
Remediation Capacity of Different Microalgae in Effluents Derived from the Cigarette Butt Cleaning Process

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. CB Collection and Cleaning Process
2.2. Microalgal Strains and Growth Conditions
2.3. Characterization of LG1 Strain and Phylogenetic Analysis
2.4. Evaluation of Microalgal Strains in Remediation
2.5. Analytical Determinations
2.6. Photosynthetic Pigments of Microalgal Strains
2.7. Statistical Analyses
3. Results
3.1. Identification of the LG1 Strain
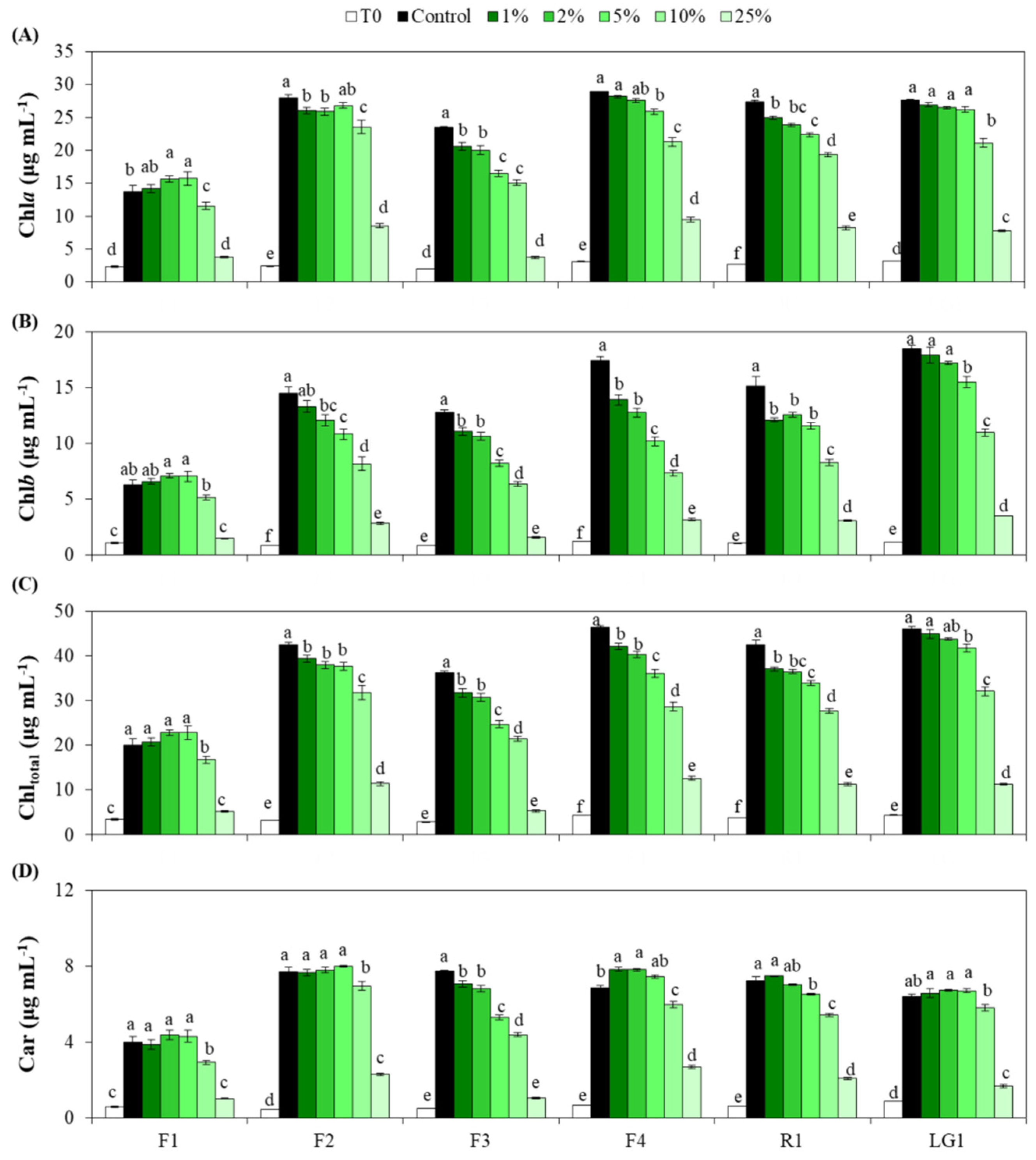
3.2. Photosynthetic Pigments of Microalgal Strains
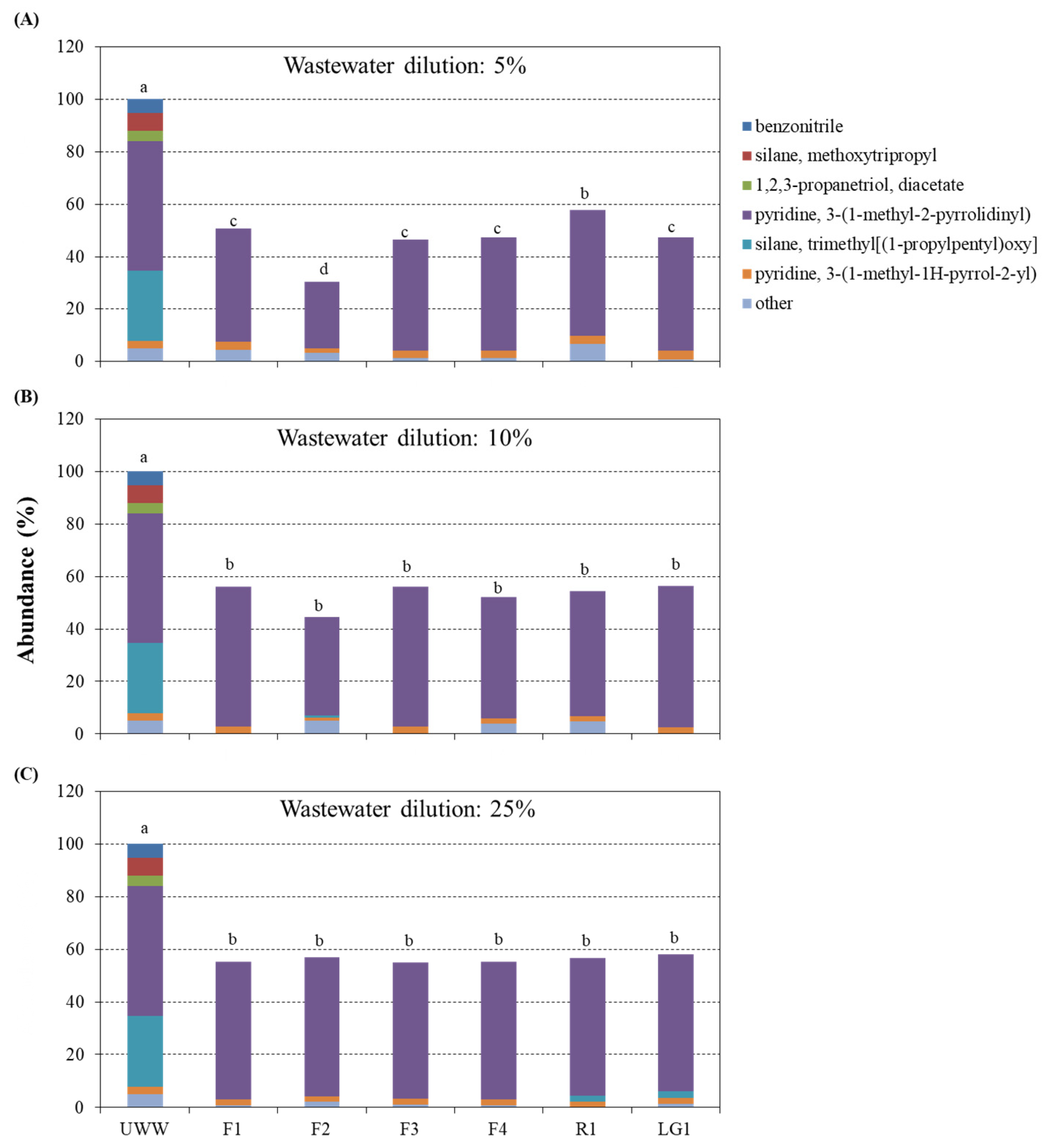
3.3. CB Wastewater Subjected to Microalgal-Based Remediation
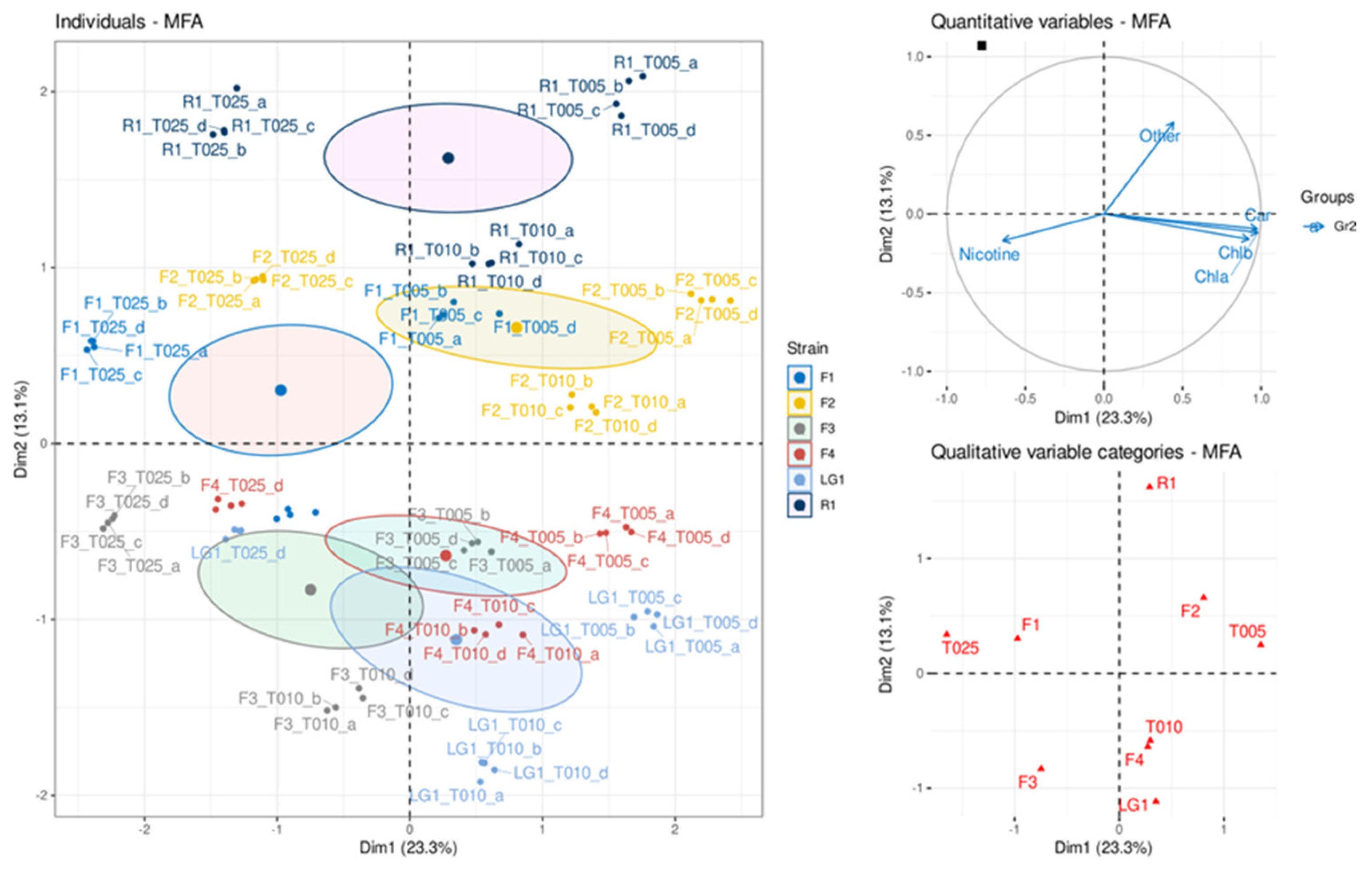
3.4. Multiple Factor Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurmus, H.; Mohajerani, A. The toxicity and valorization options of cigarette butts. Waste Manag. 2020, 104, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Green, D.S.; Tongue, A.D.W.; Boots, B. The ecological impacts of discarded cigarette butts. Trends Ecol. Evol. 2022, 37, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Rebischung, F.; Chabot, L.; Biaudet, H.; Pandard, P. Cigarette butts: A small but hazardous waste, according to European regulation. Waste Manag. 2018, 82, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Marinello, S.; Lolli, F.; Gamberini, R.; Rimini, B. A second life for cigarette butts? A review of recycling solutions. J. Hazard. Mater. 2020, 384, 121245. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, L.; Huarancca Reyes, T.; Curadi, M.; Guglielminetti, L. Recycling cigarette filters as plant growing substrate in soilless system. Horticulturae 2022, 8, 135. [Google Scholar] [CrossRef]
- Sharma, N.K.; Rai, A.K. Biodiversity and biogeography of microalgae: Progress and pitfalls. Environ. Rev. 2011, 19, 1–15. [Google Scholar] [CrossRef]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Lutzu, G.A.; Ciurli, A.; Chiellini, C.; Di Caprio, F.; Concas, A.; Dunford, N.T. Latest developments in wastewater treatment and biopolymer production by microalgae. J. Environ. Chem. Eng. 2021, 9, 104926. [Google Scholar] [CrossRef]
