
Effect of Green Synthesized ZnO-NPs on Growth, Antioxidant System Response and Bioactive Compound Accumulation in Echinops macrochaetus, a Potential Medicinal Plant, and Assessment of Genome Size (2C DNA Content)
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
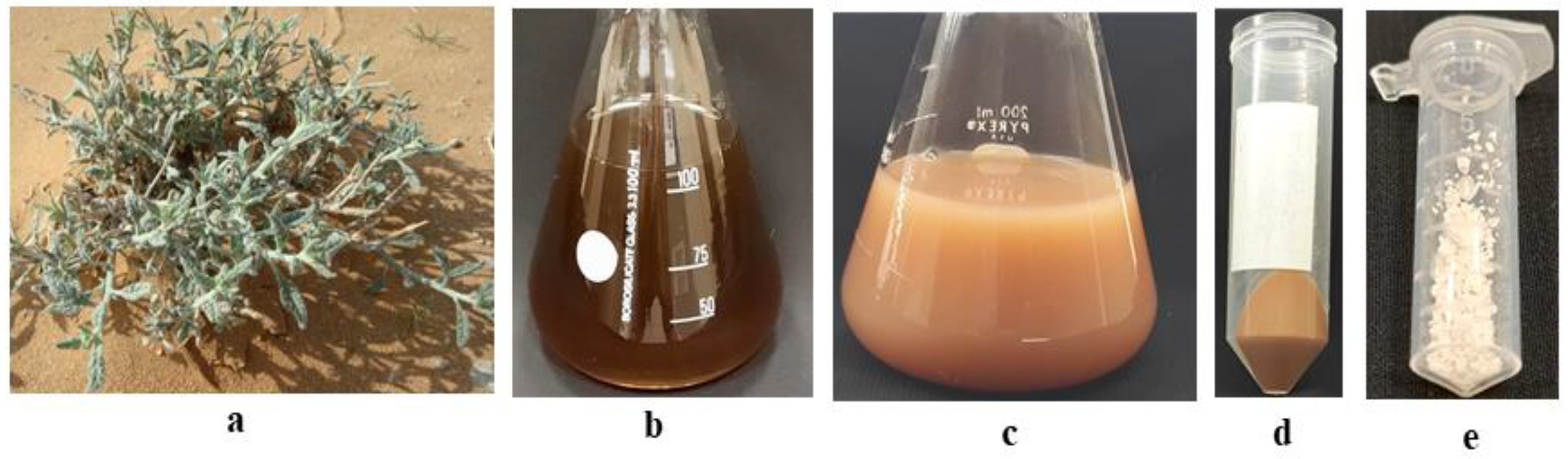
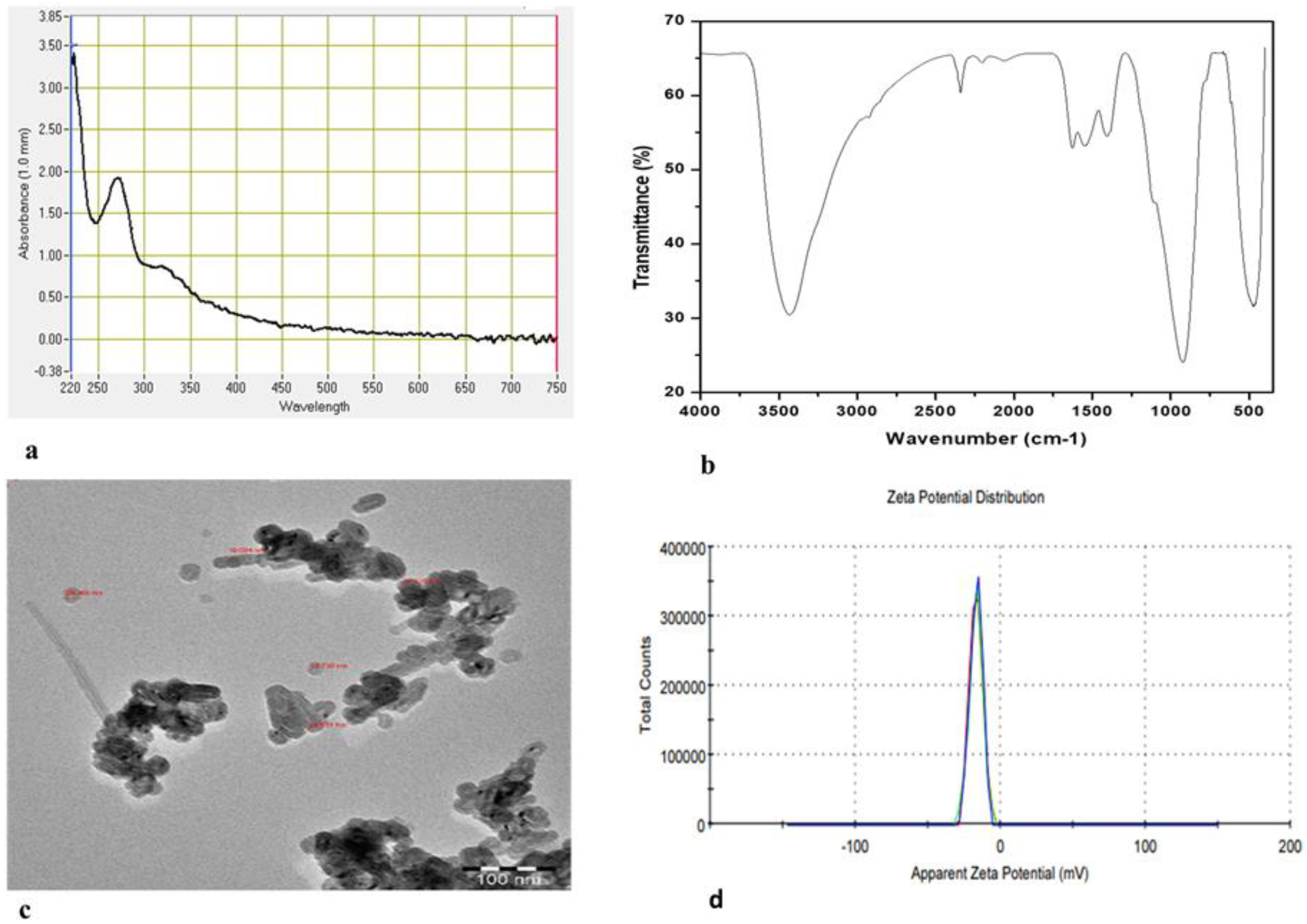
