
Molecular Characterization of a New Ecotype of Holoparasitic Plant Orobanche L. on Host Weed Xanthium spinosum L.

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Morphological Characters
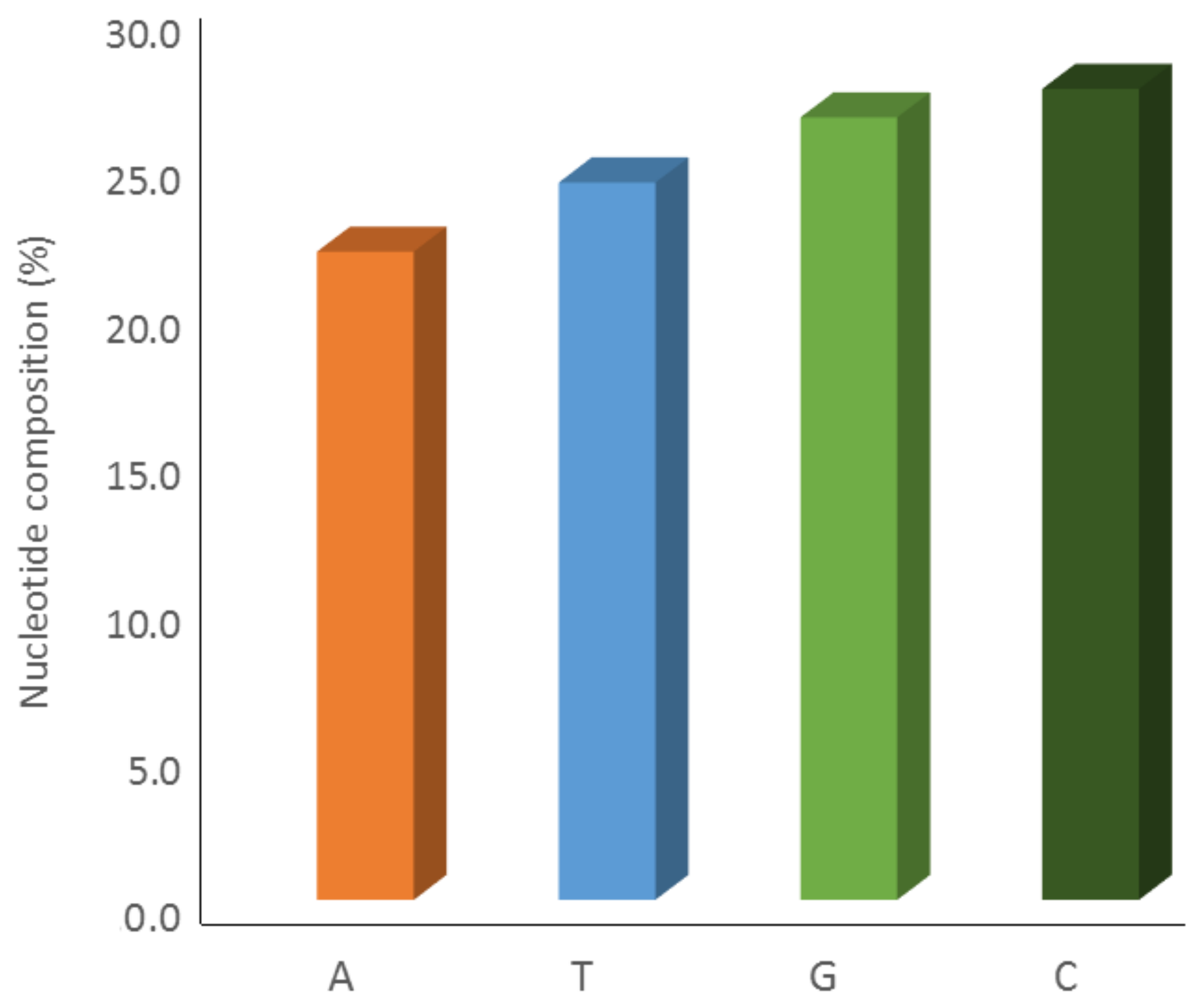
2.2. Sequence Analysis and Identities
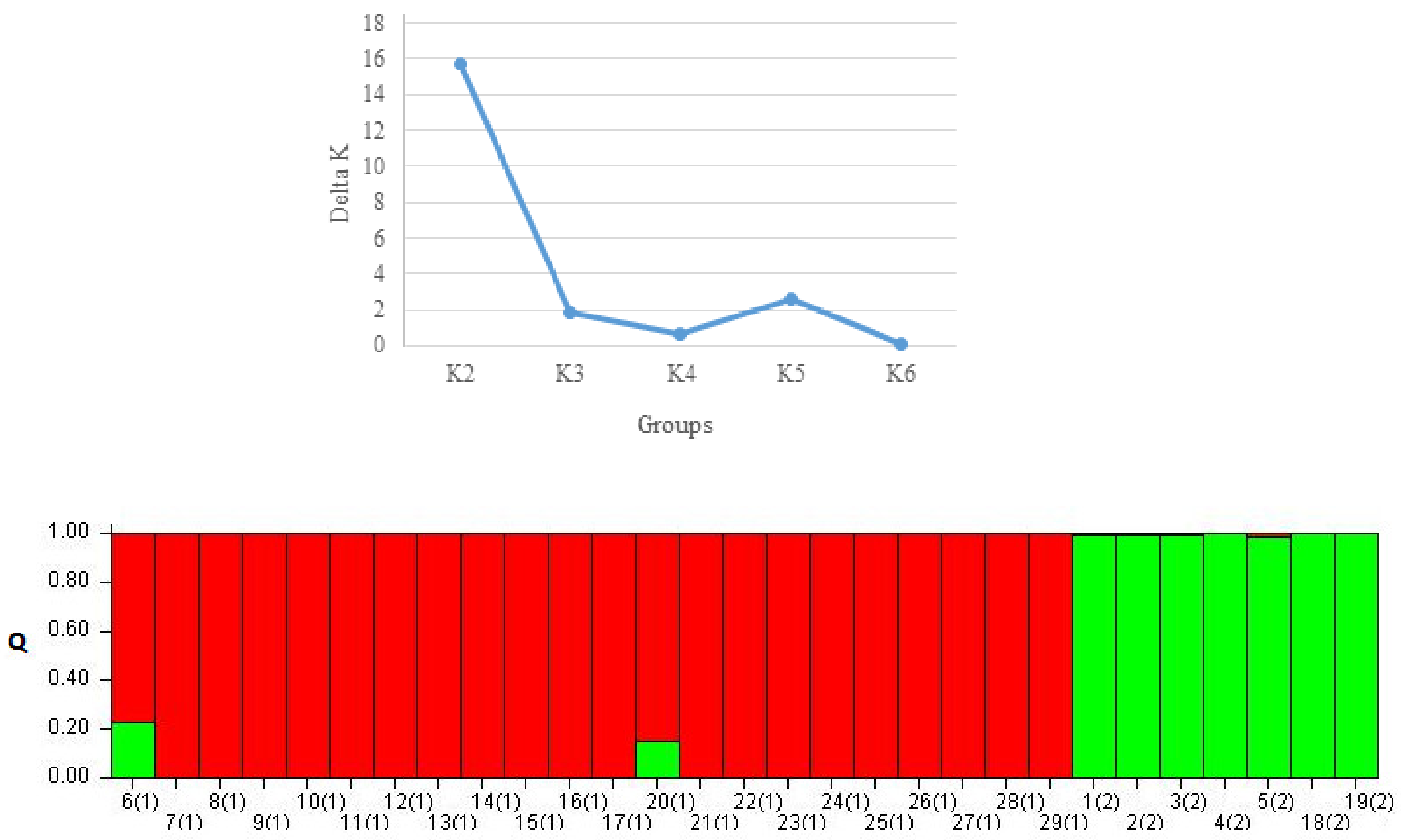
2.3. Genetic Diversity Analysis
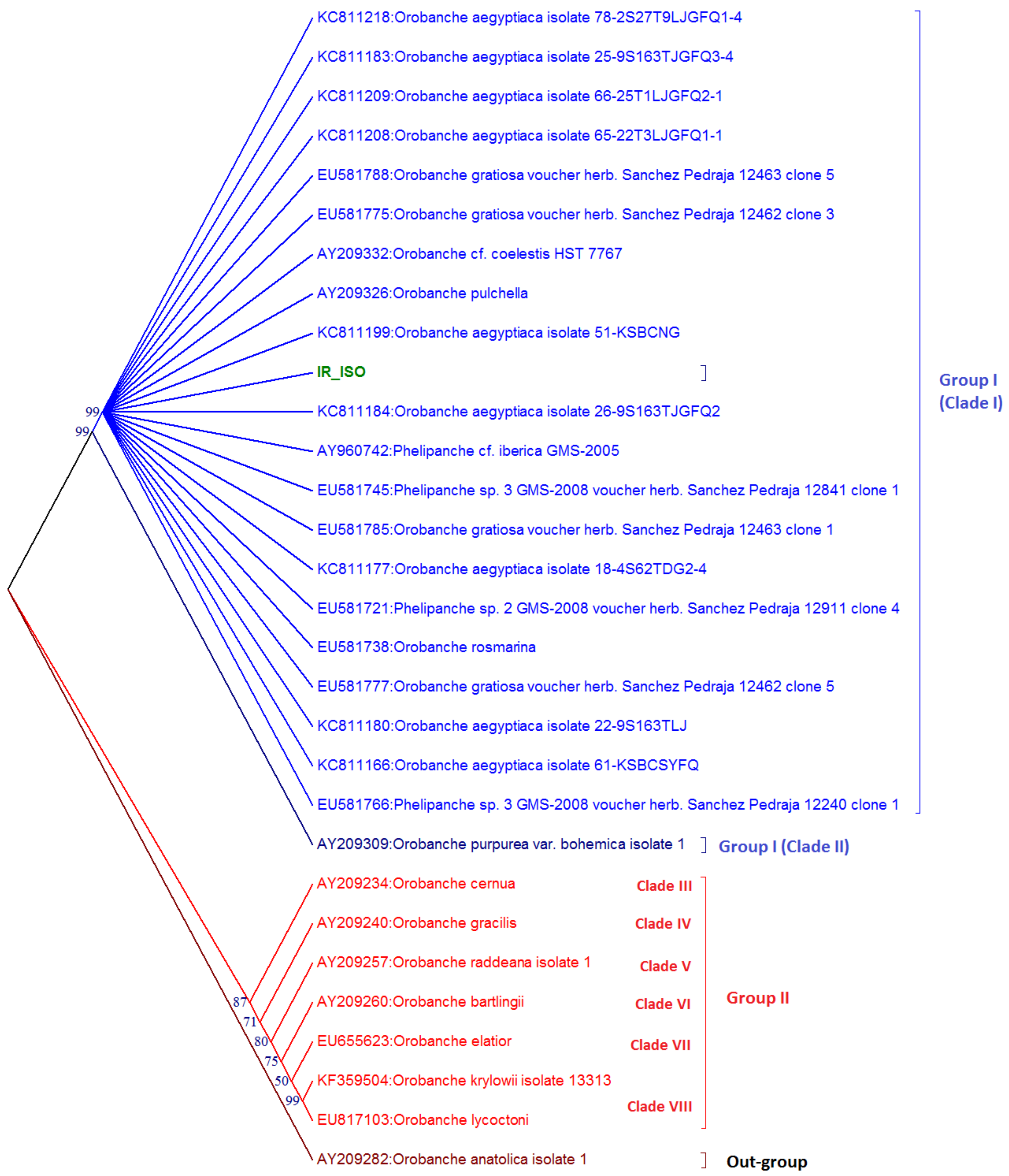
2.4. Cluster Analysis
3. Discussion
4. Materials and Methods
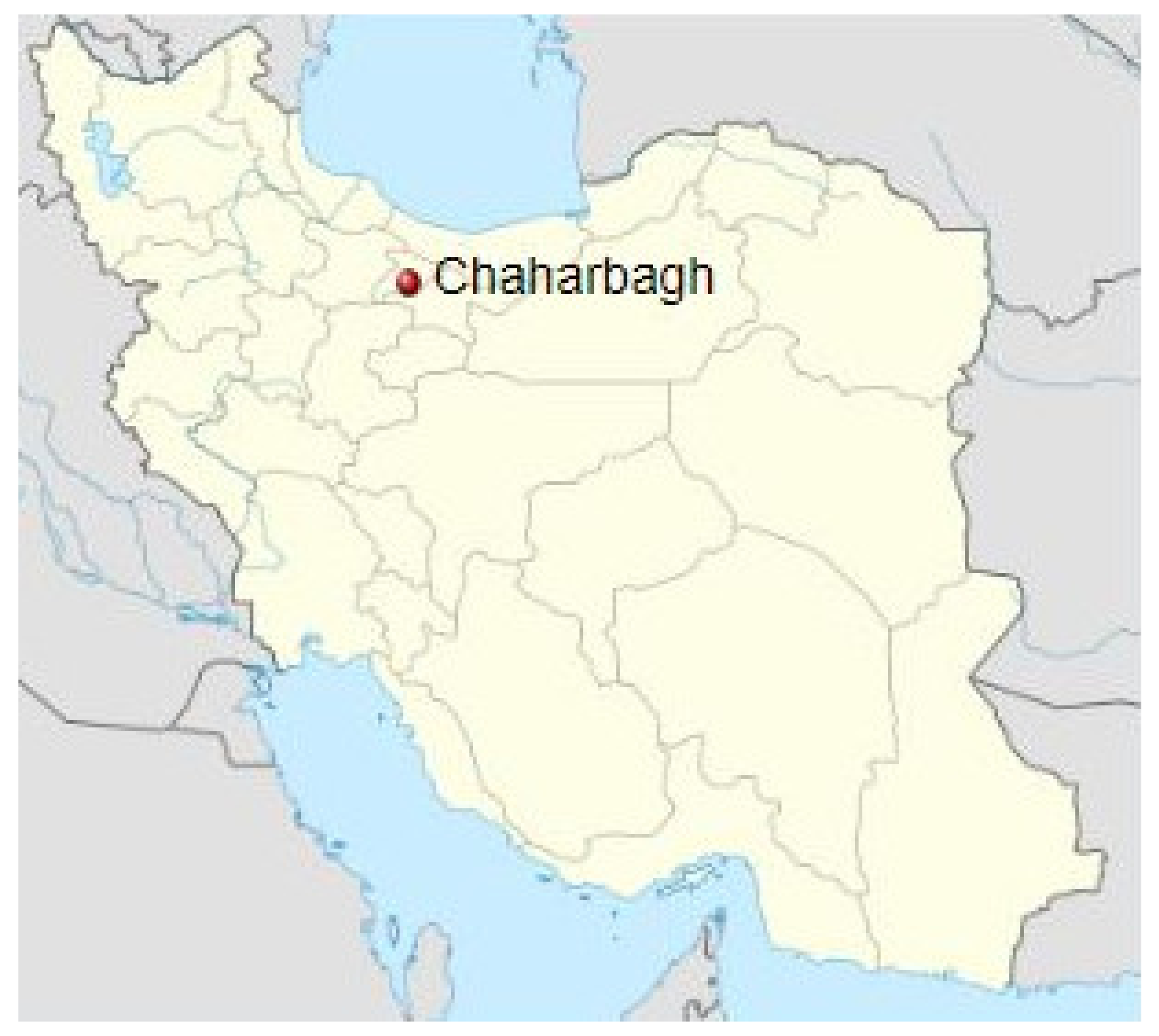
4.1. Plant Material and Sampling
4.2. Extraction of DNA, PCR Condition and DNASequencing
4.3. Sequence Analyses and Phylogenetic Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMOVA | Analysis of molecular variance |
| BLAST | basic local alignment search tool |
| CTAB | cetyltrimethyl ammonium bromide |
| InDel | insertion-deletion mutations |
| ITS | internal transcribed spacer |
| MCL | Maximum composite likelihood |
| MEGA | Molecular Evolutionary Genetics Analysis |
| PCR | polymerase chain reaction. The sequence of targeted region spanning ITS1, 5.8S rRNA and ITS2 was deposited in Gene bank (www.ncbi.nlm.nih.gov) and it’s accessible with accession number MG948171.1. |
References
- McNeal, J.R.; Bennett, J.R.; Wolfe, A.D.; Mathews, S. Phylogeny and origins of holoparasitism in Orobanchaceae. Am. J. Bot. 2013, 100, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.G. Diversity and Classification of Flowering Plants: Eudicots. In Plant Systematics, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 275–448. [Google Scholar] [CrossRef]
