
Antiallergic Effects of Callerya atropurpurea Extract In Vitro and in an In Vivo Atopic Dermatitis Model
 , and
, and 
Abstract
:1. Introduction
2. Results
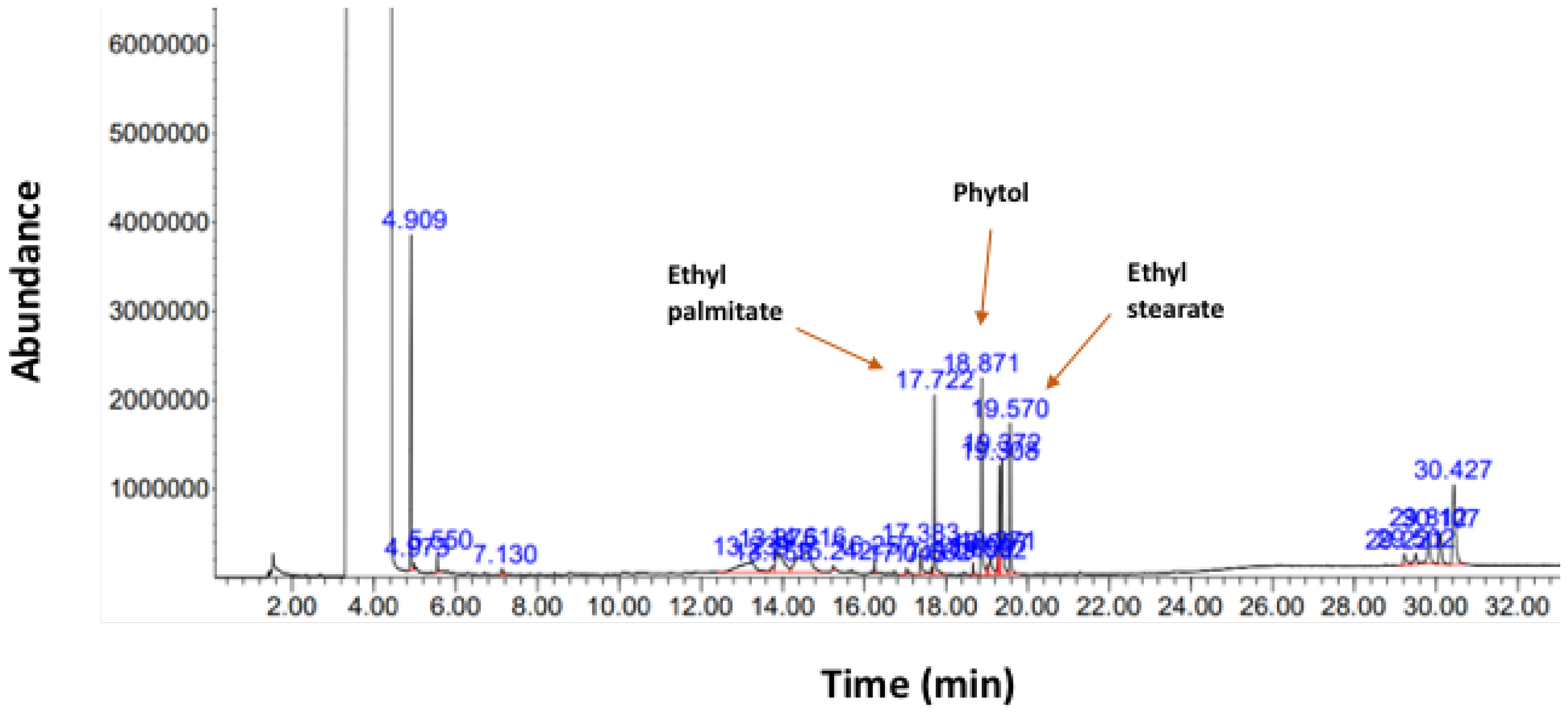
2.1. GC/MS Analysis of Callerya atropurpurea (Ca-EE)
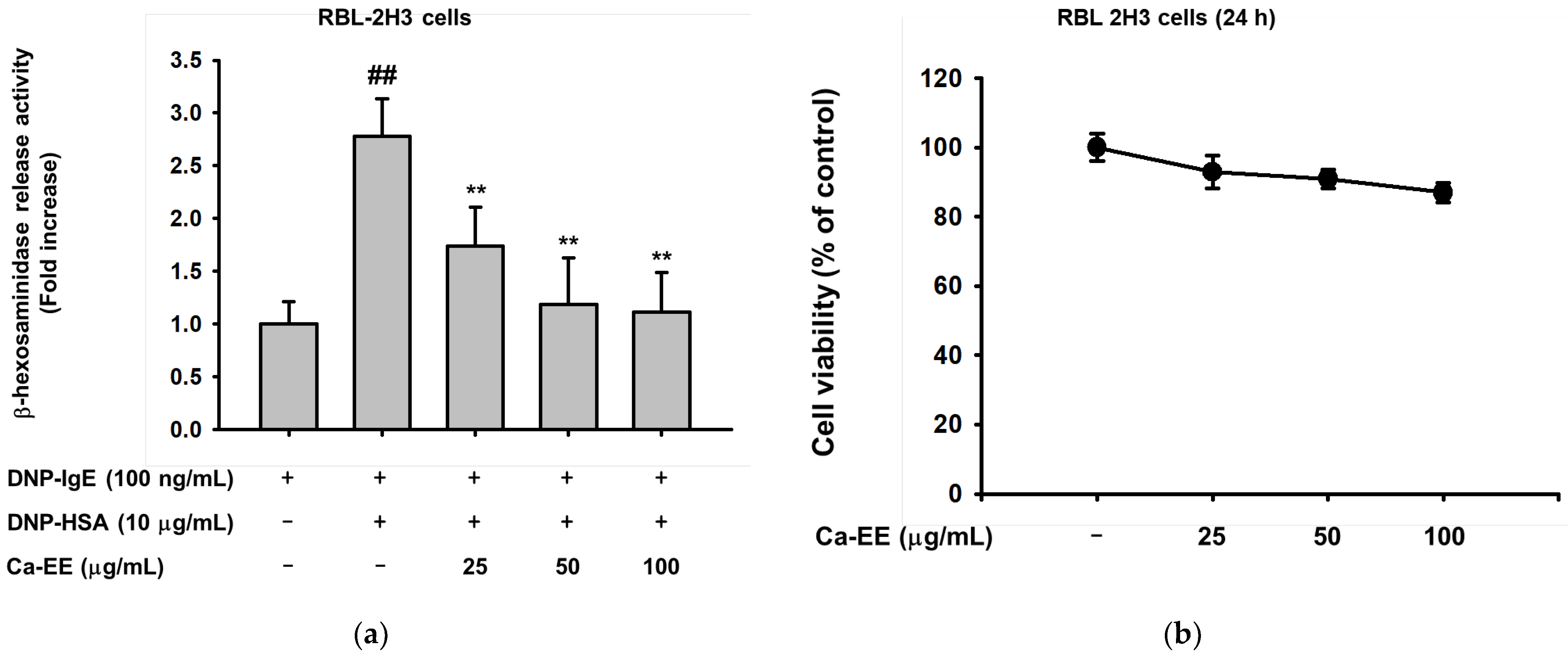
2.2. Antiallergic Activities of Ca-EE in RBL-2H3 Cells
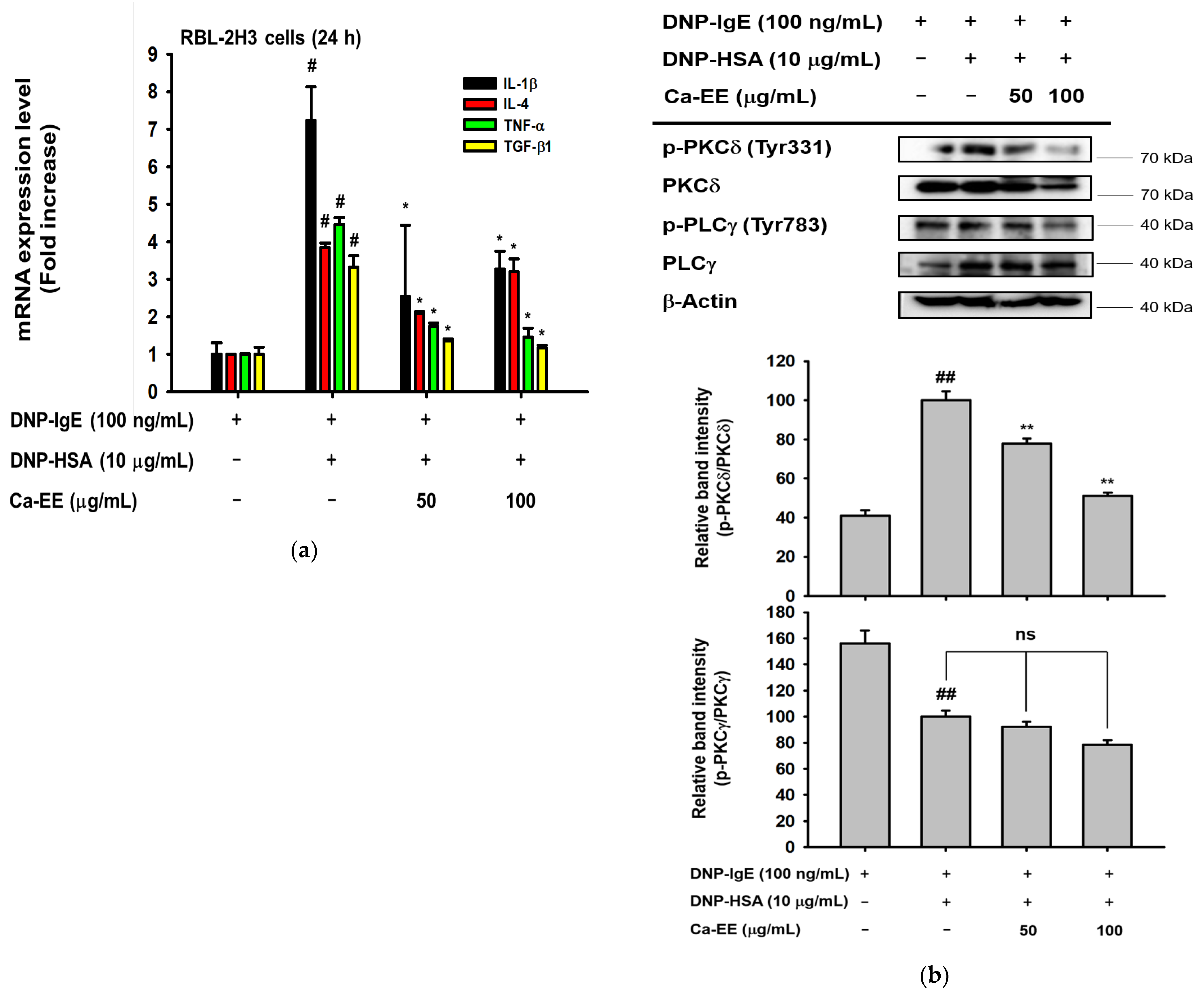
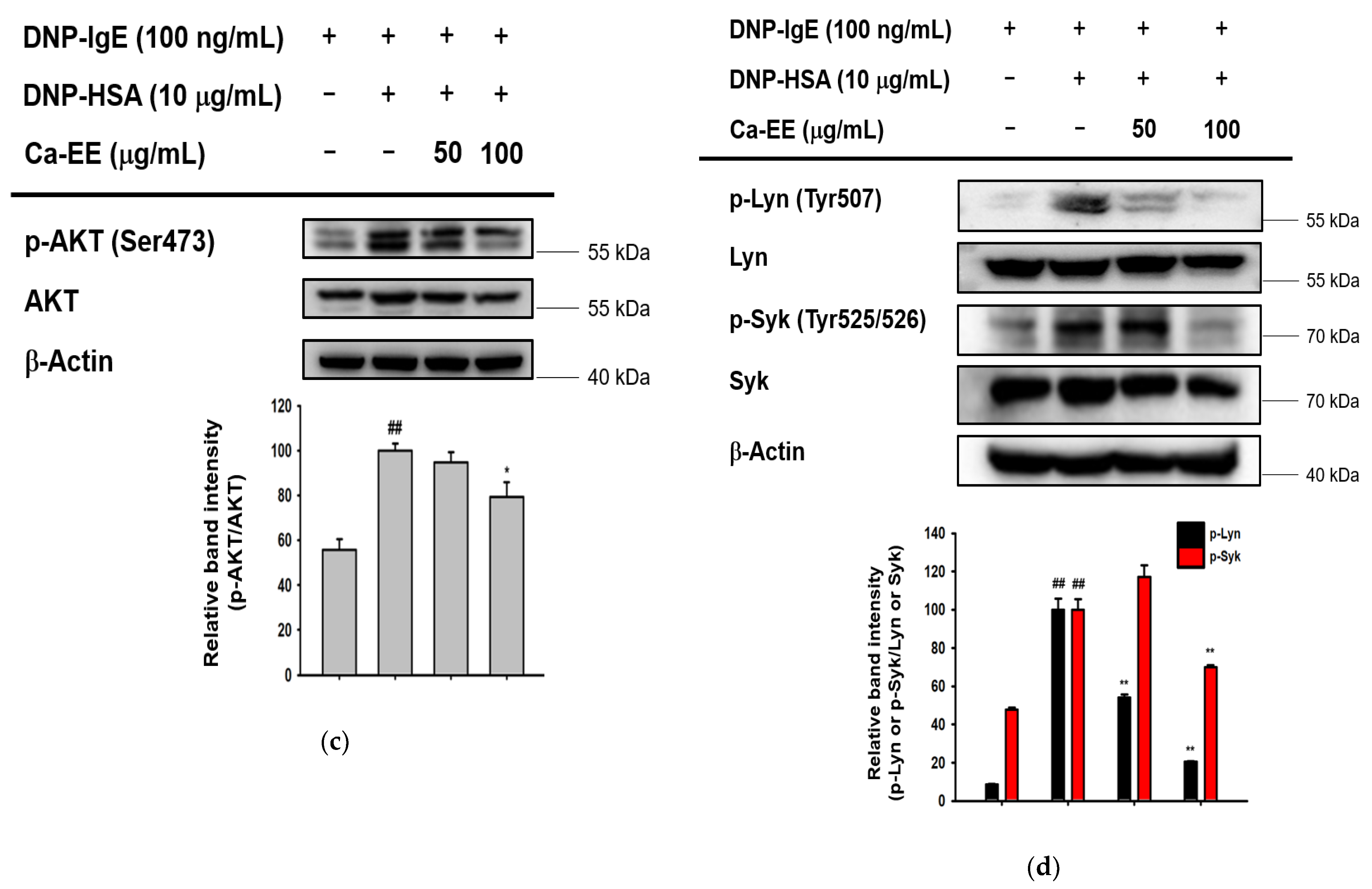
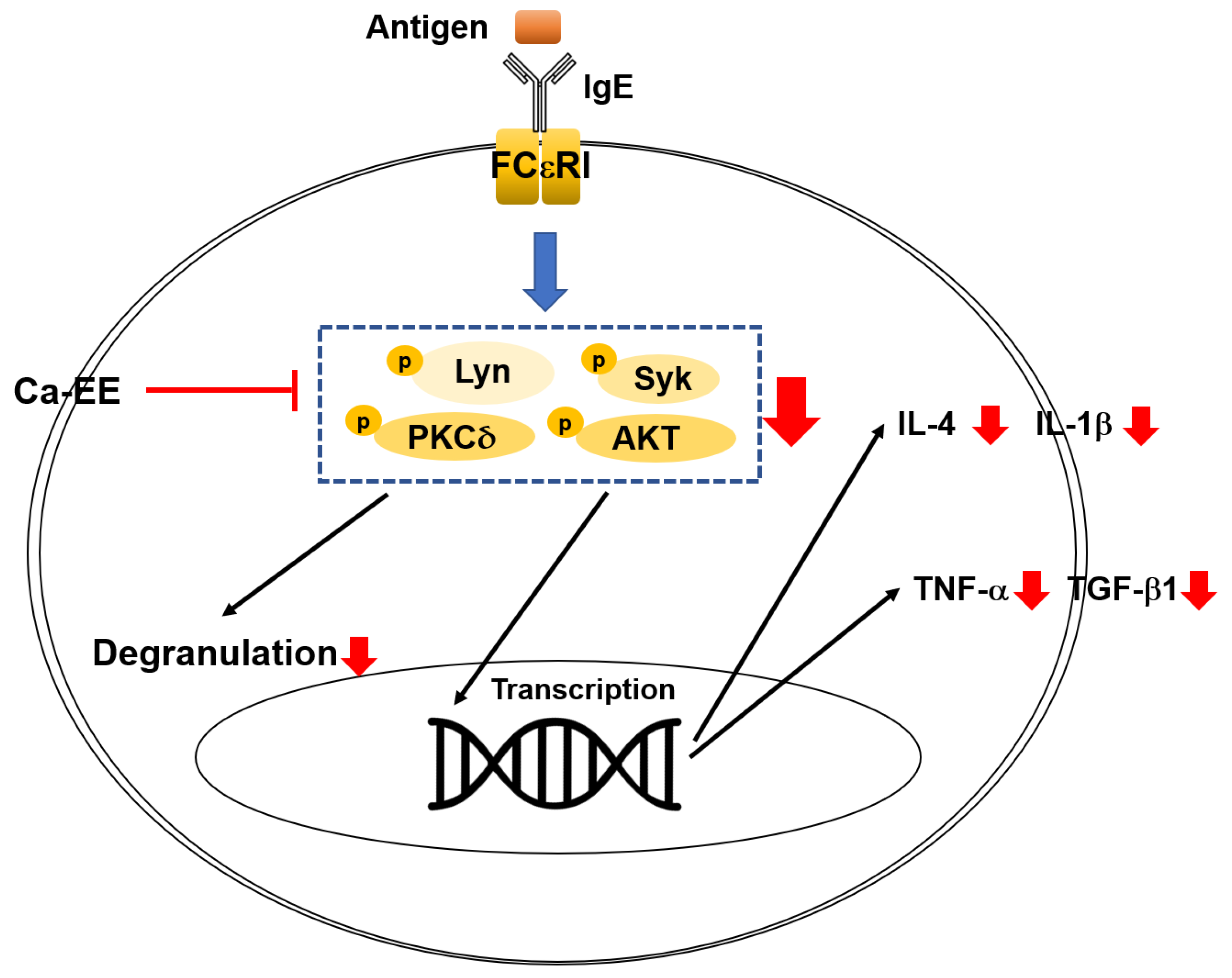
2.3. Inhibitory Action of Ca-EE on Allergic Inflammatory Cytokine Transcription and the IgE-FceRI Signaling Pathway
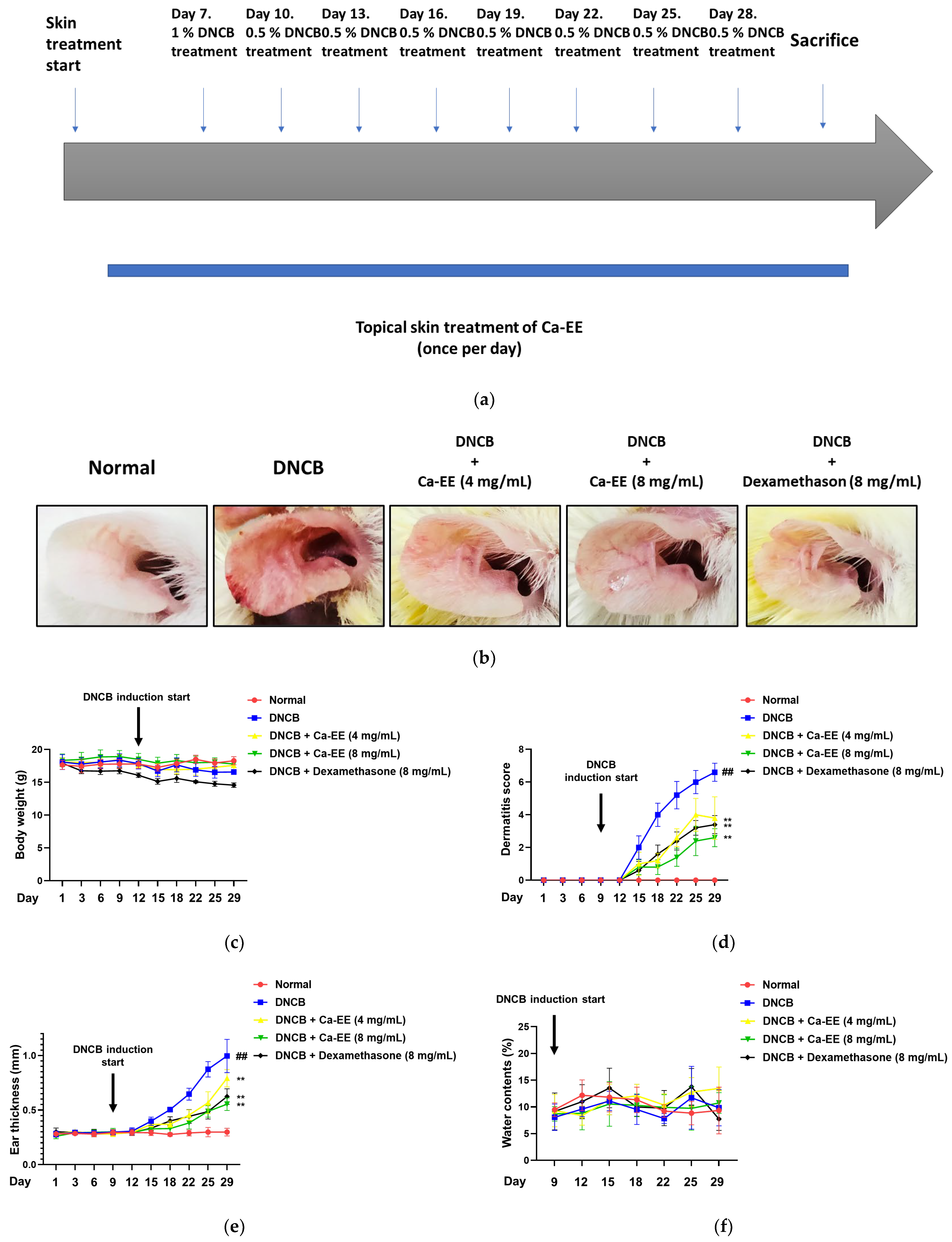
2.4. In Vivo Effects of Ca-EE in the DNCB-Induced Contact Dermatitis Model
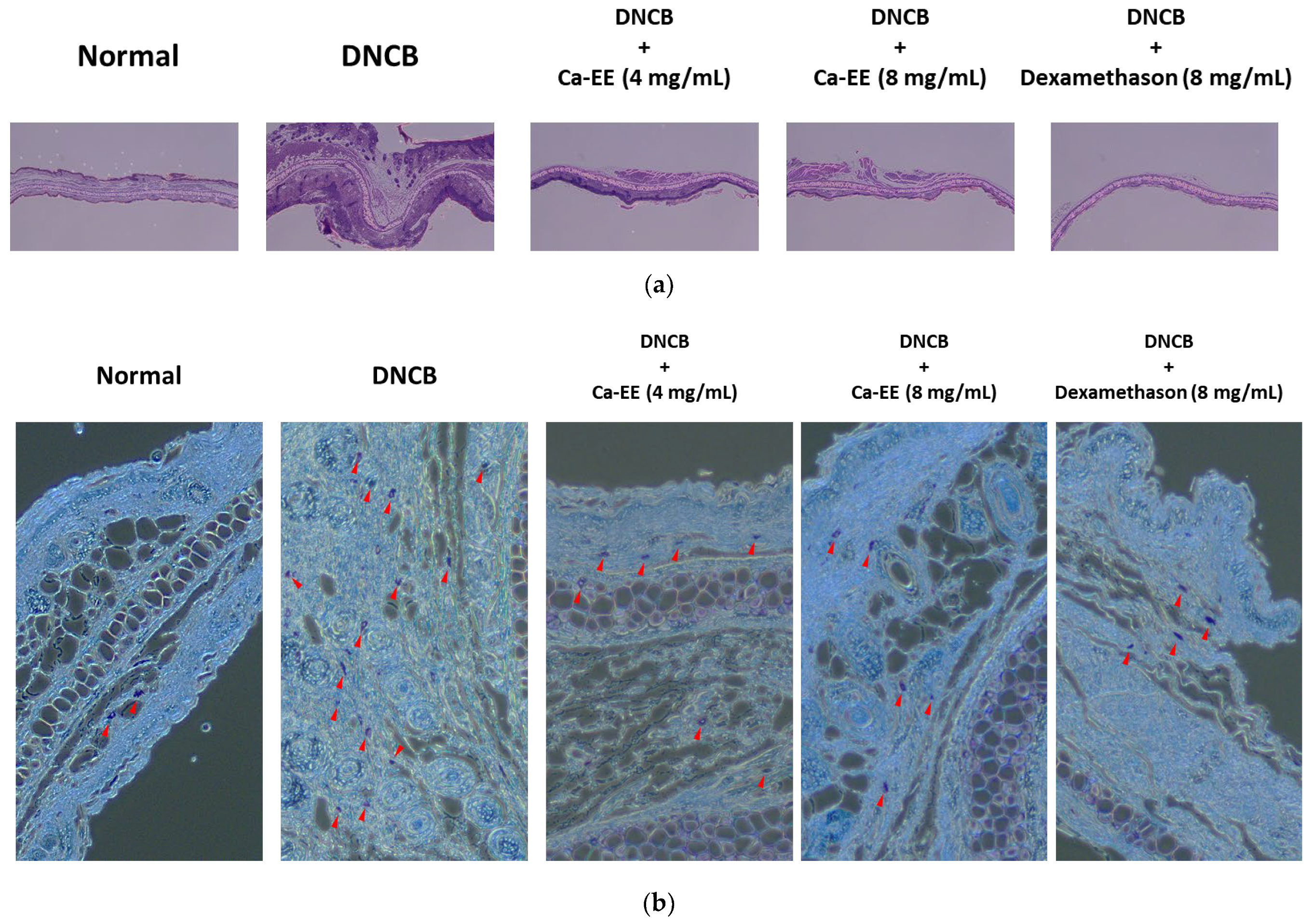
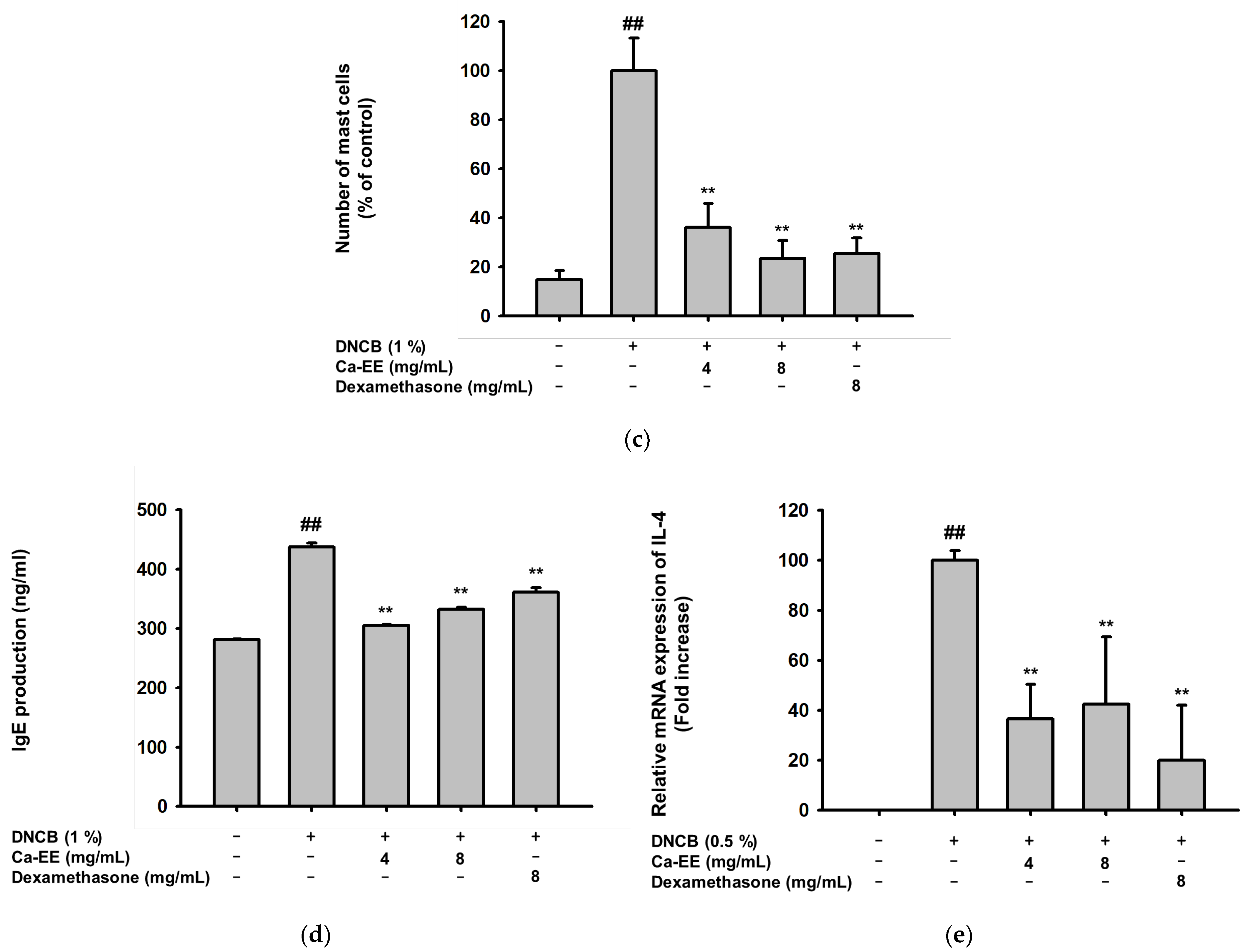
2.5. Histological and Immunological Analysis of Ca-EE-Mediated Antiatopic Effects in the DNCB-Induced Dermatitis Model
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Experiments
4.3. Cell Culture
4.4. Ca-EE Preparation
4.5. Gas Chromatography/Mass Spectrometry
4.6. Cell Viability Assay
4.7. Inhibition of 15-Lipoxygenase Activity Assay
4.8. β-Hexosaminidase Activity Assay
4.9. RNA Extraction and Quantitative Real-Time PCR
4.10. Preparation of Cell Lysates and Immunoblotting Analysis
4.11. DNCB-Induced Atopic Dermatitis Mouse Model
4.12. Measurements of Water Content
4.13. Histopathological Analysis
4.14. Quantification of IgE Level in Mouse Serum
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| AP-1 | Activator protein 1 |
| BSA | Bovine serum albumin |
| DNCB | 2,4-Dinitrochlorobenzene |
| DNP | Dinitrophenol |
| Ca-EE | Callerya atropurpurea ethanol extract |
| FceRI | High-affinity IgE receptor |
| GC/MS | Gas chromatography/mass spectrometry |
| HAS | Human serum albumin |
| H&E | Hematoxylin and eosin |
| IgE | Immunoglobulin E |
| IL-4 | Interleukin 4 |
| ITAM | Immunoreceptor tyrosine-based activation motif |
| LOX | Lipoxygenase |
| LT | Leukotriene |
| MAPK | Mitogen-activated protein kinase |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B cells |
| PCA | Passive cutaneous anaphylaxis |
| PI3K | Phosphatidylinositol 3-kinase |
| PKCδ | Protein kinase Cδ |
| PLCγ | Phospholipase Cγ |
| Syk | Spleen-associated tyrosine kinase |
| TGF-β | Transforming growth factor beta |
| TLR4 | Toll-like receptor 4 |
| TNF-α | Tumor necrosis factor alpha |
