
Genetic Variation of Zinc and Iron Concentration in Normal, Provitamin A and Quality Protein Maize under Stress and Non-Stress Conditions

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Soil Chemistry
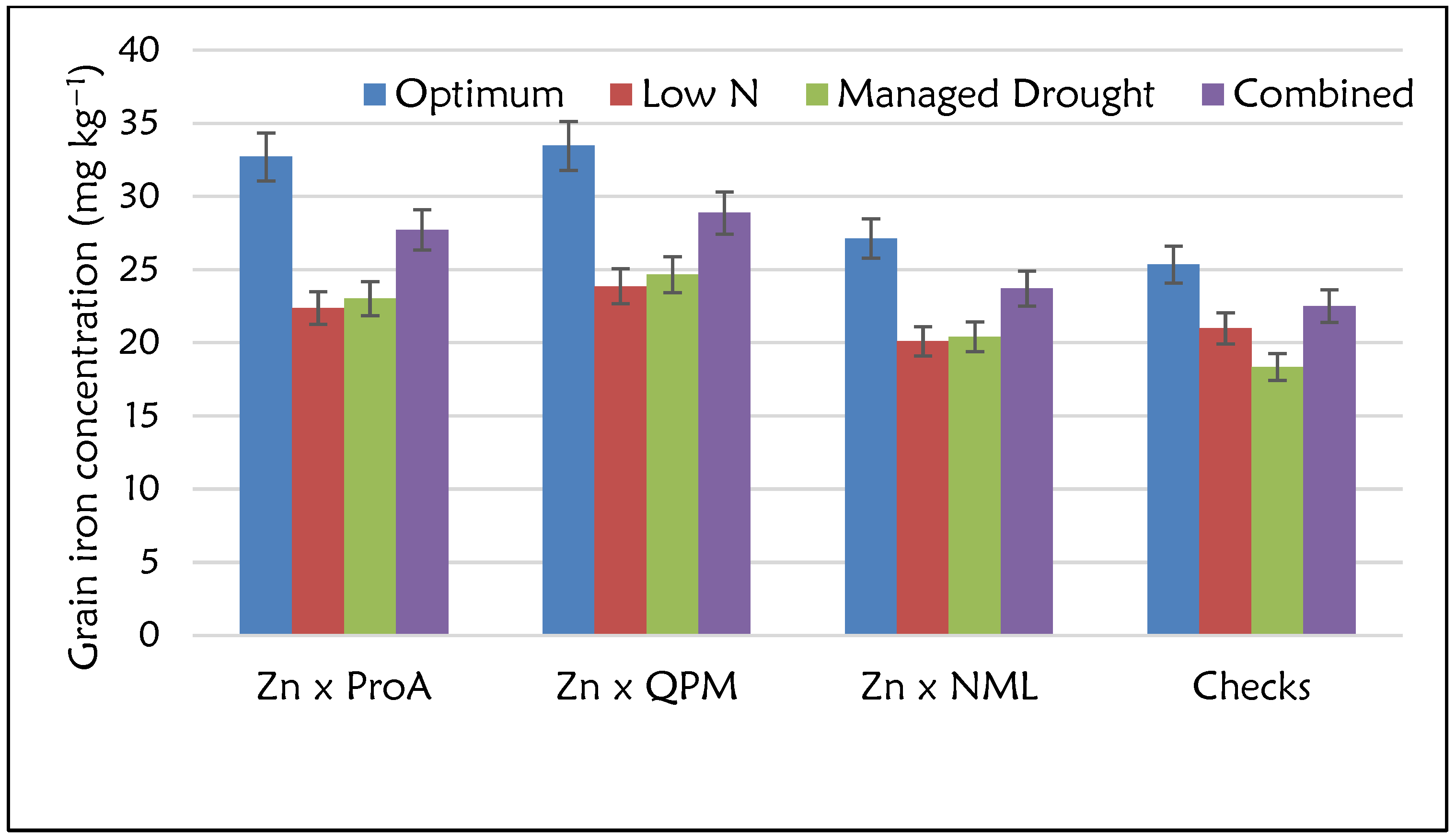
2.2. Agronomic and Micronutrient Performance of Zn-Enhanced Hybrids
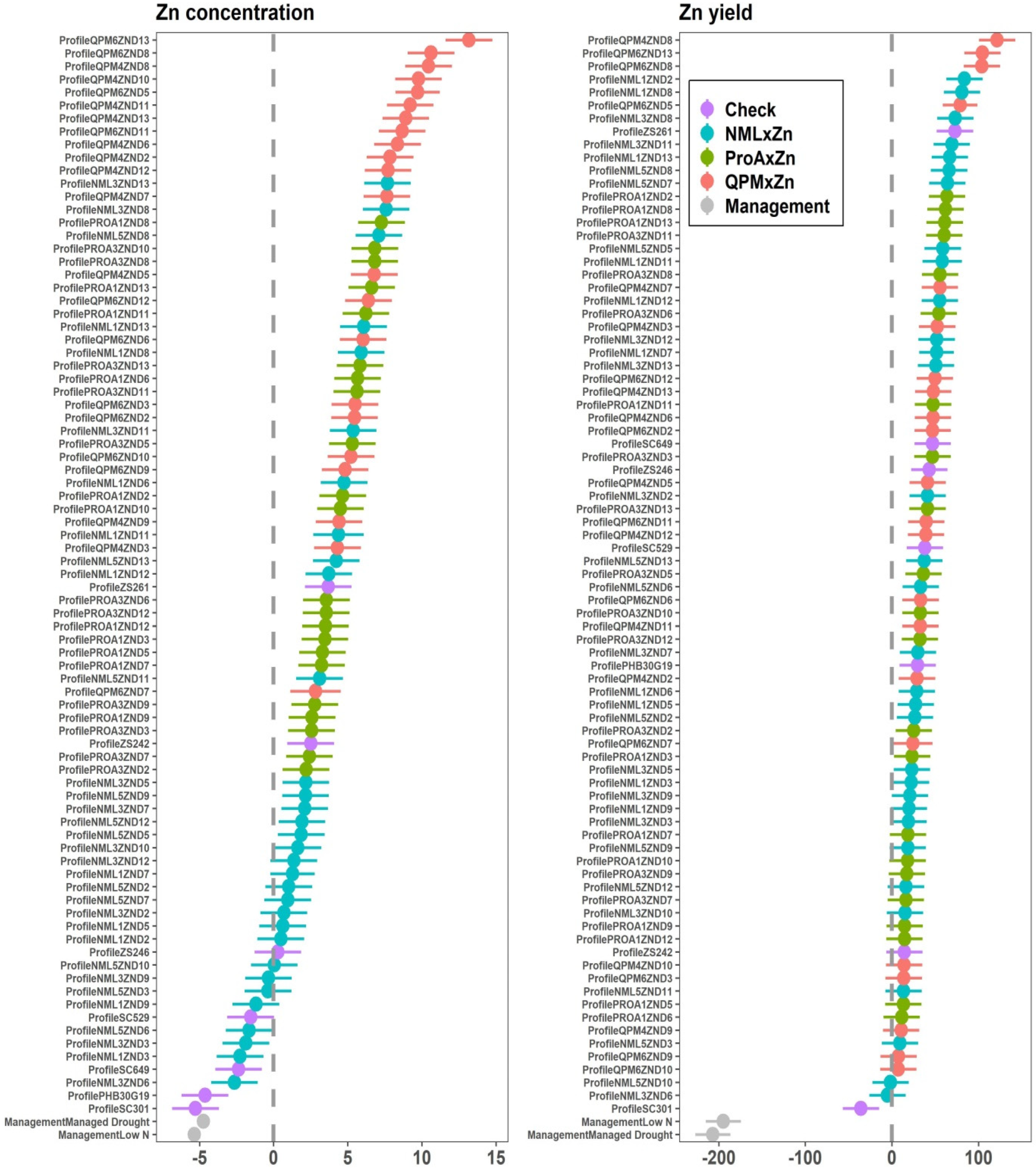
2.3. Genotypic Variance, G × E Interaction and Heritability
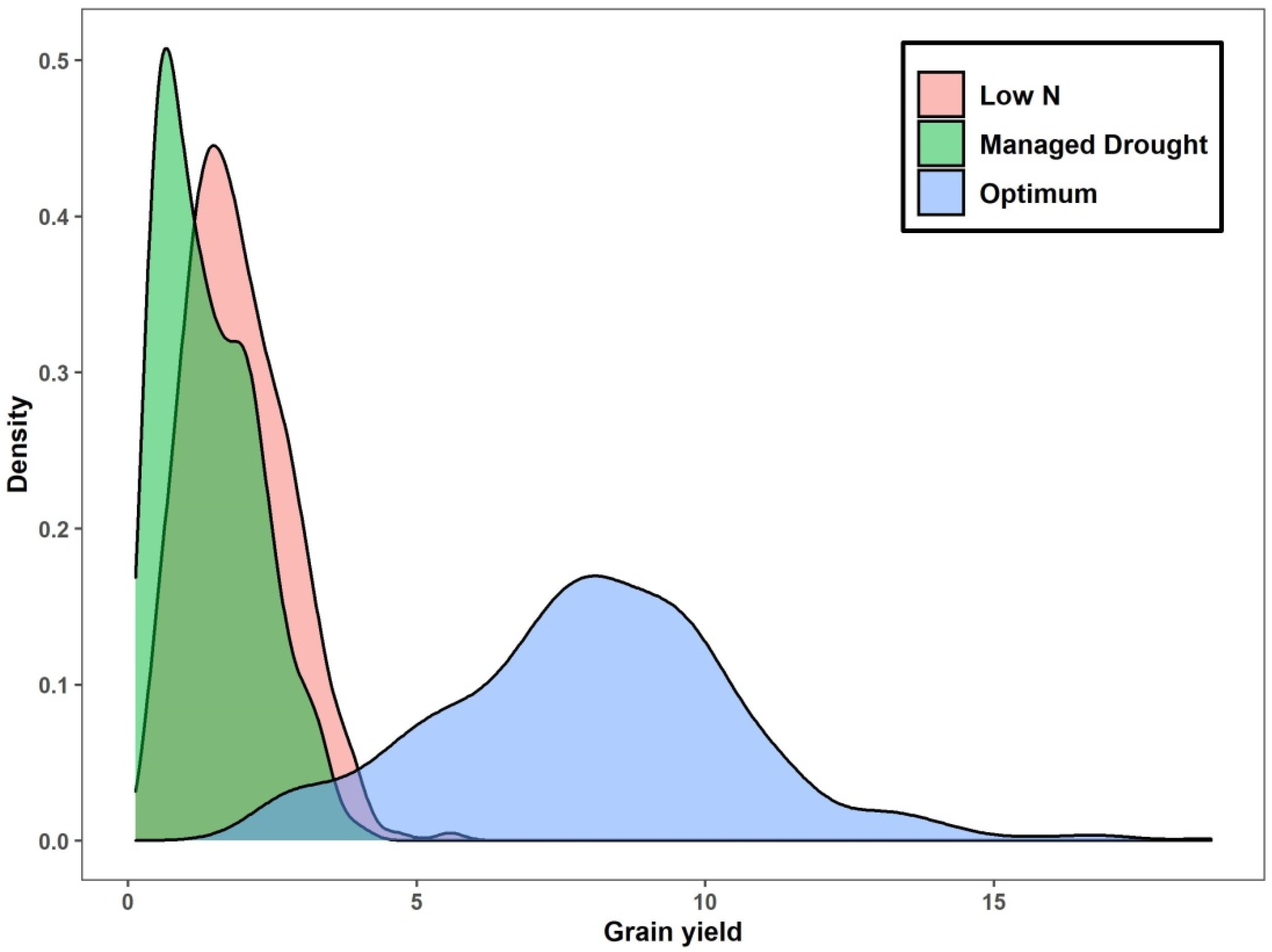
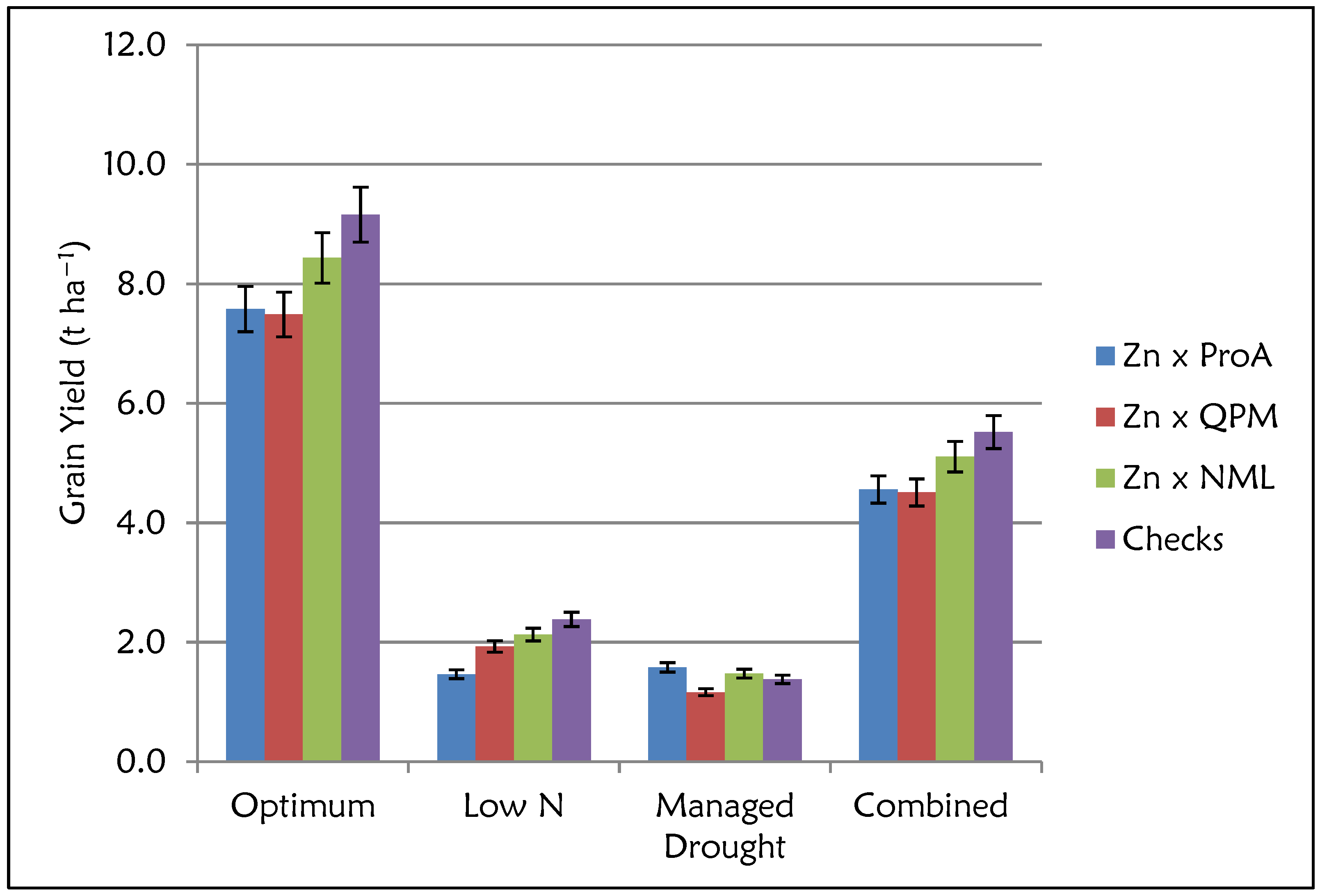
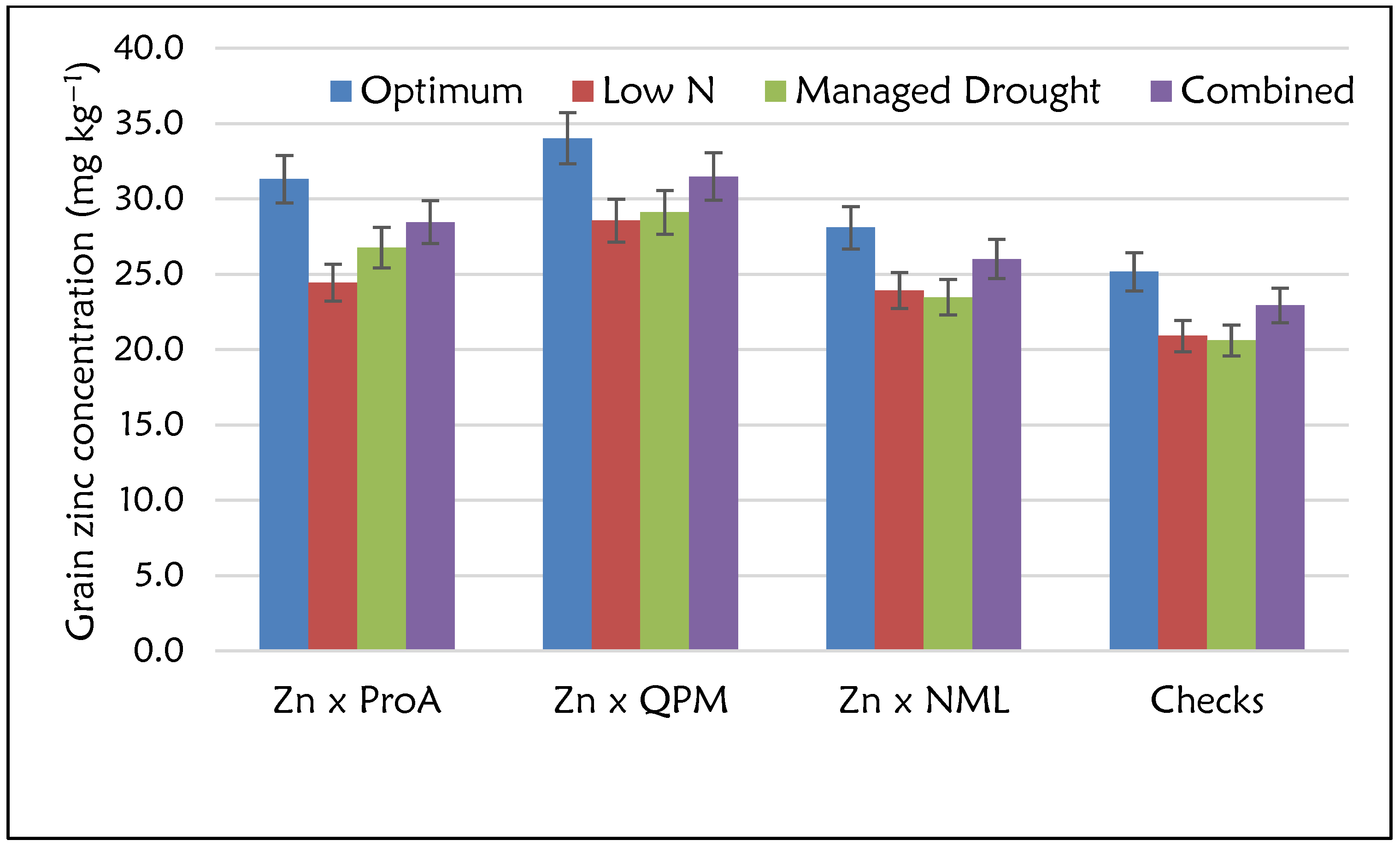
2.4. Grain Yield and Micronutrient Performance by Nutritional Type
2.5. Correlations between Agronomic and Micronutrient Traits
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Sites
4.3. Trial Layout, Management and Data Collection
4.4. Soil Micronutrient Analysis
4.5. Grain Micronutrient Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hindu, V.; Palacios-Rojas, N.; Babu, R.; Suwarno, W.B.; Rashid, Z.; Usha, R.; Saykhedkar, G.R.; Nair, S.K. Identification and validation of genomic regions influencing kernel zinc and iron in maize. Theor. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Secur. 2017, 12, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Botoman, L.; Nalivata, P.C.; Chimungu, J.G.; Munthali, M.W.; Bailey, E.H.; Ander, E.L.; Lark, R.M.; Mossa, A.W.; Young, S.D.; Broadley, M.R. Increasing zinc concentration in maize grown under contrasting soil types in Malawi through agronomic biofortification: Trial protocol for a field experiment to detect small effect sizes. Plant Direct 2020, 4, e00277. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A. Genetic variation for grain mineral content in tropical-adapted maize inbred lines. Food Chem. 2008, 110, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, A.L.; Chander, S.; Ortiz, R.; Menkir, A.; Gedil, M. Genetic basis and breeding perspectives of grain iron and zinc enrichment in cereals. Front. Plant Sci. 2018, 9, 937. [Google Scholar] [CrossRef]
- Wessells, K.R.; Brown, K.H. Estimating the global prevalence of zinc deficiency: Results based on zinc availability in national food supplies and the prevalence of stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [Green Version]
- Manzeke, G.M.; Mtambanengwe, F.; Watts, M.J.; Hamilton, E.M.; Lark, R.M.; Broadley, M.R.; Mapfumo, P. Fertilizer management and soil type influence grain zinc and iron concentration under contrasting smallholder cropping systems in Zimbabwe. Sci. Rep. 2019, 9, 6445. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Hotz, C.; Brown, K.H. Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr. Bull. 2004, 25, S94–S203. [Google Scholar]
- Krebs, N.F.; Miller, L.V.; Hambridge, K.M. Zinc deficiency in infants and children: A review of its complex and synergistic interactions. Paediatr. Int. Child Health 2014, 34, 279–288. [Google Scholar] [CrossRef]
- Cakmak, I.; Kutman, U.B. Agronomic biofortification of cereals with zinc: A review. Eur. J. Soil Sci. 2018, 69, 172–180. [Google Scholar] [CrossRef]