2.1. Biosynthesis and Characterization of ZnO-NPs
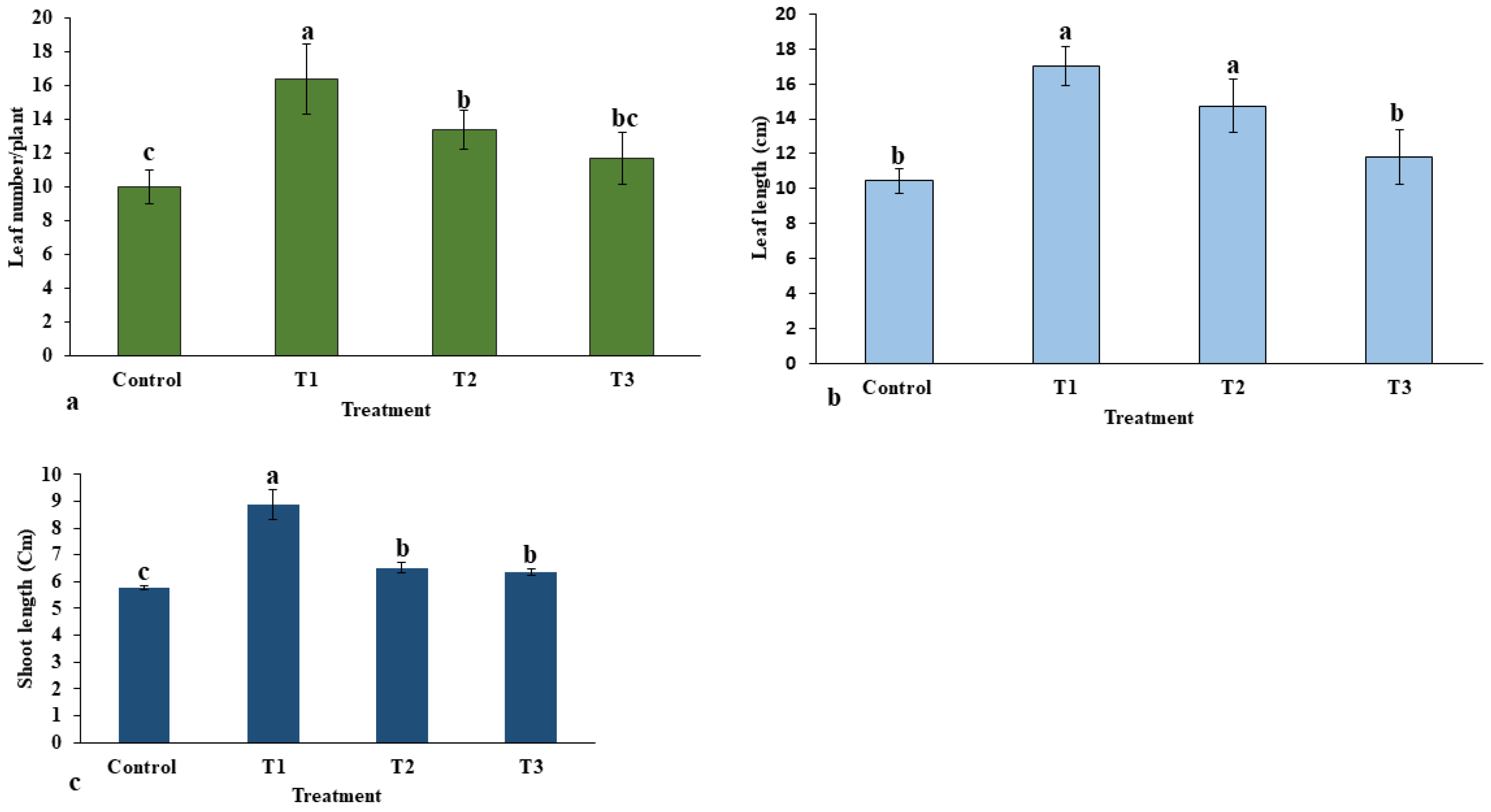
2.1.1. Effect of ZnO-NPs on Morphological Traits
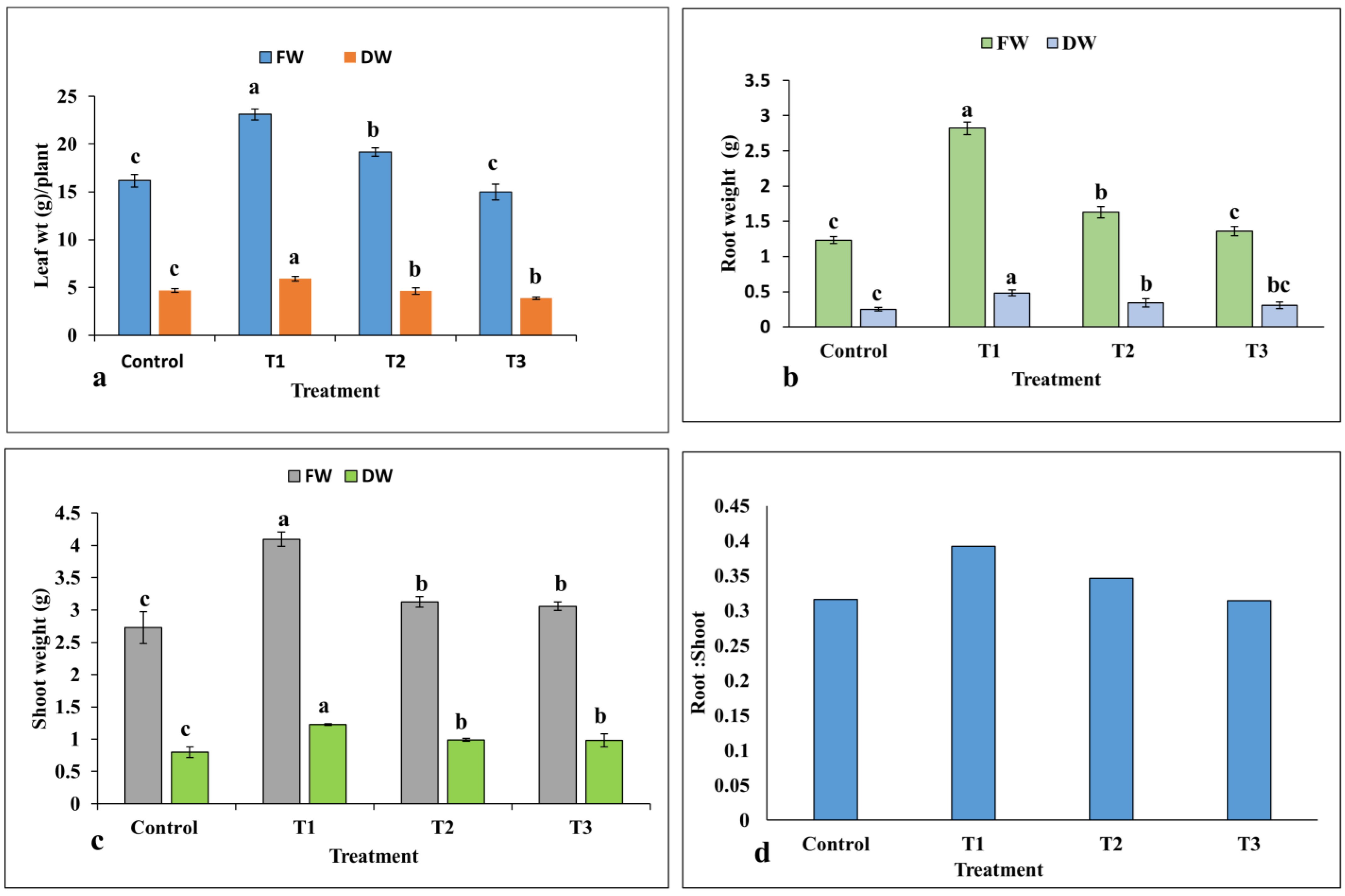
2.1.2. Effect of ZnO-NPs on Biomass Accumulation
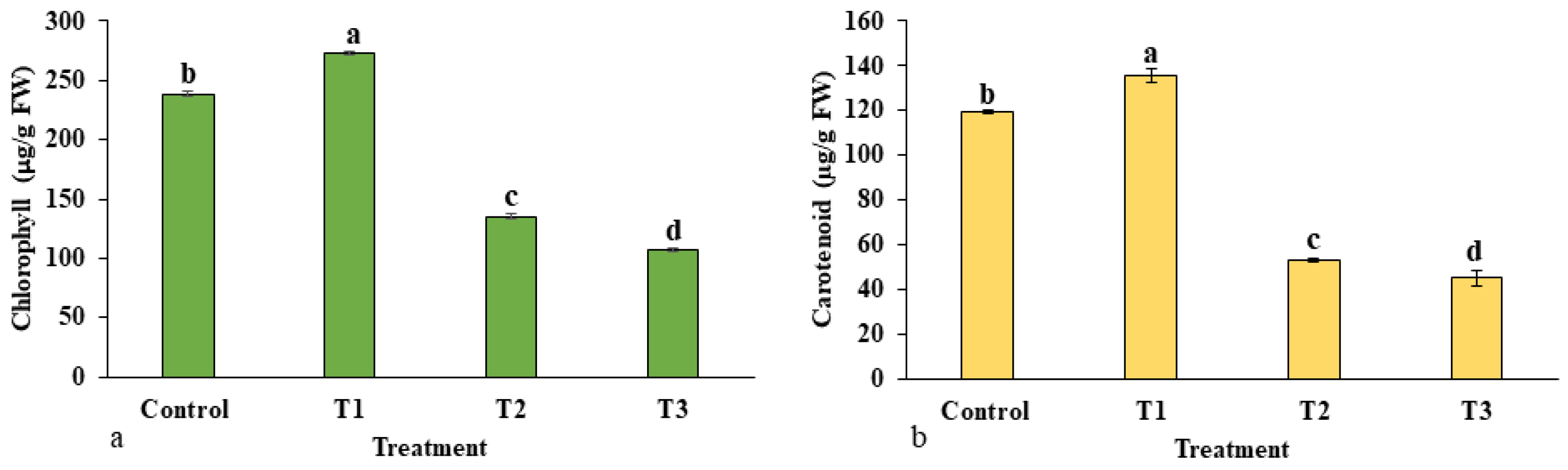
2.1.3. Photosynthetic Pigment Content
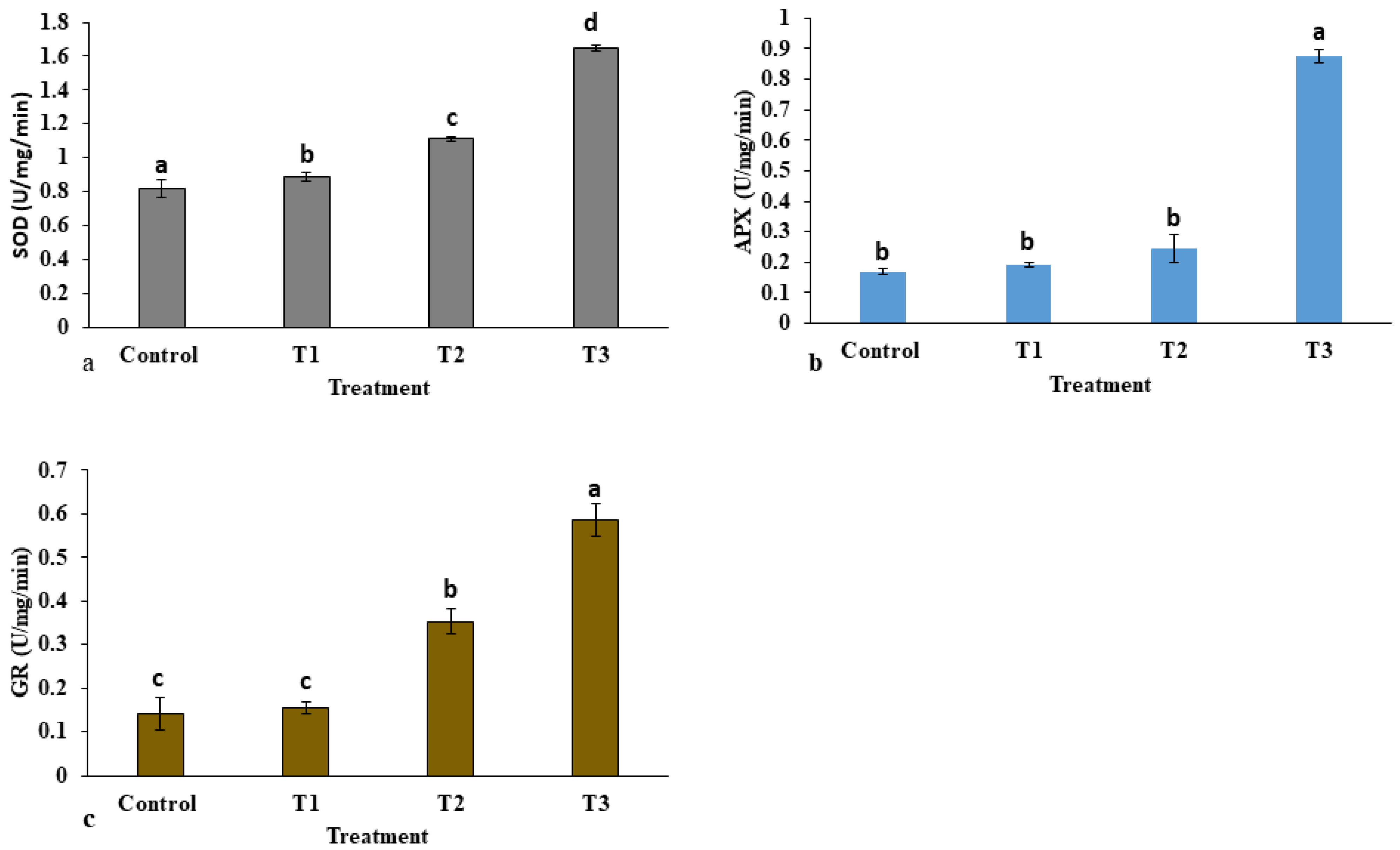
2.1.4. Antioxidant Enzyme Activity
