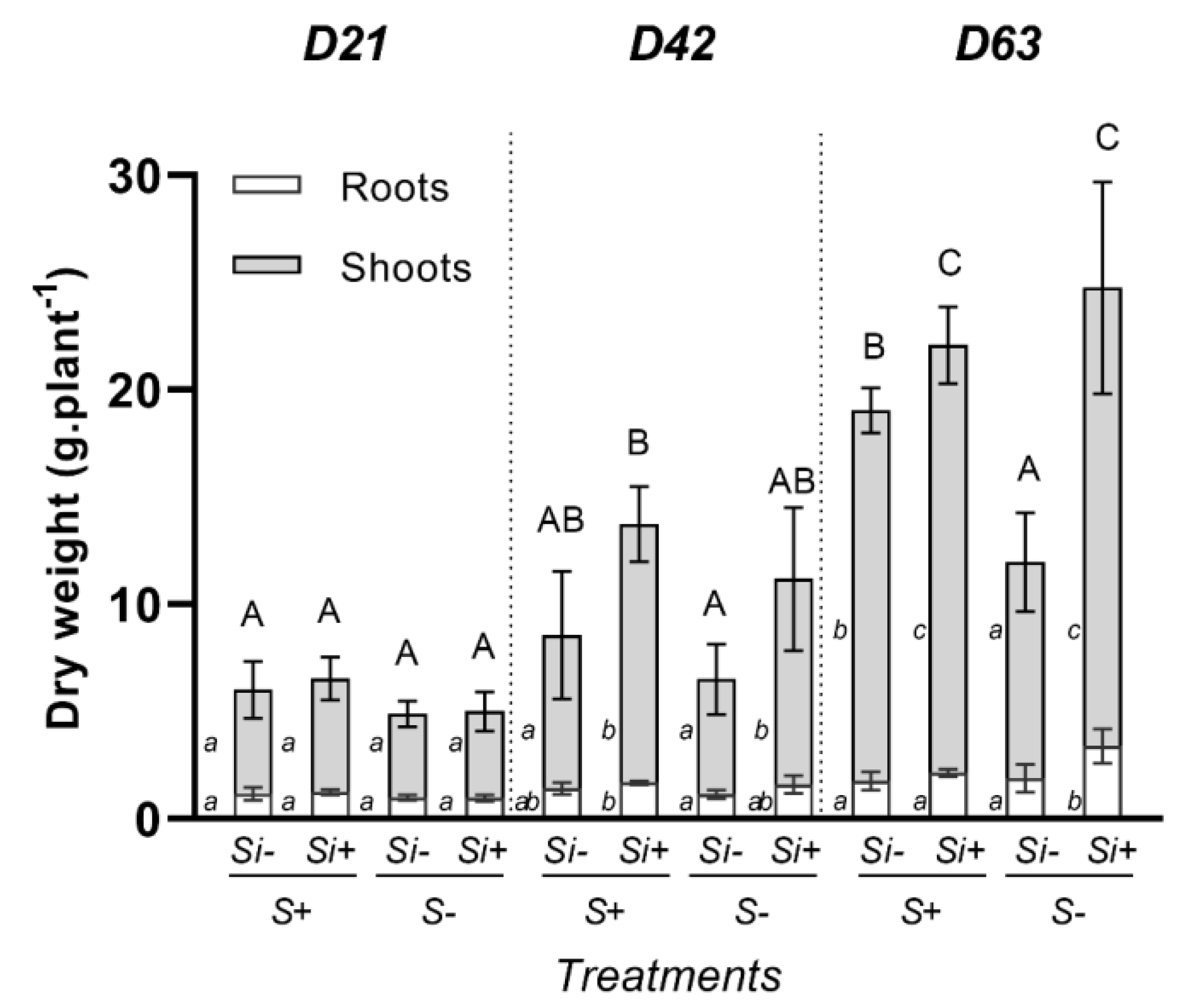
2.1. Effect of Si Supply on Growth of Trifolium incarnatum Subjected or Not to S-Deprivation
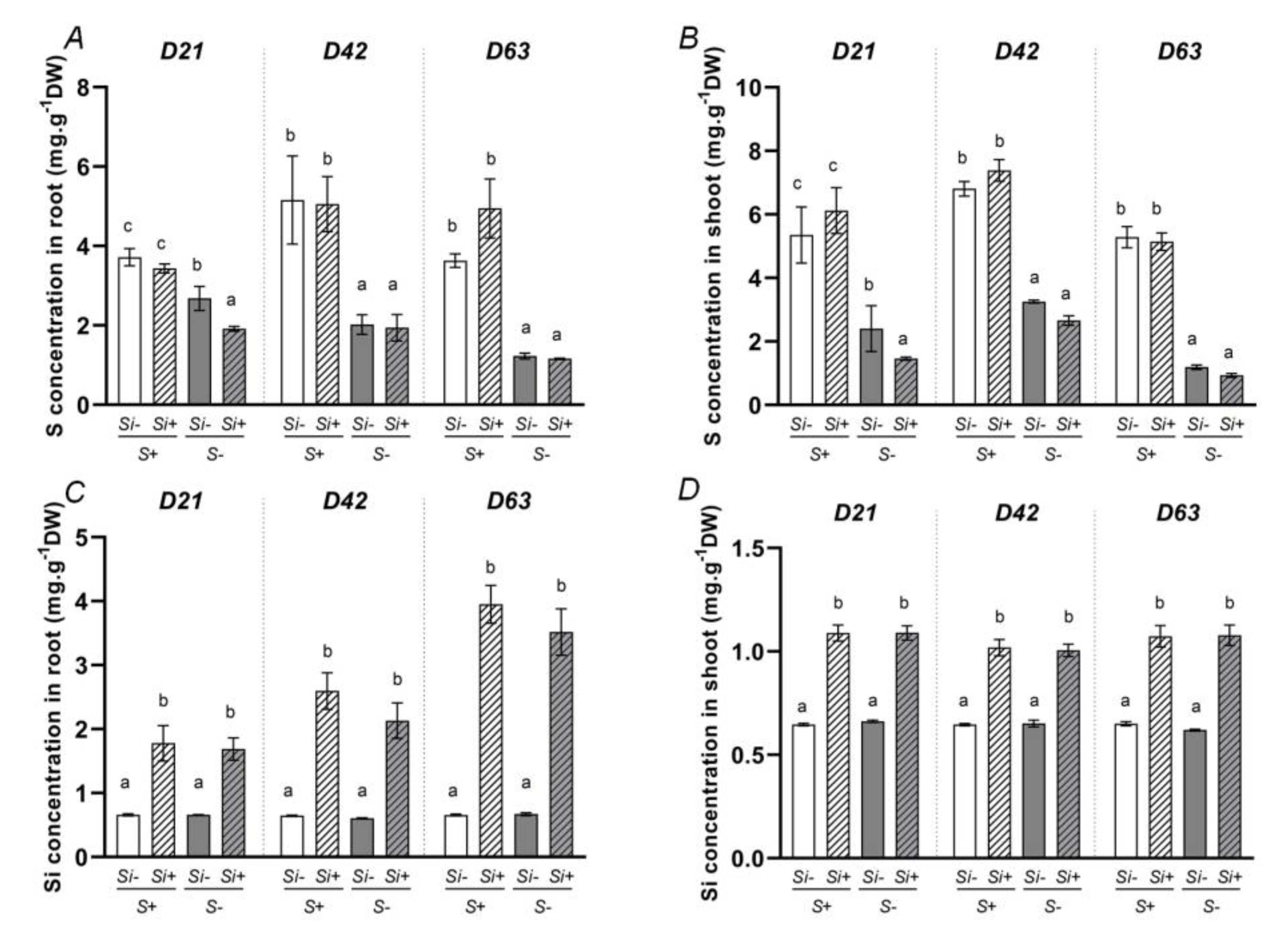
2.2. Determination of Total Sulfur (S) and Silicon (Si) Concentrations in Plants
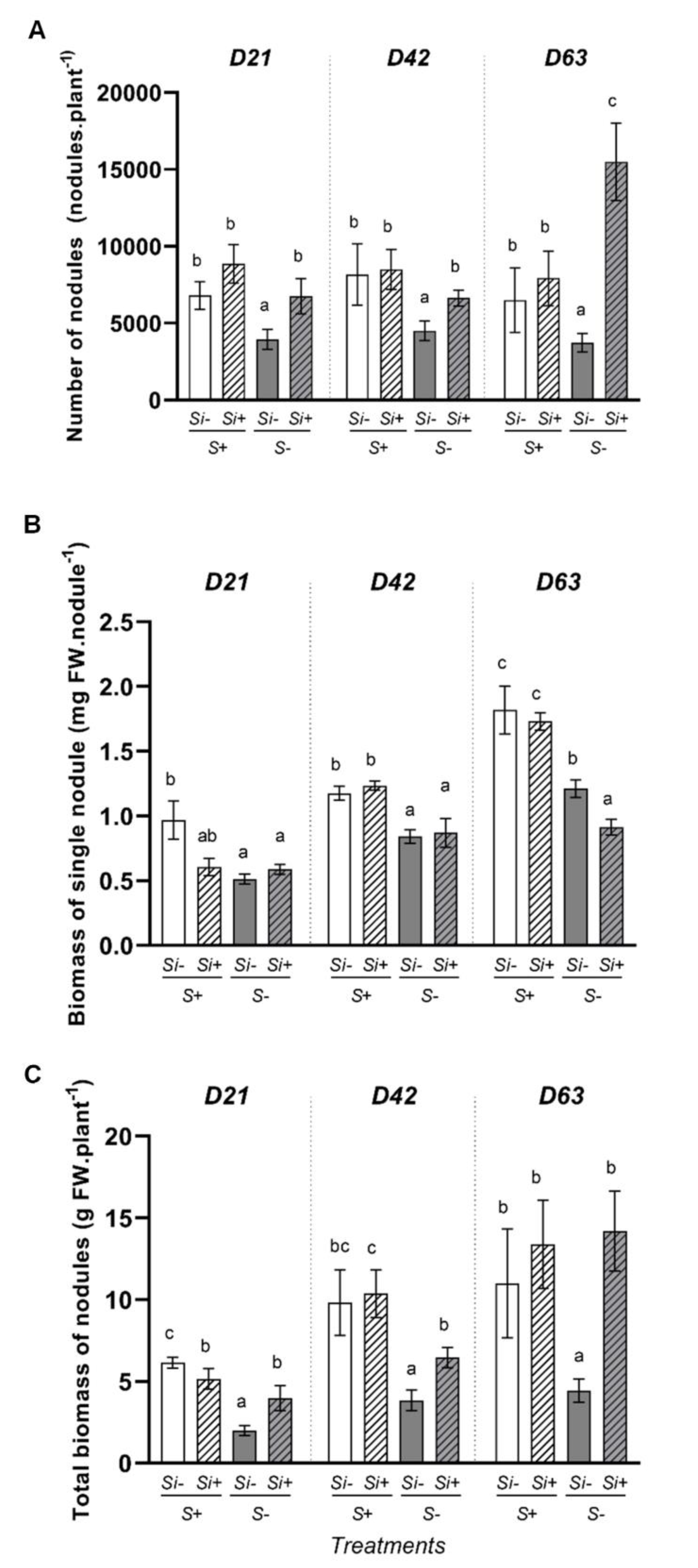
2.3. Effect of Silicon Supply on Root Nodulation
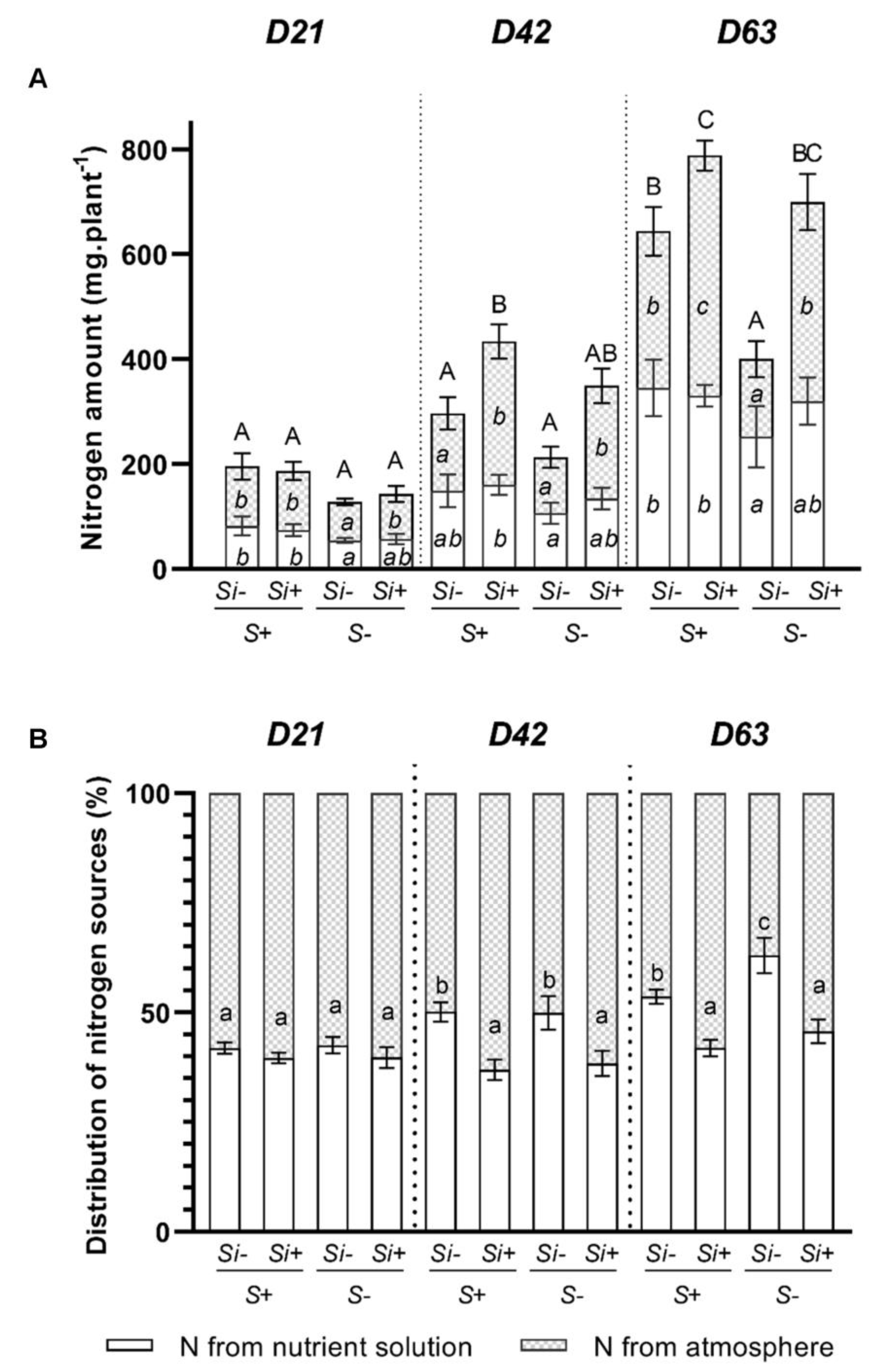
2.4. Effect of Silicon Supply on the Total Plant N Derived from Uptake and N2 Fixation
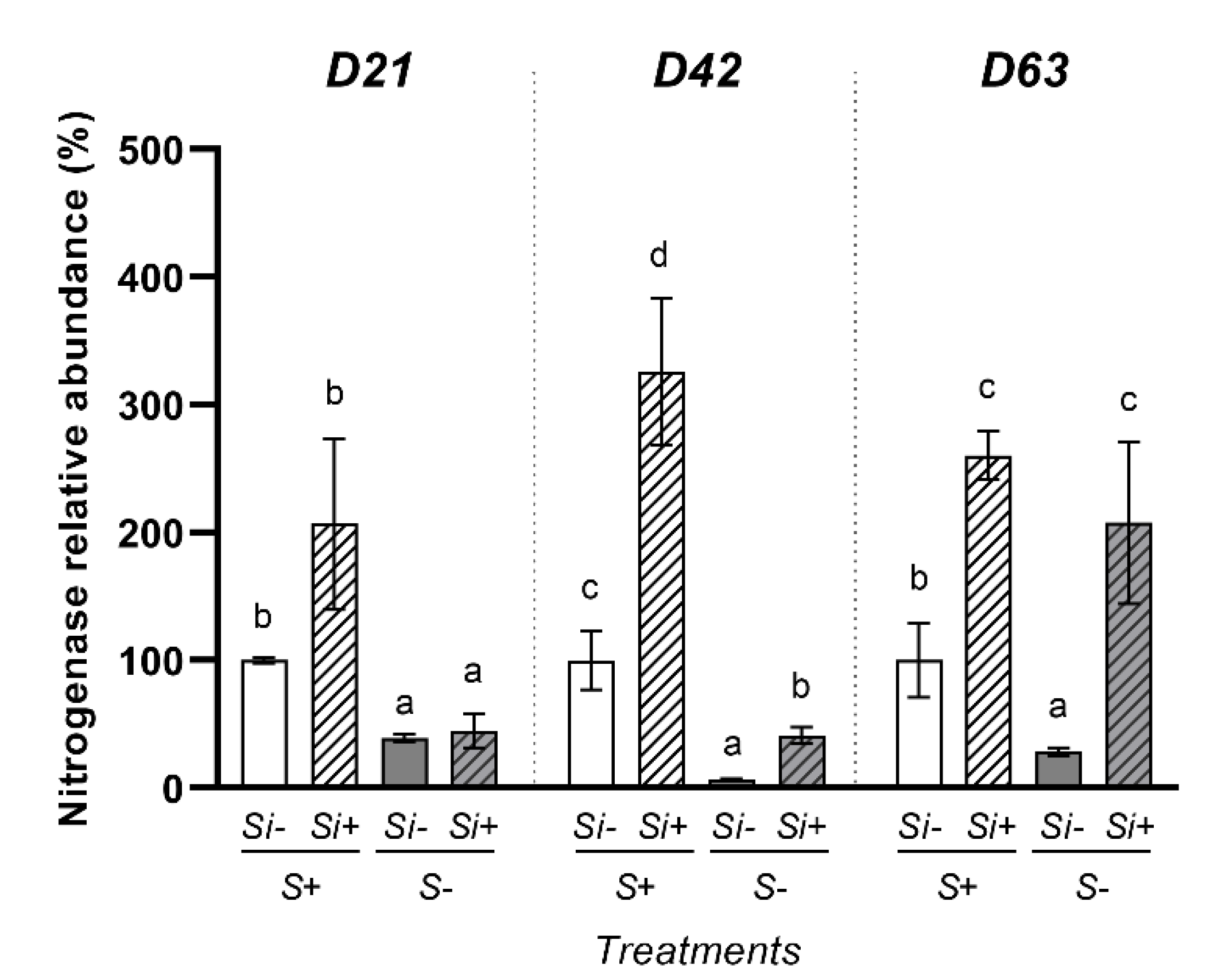
2.5. Effect of Silicon Supply on Abundance of Nitrogenase in Nodules
3. Discussion
4. Materials and Methods
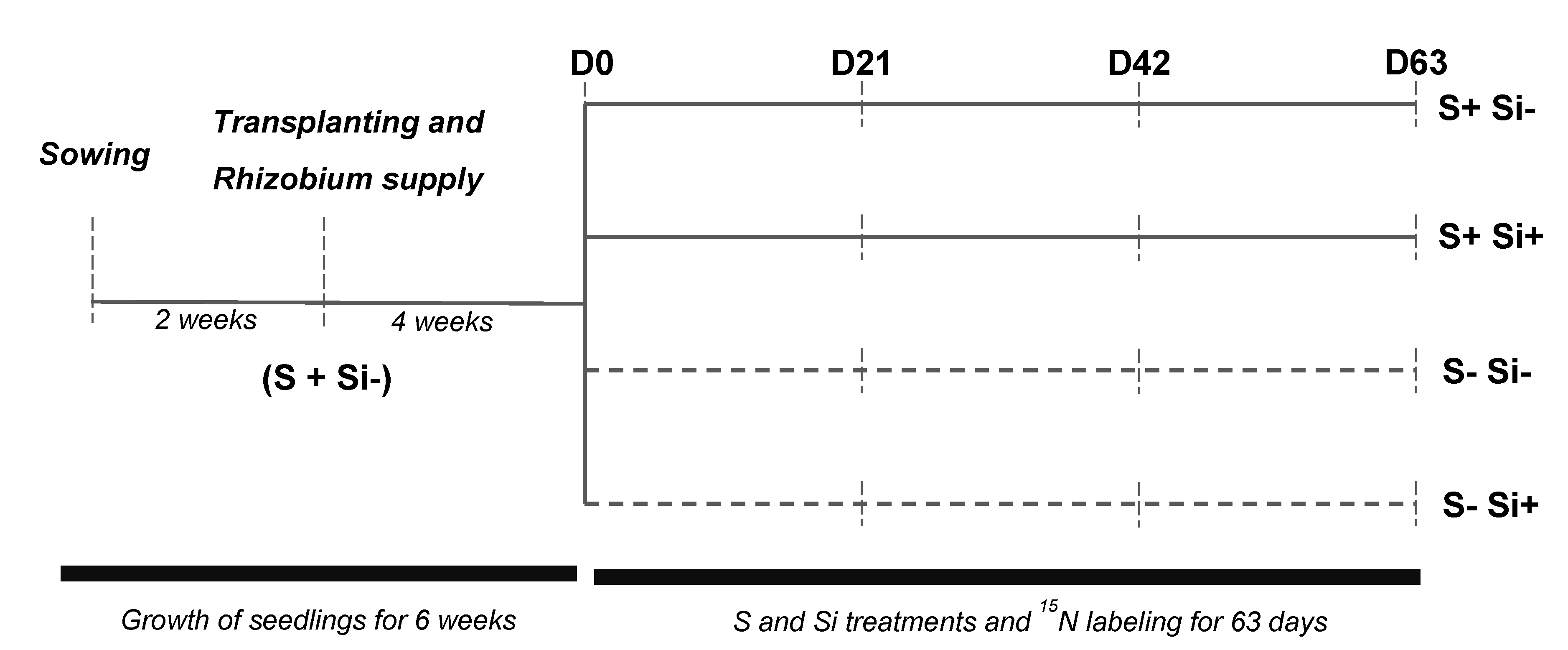
4.1. Plant Growth Conditions and Experimental Design
4.2. Determination of Total, S, Si, N and 15N Concentrations and Calculation of the Distribution of Nitrogen Sources in Plants
4.3. Extraction and Quantification of Proteins from Nodules
4.4. Immunodetection and Quantification of Nitrogenase in Root Nodules
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
International Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoesly, R.M.; Smith, S.J.; Feng, L.; Klimont, Z.; Janssens-Maenhout, G.; Pitkanen, T.; Seibert, J.J.; Vu, L.; Andres, R.J.; Bolt, R.M.; et al. Historical (1750–2014) Anthropogenic Emissions of Reactive Gases and Aerosols from the Community Emissions Data System (CEDS). Geosci. Model Dev. 2018, 11, 369–408. [Google Scholar] [CrossRef] [Green Version]
- Aas, W.; Mortier, A.; Bowersox, V.; Cherian, R.; Faluvegi, G.; Fagerli, H.; Hand, J.; Klimont, Z.; Galy-Lacaux, C.; Lehmann, C.M.B.; et al. Global and Regional Trends of Atmospheric Sulfur. Sci. Rep. 2019, 9, 953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellano, P.; Reynoso, J.; Quaranta, N. Short-Term Exposure to Sulphur Dioxide (SO2) and All-Cause and Respiratory Mortality: A Systematic Review and Meta-Analysis. Environ. Int. 2021, 150, 106434. [Google Scholar] [CrossRef]