- Nuss, E.T.; Tanumihardjo, S.A. Quality protein maize for Africa: Closing the protein inadequacy gap in vulnerable populations. Adv. Nutr. 2011, 2, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mageto, E.K.; Lee, M.; Dhliwayo, T.; Palacios-Rojas, N.; Vicente, F.S.; Burgueño, J.; Hallauer, A.R. An evaluation of kernel zinc in hybrids of elite Quality Protein Maize (QPM) and non-QPM inbred lines adapted to the tropics based on a mating design. Agronomy 2020, 10, 695. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Xia, H.; Kong, W.; Wang, L.; Xue, Y.; Liu, W.; Zhang, C.; Yang, S.; Li, C. Foliar Zn spraying simultaneously improved concentrations and bioavailability of Zn and Fe in maize grains irrespective of foliar sucrose supply. Agronomy 2019, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Vivek, B.S.; Krivanek, A.F.; Palacios-Rojas, N.; Twumasi-Afriyie, S.; Diallo, A.O. Breeding Quality Protein Maize (QPM): Protocols for Developing QPM Cultivars; CIMMYT: El Batán, Mexico, 2008. [Google Scholar]
- Maqbool, M.A.; Beshir, A.R. Zinc biofortification of maize (Zea mays L.): Status and challenges. Plant Breed. 2019, 138, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Chakraborti, M.; Prasanna, B.M.; Hossain, F.; Mazumdar, S.; Singh, A.M.; Guleria, S.; Gupta, H.S. Identification of kernel iron- and zinc-rich maize inbreds and analysis of genetic diversity using microsatellite markers. J. Plant Biochem. Biotechnol. 2011, 20, 224–233. [Google Scholar] [CrossRef]
- Phuke, R.M.; Anuradha, K.; Radhika, K.; Jabeen, F.; Anuradha, G.; Ramesh, T.; Hariprasanna, K.; Mehtre, S.P.; Deshpande, S.P.; Anil, G.; et al. Genetic variability, genotype × environment interaction, correlation, and GGE biplot analysis for grain iron and zinc concentration and other agronomic traits in RIL population of sorghum (Sorghum bicolor L. Moench). Front. Plant Sci. 2017, 8, 712. [Google Scholar] [CrossRef] [Green Version]
- Bukvic, G.; Antunovic, M.; Popovic, S.; Rastija, M. Effect of P and Zn fertilisation on biomass yield and its uptake by maize lines (Zea mays L.). Plant Soil Environ. 2003, 49, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Cheah, Z.X.; Kopittke, P.M.; Scheckel, K.G.; Noerpel, M.R.; Bell, M.J. Comparison of Zn accumulation and speciation in kernels of sweetcorn and maize differing in maturity. Ann. Bot. 2020, 125, 185–193. [Google Scholar] [CrossRef]
- Rutkowska, B.; Szulc, W.; Bomze, K.; Gozdowski, D.; Spychaj-Fabisiak, E. Soil factors affecting solubility and mobility of zinc in contaminated soils. Int. J. Env. Sci. Technol. 2015, 12, 1687–1694. [Google Scholar] [CrossRef]
- Opala, P.A. Influence of lime and phosphorus application rates on growth of maize in an acid soil. Adv. Agric. 2017, 2017, 7083206. [Google Scholar] [CrossRef] [Green Version]
- Tandzi, L.N.; Mutengwa, C.S.; Ngonkeu, E.L.M.; Woïn, N.; Gracen, V. Breeding for quality protein maize (QPM) varieties: A Review. Agronomy 2017, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Sadeghzadeh, B. A review of zinc nutrition and plant breeding. J. Soil Sci. Plant Nutr. 2013, 13, 905–927. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–448. [Google Scholar] [CrossRef]
