1. Introduction
Hypericum foliosum Aiton is a low shrub with large yellow flowers that belongs to the
Hypericaceae family. This species is endemic to the Azores and belongs to the same section (
Androsaemum) as
Hypericum androsaemum, which is also used in Portuguese traditional medicine due to its diuretic, hepatoprotective, and antihypertensive properties [
1]. Locally,
H. foliosum is known as malfurada or furalha [
2].
Currently, the
Hypericaceae family comprises nine genera:
Cratoxylum Blume,
Eliea Cambess,
Harungana Lamarck,
Hypericum L.,
Lianthus N. Robson,
Santomasia N. Robson,
Thornea Breedlove & McClintock,
Triadenum Rafinesque, and
Vismia Vand. Most of the biodiversity within this family is included in the
Hypericum genus (roughly 80%) [
3,
4]. However, this taxonomic classification has not been updated in the Flora Europea or in the Nova Flora de Portugal, in which the
Hypericum genus is still part of the
Clusiaceae (
Guttiferae) [
5].
The
Hypericum genus comprises 484 species spread across all continents except for Antarctica. These species may exist as herbaceous or bushy species—or, rarely, as trees—and they are grouped into 36 taxonomic sections constructed according to specific combinations of morphological characteristics and biogeographic distribution. They are distributed across various habitats, from temperate regions to high mountains in the tropics, avoiding areas with extreme temperature, aridity, or salinity [
6].
Many
Hypericum species have long been used in traditional medicine and had their biological activities demonstrated through pharmacological studies. However, only the flowering part of
H. perforatum is recognized as an herbal drug and described in the Portuguese and European Pharmacopoeias, and it was accepted as such by the European Medicines Agency (EMA) in 2009 [
7,
8]. In the last few years, a vast range of biological activities, such as anti-inflammatory, anticancer, antidiabetic, and antioxidant activities, related to plants from the
Hypericum genus have been described in the published literature [
9,
10,
11,
12].
H. foliosum has long been a subject of study by our team. We previously conducted botanical, chemical, pharmacological, and toxicological studies using dried extracts of the flowering aerial parts of
H. foliosum. The results from our preliminary botanical analysis led to the description of the first anatomical features necessary for identifying this medicinal plant as an herbal drug. Other specimens harvested in different areas and seasons should also be observed to confirm the obtained data. In our chemical and pharmacological work on
H. foliosum, we established the main secondary metabolites’ fingerprint profile with a methanolic extract. Phenolic compound derivatives, including biapigenin, catechin, chlorogenic acid, miquelianin, quercetin, and quinic acid, were identified. Antidepressant activity studies showed that the antidepressant activity of the
H. foliosum methanolic extract was not inferior to those of
H. perforatum and
H. androsaemum in animal models. In addition, evaluation of chronic toxicity in vivo showed no significant impact on the liver, pancreas, kidneys, or lipid profile [
1,
7].
H. foliosum methanolic extract also showed strong radical scavenging and acetylcholinesterase-inhibitory activities [
13,
14].
Previous work by other authors on
H. foliosum led to the identification of an acylphloroglucinol metabolite that demonstrated antimicrobial in vitro activity against
Staphylococcus aureus [
15]. The essential oil composition is known to mainly comprise
n-nonane, limonene, terpinolene, and other terpene derivatives [
2,
7,
13]. Conservation of the species was also a focus of study, with micropropagation methodologies showing good results [
16].
To legitimize the use of
H. foliosum as an herbal drug, it is essential to describe its botanical characteristics and establish a clear and universal quality control methodology through a quality herbal drug monograph, as well as deepen knowledge regarding its biological activity. Several secondary metabolites with diverse biological activities have previously been identified in
Hypericum genus extracts. These compounds generally belong to classes such as acylphloroglucinols, flavonoids, phloroglucinols, and xanthones and exhibit in vitro anticancer, cell protection, anti-inflammatory, antimicrobial, and antidepressant activities [
17]. Since the selective cytotoxicity and antioxidant activities related, respectively, to anticancer and cell protection mechanisms were among the most frequent biological activities described in studies focusing on this botanical genus, we decided to study these activities in
H. foliosum ethanolic, dichloromethane:ethanol, and water extracts using in vitro models. We also aimed to continue our previous work and fully describe the distinctive macroscopic and microscopic characteristics of the raw material of
