
Grapevine Badnavirus 1: Detection, Genetic Diversity, and Distribution in Croatia

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of Real-Time and Conventional PCR Detection Protocols
2.2. Virus Detection during Dormancy and the Growing Season
2.3. Virus Screening in Collection Plantations and Commercial Vineyards
2.4. Direct Sequencing and Phylogenetic Analysis
3. Results
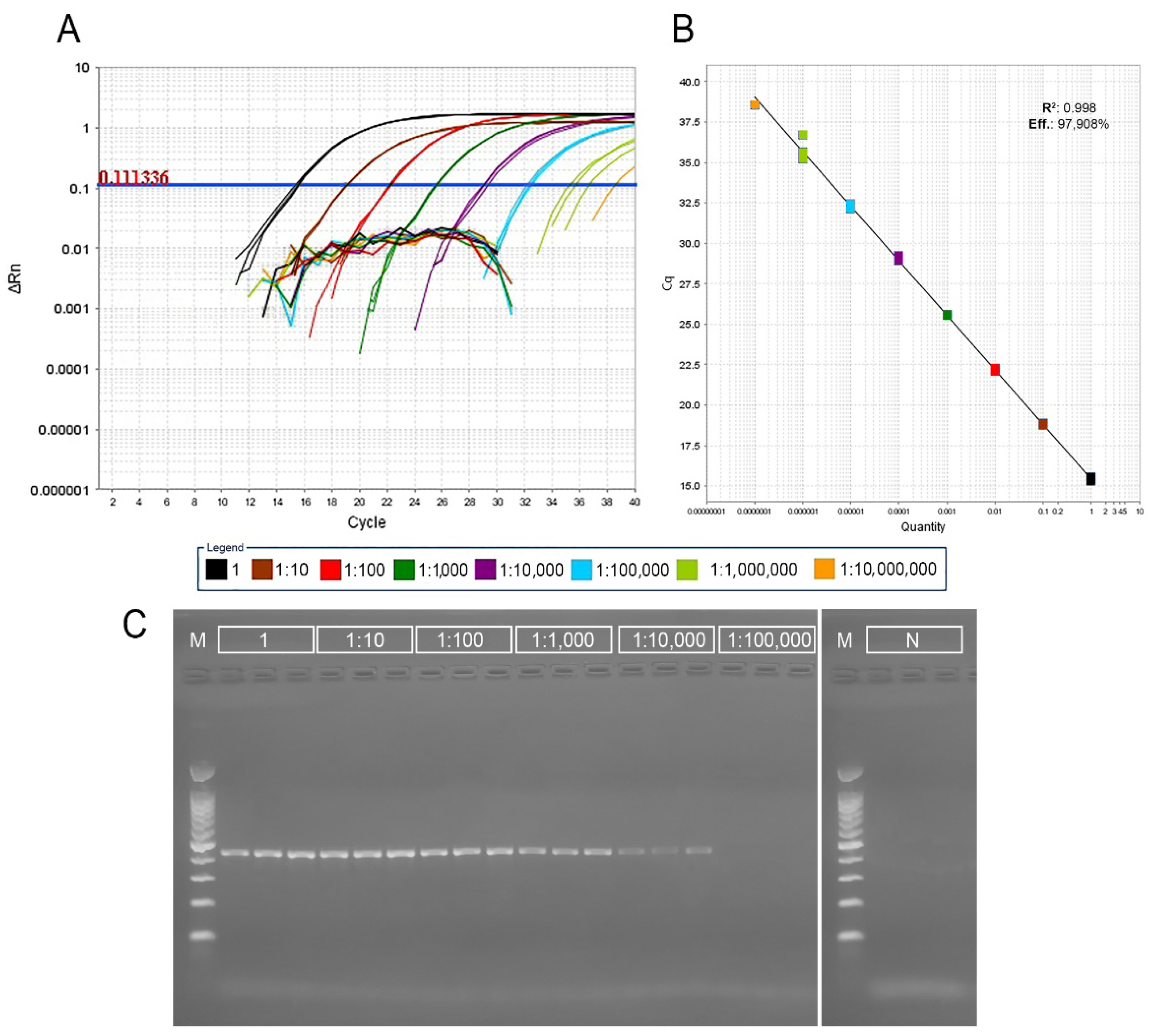
3.1. Real-Time and Conventional PCR Detection
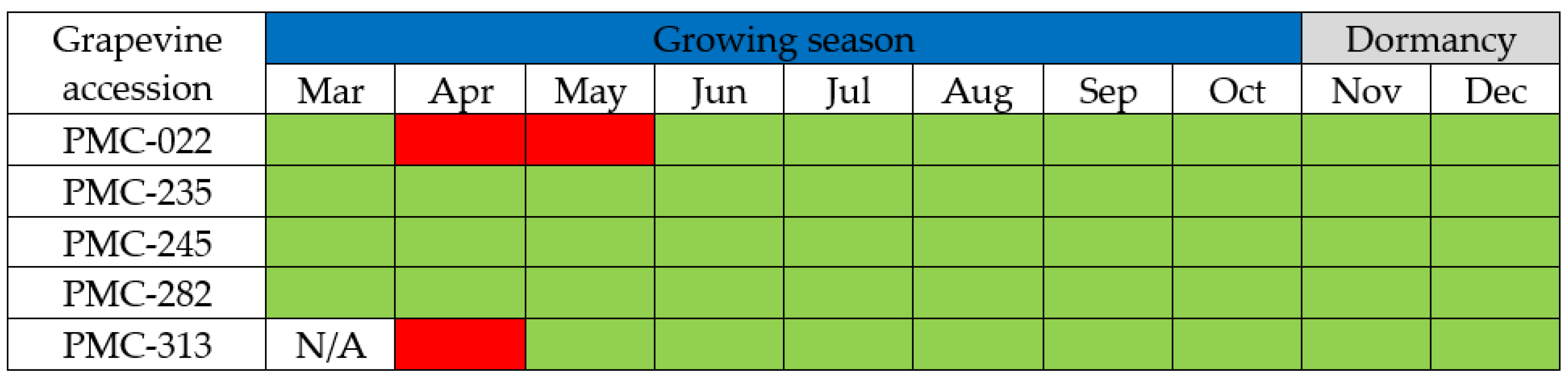
3.2. Virus Detection during Dormancy and the Growing Season
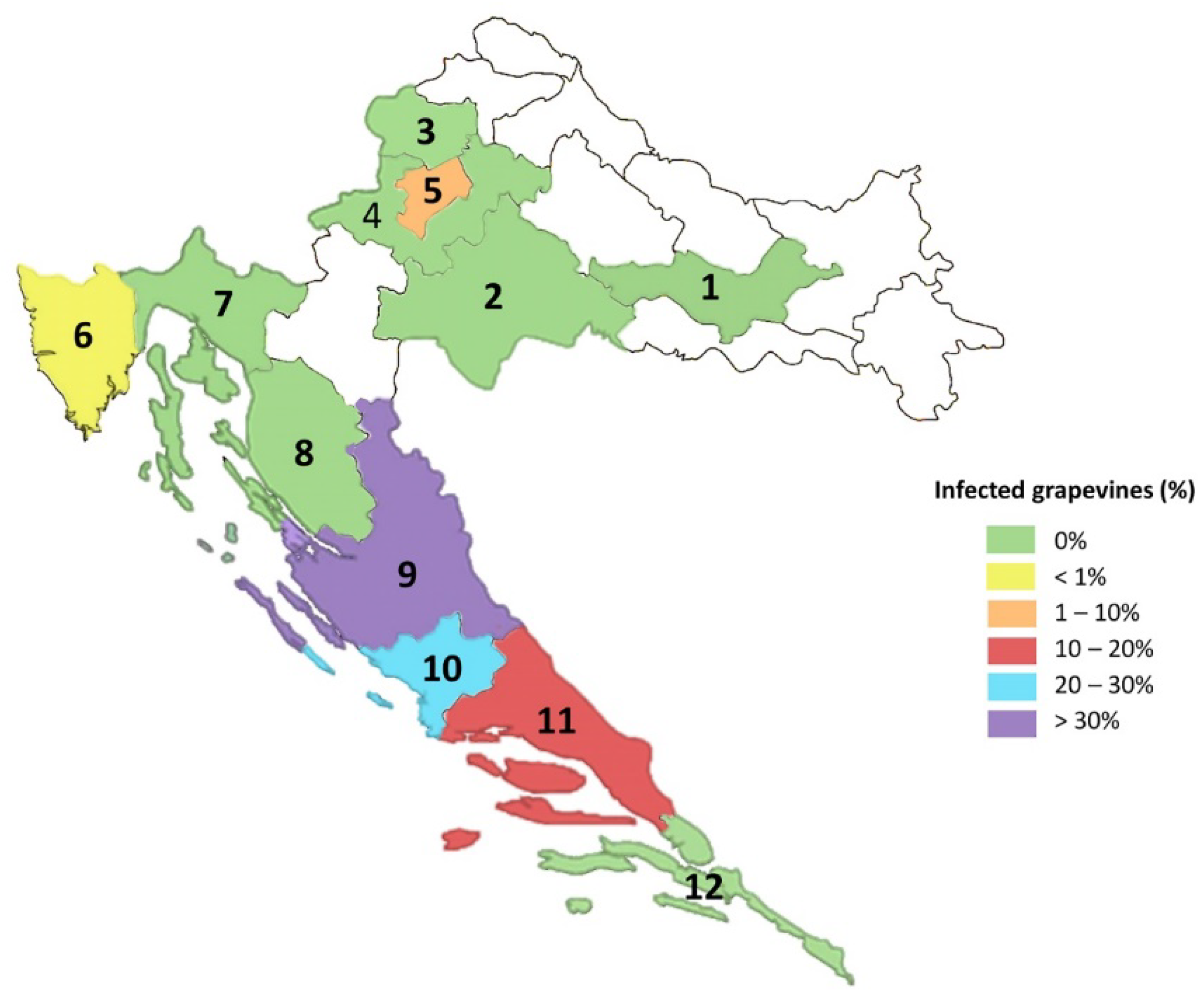
3.3. Virus Screening in Collection Plantations and Commercial Vineyards
3.4. Direct Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maletić, E.; Karoglan Kontić, J.; Pejić, I.; Preiner, D.; Zdunić, G.; Bubola, M.; Stupić, D.; Andabaka, Ž.; Marković, Z.; Šimon, S.; et al. Green Book: Indigeneous Grapevine Cultivars of Croatia; Državni Zavod za Zaštitu Prirode: Zagreb, Croatia, 2015. [Google Scholar]
- Karoglan Kontić, J.; Pejić, I.; Maletić, E.; Sladonja, B.; Poljuha, D.; Vokurka, A.; Zdunić, G.; Preiner, D.; Šimon, S.; Rühl, E. Virus diseases screening in clonal selection of Croatian grapevine cultivars. Acta Hortic. 2009, 827, 623–626. [Google Scholar] [CrossRef]
- Vončina, D.; Badurina, D.; Preiner, D.; Cvjetković, B.; Maletić, E.; Karoglan Kontić, J. Incidence of virus infections in grapevines from Croatian collection plantations. Phytopathol. Mediterr. 2011, 50, 316–326. [Google Scholar]