- Chiellini, C.; Guglielminetti, L.; Sarrocco, S.; Ciurli, A. Isolation of four microalgal strains from the lake Massaciuccoli: Screening of common pollutants tolerance pattern and perspectives for their use in biotechnological applications. Front. Plant Sci. 2020, 11, 607651. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.-J.; Chen, C.-Y.; Chang, J.-S. Resource recovery from wastewaters using microalgae-based approaches: A circular bioeconomy perspective. Bioresour. Technol. 2020, 302, 122817. [Google Scholar] [CrossRef]
- Chiellini, C.; Serra, V.; Gammuto, L.; Ciurli, A.; Longo, V.; Gabriele, M. Evaluation of nutraceutical properties of eleven microalgal strains isolated from different freshwater aquatic environments: Perspectives for their application as nutraceuticals. Foods 2022, 11, 654. [Google Scholar] [CrossRef] [PubMed]
- Gorman, D.S.; Levine, R.P. Cytochrome f and plastocyanin: Their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huarancca Reyes, T.; Pompeiano, A.; Ranieri, A.; Volterrani, M.; Guglielminetti, L.; Scartazza, A. Photosynthetic performance of five cool-season turfgrasses under UV-B exposure. Plant Physiol. Biochem. 2020, 151, 181–187. [Google Scholar] [CrossRef]
- Huarancca Reyes, T.; Scartazza, A.; Pompeiano, A.; Guglielminetti, L. Physiological responses of Lepidium meyenii plants to ultraviolet-B radiation challenge. BMC Plant Biol. 2019, 19, 186. [Google Scholar] [CrossRef] [PubMed]
- Team R Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Chen, B.; Dong, J.; Li, B.; Xue, C.; Tetteh, P.A.; Li, D.; Gao, K.; Deng, X. Using a freshwater green alga Chlorella pyrenoidosa to evaluate the biotoxicity of ionic liquids with different cations and anions. Ecotoxicol. Environ. Saf. 2020, 198, 110604. [Google Scholar] [CrossRef]
- Khoshnamvand, M.; Hanachi, P.; Ashtiani, S.; Walker, T.R. Toxic effects of polystyrene nanoplastics on microalgae Chlorella vulgaris: Changes in biomass, photosynthetic pigments and morphology. Chemosphere 2021, 280, 130725. [Google Scholar] [CrossRef]
- Werdel, G.M.; Pandey, L.K.; Bergey, E.A. Cigarette butt effects on diatom health in a stream ecosystem. Aquat. Ecol. 2021, 55, 999–1010. [Google Scholar] [CrossRef]