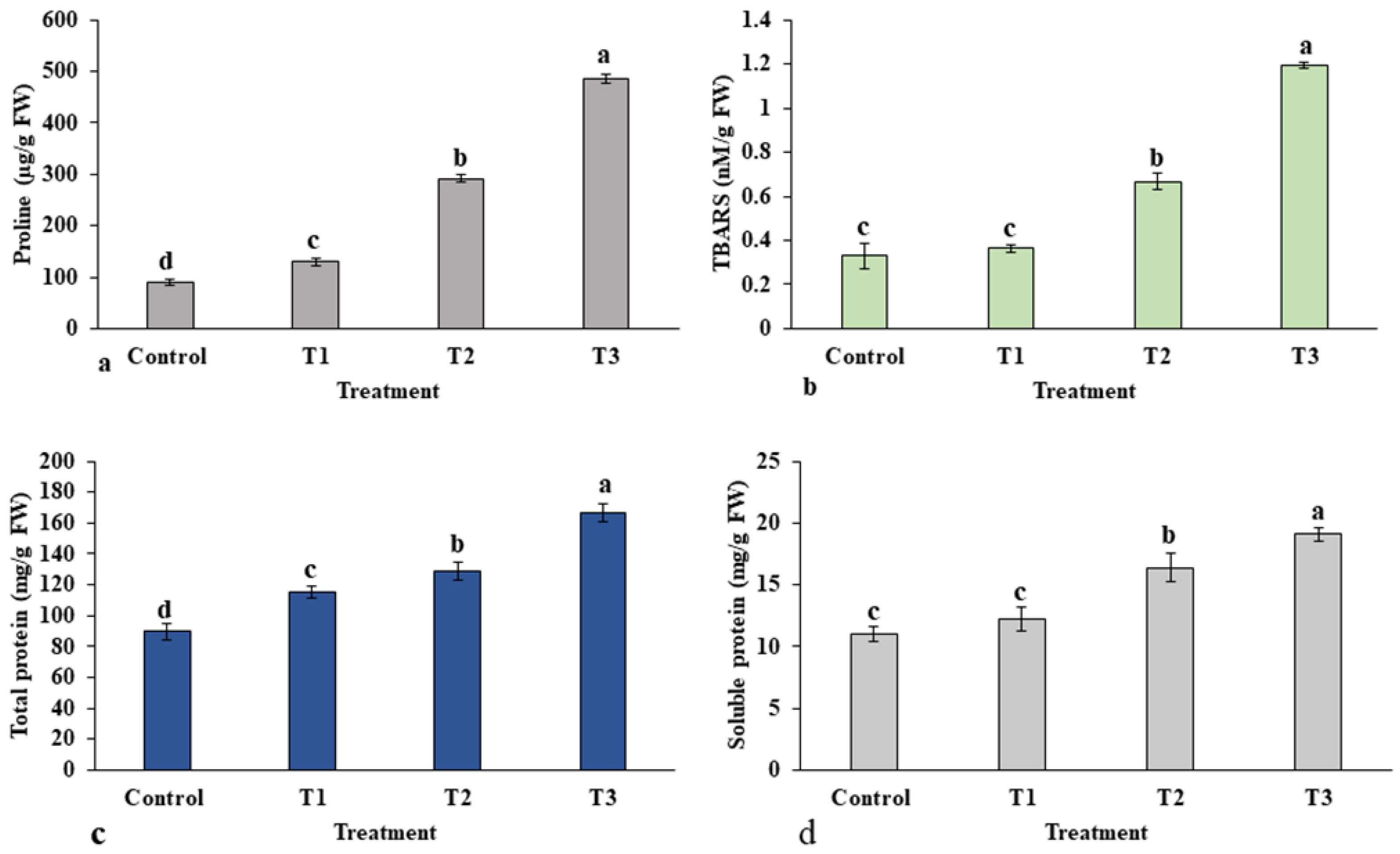
2.1.5. Estimation of Proline and TBARS Content
2.1.6. Total Protein Content
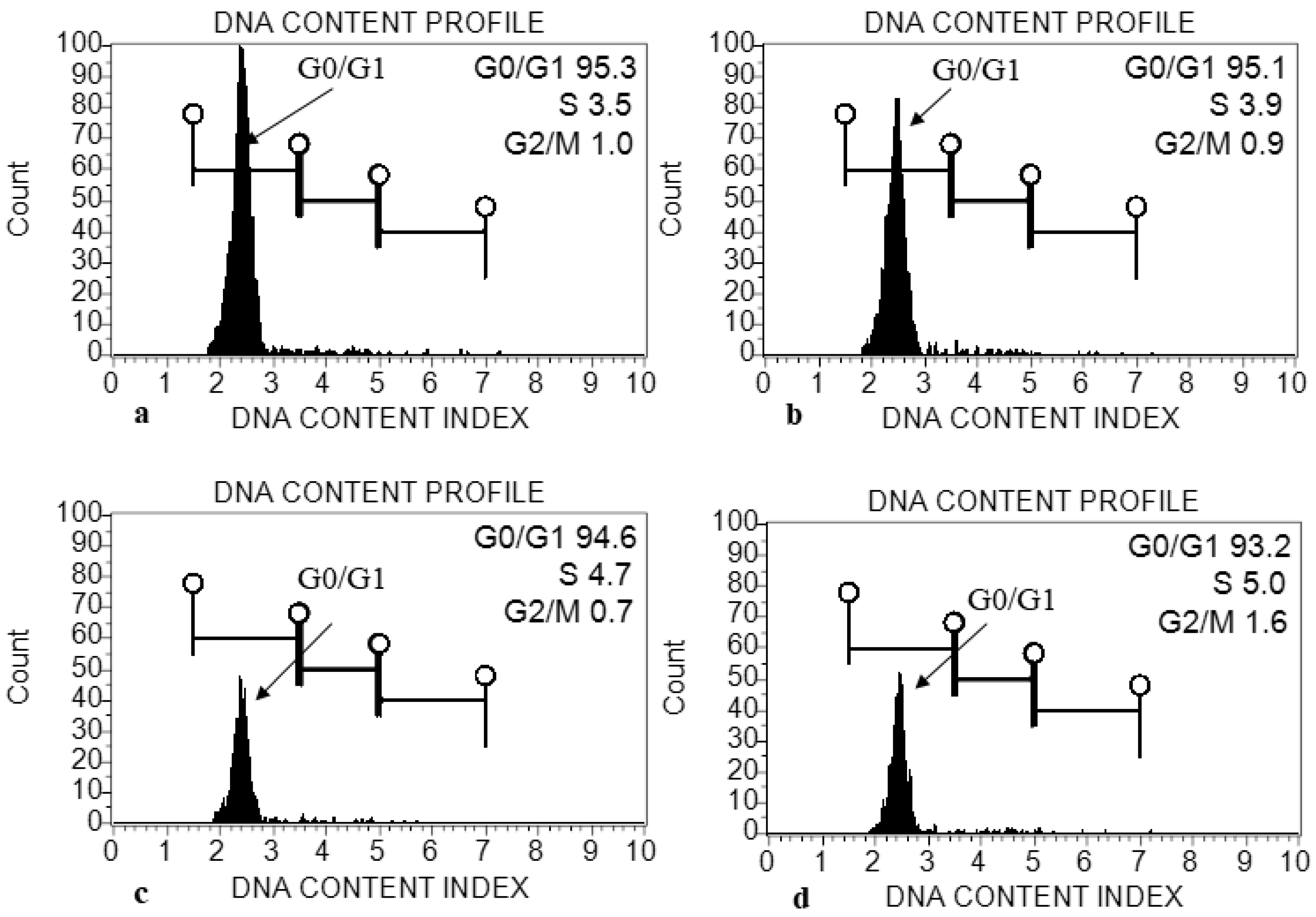
2.1.7. Genome Size Estimation in E. macrochaetus under Different Treatments of ZnO-NPs
2.1.8. Estimation of Bioactive Compound Contents in Different Parts of the Plant under Various Treatments of ZnO-NPs
3. Discussion