- Aref, F. Manganese, iron and copper contents in leaves of maize plants (Zea mays L.) grown with different boron and zinc micronutrients. Afr. J. Biotechnol. 2012, 11, 896–903. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morphophysiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Abdel-Lattif, H.; Shahba, M. Ameliorative effects of calcium sprays on yield and grain nutritional composition of maize (Zea mays L.) cultivars under drought stress. Agriculture 2021, 11, 285. [Google Scholar] [CrossRef]
- Xue, Y.F.; Yue, S.C.; Liu, D.Y.; Zhang, W.; Chen, X.P.; Zou, C.Q. Dynamic zinc accumulation and contributions of pre-and/or post-silking zinc uptake to grain zinc of maize as affected by nitrogen supply. Front. Plant Sci. 2019, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Manzeke, G.M.; Mtambanengwe, F.; Watts, M.J.; Broadley, M.R.; Lark, R.M.; Mapfumo, P. Nitrogen effect on zinc biofortification of maize and cowpea in Zimbabwean smallholder farms. Agron. J. 2020, 112, 2256–2274. [Google Scholar] [CrossRef]
- Grujcic, D.; Yazici, A.M.; Tutus, Y.; Cakmak, I.; Singh, B.R. Biofortification of silage maize with zinc, iron and selenium as affected by nitrogen fertilization. Plants 2021, 10, 391. [Google Scholar] [CrossRef]
- Bojtor, C.; Illès, A.; Mousavi, S.M.; Szèles, A.; Tóth, B.; Nagy, J.; Marton, C.L. Evaluation of the nutrient composition of maize in different NPK fertilizer levels based on multivariate method analysis. Int. J. Agron. 2021, 2021, 5537549. [Google Scholar] [CrossRef]
- Nanganoa, L.T.; Ngome, F.A.; Suh, C.; Basga, S.D. Assessing soil nutrients variability and adequacy for the cultivation of maize, cassava, and sorghum in selected agro-ecological zones of Cameroon. Int. J. Agron. 2020, 2020, 8887318. [Google Scholar] [CrossRef]
- Rengel, Z.; Batten, G.G.; Crowley, D.E. Agronomic approaches for improving the micronutrient density in edible portions of field crops. Field Crops Res. 1999, 60, 27–40. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Naresh, R.K.; Mandal, A.; Singh, R.; Dhaliwal, M.K. Dynamics and transformations of micronutrients in agricultural soils as influenced by organic matter build-up: A review. Environ. Sustain. Indic. 2019, 1–2, 100007. [Google Scholar] [CrossRef]
- Gwenzi, W.; Munondo, R. Long-term impacts of pasture irrigation with treated sewage effluent on nutrient status of a sandy soil in Zimbabwe. Nutr. Cycl. Agroecosys. 2019, 82, 197–207. [Google Scholar] [CrossRef]
- Ortiz-Monasterio, J.I.; Palacios-Rojas, N.; Meng, E. Enhancing the mineral and vitamin content of wheat and maize through plant breeding. J. Cereal Sci. 2007, 46, 293–307. [Google Scholar] [CrossRef]