- Sanders, G.E.; Pallett, K.E. Studies into the differential activity of the hydroxybenzonitrile herbicides. Pestic. Biochem. Physiol. 1986, 26, 116–127. [Google Scholar] [CrossRef]
- Ma, J.; Lin, F.; Wang, S.; Xu, L. Toxicity of 21 herbicides to the green alga Scenedesmus quadricauda. Bull. Environ. Contam. Toxicol. 2003, 71, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, J.; Bai, R.; Ohandja, D.G.; Wong, F.S. Biodegradation of organonitriles by adapted activated sludge consortium with acetonitrile-degrading microorganisms. Water Res. 2007, 41, 3465–3473. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; De Luca, D.; Amoroso, M.; Benfatto, S.; Maestri, S.; Racioppi, C.; Esposito, F.; Ianora, A. New molecular insights on the response of the green alga Tetraselmis suecica to nitrogen starvation. Sci. Rep. 2019, 9, 3336. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Li, Q.; Chen, C.; Zhang, Y.; Liu, Y.; Xu, L.; Zhou, Y.; Li, C.; Zhou, D.; Rittmann, B.E. Benzoic and salicylic acid are the signaling molecules of Chlorella cells for improving cell growth. Chemosphere 2021, 265, 129084. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Jacinto, M.; Guieysse, B.; Mattiasson, B. Combined carbon and nitrogen removal from acetonitrile using algal-bacterial bioreactors. Appl. Microbiol. Biotechnol. 2005, 67, 699–707. [Google Scholar] [CrossRef]
- Rastegari, H.; Ghaziaskar, H.S.; Yalpani, M. Valorization of biodiesel derived glycerol to acetins by continuous esterification in acetic acid: Focusing on high selectivity to diacetin and triacetin with no byproducts. Ind. Eng. Chem. Res. 2015, 54, 3279–3284. [Google Scholar] [CrossRef]
- Margesin, R. Determination of enzyme activities in contaminated soil. In Monitoring and Assessing Soil Bioremediation; Springer: Berlin/Heidelberg, Germany, 2005; Volume 5, pp. 309–320. [Google Scholar]
- Nalder, T.D. Microalgal Lipids, Lipases and Lipase Screening Methods. Ph.D. Thesis, Deakin University, Victoria, Australia, 2014. [Google Scholar]