| Th2 | Type 2 helper T |
References
- Abbas, M.; Moussa, M.; Akel, H. Type I Hypersensitivity Reaction; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Wu, C.-Y.; Huang, H.-Y.; Pan, W.-C.; Liao, S.-L.; Hua, M.-C.; Tsai, M.-H.; Lai, S.-H.; Yeh, K.-W.; Chen, L.-C.; Huang, J.-L. Allergic diseases attributable to atopy in a population sample of Asian children. Sci. Rep. 2021, 11, 16052. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Cho, J.Y. The regulatory role of Korean ginseng in skin cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ellmeier, W.; Abramova, A.; Schebesta, A. Tec family kinases: Regulation of FcεRI-mediated mast-cell activation. FEBS J. 2011, 278, 1990–2000. [Google Scholar] [CrossRef]
- Tong, P.; Wesemann, D.R. Molecular mechanisms of IgE class switch recombination. IgE Antibodies Gener. Funct. 2015, 388, 21–37. [Google Scholar]
- Lee, J.-H.; Mun, S.-H.; Ko, N.-Y.; Kim, J.-W.; Kim, D.-K.; Kim, J.-D.; Her, E.; Choi, W.-S. Roles of Src-family kinase isoforms, Lyn, Fyn, Fgr, and c-Src on degranulation in RBL-2H3 mast cells. J. Life Sci. 2007, 17, 350–355. [Google Scholar] [CrossRef]
- Krause, K.; Metz, M.; Makris, M.; Zuberbier, T.; Maurer, M. The role of interleukin-1 in allergy-related disorders. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 477–484. [Google Scholar] [CrossRef]
- Nakai, Y.; Ohashi, Y.; Kakinoki, Y.; Tanaka, A.; Washio, Y.; Nasako, Y.; Masamoto, T.; Sakamoto, H.; Ohmoto, Y. Allergen-induced mRNA expression of IL-5, but not of IL-4 and IFN-γ, in peripheral blood mononuclear cells is a key feature of clinical manifestation of seasonal allergic rhinitis. Arch. Otorhinolaryngol. Head Neck Surg. 2000, 126, 992–996. [Google Scholar] [CrossRef]
- Ryan, J.J. Interleukin-4 and its receptor: Essential mediators of the allergic response. J. Allergy Clin. Immunol. 1997, 99, 1–5. [Google Scholar] [CrossRef]
- Tirado-Rodriguez, B.; Ortega, E.; Segura-Medina, P.; Huerta-Yepez, S. TGF-β: An important mediator of allergic disease and a molecule with dual activity in cancer development. J. Immunol. Res. 2014, 2014, 318481. [Google Scholar] [CrossRef]
- Smirnova, M.G.; Birchall, J.P.; Pearson, J.P. The immunoregulatory and allergy-associated cytokines in the aetiology of the otitis media with effusion. Mediat. Inflamm 2004, 13, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Kato, A. Group 2 Innate Lymphoid Cells in Airway Diseases. Chest 2019, 156, 141–149. [Google Scholar] [CrossRef]