H. foliosum aerial parts.
3. Discussion
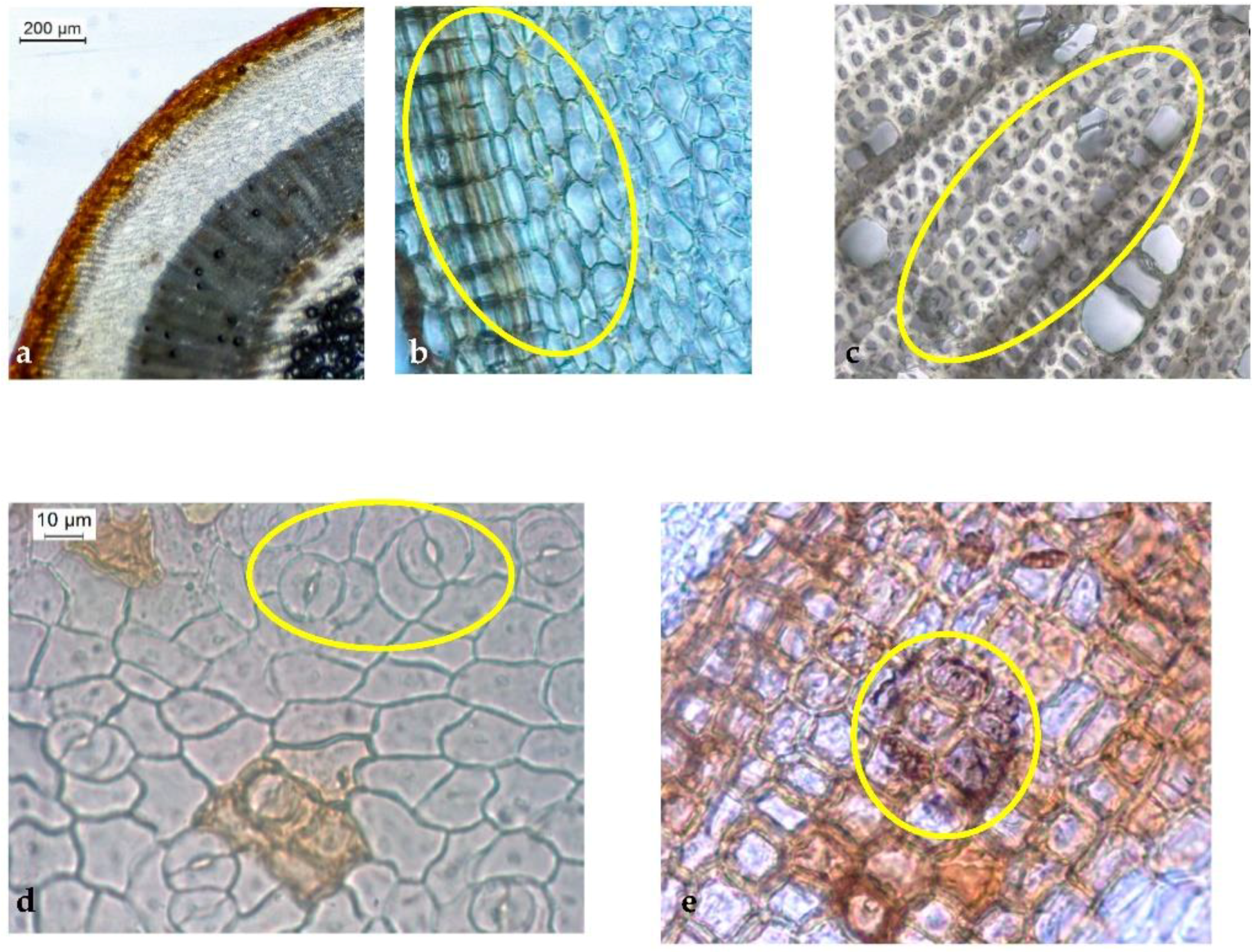
In the present study, we describe the first complete and comprehensive macroscopic and microscopic characterization of herbal medicine produced from H. foliosum aerial part.