- Urzica, E.I.; Vieler, A.; Hong-Hermesdorf, A.; Page, M.D.; Casero, D.; Gallaher, S.D.; Kropat, J.; Pellegrini, M.; Benning, C.; Merchant, S.S. Remodeling of membrane lipids in iron-starved chlamydomonas. J. Biol. Chem. 2013, 288, 30246–30258. [Google Scholar] [CrossRef] [Green Version]
- Rana, M.S.; Prajapati, S.K. Stimulating effects of glycerol on the growth, phycoremediation and biofuel potential of Chlorella pyrenoidosa cultivated in wastewater. Environ. Technol. Innov. 2021, 24, 102082. [Google Scholar] [CrossRef]
- Ortega Ramírez, C.A.; Ching, T.; Yoza, B.; Li, Q.X. Glycerol-assisted degradation of dibenzothiophene by Paraburkholderia sp. C3 is associated with polyhydroxyalkanoate granulation. Chemosphere 2022, 291, 133054. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, A.J.; Eliseeva, L.G.; Ibragimova, N.A.; Zelenkov, V.N.; Latushkin, V.V.; Nicheva, D.V. Dataset on the effect of foliar application of different concentrations of silicon dioxide and organosilicon compounds on the growth and biochemical contents of oak leaf lettuce (Lactuca sativa var. crispa) grown in phytotron conditions. Data Br. 2021, 38, 107328. [Google Scholar] [CrossRef] [PubMed]
- Lucie, A.-T.; Solange Patricia, W.; Ephrem, K.-K.; Salomon, N.; Serge Florent, B.-O.; Ponel-Béranger, L.D.; Silla, S.; Olga-Diane, Y.; Jean-Laurent, S.-M.; SECK, D.; et al. The effective insecticidal activity of the two extracts ethyl acetate and hexan of Trichilia gilgiana against Sitophilus zeamaïs. Int. J. Biol. 2016, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Barik, M.; Rawani, A.; Laskar, S.; Chandra, G. Evaluation of mosquito larvicidal activity of fruit extracts of Acacia auriculiformis against the Japanese encephalitis vector Culex vishnui. Nat. Prod. Res. 2019, 33, 1682–1686. [Google Scholar] [CrossRef]
- Jeffryes, C.; Rosenberger, J.; Rorrer, G.L. Fed-batch cultivation and bioprocess modeling of Cyclotella sp. for enhanced fatty acid production by controlled silicon limitation. Algal Res. 2013, 2, 16–27. [Google Scholar] [CrossRef]
- Moore, L.F.; Traquair, J.A. Silicon, a required nutrient for Cladophora glomerata (L) Kütz. (Chlorophyta). Planta 1976, 128, 179–182. [Google Scholar] [CrossRef]