- Feinberg, A.; Stenke, A.; Peter, T.; Hinckley, E.-L.S.; Driscoll, C.T.; Winkel, L.H.E. Reductions in the Deposition of Sulfur and Selenium to Agricultural Soils Pose Risk of Future Nutrient Deficiencies. Commun. Earth Env. 2021, 2, 101. [Google Scholar] [CrossRef]
- D’Hooghe, P.; Escamez, S.; Trouverie, J.; Avice, J.-C. Sulphur Limitation Provokes Physiological and Leaf Proteome Changes in Oilseed Rape That Lead to Perturbation of Sulphur, Carbon and Oxidative Metabolisms. BMC Plant Biol. 2013, 13, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorin, E.; Etienne, P.; Maillard, A.; Zamarreño, A.-M.; Garcia-Mina, J.-M.; Arkoun, M.; Jamois, F.; Cruz, F.; Yvin, J.-C.; Ourry, A. Effect of Sulphur Deprivation on Osmotic Potential Components and Nitrogen Metabolism in Oilseed Rape Leaves: Identification of a New Early Indicator. J. Exp. Bot. 2015, 66, 6175–6189. [Google Scholar] [CrossRef] [Green Version]
- Etienne, P.; Sorin, E.; Maillard, A.; Gallardo, K.; Arkoun, M.; Guerrand, J.; Cruz, F.; Yvin, J.-C.; Ourry, A. Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves. Plants 2018, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Varin, S.; Cliquet, J.-B.; Personeni, E.; Avice, J.-C.; Lemauviel-Lavenant, S. How Does Sulphur Availability Modify N Acquisition of White Clover (Trifolium repens L.)? J. Exp. Bot. 2010, 61, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Zuber, H.; Poignavent, G.; Le Signor, C.; Aimé, D.; Vieren, E.; Tadla, C.; Lugan, R.; Belghazi, M.; Labas, V.; Santoni, A.-L.; et al. Legume Adaptation to Sulfur Deficiency Revealed by Comparing Nutrient Allocation and Seed Traits in Medicago Truncatula. Plant J. 2013, 76, 982–996. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.; Tian, Q.-Y.; Wang, B.-L.; Zhang, W.-H. Sulfur Deficiency Had Different Effects on Medicago truncatula Ecotypes A17 and R108 in Terms of Growth, Root Morphology and Nutrient Contents. J. Plant Nutr. 2016, 39, 301–314. [Google Scholar] [CrossRef]
- Becana, M.; Wienkoop, S.; Matamoros, M.A. Sulfur Transport and Metabolism in Legume Root Nodules. Front. Plant Sci. 2018, 9, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ROS Metabolism in Plants under Environmental Stress: A Review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef] [PubMed]
- Ristova, D.; Kopriva, S. Sulfur Signaling and Starvation Response in Arabidopsis. iScience 2022, 25, 104242. [Google Scholar] [CrossRef] [PubMed]
- Scherer, H.W.; Pacyna, S.; Spoth, K.R.; Schulz, M. Low Levels of Ferredoxin, ATP and Leghemoglobin Contribute to Limited N2 Fixation of Peas (Pisum sativum L.) and Alfalfa (Medicago sativa L.) under S Deficiency Conditions. Biol. Fertil. Soils 2008, 44, 909–916. [Google Scholar] [CrossRef]
- Karmoker, J.; Clarkson, D.T.; Saker, L.R.; Rooney, J.M.; Purves, J.V. Sulphate Deprivation Depresses the Transport of Nitrogen to the Xylem and the Hydraulic Conductivity of Barley (Hordeum vulgare L.) Roots. Planta 1991, 185, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Girondé, A.; Etienne, P.; Trouverie, J.; Bouchereau, A.; Le Cahérec, F.; Leport, L.; Orsel, M.; Niogret, M.-F.; Nesi, N.; Carole, D.; et al. The Contrasting N Management of Two Oilseed Rape Genotypes Reveals the Mechanisms of Proteolysis Associated with Leaf N Remobilization and the Respective Contributions of Leaves and Stems to N Storage and Remobilization during Seed Filling. BMC Plant Biol. 