- Bachert, C. The role of histamine in allergic disease: Re-appraisal of its inflammatory potential. Allergy 2002, 57, 287–296. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The role of histamine and histamine receptors in mast cell-mediated allergy and inflammation: The hunt for new therapeutic targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef]
- Eldanasory, O.A.; Eljaaly, K.; Memish, Z.A.; Al-Tawfiq, J.A. Histamine release theory and roles of antihistamine in the treatment of cytokines storm of COVID-19. Travel Med. Infect. Dis. 2020, 37, 101874. [Google Scholar] [CrossRef]
- Guo, R.H.; Park, J.U.; Jo, S.J.; Ahn, J.H.; Park, J.H.; Yang, J.Y.; Lee, S.S.; Park, M.J.; Kim, Y.R. Anti-allergic inflammatory effects of the essential oil from fruits of Zanthoxylum coreanum Nakai. Front. Pharmacol. 2018, 9, 1441. [Google Scholar] [CrossRef]
- Marone, G.; Sven, H.; Lichtenstein, L.M. An inhibitor of lipoxygenase inhibits histamine release from human basophils. Clin. Immunol. Immunopathol. 1980, 17, 117–122. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.R.; Kang, H.; Choi, J.; Yang, H.; Lee, P.; Kim, J.; Lee, K.W. 7, 8, 4′-Trihydroxyisoflavone attenuates DNCB-induced atopic dermatitis-like symptoms in NC/Nga mice. PLoS ONE 2014, 9, e104938. [Google Scholar] [CrossRef]
- Lee, J.O.; Hwang, S.H.; Shen, T.; Kim, J.H.; You, L.; Hu, W.; Cho, J.Y. Enhancement of skin barrier and hydration-related molecules by protopanaxatriol in human keratinocytes. J. Ginseng Res. 2021, 45, 354–360. [Google Scholar] [CrossRef]
- Ratan, Z.A.; Haidere, M.F.; Hong, Y.H.; Park, S.H.; Lee, J.O.; Lee, J.; Cho, J.Y. Pharmacological potential of ginseng and its major component ginsenosides. J. Ginseng Res. 2021, 45, 199–210. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis: An expanding therapeutic pipeline for a complex disease. Nat. Rev. Drug Discov. 2022, 21, 21–40. [Google Scholar] [CrossRef]
- Baeck, M.; Marot, L.; Nicolas, J.F.; Pilette, C.; Tennstedt, D.; Goossens, A. Allergic hypersensitivity to topical and systemic corticosteroids: A review. Allergy 2009, 64, 978–994. [Google Scholar] [CrossRef] [PubMed]