4. Materials and Methods
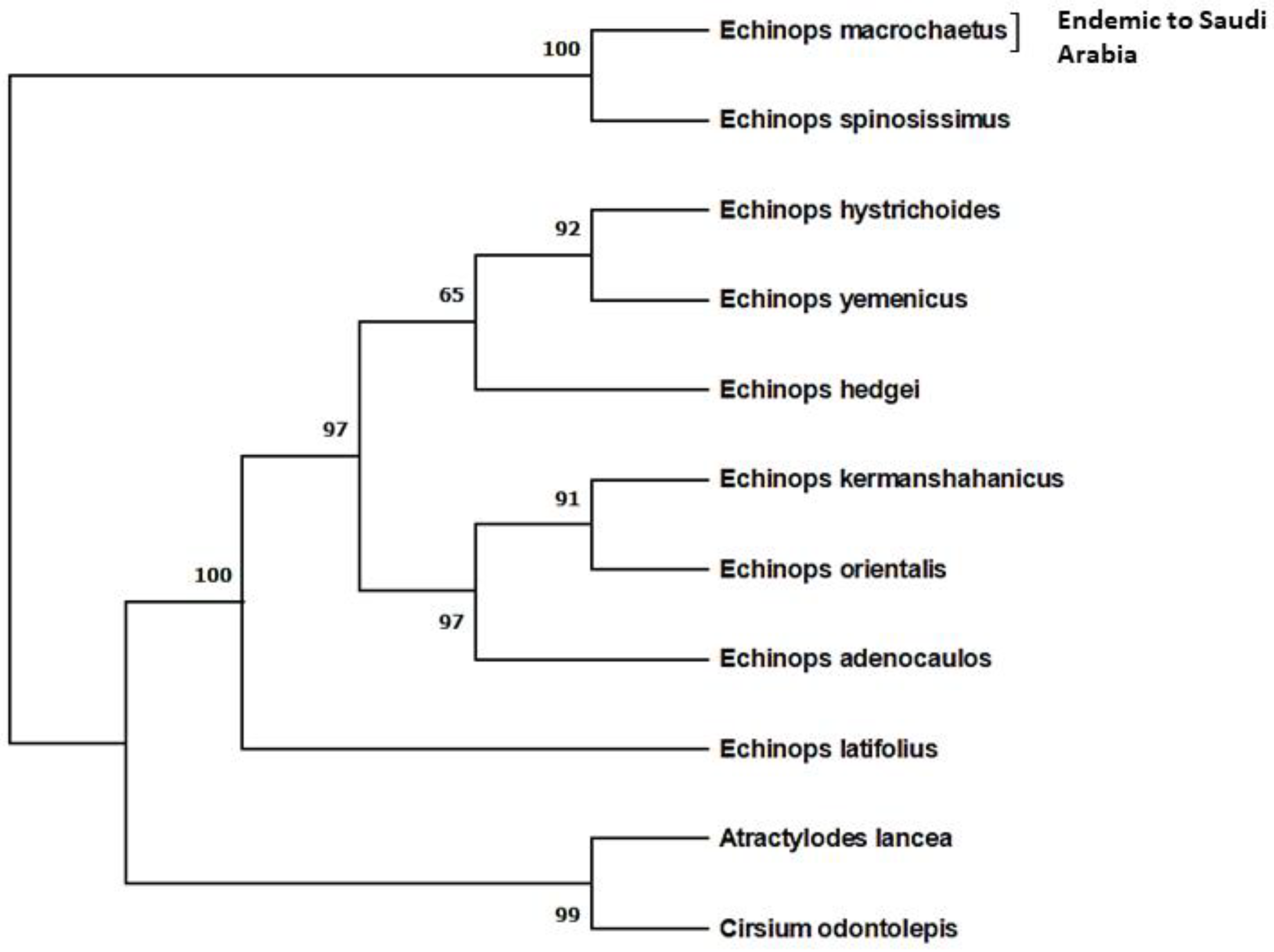
4.1. Plant Collection and Identification
4.2. Isolation of Genomic DNA, PCR and Sequencing
4.3. Aqueous Leaf Extract Preparation for Synthesis of ZnO-NPs
4.4. Natural-Extract-Mediated ZnO-NP Synthesis
4.5. ZnO-NP Characterization
4.6. Seed Sterilization and Pot Experiment
4.7. Treatment of Plant with ZnO-NPs
4.8. Morpho-Physiochemical Analysis
4.8.1. Biomass Determination
4.8.2. Morphological Traits
4.8.3. Determination of Total Chlorophyll and Carotenoids
- Chlorophyll a: Ca = 12.25 A663.2 − 2.79 A646.8 (µg per ml solution).
- Chlorophyll b: Cb = 21.50 A646.8 − 5.10 A663.2 (µg per ml solution).
- Total carotenoids: Cx + c = (1000 A470 − 1.82 Ca − 85.02 Cb)/198 (µg per ml solution).