2015, 15, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; She, M.; Zheng, T.; Diepeveen, D.; Islam, S.; Zhao, Y.; Zhang, Y.; Tang, G.; Zhang, Y.; Zhang, J.; et al. Impact and Mechanism of Sulphur-Deficiency on Modern Wheat Farming Nitrogen-Related Sustainability and Gliadin Content. Commun. Biol. 2021, 4, 945. [Google Scholar] [CrossRef]
- Génard, T.; Laîné, P.; Diquélou, S.; Nési, N.; Yvin, J.-C.; Etienne, P. Impact of Sulfur Applications on the Agronomic Performance of Rapeseed–Clover Mixtures. J. Plant Nutr. Soil Sci. 2017, 180, 676–682. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Tian, D.-D.; Guo, D.-J.; Chen, Z.-L.; Zhong, C.-S.; Nikpay, A.; Singh, M.; Rajput, V.D.; Singh, R.K.; et al. Influence of Silicon on Biocontrol Strategies to Manage Biotic Stress for Crop Protection, Performance, and Improvement. Plants 2021, 10, 2163. [Google Scholar] [CrossRef]
- Bakhat, H.F.; Bibi, N.; Zia, Z.; Abbas, S.; Hammad, H.M.; Fahad, S.; Ashraf, M.R.; Shah, G.M.; Rabbani, F.; Saeed, S. Silicon Mitigates Biotic Stresses in Crop Plants: A Review. Crop Prot. 2018, 104, 21–34. [Google Scholar] [CrossRef]
- Moraes, S.P.D.O.; Rodrigues, J.F.M.; Escobar, M.E.O.; Araújo, F.S.; Oliveira, T.S.D. Beneficial Effects of Silicon in Plants under Abiotic Stress Conditions: A New Approach. RCA 2022, 53, e2021821. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Ksiaa, M.; Zorrig, W.; Cabassa, C.; Abdelly, C.; Farissi, M.; Savoure, A. How Silicon Alleviates the Effect of Abiotic Stresses During Seed Germination: A Review. J. Plant Growth Regul. 2022, 42, 3323–3341. [Google Scholar] [CrossRef]