- Van Hoecke, K.; De Schamphelaere, K.A.C.; Van der Meeren, P.; Lucas, S.; Janssen, C.R. Ecotoxicity of silica nanoparticles to the green alga Pseudokirchneriella subcapitata: Importance of surface area. Environ. Toxicol. Chem. 2008, 27, 1948–1957. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Hamouda, R.A.; Nizam, A.A. Biodegradation of crude oil by Scenedesmus obliquus and Chlorella vulgaris growing under heterotrophic conditions. Int. Biodeterior. Biodegradation 2013, 82, 67–72. [Google Scholar] [CrossRef]
- Xaaldi Kalhor, A.; Movafeghi, A.; Mohammadi-Nassab, A.D.; Abedi, E.; Bahrami, A. Potential of the green alga Chlorella vulgaris for biodegradation of crude oil hydrocarbons. Mar. Pollut. Bull. 2017, 123, 286–290. [Google Scholar] [CrossRef]
- Luo, J.; Deng, J.; Cui, L.; Chang, P.; Dai, X.; Yang, C.; Li, N.; Ren, Z.; Zhang, X. The potential assessment of green alga Chlamydomonas reinhardtii CC-503 in the biodegradation of benz(a)anthracene and the related mechanism analysis. Chemosphere 2020, 249, 126097. [Google Scholar] [CrossRef]
- Nydahl, A.C.; King, C.K.; Wasley, J.; Jolley, D.F.; Robinson, S.A. Toxicity of fuel-contaminated soil to Antarctic moss and terrestrial algae. Environ. Toxicol. Chem. 2015, 34, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, K.; Cui, M.; Wu, Y.; Huang, X.; Xue, A.; Deng, X.; Luo, L. Effect of Dibutyl Phthalate on the Tolerance and Lipid Accumulation in the Green Microalgae Chlorella vulgaris. Bull. Environ. Contam. Toxicol. 2018, 101, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-S.; Hong, Y.-H.; Nishikawa, Y.; Kage-Nakadai, E.; Chiou, T.-Y.; Wu, C.-C. Impacts of endocrine disruptor di-n-butyl phthalate ester on microalga Chlorella vulgaris verified by approaches of proteomics and gene ontology. Molecules 2020, 25, 4304. [Google Scholar] [CrossRef]
- Cunha, C.; Paulo, J.; Faria, M.; Kaufmann, M.; Cordeiro, N. Ecotoxicological and biochemical effects of environmental concentrations of the plastic-bond pollutant dibutyl phthalate on Scenedesmus sp. Aquat. Toxicol. 2019, 215, 105281. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Li, Y.; Gao, J. Interaction between three marine microalgae and two phthalate acid esters. Ecotoxicol. Environ. Saf. 2019, 170, 407–411. [Google Scholar] [CrossRef]