- Piwowarczyk, R.; Kwolek, D.; Denysenko, M.; Cygan, M.; Goralski, G.; Slesak, H.; Tuleja, M.; Joachimiak, A.J. Orobanche grenieri (Orobanchaceae), a southwestern European species newly found in Asia. Ann. Bot. Fenn. 2015, 52, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Schneeweiss, G.M.; Frajman, B.; Dakskobler, I. Orobanche lycoctoni Rhiner (Orobanchaceae), a poorly known species of the Central European flora. Candollea 2009, 64, 91–99. [Google Scholar]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWRs) Threatened and Endemic to Italy: Urgent Actions for Protection and Use. Biology 2022, 11, 193. [Google Scholar] [CrossRef]
- Nosratti, I.; Mobli, A.; Mohammadi, G.; Yousefi, A.R.; Sabeti, P.; Chauhan, B.S. The problem of Orobanche spp. and Phelipanche spp. and their management in Iran. Weed Sci. 2020, 68, 555–564. [Google Scholar] [CrossRef]
- Rubiales, D.; Verkleij, J.; Vurro, M.; Murdoch, A.J.; Joel, D.M. Parasitic plant management in sustainable agriculture. Weed Res. 2009, 49, 1–5. [Google Scholar] [CrossRef]
- Erdogan, P. Parasitic Plants in Agriculture and Management. In Parasitic Plants; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar] [CrossRef]
- Habimana, S.; Nduwumuremyi, A.; Chinama, R.J.D. Managementof orobanche in field crops: A review. J. Soil Sci. Plant Nutr. 2014, 14, 43–62. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, W.; Dong, S.; Ren, X.; An, Y.; Lang, M. Induction of seed germination in Orobanche spp. by extracts of traditional Chinese medicinal herbs. Sci. China Life Sci. 2012, 55, 250–260. [Google Scholar] [CrossRef]