- Marković, Z.; Preiner, D.; Bošnjak, A.; Safner, T.; Stupić, D.; Andabaka, Ž.; Maletić, E.; Chatelet, P.; Engelmann, F.; Karoglan Kontić, J. In vitro introduction of healthy and virus-infected genotypes of native Croatian grapevine cultivars. Open Life Sci. 2014, 9, 1087–1098. [Google Scholar] [CrossRef]
- Vončina, D.; Preiner, D.; Šimon, S.; Cvjetković, B.; Maletić, E.; Pejić, I.; Karoglan Kontić, J. Distribution of nine viruses in Croatian autochthonous grapevine (Vitis vinifera L.) cultivars from Dalmatian region included in clonal selection. J. Cent. Eur. Agric. 2019, 20, 262–273. [Google Scholar] [CrossRef]
- Martelli, G.P. An overview on grapevine viruses, viroids, and the diseases they cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 31–46. [Google Scholar] [CrossRef]
- Martelli, G.P. Where grapevine virology is heading to. In Proceedings of the 19th Meeting of the ICVG, Santiago, Chile, 9–12 April 2018; pp. 10–15. [Google Scholar]
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Saldarelli, P.; Giampetruzzi, A.; Maree, H.J.; Rwahnih, A. High throughput sequencing: Advantages beyong virus identification. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 625–642. [Google Scholar]
- Teycheney, P.-Y.; Geering, A.D.W.; Dasgupta, I.; Hull, R.; Kreuze, J.F.; Lockhart, B.; Muller, E.; Olszewski, N.; Pappu, H.; Pooggin, M.M.; et al. ICTV virus taxonomy profile: Caulimoviridae. J. Gen. Virol. 2020, 101, 1025–1026. [Google Scholar] [CrossRef]