- Wang, D.; Hou, L.; Zhang, L.; Liu, P. The Mechanisms of Silicon on Maintaining Water Balance under Water Deficit Stress. Physiol. Plant. 2021, 173, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, T.; Sattar, A.; Sher, A.; Ul-Allah, S.; Ijaz, M.; Irfan, M.; Butt, M.; Cheema, M. Exogenous Application of Silicon Improves the Performance of Wheat under Terminal Heat Stress by Triggering Physio-Biochemical Mechanisms. Sci. Rep. 2021, 11, 23170. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Crilat, E.; Carol, P.; Lamsaadi, N.; Hidri, R.; Farissi, M.; Savouré, A. Salt Stress Is Alleviated by Either Proline or Silicon But Not by Their Combination in Alfalfa (Medicago sativa L.) Inoculated with a Salt-Tolerant Ensifer Meliloti Strain. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Haddad, C.; Arkoun, M.; Jamois, F.; Schwarzenberg, A.; Yvin, J.-C.; Etienne, P.; Laîné, P. Silicon Promotes Growth of Brassica napus L. and Delays Leaf Senescence Induced by Nitrogen Starvation. Front. Plant Sci. 2018, 9, 516. [Google Scholar] [CrossRef]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of Silicon With Essential and Beneficial Elements in Plants. Front. Plant Sci. 2021, 12, 697592. [Google Scholar] [CrossRef]
- Laîné, P.; Coquerel, R.; Arkoun, M.; Trouverie, J.; Etienne, P. Assessing the Effect of Silicon Supply on Root Sulfur Uptake in S-Fed and S-Deprived Brassica napus L. Plants 2022, 11, 1606. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Lamsaadi, N.; Oubenali, A.; Mouradi, M.; Savoure, A.; Farissi, M. Exogenous Silicon Application Promotes Tolerance of Legumes and Their N2 Fixing Symbiosis to Salt Stress. Silicon 2022, 14, 6517–6534. [Google Scholar] [CrossRef]
- Putra, R.; Vandegeer, R.K.; Karan, S.; Powell, J.R.; Hartley, S.E.; Johnson, S.N. Silicon Enrichment Alters Functional Traits in Legumes Depending on Plant Genotype and Symbiosis with Nitrogen-fixing Bacteria. Funct. Ecol. 2021, 35, 2856–2869. [Google Scholar] [CrossRef]