- Negewo, T.; Ahmed, S.; Tessema, T.; Tana, T. Biological Characteristics, Impacts, and Management of Crenate Broomrape (Orobanche crenata) in Faba Bean (Vicia faba): A Review. Front. Agron. 2022, 4, 708187. [Google Scholar] [CrossRef]
- Antonova, T.S.; Araslanova, N.M.; Strelnikov, E.A.; Ramazanova, S.A.; Guchetl, S.Z.; Chelyustnikova, T.A. Distribution of highly virulent races of sunflower broomrape (Orobanche cumana Wallr.) in the southern regions of the Russian Federation. Russ. Agric. Sci. 2013, 39, 46–50. [Google Scholar] [CrossRef]
- Fetahaj, R.; Kabashi, B.; Mehmeti, A. Parasitic Plants in Forage Legumes—Medicago sativa L. In Parasitic Plants; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar] [CrossRef]
- Croes, A.F.; Schapendonk, W.; Jansen, F.; Stommen, P.; Bentsink, L.; Zwanenburg, B.; Wullems, G.J. Germination of Orobanche-Aegyptiaca and O-Crenata Is Stimulated Invitro by Isolated Roots—A Comparative Study. In Biology and Management of Orobanche; KIT Press: Amsterdam, The Netherlands, 1994; pp. 157–168. ISBN 90-6832-093-9. [Google Scholar]
- Teryokhin, E.S. Some Biological Peculiarities of Orobanche and Its Host Plants and Their Possible Significance for Control Strategies. In Biology and Management of Orobanche; KIT Press: Amsterdam, The Netherlands, 1994; pp. 94–98. ISBN 90-6832-093-9. [Google Scholar]
- Yousefi, A.R.; Jamshidi, K.; Oveisi, M.; Karimojeni, H.; Pouryosef, M. First Report of Orobanche purpurea on Achillea wilhelmsii in Iran. Plant Dis. 2013, 97, 694. [Google Scholar] [CrossRef]
- Manen, J.F.; Habashi, C.; Jeanmonod, D.; Park, J.M.; Schneeweiss, G.M. Phylogeny and intraspecific variability of holoparasitic Orobanche (Orobanchaceae) inferred from plastid rbcL sequences. Mol. Phylogenet. Evol. 2004, 33, 482–500. [Google Scholar] [CrossRef]