- Tucker, S.L.; Dohleman, F.G.; Grapov, D.; Flagel, L.; Yang, S.; Wegener, K.M.; Kosola, K.; Swarup, S.; Rapp, R.A.; Bedair, M.; et al. Evaluating maize phenotypic variance, heritability, and yield relationships at multiple biological scales across agronomically relevant environments. Plant Cell Environ. 2019, 43, 880–902. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, H.; Wang, H.; Guo, Z.; Xu, X.; Liu, J.; Wang, S.; Li, W.; Zou, C.; Prasanna, B.M.; et al. Factors affecting genomic selection revealed by empirical evidence in maize. Crop J. 2018, 6, 341–352. [Google Scholar] [CrossRef]
- Govindaraj, M.; Rai, K.N.; Kanatti, A.; Upadhyaya, H.D.; Shivade, H.; Rao, A.S. Exploring the genetic variability and diversity of pearl millet core collection germplasm for grain nutritional traits improvement. Sci. Rep. 2020, 10, 21177. [Google Scholar] [CrossRef]
- Pujar, M.; Govindaraj, M.; Gangaprasad, S.; Kanatti, A.; Shivade, H. Genetic variation and diversity for grain iron, zinc, protein and agronomic traits in advanced breeding lines of pearl millet [Pennisetum glaucum (L.) R. Br.] for biofortification breeding. Genet. Resour. Crop Evol. 2020, 67, 2009–2022. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Mazumdar, S.; Chakraborti, M.; Hossain, F.; Manjaiah, K.M.; Agrawal, P.K. Genetic variability and genotype × environment interactions for kernel Fe and Zn concentrations in maize (Zea mays L.) genotypes. Indian J. Agric. Sci. 2011, 81, 704–711. [Google Scholar]
- Meseka, S.; Menkir, A.; Bossey, B.; Mengesha, W. Performance assessment of drought tolerant maize hybrids under combined drought and heat stress. Agronomy 2018, 8, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutman, U.B.; Yildiz, B.; Ozturk, L.; Cakmak, I. Biofortification of durum wheat with zinc through soil and foliar applications of nitrogen. Cereal Chem. 2010, 87, 1–9. [Google Scholar] [CrossRef]
- Manzeke-Kangara, M.G.; Mtambanengwe, F.; Watts, M.J.; Broadley, M.R.; Lark, R.M.; Mapfumo, P. Can nitrogen fertilizer management improve grain iron concentration of agro-biofortified crops in Zimbabwe? Agronomy 2021, 11, 124. [Google Scholar] [CrossRef]
- Maqbool, M.A.; Issa, A.B.; Khokhar, E.S. Quality protein maize (QPM): Importance, genetics, timeline of different events, breeding strategies and varietal adoption. Plant Breed. 2021, 140, 375–399. [Google Scholar] [CrossRef]
- Queiroz, V.A.V.; Guimarães, P.E.O.; Queiroz, L.R.; Guedes, E.O.; Vasconcelos, V.D.B.; Guimarães, L.J.; Ribeiro, P.E.A.; Schaffert, R.E. Iron and zinc availability in maize lines. Ciência Tecn. Alim. 2011, 31, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.Y.; Zhang, W.; Liu, Y.M.; Chen, X.P.; Zou, C.Q. Soil application of zinc fertilizer increases maize yield by enhancing the kernel number and kernel weight of inferior grains. Front. Plant Sci. 2020, 11, 188. [Google Scholar] [CrossRef]
- Bänziger, M.; Edmeades, G.O.; Beck, D.; Bellon, M. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice; CIMMYT: El-Batán, Mexico, 2000. [Google Scholar]
- Mugandani, M.; Wuta, A.; Makarau, A.; Chipindu, B. Re-classification of agro-ecological regions of Zimbabwe in conformity with climate variability and change. Afr. Crop Sci. J. 2012, 20, 361–369. [Google Scholar]