4.8.4. Proline Estimation
4.8.5. Thiobarbituric Acid Reactive Substances (TBARS)
4.8.6. Total Crude and Soluble Protein Estimation
4.8.7. Superoxide Dismutase (EC 1.15.1.1)
4.8.8. Ascorbate Peroxidase (EC 1.11.1.11)
4.8.9. Glutathione Reductase (EC 1.6.4.2)
4.9. Assessment of Genome Size (2C DNA Content)
4.9.1. Nuclei Extraction
4.9.2. Flow Cytometry Analysis
4.10. Extraction and Estimation of Bioactive Compounds
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Funk, V.A.; Bayer, R.J.; Keeley, S.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panero, J.L.; Baldwin, B.G.; Garcia-Jacas, N. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr. 2005, 55, 343–373. [Google Scholar]
- Bitew, H.; Hymete, A. The genus Echinops: Phytochemistry and biological activities: A review. Front. Pharmacol. 2019, 10, 1234. [Google Scholar] [CrossRef] [PubMed]
- Arroo, R.; Jacobs, J.; Van Gestel, J.; Kenkel, H.; Jannink, W.; Croes, A.; Wullems, G. Regulation of thiophene biosynthesis by sulphate in roots of marigolds. New Phytol. 1997, 135, 175–181. [Google Scholar] [CrossRef]
- Zamzami, T.A.; Abdallah, H.M.; Shehata, I.A.; Mohamed, G.A.; Alfaifi, M.Y.; Elbehairi, S.E.I.; Koshak, A.E.; Ibrahim, S.R. Macrochaetosides A and B, new rare sesquiterpene glycosides from Echinops macrochaetus and their cytotoxic activity. Phytochem. Lett. 2019, 30, 88–92. [Google Scholar] [CrossRef]
- Abegaz, B.M. Polyacetylenic thiophenes and terpenoids from the roots of Echinops pappii. Phytochemistry 1991, 30, 879–881. [Google Scholar] [CrossRef]
- Muir, A.D.; Westcott, N.D. Flax: The Genus Linum; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Zahir, A.; Ahmad, W.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. In vitro cultures of Linum usitatissimum L.: Synergistic effects of mineral nutrients and photoperiod regimes on growth and biosynthesis of lignans and neolignans. J. Photochem. Photobiol. B Biol. 2018, 187, 141–150. [Google Scholar] [CrossRef]
- Pappi, P.; Nikoloudakis, N.; Fanourakis, D.; Zambounis, A.; Delis, C.; Tsaniklidis, G. Differential triggering of the phenylpropanoid biosynthetic pathway key genes transcription upon cold stress and viral infection in tomato leaves. Horticulturae 2021, 7, 448. [Google Scholar] [CrossRef]
- Mocanu, M.-M.; Nagy, P.; Szöllősi, J. Chemoprevention of breast cancer by dietary polyphenols. Molecules 2015, 20, 22578–22620. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Rupasinghe, H. Polyphenols: Multipotent therapeutic agents in neurodegenerative diseases. Oxidative Med. Cell. Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, K.; Pandey, V. Constituents of Echinops niveus. Fitoterapia 1990, 61, 279. [Google Scholar]
- Mahmood, A.A.R.; Khadeem, E.J. Phytochemical investigation of flavonoids glycoside in the Iraqi Echinops heterophyllus (Compositae). Pharm. Glob. 2013, 4, 1–8. [Google Scholar]
- Tene, M.; Tane, P.; Sondengam, B.L.; Connolly, J.D. Lignans from the roots of Echinops giganteus. Phytochemistry 2004, 65, 2101–2105. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Ning, L.; Wan, X.; Peng, Z.; Zhong-jun, M.; Xian, L. Chemical constituents of the root of echinops grijisii Hance. Shenyang Yao Ke Da Xue Xue Bao 2008, 8. [Google Scholar]
- Francois, S.; Celine, D.; Ulugbek, K.; Frederic, M.; Parhat, K.; Akber, A.H.; Corine, G.-T. Chemical constituents from Echinops integrifolius. Biochem. Syst. Ecol. 2013, 47, 42–44. [Google Scholar]
- Pragasam, S.J.; Venkatesan, V.; Rasool, M. Immunomodulatory and anti-inflammatory effect of p-coumaric acid, a common dietary polyphenol on experimental inflammation in rats. Inflammation 2013, 36, 169–176. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Nawaz, M.A.; Rahman, K. Potassium and zinc increase tolerance to salt stress in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2017, 116, 139–149. [Google Scholar] [CrossRef]
- Sheoran, P.; Grewal, S.; Kumari, S.; Goel, S. Enhancement of growth and yield, leaching reduction in Triticum aestivum using biogenic synthesized zinc oxide nanofertilizer. Biocatal. Agric. Biotechnol. 2021, 32, 101938. [Google Scholar] [CrossRef]
- Ruszkiewicz, J.A.; Pinkas, A.; Ferrer, B.; Peres, T.V.; Tsatsakis, A.; Aschner, M. Neurotoxic effect of active ingredients in sunscreen products, a contemporary review. Toxicol. Rep. 2017, 4, 245–259. [Google Scholar] [CrossRef]
- Jafari, S.; Mousavi-Fard, S.; Rezaei Nejad, A.; Mumivand, H.; Sorkheh, K.; Nikoloudakis, N.; Fanourakis, D. Chitosan and Titanium Dioxide Are More Effective in Improving Seed Yield and Quality in Nanoparticle Compared to Non-Structured Form: A Case Study in Five Milk Thistle Ecotypes (Silybum marianum (L.) Gaertn.). Agronomy 2022, 12, 1827. [Google Scholar] [CrossRef]
- Aisida, S.O.; Onwujiobi, C.; Ahmad, I.; Zhao, T.-K.; Maaza, M.; Ezema, F.I. Biogenic synthesis of zinc oxide nanorods for biomedical applications and photodegradation of Rhodamine B. Mater. Today Commun. 2022, 33, 104660. [Google Scholar] [CrossRef]
- Onyedikachi, O.A.; Aisida, S.O.; Agbogu, A.; Rufus, I.; Ahmad, I.; Maaza, M.; Ezema, F.I. Zinc ferrite nanoparticles capped with Gongronema latifolium for moderate hyperthermia applications. Appl. Phys. A 2022, 128, 95. [Google Scholar] [CrossRef]
- Fraceto, L.F.; Grillo, R.; de Medeiros, G.A.; Scognamiglio, V.; Rea, G.; Bartolucci, C. Nanotechnology in agriculture: Which innovation potential does it have? Front. Environ. Sci. 2016, 4, 20. [Google Scholar] [CrossRef]
- Teske, S.S.; Detweiler, C.S. The biomechanisms of metal and metal-oxide nanoparticles’ interactions with cells. Int. J. Environ. Res. Public Health 2015, 12, 1112–1134. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Rezaei Nejad, A.; Mousavi-Fard, S.; Fanourakis, D. Postharvest application of single, multi-walled carbon nanotubes and nanographene oxide improves rose keeping quality. J. Hortic. Sci. Biotechnol. 2022, 97, 346–360. [Google Scholar] [CrossRef]
- Hussain, A.; Ali, S.; Rizwan, M.; ur Rehman, M.Z.; Javed, M.R.; Imran, M.; Chatha, S.A.S.; Nazir, R. Zinc oxide nanoparticles alter the wheat physiological response and reduce the cadmium uptake by plants. Environ. Pollut. 2018, 242, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Faraz, A.; Hayat, S. Effective use of zinc oxide nanoparticles through root dipping on the performance of growth, quality, photosynthesis and antioxidant system in tomato. J. Plant Biochem. Biotechnol. 2020, 29, 553–567. [Google Scholar] [CrossRef]