- Ye, X.; Lu, Q.; Li, W.; Gao, P.; Hu, B.; Zhang, Z.; Dong, C. Selective production of nicotyrine from catalytic fast pyrolysis of tobacco biomass with Pd/C catalyst. J. Anal. Appl. Pyrolysis 2016, 117, 88–93. [Google Scholar] [CrossRef]
- Jiang, H.J.; Ma, Y.; Qiu, G.J.; Wu, F.L.; Chen, S.L. Biodegradation of nicotine by a novel Strain Shinella sp. HZN1 isolated from activated sludge. J. Environ. Sci. Health. B 2011, 46, 703–708. [Google Scholar] [CrossRef]
- Bonanomi, G.; Maisto, G.; De Marco, A.; Cesarano, G.; Zotti, M.; Mazzei, P.; Libralato, G.; Staropoli, A.; Siciliano, A.; De Filippis, F.; et al. The fate of cigarette butts in different environments: Decay rate, chemical changes and ecotoxicity revealed by a 5-years decomposition experiment. Environ. Pollut. 2020, 261, 114108. [Google Scholar] [CrossRef]
- Oliva, M.; De Marchi, L.; Cuccaro, A.; Pretti, C. Bioassay-based ecotoxicological investigation on marine and freshwater impact of cigarette butt littering. Environ. Pollut. 2021, 288, 117787. [Google Scholar] [CrossRef]
- Green, D.S.; Kregting, L.; Boots, B. Effects of cigarette butts on marine keystone species (Ulva lactuca L. and Mytilus edulis L.) and sediment microphytobenthos. Mar. Pollut. Bull. 2021, 165, 112152. [Google Scholar] [CrossRef]
- Fazeli, M.R.; Tofighi, H.; Madadkar-Sobhani, A.; Shahverdi, A.R.; Nejad-Sattari, T.; Mirzaie, S.; Jamalifar, H. Nicotine inhibition of lycopene cyclase enhances accumulation of carotenoid intermediates by Dunaliella salina CCAP 19/18. Eur. J. Phycol. 2009, 44, 215–220. [Google Scholar] [CrossRef]
- Ishikawa, E.; Abe, H. Lycopene accumulation and cyclic carotenoid deficiency in heterotrophic Chlorella treated with nicotine. J. Ind. Microbiol. Biotechnol. 2004, 31, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Rise, M.; Prengler, M.; Malis Arad, S. Characterization of a nicotine-resistant mutant of Chlorella emersonii. J. Plant Physiol. 1998, 152, 583–585. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Hong, Y.; Zhao, G.-P.; Zhang, H.-K.; Zhai, Q.-Y.; Wang, Q. Microalgae-based swine wastewater treatment: Strain screening, conditions optimization, physiological activity and biomass potential. Sci. Total Environ. 2022, 807, 151008. [Google Scholar] [CrossRef] [PubMed]