- Borah, B.K.; Sharma, S.; Kant, R.; Anthony-Johnson, A.M.; Saigopal, D.V.R.; Dasgupta, I. Bacilliform DNA containing plant viruses in the tropics: Commonalities within a genetically diverse group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef]
- Lockhart, B.E.L.; Olsweski, N.E. Plant virus badnavirus group. In The Encyclopedia of Virology, 1st ed.; Webster, R.G., Granoff, A., Eds.; Academic Press: New York, NY, USA, 1994; Volume 1, pp. 139–143. [Google Scholar]
- Bhat, A.I.; Hohn, T.; Selvarajan, R. Badnaviruses: The current global scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef]
- Zhang, Y.; Singh, K.; Kaur, R.; Qiu, W. Association of a novel DNA virus with the grapevine vein-clearing and vine decline syndrome. Phytopathology 2011, 101, 1081–1090. [Google Scholar] [CrossRef]
- Qiu, W.P.; Avery, J.D.; Lunden, S. Characterization of a severe virus-like disease in Chardonnay grapevines in Missouri. Plant Health Prog. 2007, 8, 39. [Google Scholar] [CrossRef]
- Petersen, S.; Keith, C.; Austin, K.; Howard, S.; Su, L.; Qiu, W. A natural reservoir and transmission vector of grapevine vein clearing virus. Plant Dis. 2018, 103, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Schoelz, J. Grapevine vein clearing virus: Diagnostics, genome, genetic diversity, and management. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 315–330. [Google Scholar] [CrossRef]