- Saag, K.G.; Koehnke, R.; Caldwell, J.R.; Brasington, R.; Burmeister, L.F.; Zimmerman, B.; Kohler, J.A.; Furst, D.E. Low dose long-term corticosteroid therapy in rheumatoid arthritis: An analysis of serious adverse events. Am. J. Med. 1994, 96, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.A.; Jacobs, J.W.; Kirwan, J.R.; Boers, M.; Saag, K.G.; Inês, L.B.; de Koning, E.J.; Buttgereit, F.; Cutolo, M.; Capell, H. Safety of low dose glucocorticoid treatment in rheumatoid arthritis: Published evidence and prospective trial data. Ann. Rheum. Dis. 2006, 65, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Hao, L.; Zhu, S.; Yang, X.; Shi, W.; Zheng, K.; Wang, T.; Chen, H. Potential adverse effects of dexamethasone therapy on COVID-19 patients: Review and recommendations. Infect. Dis. Ther. 2021, 10, 1907–1931. [Google Scholar] [CrossRef]
- Nagai, H.; Teramachi, H.; Tuchiya, T. Recent advances in the development of anti-allergic drugs. Allergol. Int. 2006, 55, 35–42. [Google Scholar] [CrossRef]
- Lim, S.; Jeong, I.; Cho, J.; Shin, C.; Kim, K.-I.; Shim, B.-S.; Ko, S.-G.; Kim, B. The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery. Antioxidants 2021, 10, 1524. [Google Scholar] [CrossRef]
- Hu, J.-m.; Lavin, M.; Wojciechowski, M.F.; Sanderson, M.J. Phylogenetic analysis of nuclear ribosomal ITS/5.8 S sequences in the tribe Millettieae (Fabaceae): Poecilanthe-Cyclolobium, the core Millettieae, and the Callerya group. Syst. Bot. 2002, 27, 722–733. [Google Scholar]
- Schot, A. A Revision of Callerya Endl.(including Padbruggea and Whitfordiodendron)(Papilionaceae: Millettieae). Blumea Biodivers. Evol. Biogeogr. Plants 1994, 39, 1–40. [Google Scholar]
- You, L.; Huang, L.; Jang, J.; Hong, Y.H.; Kim, H.G.; Chen, H.; Shin, C.Y.; Yoon, J.H.; Manilack, P.; Sounyvong, B. Callerya atropurpurea suppresses inflammation in vitro and ameliorates gastric injury as well as septic shock in vivo via TLR4/MyD88-dependent cascade. Phytomedicine 2022, 105, 154338. [Google Scholar] [CrossRef]
- Kitao, M.; Yoneda, R.; Tobita, H.; Matsumoto, Y.; Maruyama, Y.; Arifin, A.; Azani, A.M.; Muhamad, M.N. Susceptibility to photoinhibition in seedlings of six tropical fruit tree species native to Malaysia following transplantation to a degraded land. Trees 2006, 20, 601–610. [Google Scholar] [CrossRef]
- Duchesne, M.; Okoye, I.; Lacy, P. Epithelial cell alarmin cytokines: Frontline mediators of the asthma inflammatory response. Front. Immunol. 2022, 13, 975914. [Google Scholar] [CrossRef]