- Mysore Doddaiah, K.; Narayan, A.; Gokare Aswathanarayana, R.; Ravi, S. Effect of metabolic inhibitors on growth and carotenoid production in Dunaliella bardawil. J. Food Sci. Technol. 2013, 50, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- King, I.C.; Lorenzi, V.; Blasius, M.E.; Gossett, R. Leachates from cigarette butts can persist in marine sediment. Water Air Soil Pollut. 2021, 232, 38. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Isolation Source | Taxonomic Affiliation | Accession Number | Reference |
|---|---|---|---|---|
| F1 | “Le Morette”, Fucecchio Marshland | Scenedesmaceae | OM311002 and OM310999 | [11] |
| F2 | “Le Morette”, Fucecchio Marshland | Chlamydomonas debaryana | OM311003 | [11] |
| F3 | “Le Morette”, Fucecchio Marshland | Chlamydomonas debaryana | OM311004 | [11] |
| F4 | “Le Morette”, Fucecchio Marshland | Chlorella sorokiniana | OM311005 and OM311000 | [11] |
| R1 | Private terrace in Pisa, water sample | Chlorella sorokiniana | OM311006 | [11] |
| LG1 | Recycle cigarette butts substrate | Chlorella sorokiniana | ON065975 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiellini, C.; Mariotti, L.; Huarancca Reyes, T.; de Arruda, E.J.; Fonseca, G.G.; Guglielminetti, L. Remediation Capacity of Different Microalgae in Effluents Derived from the Cigarette Butt Cleaning Process. Plants 2022, 11, 1770. https://doi.org/10.3390/plants11131770
Chiellini C, Mariotti L, Huarancca Reyes T, de Arruda EJ, Fonseca GG, Guglielminetti L. Remediation Capacity of Different Microalgae in Effluents Derived from the Cigarette Butt Cleaning Process. Plants. 2022; 11(13):1770. https://doi.org/10.3390/plants11131770
Chicago/Turabian StyleChiellini, Carolina, Lorenzo Mariotti, Thais Huarancca Reyes, Eduardo José de Arruda, Gustavo Graciano Fonseca, and Lorenzo Guglielminetti. 2022. "Remediation Capacity of Different Microalgae in Effluents Derived from the Cigarette Butt Cleaning Process" Plants 11, no. 13: 1770. https://doi.org/10.3390/plants11131770