- Motsara, M.R.; Roy, R.N. Guide to Laboratory Establishment for Plant Nutrient Analysis; FAO Fertilizer and Plant Nutrition Bulletin: Rome, Italy, 2008; Volume 19. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of available phosphorus in water and sodium bicarbonate extracts from soils. Soil Sci. Am. Proc. 1954, 29, 677–678. [Google Scholar]
- Tucker, B.M. The determination of exchangeable calcium and magnesium in carbonate soils. Austr. J. Agric. Res. 1954, 5, 705–715. [Google Scholar] [CrossRef]
- Etheridge, R.D.; Pesti, G.M.; Foster, E.H. A comparison of nitrogen values obtained utilizing the Kjeldahl nitrogen and Dumas combustion methodologies (Leco CNS 2000) on samples typical of an animal nutrition analytical laboratory. Anim. Feed. Sci. Technol. 1998, 73, 21–28. [Google Scholar] [CrossRef]
- Galicia, L.; Nurit, E.; Rosales, A.; Palacios-Rojas, N. Laboratory Protocols. Maize Nutrition Quality and Plant Tissue Analysis Laboratory; CIMMYT: El Batán, Mexico, 2009. [Google Scholar]
- Zarcinas, B.A.; Cartwright, B.; Spouncer, L.R. Nitric acid digestion and multi element analysis of plant material by inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1987, 18, 131–146. [Google Scholar] [CrossRef]
- Alvarado, G.; López, M.; Vargas, M.; Pacheco, A.; Rodríguez, F.; Burgueño, J.; Crossa, J. Multi-Environment Trial Analysis with R (META-R) Version 6.04; CIMMYT: El Batán, Mexico, 2015. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley and Sons: New York, NY, USA, 1984. [Google Scholar]
- VSN International. Genstat for Windows, 17th ed.; VSN International: Hemel Hempstead, UK, 2020. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | Management | pH | TC (%) | IC (%) | TN (%) | SOC (%) | Olsen P mg kg−1 | CEC meq−100 g | Zn mg kg−1 | Fe mg kg−1 | % Clay | % Silt | % Sand |
| ART farm | Optimum | 5.80 | 1.40 | 0.02 | 0.15 | 1.30 | 25.9 | 14.2 | 5.10 | 57.8 | 39.5 | 49.5 | 11.0 |
| CIMMYT | Optimum | 6.40 | 2.20 | 0.01 | 0.20 | 2.20 | 10.4 | 24.5 | 2.70 | 76.1 | 40.5 | 40.5 | 19.0 |
| DR&SS | Low N | 6.10 | 0.80 | 0.02 | 0.04 | 0.70 | 17.8 | 16.4 | 4.70 | 56.1 | 52.0 | 26.5 | 21.5 |
| Chisumbanje | Managed drought | 7.60 | 1.70 | 0.03 | 0.15 | 1.70 | 15.7 | 28.1 | 1.90 | 41.1 | 73.0 | 26.0 | 1.00 |
| † F-test | * | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| 2020 | |||||||||||||
| ART farm | Optimum | 5.70 | 1.40 | 0.02 | 0.15 | 1.40 | 23.4 | 13.2 | 5.70 | 52.3 | 37.5 | 48.5 | 14.0 |
| CIMMYT | Optimum | 6.30 | 2.30 | 0.01 | 0.18 | 2.30 | 12.1 | 25.8 | 2.70 | 77.9 | 41.5 | 39.5 | 19.0 |
| DR&SS | Low N | 6.20 | 0.60 | 0.02 | 0.05 | 0.60 | 18.5 | 14.9 | 4.80 | 57.6 | 51.5 | 26.0 | 22.5 |
| Chisumbanje | Managed drought | 7.60 | 1.80 | 0.02 | 0.17 | 1.70 | 16.0 | 29.1 | 2.00 | 39.1 | 72.5 | 25.0 | 2.50 |
| † F-test | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Environments | Management | Statistics | Traits Ұ | |||||
|---|---|---|---|---|---|---|---|---|
| AD | ASI | PH | GY | Zn | Fe | |||
| CIMMYT 2018/19 (E1) | Optimum | Mean | 69.2 | 1.40 | 218 | 6.80 | 30.7 | 29.7 |
| Range | 60.0–74.0 | −1.00–3.00 | 163–253 | 2.90–13.7 | 12.9–46.7 | 11.3–55.1 | ||