- Poto, R.; Gambardella, A.R.; Marone, G.; Schroeder, J.T.; Mattei, F.; Schiavoni, G.; Varricchi, G. Basophils from allergy to cancer. Front. Immunol. 2022, 13, 1056838. [Google Scholar] [CrossRef]
- Mihály, J.; Gericke, J.; Törőcsik, D.; Gáspár, K.; Szegedi, A.; Rühl, R. Reduced lipoxygenase and cyclooxygenase mediated signaling in PBMC of atopic dermatitis patients. Prostaglandins Other Lipid Mediat. 2013, 107, 35–42. [Google Scholar] [CrossRef]
- Kim, H.; Mani, I.; Iversen, L.; Ziboh, V. Effects of naturally-occurring flavonoids and biflavonoids on epidermal cyclooxygenase and lipoxygenase from guinea-pigs. Prostaglandins Leukot. Essent. Fat. Acids 1998, 58, 17–24. [Google Scholar] [CrossRef]
- Lee, H.P.; Choi, W.; Kwon, K.W.; You, L.; Rahmawati, L.; Luong, V.D.; Kim, W.; Lee, B.H.; Lee, S.; Kim, J.H.; et al. Inhibitory Effects of Grewia tomentosa Juss. on IgE-Mediated Allergic Reaction and DNCB-Induced Atopic Dermatitis. Plants 2022, 11, 2540. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Hattori, K.; Nishikawa, M.; Watcharanurak, K.; Ikoma, A.; Kabashima, K.; Toyota, H.; Takahashi, Y.; Takahashi, R.; Watanabe, Y.; Takakura, Y. Sustained exogenous expression of therapeutic levels of IFN-γ ameliorates atopic dermatitis in NC/Nga mice via Th1 polarization. J. Immunol. 2010, 184, 2729–2735. [Google Scholar] [CrossRef]
- Vasconcelos, J.F.; Teixeira, M.M.; Barbosa-Filho, J.M.; Lucio, A.S.; Almeida, J.R.; de Queiroz, L.P.; Ribeiro-Dos-Santos, R.; Soares, M.B. The triterpenoid lupeol attenuates allergic airway inflammation in a murine model. Int. Immunopharmacol. 2008, 8, 1216–1221. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra Shill, M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, Y.; Li, C.; Gao, J.; Wang, X.; An, H. Alpha-linolenic acid inhibits IgE-mediated anaphylaxis by inhibiting Lyn kinase and suppressing mast cell activation. Int. Immunopharmacol. 2022, 103, 108449. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed]
- León, B.; Ballesteros-Tato, A. Modulating Th2 cell immunity for the treatment of asthma. Front. Immunol. 2021, 12, 637948. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, A.; Russo, M. Dual role of toll-like receptors in human and experimental asthma models. Front. Immunol. 2018, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Z.; Wang, G.; Xu, S. The Effects of Saposhnikovia divaricata Aqueous Extracts on the Inflammation and Intestinal Microflora in Allergic Rhinitis Mice. Evid. Regarding Complement. Altern. Med. 2022, 2022, 1052359. [Google Scholar] [CrossRef]