Our macroscopic characterization of dried, fragmented raw material from
H. foliosum aerial parts was consistent with the morphological botanical description of the species in the Nova Flora de Portugal by Franco et al. related to the species’ taxonomic identification [
5]. The crenated stem with longitudinal ridges and dark pits; reticulated venation; stipules in the leaf’s basis; the number and color of sepals, petals, and carpels; and the absence of dark glands were the main features used for the species’ initial identification.
Our microscopic analysis revealed distinct and useful characteristics for accurately identifying herbal medicine produced from
H. foliosum aerial parts. For instance, in the transverse section view, we could see the stem with a thick cuticle; the number of collenchymatous layers, type A secretory canal, circularly shaped vascular bundles, and uniseriate medullary rays also contributed to a clear identification of
H. foliosum. With regard to the leaf, the bifacial mesophyll with translucent glands, the open collateral vascular bundle, and the type B secretory canals in the protruded midrib were the most useful characteristics. As anticipated, our results shared several similarities with Serrano et al.’s findings [
21].
To the best of our knowledge, this is the first full description of the stem, leaf, and flower of
H. foliosum. No calcium oxalate crystals were observed. The outlines and proportions of vascular bundles in the stem and leaf are also important characteristics in distinguishing some
Hypericum species. For instance, the xylem of the
Hipericum thymopsis Boiss. fills the major part of the stem, whereas the pith is in the small region of the transverse section’s center [
22], which can be observed easily. Our analysis of
H. foliosum—specifically, the observation of circularly shaped vascular bundles in the stem and an open collateral vascular bundle type in the midrib of the leaf—can be used as important microscopic characteristics to differentiate
Hypericum species and thus to identify
H. foliosum aerial parts properly.
A growing body of literature is focused on the secretory structure of
Hypericum species [
19,
23,
24,
25,
26]. Secretory canals seem to be a common secretory structure in the
Hypericum sp. and play an important role in its anatomy and distinction. Generally, each species is characterized by the presence of different types of secretory structures, including translucent glands, black nodules, and secretory canals with different shapes, ontogenesis routes, and localizations [
25]. In
H. foliosum, the translucent glands were present within the lamina of the leaf, close to the abaxial surface; type A and type B secretory canals were found in the cortical parenchyma of the stem and associated with phloem in the midrib, respectively. These secretory structures and their localization have previously been described in other
Hypericum sp., such as
Hypericum elodes L. [
20],
Hypericum perforatum L. [
16],
Hypericum inodorum Mill.,
Hypericum olympicum L., and
Hypericum forrestii (Chitt.) N. Robson, as well as in other families of Angiosperms [
27].
In addition, even though the representative metabolic naphthodianthrones (such as hypericin) are only present in black nodules [
22], histochemical tests have been used to establish a correlation between the function of secretory structures and the localization of some secondary metabolites [
21]. From a pharmacological point of view, translucent glands and type B canals produce biologically secondary metabolites with potential pharmacological activity (such as alkaloids, lipids, and resins), which may protect the plant against herbivores and parasites. As tannins accumulate in stems [
25], type A canals can act as storage cavities.
Macroscopic analysis of new plant material harvested in 2021 confirmed the botanical characteristics previously observed, validating their use as monographic identification elements in a future quality monograph on H. foliosum aerial parts as an herbal drug.