| SD | 2.16 | 0.74 | 20.8 | 2.10 | 6.39 | 8.14 | ||
| ART farm 2018/19 (E2) | Optimum | Mean | 68.6 | 1.40 | 245 | 7.50 | 29.9 | 30.1 |
| Range | 58.0–74.0 | −3.00–5.00 | 159–288 | 1.90–18.8 | 16.4–49.2 | 12.2–54.9 | ||
| SD | 3.06 | 1.09 | 22.3 | 3.53 | 5.80 | 7.91 | ||
| DR&SS 2018/19 (E3) | Low N | Mean | 71.0 | 3.00 | 186 | 1.80 | 24.8 | 20.4 |
| Range | 63.0–76.0 | 0.00–6.00 | 148–226 | 0.40–4.10 | 13.9–39.5 | 7.81–34.9 | ||
| SD | 2.81 | 1.65 | 17.4 | 0.86 | 5.24 | 6.08 | ||
| CHISUMBANJE 2018/19 (E4) | Managed Drought | Mean | 68.0 | 2.70 | 174 | 1.80 | 26.2 | 23.0 |
| Range | 64.0–73.0 | 0.00–7.00 | 140–205 | 0.40–4.10 | 10.7–39.4 | 7.10–37.6 | ||
| SD | 1.88 | 1.50 | 14.1 | 0.82 | 5.09 | 4.85 | ||
| CIMMYT 2019/20 (E5) | Optimum | Mean | 69.6 | 1.40 | 207 | 8.50 | 30.5 | 29.9 |
| Range | 62.0–76.0 | −5.00–4.00 | 150–245 | 4.30–14.3 | 17.3–45.3 | 12.6–58.4 | ||
| SD | 3.14 | 0.90 | 22.3 | 2.07 | 5.87 | 8.06 | ||
| ART farm 2019/20 (E6) | Optimum | Mean | 72.6 | 2.00 | 206 | 9.30 | 30.4 | 30.8 |
| Range | 65.0–84.0 | −5.00–6.00 | 150–240 | 6.50–13.0 | 15.6–57.8 | 12.5–54.4 | ||
| SD | 3.75 | 1.28 | 18.8 | 1.30 | 6.32 | 7.69 | ||
| DR&SS 2019/20 (E7) | Low N | Mean | 71.1 | 2.90 | 182 | 2.00 | 25.2 | 23.1 |
| Range | 64.0–76.0 | 0.00–7.00 | 145–220 | 0.40–5.60 | 12.4–36.6 | 9.20–37.1 | ||
| SD | 2.45 | 1.56 | 16.4 | 0.89 | 4.92 | 6.12 | ||
| CHISUMBANJE 2019/20 (E8) | Managed Drought | Mean | 69.4 | 2.8.0 | 161 | 1.10 | 25.1 | 21.1 |
| Range | 64.0–77.0 | 0.00–6.00 | 116–214 | 0.10–3.80 | 12.1–36.8 | 7.90–42.8 | ||
| SD | 2.71 | 1.51 | 18.1 | 0.72 | 5.23 | 6.10 | ||
| Trait Ұ | CIMMYT 2018/19 (E1) | ART Farm 2018/19 (E2) | DR&SS 2018/19 (E3) | Chisumbanje 2018/19 (E4) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ2g | SE | H2 | δ2g | SE | H2 | δ2g | SE | H2 | δ2g | SE | H2 | |
| AD | 3.05 ** | 0.17 | 0.79 | 1.79 | 0.24 | 0.55 | 6.75 ** | 0.22 | 0.92 | 1.89 ** | 0.15 | 0.72 |
| ASI | 0.19 ** | 0.06 | 0.51 | 0.16 | 0.08 | 0.53 | 2.07 ** | 0.13 | 0.86 | 1.46 ** | 0.12 | 0.78 |
| PH | 20.4 ** | 1.61 | 0.68 | 305 ** | 1.72 | 0.79 | 202 * | 1.35 | 0.81 | 83.2 ** | 1.09 | 0.65 |
| GY | 3.14 ** | 0.16 | 0.85 | 9.57 ** | 0.27 | 0.87 | 0.47 ** | 0.07 | 0.78 | 0.38 ** | 0.06 | 0.73 |
| Zn | 38.9 ** | 0.49 | 0.97 | 27.9 ** | 0.45 | 0.89 | 24.5 ** | 0.40 | 0.94 | 22.8 ** | 0.39 | 0.94 |
| Fe | 49.8 ** | 0.63 | 0.87 | 58.3 ** | 0.61 | 0.97 | 34.6 ** | 0.47 | 0.96 | 20.6 ** | 0.37 | 0.94 |
| Trait | CIMMYT 2019/20 (E5) | ART Farm 2019/20 (E6) | DR&SS 2019/20 (E7) | Chisumbanje 2019/20 (E8) | ||||||||
| δ2g | SE | H2 | δ2g | SE | H2 | δ2g | SE | H2 | δ2g | SE | H2 | |
| AD | 7.72 ** | 0.24 | 0.90 | 11.5 * | 0.29 | 0.92 | 3.83 * | 0.19 | 0.78 | 5.98 ** | 0.21 | 0.89 |
| ASI | 0.24 * | 0.25 | 0.56 | 0.15 ns | 0.10 | 0.51 | 1.41 * | 0.12 | 0.76 | 1.27 ** | 0.12 | 0.73 |
| PH | 274 ** | 1.72 | 0.73 | 217 ** | 1.45 | 0.76 | 142 ** | 1.27 | 0.69 | 86.4 * | 1.39 | 0.63 |
| GY | 3.59 ** | 0.16 | 0.92 | 1.19 ** | 0.10 | 0.83 | 0.47 ** | 0.07 | 0.75 | 0.29 ** | 0.06 | 0.73 |
| Zn | 32.0 ** | 0.45 | 0.96 | 34.7 ** | 0.49 | 0.93 | 21.3 ** | 0.38 | 0.93 | 23.4 ** | 0.40 | 0.92 |