- Cieniewicz, E.J.; Qiu, W.; Saldarelli, P.; Fuchs, M. Believing is seeing: Lessons from emerging viruses in grapevine. J. Plant Pathol. 2020, 102, 619–632. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Avgelis, A.D. A novel grapevine badnavirus is associated with the Roditis leaf discoloration disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Rumbos, I.C.; Avgelis, A.D. Roditis leaf discoloration—A new virus disease of grapevine: Symptomatology and transmission to indicators plants. J. Phytopathol. 1989, 152, 274–278. [Google Scholar] [CrossRef]
- Chiumenti, M.; Morelli, M.; Giampetruzzi, A.; Palmisano, F.; Savino, V.N.; La Notte, P.; Martelli, G.P.; Saldarelli, P. First report of grapevine Roditis leaf discoloration-associated virus in Italy. J. Plant Pathol. 2015, 97, 551. [Google Scholar] [CrossRef]
- Vončina, D.; Almeida, R.P.P. Screening of some Croatian autochthonous grapevine varieties reveals multitude of viruses including novel ones. Arch. Virol. 2018, 163, 2239–2243. [Google Scholar] [CrossRef]
- Ulubaş Serçe, Ç.; Altan, B.; Bolat, V.; Ayyaz, M.; Çifçi, O.; Önder, S.; Öztürk Gökçe, Z.N.; Maliogka, V.I. First Report of grapevine Roditis leaf discoloration-associated virus infecting grapevine (Vitis vinifera) in Turkey. Plant Dis. 2018, 2, 256. [Google Scholar] [CrossRef]
- Ekemen, M. Investigation of Etiology on Grapevine Roditis Leaf Discoloration-Associated Virus. Master Thesis, Niğde Ömer Halisdemir University, Merkez Yerleşke, Bor Yolu Üzeri, Merkez/Niğde, Turkey, February 2021. Available online: http://acikerisim.ohu.edu.tr/xmlui/handle/11480/8548 (accessed on 5 August 2019).