- Park, J.M.; Manen, J.F.; Schneeweiss, G.M. Horizontal gene transfer of a plastid gene in the non-photosynthetic flowering plants Orobanche and Phelipanche (Orobanchaceae). Mol. Phylogenet. Evol. 2007, 43, 974–985. [Google Scholar] [CrossRef]
- Schneeweiss, G.M.; Colwell, A.; Park, J.M.; Jang, C.G.; Stuessy, T.F. Phylogeny of holoparasitic Orobanche (Orobanchaceae) inferred from nuclear ITS sequences. Mol. Phylogenet. Evol. 2004, 30, 465–478. [Google Scholar] [CrossRef]
- Li, X.; Feng, T.; Randle, C.; Schneeweiss, G.M. Phylogenetic Relationships in Orobanchaceae Inferred From Low-Copy Nuclear Genes: Consolidation of Major Clades and Identification of a Novel Position of the Non-photosynthetic Orobanche Clade Sister to All Other Parasitic Orobanchaceae. Front. Plant Sci. 2019, 10, 902. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.R.; Mathews, S. Phylogeny of the parasitic plant family Orobanchaceae inferred from phytochrome A. Am. J. Bot. 2006, 93, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Gaudeul, M.; Siljak-Yakovlev, S.; Jang, T.S.; Rouhan, G. Reconstructing Species Relationships within the Recently Diversified Genus Odontites Ludw. (Orobanchaceae): Evidence for Extensive Reticulate Evolution. Int. J. Plant Sci. 2018, 179, 1–20. [Google Scholar] [CrossRef]