| Fe | 59.8 ** | 0.62 | 0.96 | 52.9 ** | 0.59 | 0.94 | 35.4 ** | 0.47 | 0.97 | 34.1 ** | 0.47 | 0.95 |
| Trait Ұ | Combined Data | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| δ2g | SE | δ2gy | SE | δ2ge | SE | δ2gye | SE | H2 | |
| AD | 0.94 ** | 0.29 | 0.12 | 0.25 | 1.63 * | 0.41 | 2.59 * | 0.21 | 0.59 |
| ASI | 0.08 * | 0.04 | 0.09 | 0.04 | 0.01 | 0.11 | 0.87 * | 0.15 | 0.56 |
| PH | 64.0 ** | 13.8 | 11.0 | 7.7 | 41.3 | 20.0 | 174.7 ** | 0.42 | 0.69 |
| GY | 0.30 ** | 0.11 | 0.01 | 0.10 | 1.23 ** | 0.17 | 0.84 * | 0.17 | 0.89 |
| Zn | 11.2 ** | 2.01 | 1.28 | 0.66 | 14.0 ** | 1.31 | 3.99 ** | 0.32 | 0.81 |
| Fe | 9.36 ** | 2.24 | 0.55 | 1.49 | 29.3 ** | 2.40 | 5.35 ** | 0.05 | 0.66 |
| Traits Ұ | CIMMYT 2018/19 (E1) | Art Farm 2018/19 (E2) | DR&SS 2018/19 (E3) | Chisumbanje 2018/19 (E4) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GY | Zn | Fe | GY | Zn | Fe | GY | Zn | Fe | GY | Zn | Fe | |
| AD | 0.42 ** | 0.07 | 0.22 | 0.18 ** | 0.31 ** | 0.25 | −0.72 ** | 0.11 | −0.01 | −0.48 ** | 0.20 | −0.03 |
| ASI | 0.50 ** | −0.02 | 0.29 * | −0.35 ** | 0.14 | 0.38 ** | −0.88 ** | 0.08 | −0.00 | −0.43 ** | 0.04 | 0.15 |
| PH | 0.58 ** | −0.14 | −0.16 | 0.66 ** | −0.04 | −0.19 | 0.57 ** | −0.11 | −0.22 * | 0.28 * | −0.30 * | −0.11 |
| GY | – | −0.01 | 0.04 | – | −0.12 | −0.18 | – | −0.00 | 0.06 | – | −0.18 | −0.12 |
| Fe | – | 0.75 ** | – | – | 0.72 ** | – | – | 0.52 ** | – | – | 0.67 ** | – |
| Traits | CIMMYT 2019/20 (E5) | Art farm 2019/20 (E6) | DR&SS 2019/20 (E7) | Chisumbanje 2019/20 (E8) | ||||||||
| GY | Zn | Fe | GY | Zn | Fe | GY | Zn | Fe | GY | Zn | Fe | |
| AD | 0.41 ** | 0.05 | −0.09 | −0.21 | 0.14 | −0.02 | −0.52 ** | 0.11 | 0.18 | −0.32 ** | 0.16 | 0.23 * |
| ASI | 0.16 | −0.08 | −0.09 | −0.07 | 0.06 | 0.24 | −0.64 ** | 0.08 | 0.19 | −0.67 ** | −0.16 | −0.07 |
| PH | 0.74 ** | −0.25 * | −0.29 * | 0.58 ** | −0.01 | −0.06 | 0.61 ** | −0.09 | 0.04 | 0.83 ** | −0.11 | 0.02 |
| GY | – | −0.01 | −0.14 | – | −0.14 | −0.16 | – | 0.05 | 0.03 | – | 0.11 | 0.12 |
| Fe | – | 0.56 ** | – | – | 0.70 ** | – | – | 0.54 ** | – | – | 0.51 ** | – |
| Across environments | ||||||||||||
| Traits | GY | Zn | Fe | |||||||||
| AD | −0.09 | 0.43 ** | 0.25 * | |||||||||
| ASI | −0.57 ** | 0.34 ** | 0.60 ** | |||||||||
| PH | 0.99 ** | −0.39 ** | −0.51 ** | |||||||||
| GY | – | −0.44 ** | −0.43 ** | |||||||||
| Fe | – | 0.97 ** | – | |||||||||
| NO. | Code | Parent Type | Nutritional Type | Origin | Zn (mg kg−1) | Fe (mg kg−1) |
|---|---|---|---|---|---|---|
| 1 | D2 | Male | Zinc donor | CIMMYT-Mexico | 33.85 | 28.27 |
| 2 | D3 | Male | Zinc donor | CIMMYT-Mexico | 33.72 | 28.98 |
| 3 | D5 | Male | Zinc donor | CIMMYT-Mexico | 30.36 | 35.00 |
| 4 | D6 | Male | Zinc donor | CIMMYT-Mexico | 28.68 | 31.52 |
| 5 | D7 | Male | Zinc donor | CIMMYT-Mexico | 32.18 | 28.06 |
| 6 | D8 | Male | Zinc donor | IITA | 30.52 | 34.40 |
| 7 | D9 | Male | Zinc donor | IITA | 30.25 | 26.17 |
| 8 | D10 | Male | Zinc donor | IITA | 34.29 | 28.77 |
| 9 | D11 | Male | Zinc donor | IITA | 30.09 | 32.14 |