- Tian, B.; Ma, X.; Jiang, R. Daphnetin mitigates ovalbumin-induced allergic rhinitis in mice by regulating Nrf2/HO-1 and TLR4/NF-kB signaling. Am. J. Rhinol. Allergy 2022, 37, 19–25. [Google Scholar] [CrossRef]
- Kwon, Y.; Cho, S.-Y.; Kwon, J.; Hwang, M.; Hwang, H.; Kang, Y.J.; Lee, H.-S.; Kim, J.; Kim, W.K. Anti-atopic dermatitis effects of Parasenecio auriculatus via simultaneous inhibition of multiple inflammatory pathways. BMB Rep. 2022, 55, 275–280. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, Y.; Zhao, Y.; Li, X.; Zhang, Y.; Liu, Z. The role of the tyrosine kinase Lyn in allergy and cancer. Mol. Immunol. 2021, 131, 121–126. [Google Scholar] [CrossRef]
- Ding, Y.; Li, C.; Zhang, Y.; Ma, P.; Zhao, T.; Che, D.; Cao, J.; Wang, J.; Liu, R.; Zhang, T. Quercetin as a Lyn kinase inhibitor inhibits IgE-mediated allergic conjunctivitis. Food Chem. Toxicol. 2020, 135, 110924. [Google Scholar] [CrossRef]
- Lee, D.; Park, Y.H.; Lee, J.E.; Kim, H.S.; Min, K.Y.; Jo, M.G.; Kim, H.S.; Choi, W.S.; Kim, Y.M. Dasatinib inhibits Lyn and Fyn Src-family kinases in mast cells to suppress type I hypersensitivity in mice. Biomol. Ther. 2020, 28, 456. [Google Scholar] [CrossRef]
- Rivera, J.; Olivera, A. A current understanding of Fc epsilon RI-dependent mast cell activation. Curr. Allergy Asthma Rep. 2008, 8, 14–20. [Google Scholar] [CrossRef]
- Xu, C.; Li, L.; Wang, C.; Jiang, J.; Li, L.; Zhu, L.; Jin, S.; Jin, Z.; Lee, J.J.; Li, G.; et al. Effects of G-Rh2 on mast cell-mediated anaphylaxis via AKT-Nrf2/NF-kappaB and MAPK-Nrf2/NF-kappaB pathways. J. Ginseng Res. 2022, 46, 550–560. [Google Scholar] [CrossRef]
- Skowron, P.; Turner, J.; McNaughton, G. The use of corticosteroid (dexamethasone) in the treatment of acute laryngotracheitis. Can. Med. Assoc. J. 1966, 94, 528. [Google Scholar]
- Yasir, M.; Goyal, A.; Bansal, P.; Sonthalia, S. Corticosteroid Adverse Effects; StatPearls: Tampa, FL, USA, 2018. [Google Scholar]
- Jutel, M.; Kosowska, A.; Smolinska, S. Allergen immunotherapy: Past, present, and future. Allergy Asthma Immunol. Res. 2016, 8, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-S.; Wang, G.-Y.; Song, H.-B.; Lei, Y.-H.; Wang, H.-T. Advances in targeted drugs for allergic diseases. Chin. Med. J. 2021, 134, 2006–2008. [Google Scholar] [CrossRef]