- Maillard, A.; Ali, N.; Schwarzenberg, A.; Jamois, F.; Yvin, J.-C.; Hosseini, S.A. Silicon Transcriptionally Regulates Sulfur and ABA Metabolism and Delays Leaf Senescence in Barley under Combined Sulfur Deficiency and Osmotic Stress. Environ. Exp. Bot. 2018, 155, 394–410. [Google Scholar] [CrossRef]
- Réthoré, E.; Ali, N.; Yvin, J.-C.; Hosseini, S.A. Silicon Regulates Source to Sink Metabolic Homeostasis and Promotes Growth of Rice Plants under Sulfur Deficiency. Int. J. Mol. Sci. 2020, 21, 3677. [Google Scholar] [CrossRef] [PubMed]
- Génard, T.; Etienne, P.; Diquélou, S.; Yvin, J.-C.; Revellin, C.; Laîné, P. Rapeseed-Legume Intercrops: Plant Growth and Nitrogen Balance in Early Stages of Growth and Development. Heliyon 2017, 3, e00261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Génard, T.; Etienne, P.; Laîné, P.; Yvin, J.-C.; Diquélou, S. Nitrogen Transfer from Lupinus albus L., Trifolium incarnatum L. and Vicia sativa L. Contribute Differently to Rapeseed (Brassica napus L.) Nitrogen Nutrition. Heliyon 2016, 2, e00150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Keya, S.S.; Abdelrahman, M.; Miah, M.G.; Phan Tran, L.-S. Silicon in Mitigation of Abiotic Stress-Induced Oxidative Damage in Plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef]
- Laîné, P.; Haddad, C.; Arkoun, M.; Yvin, J.-C.; Etienne, P. Silicon Promotes Agronomic Performance in Brassica Napus Cultivated under Field Conditions with Two Nitrogen Fertilizer Inputs. Plants 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Réthoré, E.; Yvin, J.-C.; Hosseini, S.A. The Regulatory Role of Silicon in Mitigating Plant Nutritional Stresses. Plants 2020, 9, 1779. [Google Scholar] [CrossRef]