- Morán, F.; Sassalou, C.-L.; Canales, C.; Maliogka, V.I.; Olmos, A.; Ruiz-García, A.B. Specific real-time PCR for the detection and absolute quantitation of grapevine Roditis leaf discoloration-associated virus, an EPPO alert pathogen. Plants 2020, 9, 1151. [Google Scholar] [CrossRef]
- Čarija, M.; Radić, T.; Černi, S.; Mucalo, A.; Zdunić, G.; Vončina, D.; Jagunić, M.; Hančević, K. Prevalence of virus infections and GLRaV-3 genetic diversity in selected clones of Croatian indigenous grapevine cultivar Plavac Mali. Pathogens 2022, 11, 176. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Diaz-Lara, A.; Arnold, K.; Cooper, M.L.; Smith, R.J.; Zhuang, G.; Battany, M.C.; Bettiga, L.J.; Rowhani, A.; Golino, D. Incidence and genetic diversity of grapevine Pinot gris virus in California. Am. J. Enol. Vitic. 2021, 72, 164–169. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Rowhani, A.; Biardi, L.; Johnson, R.; Saldarelli, P.; Zhang, Y.P.; Chin, J.; Green, M. Simplified sample preparation method and one-tube RT-PCR for grapevine viruses. In Proceedings of the 13th Meeting of the ICVG, Adelaide, Australia, 12–18 March 2000; p. 148. [Google Scholar]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y.; et al. Characterization of grapevine leafroll-associated virus 3 genetic variants and application towards RT-qPCR assay design. PLoS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Vončina, D.; Simon, S.; Đermic, E.; Cvjetkovic, B.; Pejic, I.; Maletic, E.; Karoglan Kontić, J. Distribution and partial molecular characterization of Grapevine leafroll-associated virus 2 (GLRaV-2) found in Croatian autochthonous grapevine (Vitis vinifera L.) germplasm. J. Plant Dis. Prot. 2010, 117, 194–200. [Google Scholar] [CrossRef]
- Hančević, K.; Saldarelli, P.; Čarija, M.; Černi, S.; Zdunić, G.; Mucalo, A.; Radić, T. Predominance and Diversity of GLRaV-3 in Native Vines of Mediterranean Croatia. Plants 2021, 10, 17. [Google Scholar] [CrossRef]
- Vončina, D.; Šimon, S.; Đermić, E.; Cvjetkovic, B.; Pejić, I.; Maletić, E.; Karoglan Kontić, J. Differential properties of grapevine virus B isolates from Croatian autochthonous grapevine cultivars. Eur. J. Plant Pathol. 2011, 93, 283–289. [Google Scholar]
- Tsai, C.W.; Daugherty, M.P.; Almeida, R.P.P. Seasonal dynamics and virus translocation of grapevine leafroll-associated virus 3 in grapevine cultivars. Plant Pathol. 2011, 61, 977–985. [Google Scholar] [CrossRef]
- Shabanian, M.; Xiao, H.; Meng, B. Seasonal dynamics and tissue distribution of two major viruses associated with grapevine leafroll under cool climate condition. Eur. J. Plant Pathol. 2020, 158, 1017–1031. [Google Scholar] [CrossRef]
- Teliz, D.; Tanne, E.; Gonsalves, D.; Zee, F. Field serological detection of viral antigens associated with grapevine leafroll disease. Plant Dis. 1987, 71, 704–709. [Google Scholar] [CrossRef]
- Kahl, D.; Lowery, D.T.; Hart, M.; Úrbez-Torres, J.R. Seasonal dynamics and optimal diagnostics of grapevine red blotch virus in a British Columbian vineyard. Can. J. Plant Pathol. 2021, 44, 453–464. [Google Scholar] [CrossRef]
- Rowhani, A.; Walker, M.A.; Rokni, S. Sampling strategies for the detection of grapevine fanleaf virus and the grapevine strain of tomato ringspot virus. Vitis 1992, 31, 35–44. [Google Scholar]
- Krebelj, A.J.; Čepin, U.; Ravnikar, M.; Novak, M.P. Spatio-temporal distribution of grapevine fanleaf virus (GFLV) in grapevine. Eur. J. Plant Pathol. 2015, 142, 159–171. [Google Scholar] [CrossRef]
- Bertazzon, N.; Forte, V.; Filippin, L.; Causin, R.; Maixner, M.; Angelini, E. Association between genetic variability and titre of grapevine pinot gris virus with disease symptoms. Plant Pathol. 2016, 66, 949–959. [Google Scholar] [CrossRef]