- Al-Qurainy, F.; Khan, S.; Alansi, S.; Nadeem, M.; Alshameri, A.; Gaafar, A.-R.; Tarroum, M.; Shaikhaldein, H.O.; Salih, A.M.; Alenezi, N.A. Impact of Phytomediated Zinc Oxide Nanoparticles on Growth and Oxidative Stress Response of In Vitro Raised Shoots of Ochradenus arabicus. BioMed Res. Int. 2021, 2021, 6829806. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Zahir, A.; Ahmad, W.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C. Biogenic zinc oxide nanoparticles-enhanced biosynthesis of lignans and neolignans in cell suspension cultures of Linum usitatissimum L. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1367–1373. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Haliloğlu, K.; Türkoğlu, A.; Balpınar, Ö.; Nadaroğlu, H.; Alaylı, A.; Poczai, P. Effects of Zinc, Copper and Iron Oxide Nanoparticles on Induced DNA Methylation, Genomic Instability and LTR Retrotransposon Polymorphism in Wheat (Triticum aestivum L.). Plants 2022, 11, 2193. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Zinc oxide nanoparticles (ZnONPs) as a novel nanofertilizer: Influence on seed yield and antioxidant defense system in soil grown soybean (Glycine max cv. Kowsar). Sci. Total Environ. 2020, 738, 140240. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Das, A.; Singh, S.; Kumar, M.; Kumar, O. Zinc biofortification: A novel strategy for improving human health. J. Exp. Biol. Agric. Sci. 2018, 6, 751–762. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Mousavi-Fard, S.; Rezaei Nejad, A.; Fanourakis, D. Carbon nanotubes in the holding solution stimulate flower opening and prolong vase life in carnation. Chem. Biol. Technol. Agric. 2022, 9, 15. [Google Scholar] [CrossRef]
- Khoshgoftarmanesh, A.H.; Markarian, S. Antioxidant response of wheat to tire rubber ash and ZnO nanoparticles and ionic zinc exposure in nutrient solution culture. Acta Physiol. Plant. 2022, 44, 50. [Google Scholar] [CrossRef]
- Mousavi Kouhi, S.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Comparative phytotoxicity of ZnO nanoparticles, ZnO microparticles, and Zn2+ on rapeseed (Brassica napus L.): Investigating a wide range of concentrations. Toxicol. Environ. Chem. 2014, 96, 861–868. [Google Scholar] [CrossRef]
- Borm, P.; Klaessig, F.C.; Landry, T.D.; Moudgil, B.; Pauluhn, J.; Thomas, K.; Trottier, R.; Wood, S. Research strategies for safety evaluation of nanomaterials, part V: Role of dissolution in biological fate and effects of nanoscale particles. Toxicol. Sci. 2006, 90, 23–32. [Google Scholar] [CrossRef]
- Khan, S.; Al-Qurainy, F.; Nadeem, M.; Tarroum, M. Development of genetic markers for Ochradenus arabicus (Resedaceae), an endemic medicinal plant of Saudi Arabia. Genet. Mol. Res 2012, 11, 1300–1308. [Google Scholar] [CrossRef]
- Khan, S.; Al-Qurainy, F.; Nadeem, M.; Tarroum, M. Selection of chloroplast DNA markers for the development of DNA barcode and reconstruction of phylogeny of senecio asirensis boulos and jri wood. Pak. J. Bot 2013, 45, 703–710. [Google Scholar]
- Al-Qurainy, F.; Khan, S.; Nadeem, M.; Tarroum, M.; Gaafar, A. Selection of DNA barcoding loci for Nepeta deflersiana Schweinf. ex Hedge from chloroplast and nuclear DNA genomes. Genet. Mol. Res 2014, 13, 1144–1151. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, S.; Srivastava, P.K.; Singh, V.P.; Singh, S.; Prasad, S.M.; Singh, P.K.; Dubey, N.K.; Pandey, A.C. Nitric oxide alleviates silver nanoparticles (AgNps)-induced phytotoxicity in Pisum sativum seedlings. Plant Physiol. Biochem. 2017, 110, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.D.; Agarwal, A.; Pradhan, S. Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotoxicol. Environ. Saf. 2018, 161, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.A.; Parveen, A.; Ahmad, L.; Hashem, A. Effects of graphene oxide and zinc oxide nanoparticles on growth, chlorophyll, carotenoids, proline contents and diseases of carrot. Sci. Hortic. 2019, 249, 374–382. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, S.M.; Singh, S. Impact of nano ZnO on metabolic attributes and fluorescence kinetics of rice seedlings. Environ. Nanotechnol. Monit. Manag. 2018, 9, 42–49. [Google Scholar] [CrossRef]
- Salachna, P.; Mizielińska, M.; Płoszaj-Witkowska, B.; Jaszczak, A. Zinc oxide nanoparticles enhanced biomass and zinc content and induced changes in biological properties of red Perilla frutescens. Materials 2021, 14, 6182. [Google Scholar] [CrossRef]
- Zomorrodi, N.; Rezaei Nejad, A.; Mousavi-Fard, S.; Feizi, H.; Tsaniklidis, G.; Fanourakis, D. Potency of titanium dioxide nanoparticles, sodium hydrogen sulfide and salicylic acid in ameliorating the depressive effects of water deficit on periwinkle ornamental quality. Horticulturae 2022, 8, 675. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Chen, S.; Li, Q.; Wang, W.; Hou, C.; Gao, X.; Wang, L.; Wang, S. Zinc oxide nanoparticles affect biomass accumulation and photosynthesis in Arabidopsis. Front. Plant Sci. 2016, 6, 1243. [Google Scholar] [CrossRef]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef]
- García-Gómez, C.; García-Gutiérrez, S.; Obrador, A.; Fernández, M.D. Study of Zn availability, uptake, and effects on earthworms of zinc oxide nanoparticle versus bulk applied to two agricultural soils: Acidic and calcareous. Chemosphere 2020, 239, 124814. [Google Scholar] [CrossRef]
- Fanourakis, D.; Kazakos, F.; Nektarios, P.A. Allometric individual leaf area estimation in chrysanthemum. Agronomy 2021, 11, 795. [Google Scholar] [CrossRef]
- Prasad, T.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.; Sajanlal, P.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Chen, Y.; Zhao, M.; Zhang, W.-H. A novel Medicago truncatula HD-Zip gene, MtHB2, is involved in abiotic stress responses. Environ. Exp. Bot. 2012, 80, 1–9. [Google Scholar] [CrossRef]
- Gehan, M.A.; Greenham, K.; Mockler, T.C.; McClung, C.R. Transcriptional networks—Crops, clocks, and abiotic stress. Curr. Opin. Plant Biol. 2015, 24, 39–46. [Google Scholar] [CrossRef]
- Qu, Y.; Duan, M.; Zhang, Z.; Dong, J.; Wang, T. Overexpression of the Medicago falcata NAC transcription factor MfNAC3 enhances cold tolerance in Medicago truncatula. Environ. Exp. Bot. 2016, 129, 67–76. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global plant stress signaling: Reactive oxygen species at the cross-road. Front. Plant Sci. 2016, 7, 187. [Google Scholar] [CrossRef]
- Sharifi-Rad, R.; Esmaeilzadeh Bahabadi, S.; Samzadeh-Kermani, A.; Gholami, M. The Effect of Non-biological Elicitors on Physiological and Biochemical Properties of Medicinal Plant Momordica charantia L. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 1315–1326. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Hwang, H.M.; Ray, P.C.; Yu, H. Mechanisms of Nanotoxicity: Generation of Reactive Oxygen Species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef]