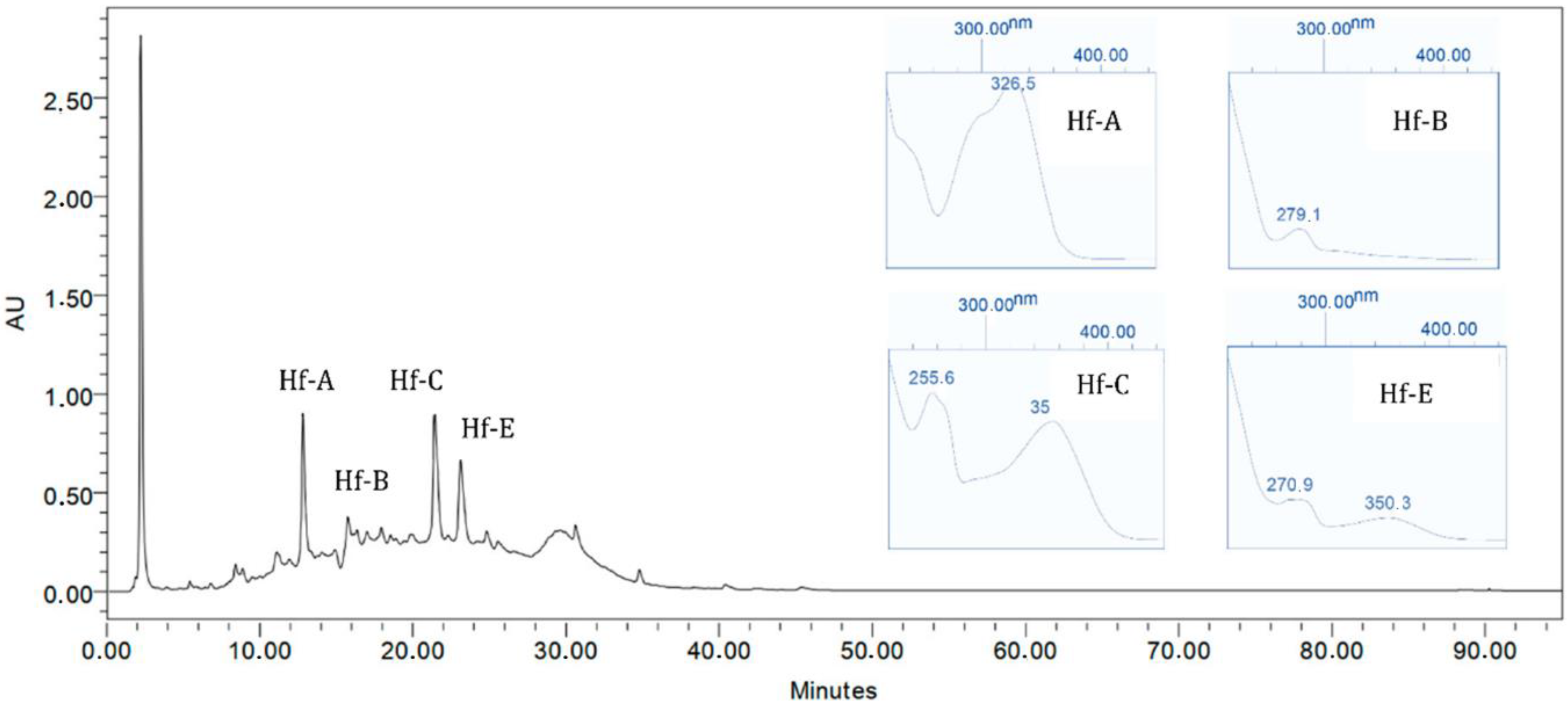
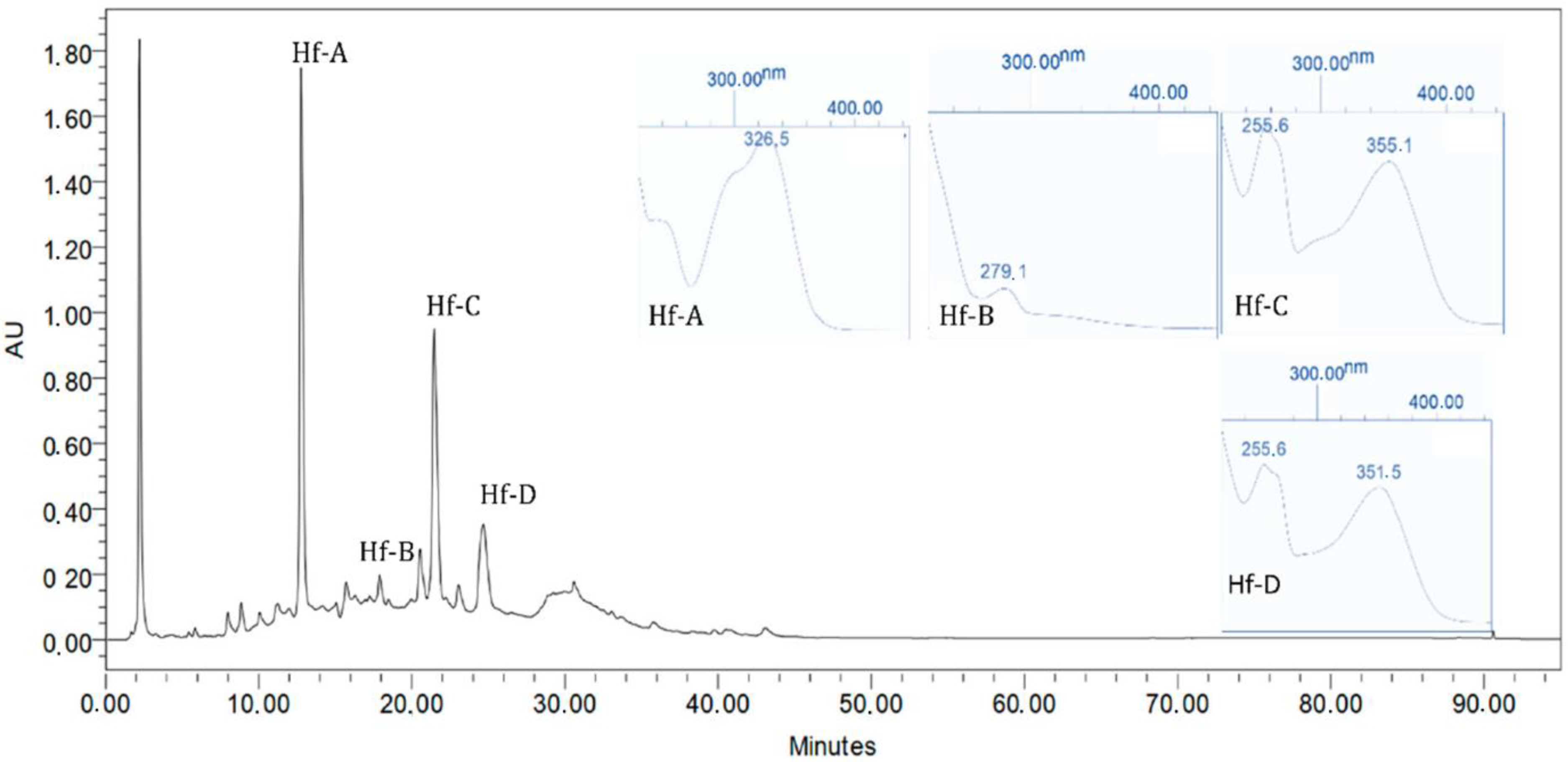
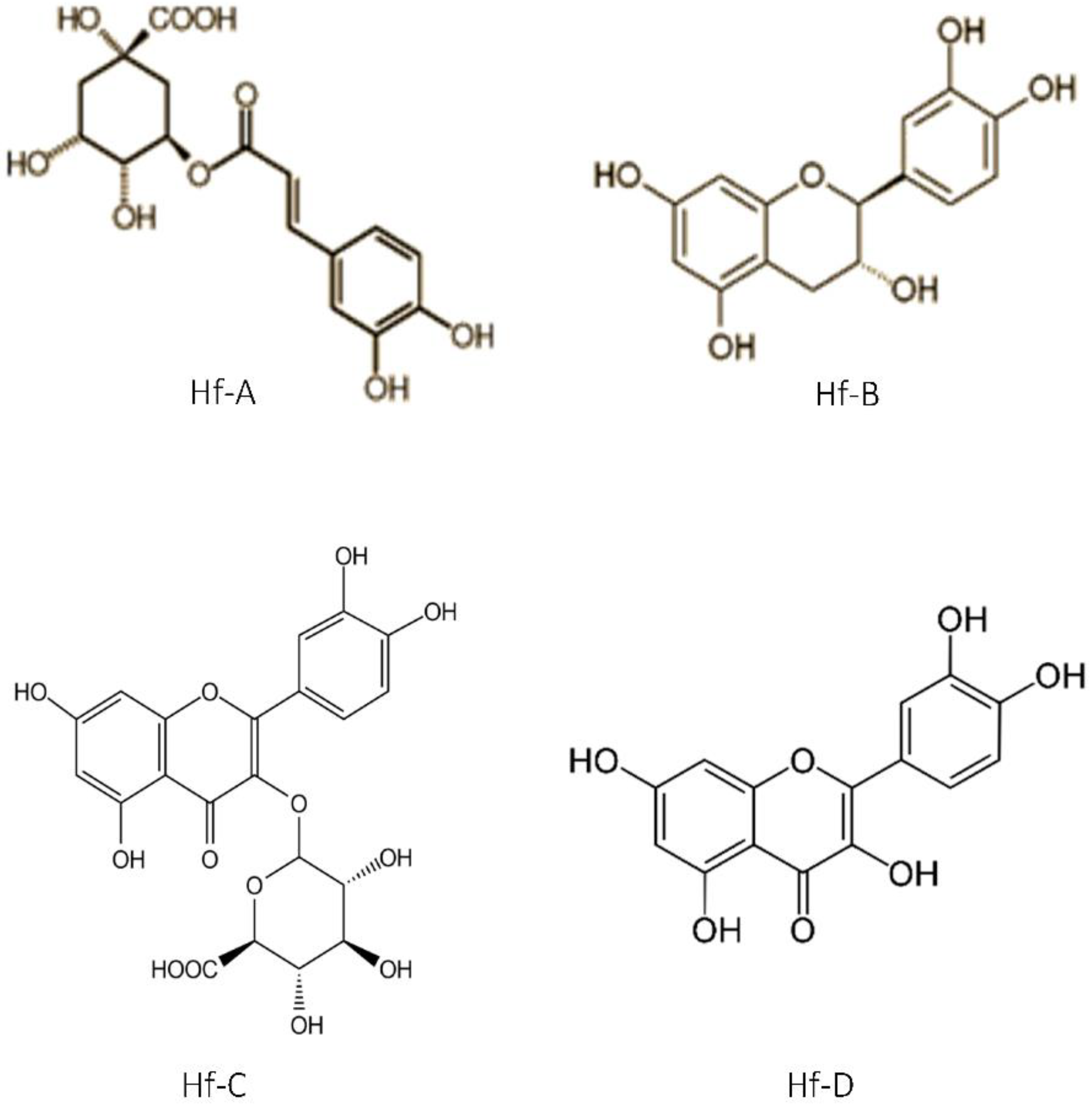
Previous
H. foliosum studies conducted by our team led to the identification of compounds such as quinic acid, catechin, chlorogenic acid, quercetin, miquelianin, and biapigenin in the methanolic extract and the establishment of a correlation between the antioxidant activity and flavonoid composition of the plant extract [
20]. The higher the flavonoid content was, the stronger the antioxidant activity observed. Therefore, considering the antioxidant activity shown in our results, we can infer that the
H. foliosum extracts under analysis in this study had different flavonoid contents, the ethanolic extract being the extract with the higher content, followed by dichloromethane/methanol 1:1.
In this work, Hf. E and HF. DM showed significant anticancer activity against all cancer cell lines tested. Selectively, these extracts showed different levels of activity against MDA-MB-231, A549, and HCT8 cells, which are, respectively, related to breast, lung, and colon cancers. These results suggest a possible efflux of the main Hf. E secondary metabolites (phenol acid derivatives) by P-glycoprotein.
The antioxidant and anticancer activities exhibited by
H. foliosum in this study were in line with the major biological activities exhibited by natural compounds isolated from plants and subject to analysis, such as quercetin and catechin [
17].
Our results are currently being investigated to achieve a better understanding of the cytotoxic activity exhibited by H. foliosum in this study. By establishing a correlation between class compounds and selective anticancer activities, we can contribute to the discovery of new compounds with potential therapeutic applications in oncology.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}