- Wu, S.; Pang, Y.; He, Y.; Zhang, X.; Peng, L.; Guo, J.; Zeng, J. A comprehensive review of natural products against atopic dermatitis: Flavonoids, alkaloids, terpenes, glycosides and other compounds. Biomed. Pharmacother. 2021, 140, 111741. [Google Scholar] [CrossRef]
- Kow, A.S.F.; Khoo, L.W.; Tan, J.W.; Abas, F.; Lee, M.-T.; Israf, D.A.; Shaari, K.; Tham, C.L. Clinacanthus nutans aqueous leaves extract exerts anti-allergic activity in preclinical anaphylactic models via alternative IgG pathway. J. Ethnopharmacol. 2022, 303, 116003. [Google Scholar] [CrossRef]
- Larsen, J.N.; Broge, L.; Jacobi, H. Allergy immunotherapy: The future of allergy treatment. Drug Discov. Today 2016, 21, 26–37. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Kim, J.W.; Kwon, K.W.; Kim, M.Y.; Cho, J.Y. Potentilla paradoxa Nutt. Ethanol Extract Exhibits Anti-Inflammatory Effects by Suppression of the Src/NF-kappaB Signaling Pathway. Plants 2022, 11, 1750. [Google Scholar] [CrossRef]
- Jang, W.Y.; Lee, H.P.; Kim, S.A.; Huang, L.; Yoon, J.H.; Shin, C.Y.; Mitra, A.; Kim, H.G.; Cho, J.Y. Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition. Plants 2022, 11, 1306. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Shin, K.K.; Kim, H.; Hong, Y.H.; Choi, W.; Kwak, Y.S.; Han, C.K.; Hyun, S.H.; Cho, J.Y. Korean Red Ginseng exerts anti-inflammatory and autophagy-promoting activities in aged mice. J. Ginseng Res. 2021, 45, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Aziz, N.; Park, J.G.; Lee, D.; Kim, J.K.; Kim, S.A.; Choi, W.; Lee, C.Y.; Lee, H.P.; Huyen Trang, H.T.; et al. The EEF1AKMT3/MAP2K7/TP53 axis suppresses tumor invasiveness and metastasis in gastric cancer. Cancer Lett. 2022, 544, 215803. [Google Scholar] [CrossRef] [PubMed]
- Yasin, M.; Shahid, W.; Ashraf, M.; Saleem, M.; Muzaffar, S.; Aziz-ur-Rehman; Ejaz, S.A.; Saeed, A.; Majer, T.; Bhattarai, K. 4-Chlorophenyl-N-furfuryl-1, 2, 4-triazole methylacetamides as significant 15-lipoxygenase inhibitors: An efficient approach for finding lead anti-inflammatory compounds. ACS Omega 2022, 7, 19721–19734. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Name of Compound | % of Total |
|---|---|---|
| 1 | Methanesulfonyl fluoride | 0.275 |
| 2 | Succinic acid, di(3-phenylprop-2-en-1-yl) ester | 0.752 |
| 3 | Oxalic acid, 4-chlorophenyl octadecyl ester | 0.569 |