- García-López, J.I.; Zavala-García, F.; Olivares-Sáenz, E.; Lira-Saldívar, R.H.; Díaz Barriga-Castro, E.; Ruiz-Torres, N.A.; Ramos-Cortez, E.; Vázquez-Alvarado, R.; Niño-Medina, G. Zinc oxide nanoparticles boosts phenolic compounds and antioxidant activity of Capsicum annuum L. during germination. Agronomy 2018, 8, 215. [Google Scholar] [CrossRef]
- Pérez-Labrada, F.; López-Vargas, E.R.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Responses of tomato plants under saline stress to foliar application of copper nanoparticles. Plants 2019, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Montejo, S.D.J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as novel elicitors to improve bioactive compounds in plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Ding, Z.; Wang, H.; Song, L.; Jia, S.; Ma, D. Zinc stress affects ionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 111, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Borovaya, S.; Klykov, A.; Barsukova, E.; Chaikina, E. Study of the Effect of Selective Media with High Doses of Zinc on Regeneration Ability and Rutin Accumulation in Common Buckwheat In Vitro. Plants 2022, 11, 264. [Google Scholar] [CrossRef] [PubMed]
- Karimi, N.; Behbahani, M.; Dini, G.; Razmjou, A. Enhancing the secondary metabolite and anticancer activity of Echinacea purpurea callus extracts by treatment with biosynthesized ZnO nanoparticles. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 045009. [Google Scholar] [CrossRef]
- Kumar, V.; Okem, A.; Moyo, M.; Gruz, J.; Doležal, K.; Van Staden, J. Effect of zinc on the production of phenolic acids and hypoxoside in micropropagated Hypoxis hemerocallidea. Plant Growth Regul. 2019, 89, 19–24. [Google Scholar] [CrossRef]
- Simon, C.; Langlois-Meurinne, M.; Bellvert, F.; Garmier, M.; Didierlaurent, L.; Massoud, K.; Chaouch, S.; Marie, A.; Bodo, B.; Kauffmann, S. The differential spatial distribution of secondary metabolites in Arabidopsis leaves reacting hypersensitively to Pseudomonas syringae pv. tomato is dependent on the oxidative burst. J. Exp. Bot. 2010, 61, 3355–3370. [Google Scholar]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, uptake, and translocation of engineered nanomaterials in vascular plants. Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef]
- Sadhu, A.; Bhadra, S.; Bandyopadhyay, M. Novel nuclei isolation buffer for flow cytometric genome size estimation of Zingiberaceae: A comparison with common isolation buffers. Ann. Bot. 2016, 118, 1057–1070. [Google Scholar]
- Bennetzen, J.L. Transposable elements, gene creation and genome rearrangement in flowering plants. Curr. Opin. Genet. Dev. 2005, 15, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Khan, S.S.; Pakrashi, S.; Mukherjee, A.; Chandrasekaran, N. Cytogenetic and genotoxic effects of zinc oxide nanoparticles on root cells of Allium cepa. J. Hazard. Mater. 2011, 190, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Qureshi, M.I.; Alam, T.; Abdin, M. Protocol for isolation of genomic DNA from dry and fresh roots of medicinal plants suitable for RAPD and restriction digestion. Afr. J. Biotechnol. 2007, 6, 175. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Hanson, A.; Nelsen, C.; Pedersen, A.; Everson, E. Capacity for proline accumulation during water stress in barley and its implications for breeding for drought resistance 1. Crop Sci. 1979, 19, 489–493. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Singh, N.; Jain, N.; Kumar, R.; Jain, A.; Singh, N.K.; Rai, V. A comparative method for protein extraction and 2-D gel electrophoresis from different tissues of Cajanus cajan. Front. Plant Sci. 2015, 6, 606. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Rao, M. Cellular detoxifying mechanisms determine the age dependent injury in tropical trees exposed to SO2. J. Plant Physiol. 1992, 140, 733–740. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DNA Content (pg) per 2C (G0/G1) (Mean ± SD) | DNA Content (pg) per 4C (S Phase) (Mean ± SD) | Mitotic Index (4C/2C) |
|---|---|---|---|
| Control (0 mg/L, ZnO-NPs) | 2.2813 ± 0.01 a | 4.01 ± 0.08 b | 1.761 |
| T1 (10 mg/L, ZnO-NPs) | 2.275778 ± 0.03 a | 3.88 ± 0.08 ab | 1.708 |
| T2 (20 mg/L, ZnO-NPs) | 2.257111 ± 0.05 a | 3.84 ± 0.10 a | 1.695 |
| T3 (40 mg/L, ZnO-NPs) | 2.249022 ± 0.04 a | 3.7 ± 0.08 a | 1.681 |
| Treatment | Quercetin-3-β-D-Glucoside (mg/g DW) | Luteolin 7-Rutinoside (mg/g DW) | p-Coumaric Acid (µg/g DW) |
|---|---|---|---|
| Leaf | |||
| Control | 4.52 ± 0.11 b | 5.99 ± 0.27 c | 270.22 ± 7.91 c |
| T1 | 5.59 ± 0.23 a | 7.01 ± 0.14 b | 313.16 ± 8.94 b |
| T2 | 5.31 ± 0.04 a | 7.53 ± 0.39 a | 348.55 ± 2.13 a |
| T3 | 4.09 ± 0.13 c | 5.63 ± 0.25 d | 216.64 ± 9.90 d |
| Shoot | |||
| Control | 4.55 ± 0.16 a | 4.94 ± 0.27 c | 200.59 ± 9.73 b |
| T1 | 4.27 ± 0.17 a | 6.01 ± 0.12 b | 277.96 ± 11.91 a |
| T2 | 3.32 ± 0.08 c | 6.50 ± 0.10 a | 263.45 ± 6.43 a |
| T3 | 3.99 ± 0.08 b | 4.63 ± 0.25 d | 176.80 ± 7.56 c |
| Root | |||
| Control | 0.88 ± 0.03 a | 1.52 ± 0.11 ab | 65.20 ± 5.91 b |
| T1 | 0.69 ± 0.07 b | 1.40 ± 0.06 ab | 86.67 ± 7.17 a |
| T2 | 0.58 ± 0.04 c | 1.46 ± 0.24 a | 97.11 ± 9.85 a |
| T3 | 0.31 ± 0.05 d | 1.27 ± 0.09 c | 56.50 ± 6.77 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Al-Qurainy, F.; Al-hashimi, A.; Nadeem, M.; Tarroum, M.; Shaikhaldein, H.O.; Salih, A.M. Effect of Green Synthesized ZnO-NPs on Growth, Antioxidant System Response and Bioactive Compound Accumulation in Echinops macrochaetus, a Potential Medicinal Plant, and Assessment of Genome Size (2C DNA Content). Plants 2023, 12, 1669. https://doi.org/10.3390/plants12081669
Khan S, Al-Qurainy F, Al-hashimi A, Nadeem M, Tarroum M, Shaikhaldein HO, Salih AM. Effect of Green Synthesized ZnO-NPs on Growth, Antioxidant System Response and Bioactive Compound Accumulation in Echinops macrochaetus, a Potential Medicinal Plant, and Assessment of Genome Size (2C DNA Content). Plants. 2023; 12(8):1669. https://doi.org/10.3390/plants12081669
Chicago/Turabian StyleKhan, Salim, Fahad Al-Qurainy, Abdulrahman Al-hashimi, Mohammad Nadeem, Mohamed Tarroum, Hassan O. Shaikhaldein, and Abdalrhaman M. Salih. 2023. "Effect of Green Synthesized ZnO-NPs on Growth, Antioxidant System Response and Bioactive Compound Accumulation in Echinops macrochaetus, a Potential Medicinal Plant, and Assessment of Genome Size (2C DNA Content)" Plants 12, no. 8: 1669. https://doi.org/10.3390/plants12081669