- Frajman, B.; Carlon, L.; Kosachev, P.; Pedraja, O.S.; Schneeweiss, G.M.; Schonswetter, P. Phylogenetic position and taxonomy of the enigmatic Orobanche krylowii (Orobanchaceae), a predominatly Asian species newly found in Albania (SE Europe). Phytotaxa 2013, 137, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, B.G. Phylogenetic Utility of the Internal Transcribed Spacers of Nuclear Ribosomal DNA in Plants: An Example from the Compositae. Mol. Phylogenet. Evol. 1992, 1, 3–16. [Google Scholar] [CrossRef]
- Baldwin, B.G. Molecular Phylogenetics of Calycadenia (Compositae) Based on Its Sequences of Nuclear Ribosomal DNA—Chromosomal and Morphological Evolution Reexamined. Am. J. Bot. 1993, 80, 222–238. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The Its Region of Nuclear Ribosomal DNA—A Valuable Source of Evidence on Angiosperm Phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Germano, J.; Klein, A.S. Species-specific nuclear and chloroplast single nucleotide polymorphisms to distinguish Picea glauca, P-mariana and P-rubens. Theor. Appl. Genet. 1999, 99, 37–49. [Google Scholar] [CrossRef]
- Maggini, F.; Frediani, M.; Gelati, M.T. Nucleotide sequence of the internal transcribed spacers of ribosomal DNA in Picea abies karst. DNA Seq. 2000, 11, 87–89. [Google Scholar] [CrossRef]
- Nashimoto, M.; Ogata, K.; Mishima, Y. In Vitro Sequence-Specific Cleavage in Transcribed Spacer of Mouse Precursor Ribosomal-Rna. J. Biochem. 1988, 103, 992–997. [Google Scholar] [CrossRef]
- Van Nues, R.W.; Rientjes, J.M.; van der Sande, C.A.; Zerp, S.F.; Sluiter, C.; Venema, J.; Planta, R.J.; Raue, H.A. Separate structural elements within internal transcribed spacer 1 of Saccharomyces cerevisiae precursor ribosomal RNA direct the formation of 17S and 26S rRNA. Nucleic Acids Res. 1994, 22, 912–919. [Google Scholar] [CrossRef]