| 10 | D12 | Male | Zinc donor | IITA | 30.02 | 26.72 |
| 11 | D13 | Male | Zinc donor | IITA | 27.25 | 28.39 |
| 12 | NML1 | Female | Normal | CIMMYT-SARO | 34.39 | 34.39 |
| 13 | NML3 | Female | Normal | CIMMYT-SARO | 28.34 | 28.34 |
| 14 | NML5 | Female | Normal | CIMMYT-SARO | 30.11 | 30.11 |
| 15 | PROA1 | Female | Provitamin A | CIMMYT-SARO | 30.82 | 28.78 |
| 16 | PROA3 | Female | Provitamin A | CIMMYT-SARO | 28.10 | 34.53 |
| 17 | QPM4 | Female | QPM | CIMMYT-SARO | 35.48 | 33.65 |
| 18 | QPM6 | Female | QPM | CIMMYT-SARO | 29.19 | 30.67 |
| 19 | C1 | Check | Provitamin A | Seed company | ND | ND |
| 20 | C2 | Check | Provitamin A | Seed company | ND | ND |
| 21 | C3 | Check | QPM | Seed company | ND | ND |
| 22 | C4 | Check | Normal | Seed company | ND | ND |
| 23 | C5 | Check | Normal | Seed company | ND | ND |
| 24 | C6 | Check | Normal | Seed company | ND | ND |
| 25 | C7 | Check | Normal | Seed company | ND | ND |
| Site/Year | Agro-Ecology | Latitude | Longitude | Altitude (masl) | Annual Rainfall (mm) | Management | Entries | Planting Time | Soil Type |
|---|---|---|---|---|---|---|---|---|---|
| CIMMYT 2018/19 (E1) | IIa | 17°48′ S | 31°03′ E | 1483 | 850 | Optimum | 84 × 2 reps | November | Ferralsol |
| ART farm 2018/19 (E2) | IIa | 17°42′ S | 31°5′ E | 1556 | 850 | Optimum | 84 × 2 reps | November | Lixisol |
| DR&SS 2018/19 (E3) | IIa | 17°13′ S | 31°03′ E | 1506 | 850 | Low N | 84 × 2 reps | November | Ferralsol |
| Chisumbanje 2018/19 (E4) | V | 20°47′ S | 32°13′ E | 480 | 450 | Managed drought | 84 × 2 reps | May | Vertisol |
| CIMMYT 2019/20 (E5) | IIa | 17°48′ S | 31°03′ E | 1483 | 850 | Optimum | 84 × 2 reps | November | Ferralsol |
| ART farm 2019/20 (E6) | IIa | 17°42′ S | 31° 5′ E | 1556 | 850 | Optimum | 84 × 2 reps | November | Lixisol |
| DR&SS 2019/20 (E7) | IIa | 17°13′ S | 31°03′ E | 1506 | 850 | Low N | 84 × 2 reps | November | Ferralsol |
| Chisumbanje 2019/20 (E8) | V | 20°47′ S | 32°13′ E | 480 | 450 | Managed drought | 84 × 2 reps | May | Vertisol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goredema-Matongera, N.; Ndhlela, T.; van Biljon, A.; Kamutando, C.N.; Cairns, J.E.; Baudron, F.; Labuschagne, M. Genetic Variation of Zinc and Iron Concentration in Normal, Provitamin A and Quality Protein Maize under Stress and Non-Stress Conditions. Plants 2023, 12, 270. https://doi.org/10.3390/plants12020270
Goredema-Matongera N, Ndhlela T, van Biljon A, Kamutando CN, Cairns JE, Baudron F, Labuschagne M. Genetic Variation of Zinc and Iron Concentration in Normal, Provitamin A and Quality Protein Maize under Stress and Non-Stress Conditions. Plants. 2023; 12(2):270. https://doi.org/10.3390/plants12020270
Chicago/Turabian StyleGoredema-Matongera, Nakai, Thokozile Ndhlela, Angeline van Biljon, Casper N. Kamutando, Jill E. Cairns, Frederic Baudron, and Maryke Labuschagne. 2023. "Genetic Variation of Zinc and Iron Concentration in Normal, Provitamin A and Quality Protein Maize under Stress and Non-Stress Conditions" Plants 12, no. 2: 270. https://doi.org/10.3390/plants12020270