- Dos Santos Sarah, M.M.; De Mello Prado, R.; De Souza Júnior, J.P.; Teixeira, G.C.M.; Dos Santos Duarte, J.C.; De Medeiros, R.L.S. Silicon Supplied via Foliar Application and Root to Attenuate Potassium Deficiency in Common Bean Plants. Sci. Rep. 2021, 11, 19690. [Google Scholar] [CrossRef]
- Hu, A.Y.; Xu, S.N.; Qin, D.N.; Li, W.; Zhao, X.Q. Role of Silicon in Mediating Phosphorus Imbalance in Plants. Plants 2020, 10, 51. [Google Scholar] [CrossRef]
- Beier, S.; Marella, N.C.; Yvin, J.-C.; Hosseini, S.A.; Von Wirén, N. Silicon Mitigates Potassium Deficiency by Enhanced Remobilization and Modulated Potassium Transporter Regulation. Environ. Exp. Bot. 2022, 198, 104849. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Marschner, H. Plant Uptake and Utilization of Nitrogen. In Nitrogen Fertilization in the Environment; Marcel Dekker, Inc.: New York, NY, USA, 1995; pp. 41–81. [Google Scholar]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.M.; HafeezLaghari, A.; MustafaBhabhan, G.; HussainTalpur, K.; Lashari, A.A. Role of Nitrogen for Plant Growth and Development: A Review. Adv. Environ. Biol. 2016, 10, 209–219. [Google Scholar]
- Jamal, A.; Moon, Y.-S.; Abdin, M. Sulphur—A General Overview and Interaction with Nitrogen. Aust. J. Crop Sci. 2010, 4, 523–529. [Google Scholar]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Revisiting Sulphur—The Once Neglected Nutrient: It’s Roles in Plant Growth, Metabolism, Stress Tolerance and Crop Production. Agriculture 2021, 11, 626. [Google Scholar] [CrossRef]
- Chandra, N.; Pandey, N. Influence of Sulfur Induced Stress on Oxidative Status and Antioxidative Machinery in Leaves of Allium Cepa L. Int. Sch. Res. Not. 2014, 2014, 568081. [Google Scholar] [CrossRef] [Green Version]