- Poljuha, D.; Sladonja, B.; Bubola, M. Incidence of viruses infecting grapevine varieties in Istria (Croatia). J. Food Agric. Environ. 2010, 8, 166–169. [Google Scholar]
- Karoglan Kontić, J.; Preiner, D.; Šimon, S.; Zdunić, G.; Poljuha, D.; Maletić, E. Sanitary status of Croatian native grapevine varieties. Agric. Conspec. Sci. 2009, 74, 99–103. [Google Scholar]
- Schoelz, J.; Volenberg, D.; Adhab, M.; Fang, Z.; Klassen, V.; Spinka, C.; Rwahnih, M.A. A survey of viruses found in grapevine cultivars grown in Missouri. Am. J. Enol. Vitic. 2020, 72, 73–84. [Google Scholar] [CrossRef]
- Minafra, A.; Mawassi, M.; Goszczynski, D.; Saldarelli, P. Grapevine vitiviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 229–256. [Google Scholar] [CrossRef]
- Herrbach, E.; Alliaume, A.; Prator, C.A.; Daane, K.M.; Cooper, M.L.; Almeida, R.P.P. Vector transmission of grapevine leafroll-associated viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 483–503. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Orientation | Assay/Target Gene | Primer/Probe Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|---|---|
| GBV-1 | GBV-1-F | Forward primer | Real-time PCR/reverse transcriptase (RT) | GGYAAGGAAAGAATGGTCTTCA | 181 |
| GBV-1-R | Reverse primer | TCCATTCTATAGAATCTGGGTGCAT | |||
| GBV-1-P | TaqMan probe | AAGATCAATATAGCCTTCCTGGA | |||
| GBV-1-F | Forward primer | Conventional PCR/reverse transcriptase (RT) | GGYAAGGAAAGAATGGTCTTCA | 419 | |
| GBV-1-R_con | Reverse primer | TTTGTTGGGCTCARGACAAGCC |
| Wine-Growing Region | County | Region | Location | Positive/Total Number of Analysed Samples (%) |
|---|---|---|---|---|
| Continental | city of Zagreb | Zagreb | Grapevine virus collection | 10/196 (5.1%) |
| Jazbina (National collection 2) | 15/113 (13.3%) | |||
| Coastal | Istria | Poreč | Kršete | 2/105 (1.9%) |
| Zadar | Rava island | Mala Rava | 66/194 (34%) | |
| Vela Rava | 85/110 (77.3%) | |||
| Pag island | Pag 2 | 10/30 (33.3%) | ||
| Pag 3 | 32/58 (55.2%) | |||
| Pag 4 | 15/30 (50%) | |||
| Nin | Queen’s beach | 48/50 (96%) | ||
| Zemunik | Zemunik Donji | 29/50 (58%) | ||
| Šibenik-Knin | Primošten | Vezac | 4/10 (40%) | |
| Bucavac | 7/30 (23.3%) | |||
| Split-Dalmatia | Kaštela | Kaštel Sućurac | 6/50 (12%) | |
| Radun | 8/54 (14.8%) | |||
| Kaštel Lukšić | 22/50 (44%) | |||
| Furnaže | 9/50 (18%) | |||
| Stomorija | 2/50 (4%) | |||
| Kaštel novi | 7/70 (10%) | |||
| Split | Collection plantation | 16/105 (15.2%) | ||
| Proložac | Vučija Draga | 5/135 (3.7%) | ||
| Hvar isand | Velo vijelo | 12/50 (24%) | ||
| Ivan Dolac 1 | 48/50 (96%) | |||
| Ivan Dolac 2 | 34/50 (68%) | |||
| Crkvenik | 24/50 (48%) | |||
| Vis island | Petričevo | 32/231 (13.8%) | ||
| Šolta island | Srednje selo | 19/130 (14.6%) | ||
| Gornje selo | 9/51 (17.6%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jagunić, M.; Diaz-Lara, A.; Rwahnih, M.A.; Preiner, D.; Stevens, K.; Zdunić, G.; Hwang, M.; Vončina, D. Grapevine Badnavirus 1: Detection, Genetic Diversity, and Distribution in Croatia. Plants 2022, 11, 2135. https://doi.org/10.3390/plants11162135
Jagunić M, Diaz-Lara A, Rwahnih MA, Preiner D, Stevens K, Zdunić G, Hwang M, Vončina D. Grapevine Badnavirus 1: Detection, Genetic Diversity, and Distribution in Croatia. Plants. 2022; 11(16):2135. https://doi.org/10.3390/plants11162135
Chicago/Turabian StyleJagunić, Martin, Alfredo Diaz-Lara, Maher Al Rwahnih, Darko Preiner, Kristian Stevens, Goran Zdunić, Minsook Hwang, and Darko Vončina. 2022. "Grapevine Badnavirus 1: Detection, Genetic Diversity, and Distribution in Croatia" Plants 11, no. 16: 2135. https://doi.org/10.3390/plants11162135