| 4 | Xylitol | 7.592 |
| 5 | d-Mannose | 0.802 |
| 6 | Trehalose | 6.904 |
| 7 | cis-Raphasatin | 12.673 |
| 8 | 1-Hydroxy-1-(4-methoxyphenyl)propan-2-one | 0.396 |
| 9 | 2-Pentadecanone, 6,10,14-trimethyl | 0.625 |
| 10 | Octahydropyrrolo[1,2-a]pyrazine | 0.351 |
| 11 | n-Hexadecanoic acid | 2.047 |
| 12 | Ethyl 9-hexadecenoate | 0.356 |
| 13 | Ethyl tridecanoate | 8.812 |
| 14 | Heptadecanoic acid, ethyl ester | 0.536 |
| 15 | Phytol | 10.508 |
| 16 | Linoelaidic acid | 0.635 |
| 17 | 9,12,15-Octadecatrienoic acid, (Z,Z,Z)- | 2.334 |
| 18 | Octadecanoic acid | 0.953 |
| 19 | Linoleic acid ethyl ester | 4.979 |
| 20 | 9,12,15-Octadecatrienoic acid, ethyl ester, (Z,Z,Z)- | 9.755 |
| 21 | Octadecanoic acid, ethyl ester | 7.126 |
| 22 | Brallobarbital | 1.655 |
| 23 | Cyclohexane-1,3-dione, 2-allylaminomethylene-5,5-dimethyl- | 1.437 |
| 24 | beta.-Amyrin | 3.852 |
| 25 | Olean-12-ene | 3.501 |
| 26 | Lupeol | 10.578 |
| Samples (μg/mL) | Inhibition of Lipoxygenase (%) | |
|---|---|---|
| Ca-EE | 50 | 40.1 ± 1.5 |
| 100 | 43.9 ± 3.8 | |
| Quercetin | 10 | 34.8 ± 3.0 |
| Gene | Species | Direction | Sequence |
|---|---|---|---|
| IL-4 | Rat | F | GTACCAGACGTCCTTACGGC |
| R | ATTCACGGTGCAGCTTCTCA | ||
| TNF-a | Rat | F | GGCTTTCGGAACTCACTGGA |
| R | GGGAACAGTCTGGGAAGCTC | ||
| IL-1β | Rat | F | TTGAGTCTGCACAGTTCCCC |
| R | TCCTGGGGAAGGCATTAGGA | ||
| TGF-β1 | Rat | F | AGGGCTACCATGCCAACTTC |
| R | CCACGTAGTAGACGATGGGC | ||
| IL-4 | Mouse | F | ATGGATGTGCCAAACGTCCT |
| R | AAGCCCGAAAGAGTCTCTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.; Lee, H.P.; Manilack, P.; Saysavanh, V.; Lee, B.-H.; Lee, S.; Kim, E.; Cho, J.Y. Antiallergic Effects of Callerya atropurpurea Extract In Vitro and in an In Vivo Atopic Dermatitis Model. Plants 2023, 12, 860. https://doi.org/10.3390/plants12040860
Choi W, Lee HP, Manilack P, Saysavanh V, Lee B-H, Lee S, Kim E, Cho JY. Antiallergic Effects of Callerya atropurpurea Extract In Vitro and in an In Vivo Atopic Dermatitis Model. Plants. 2023; 12(4):860. https://doi.org/10.3390/plants12040860
Chicago/Turabian StyleChoi, Wooram, Hwa Pyoung Lee, Philaxay Manilack, Veosavanh Saysavanh, Byoung-Hee Lee, Sarah Lee, Eunji Kim, and Jae Youl Cho. 2023. "Antiallergic Effects of Callerya atropurpurea Extract In Vitro and in an In Vivo Atopic Dermatitis Model" Plants 12, no. 4: 860. https://doi.org/10.3390/plants12040860