- Mhadhbi, H.; Chihaoui, S.; Mhamdi, R.; Mnasri, B.; Jebara, M.; Mhamdi, R. A Highly Osmotolerant Rhizobial Strain Confers a Better Tolerance of Nitrogen Fixation and Enhances Protective Activities to Nodules of Phaseolus vulgaris under Drought Stress. Afr. J. Biotechnol. 2011, 10, 4555–4563. [Google Scholar] [CrossRef]
- Putra, R.; Waterman, J.M.; Mathesius, U.; Wojtalewicz, D.; Powell, J.R.; Hartley, S.E.; Johnson, S.N. Benefits of Silicon-Enhanced Root Nodulation in a Model Legume Are Contingent upon Rhizobial Efficacy. Plant Soil 2022, 477, 201–217. [Google Scholar] [CrossRef]
- Jamont, M.; Piva, G.; Fustec, J. Sharing N Resources in the Early Growth of Rapeseed Intercropped with Faba Bean: Does N Transfer Matter? Plant Soil 2013, 371, 641–653. [Google Scholar] [CrossRef]
- Gulati, S.L. New Nonsynthetic Medium ForRhizobium Culture Production from Wastes. Biotechnol. Bioeng. 1979, 21, 1507–1515. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coquerel, R.; Arkoun, M.; Dupas, Q.; Leroy, F.; Laîné, P.; Etienne, P. Silicon Supply Improves Nodulation and Dinitrogen Fixation and Promotes Growth in Trifolium incarnatum Subjected to a Long-Term Sulfur Deprivation. Plants 2023, 12, 2248. https://doi.org/10.3390/plants12122248
Coquerel R, Arkoun M, Dupas Q, Leroy F, Laîné P, Etienne P. Silicon Supply Improves Nodulation and Dinitrogen Fixation and Promotes Growth in Trifolium incarnatum Subjected to a Long-Term Sulfur Deprivation. Plants. 2023; 12(12):2248. https://doi.org/10.3390/plants12122248
Chicago/Turabian StyleCoquerel, Raphaël, Mustapha Arkoun, Quentin Dupas, Fanny Leroy, Philippe Laîné, and Philippe Etienne. 2023. "Silicon Supply Improves Nodulation and Dinitrogen Fixation and Promotes Growth in Trifolium incarnatum Subjected to a Long-Term Sulfur Deprivation" Plants 12, no. 12: 2248. https://doi.org/10.3390/plants12122248