- Yousefi, A.R.; Soheily, E. First Report of Orobanche aegyptiaca on Kalanchoe blossfeldiana in Iran. Plant Dis. 2014, 98, 1287. [Google Scholar] [CrossRef]
- Kalbande, B.B.; Patil, A.S.; Chakrabarty, P.K. An efficient, simple and high throughput protocol for cotton genomic DNA isolation. J. Plant Biochem. Biotechnol. 2016, 25, 437–441. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Rzhetsky, A.; Nei, M. A Simple Method for Estimating and Testing Minimum-Evolution Trees. Mol. Biol. Evol. 1992, 9, 945–967. [Google Scholar]
- Felsenstein, J. Confidence-Limits on Phylogenies—An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial-DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide-Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Accession Number | Scientific Name | Origin | Host Plant | IR-ISO |
|---|---|---|---|---|---|
| 1 | IR-Iso | Orobanche sp. | Iran | Spiny cocklebur | 100.0 |
| 2 | AY209326 | Orobanche pulchella | Georgia | - | 94.46 |
| 3 | AY960742 | Phelipanche cf. iberica | Turkey | 94.30 | |
| 4 | EU581788 | Orobanche gratiosa | Spain | - | 94.30 |
| 5 | EU581777 | Orobanche gratiosa | Spain | - | 94.30 |
| 6 | KC811184 | Orobanche aegyptiaca | China | Tomato | 94.41 |
| 7 | EU581785 | Orobanche gratiosa | Spain | - | 94.14 |
| 8 | EU581775 | Orobanche gratiosa | Spain | - | 94.14 |
| 9 | AY209332 | Orobanche cf. coelestis | Turkey | - | 94.14 |
| 10 | KC811199 | Orobanche aegyptiaca | China | Pumpkin | 94.24 |
| 11 | KC811218 | Orobanche aegyptiaca | China | Tomato | 94.08 |
| 12 | KC811209 | Orobanche aegyptiaca | China | Tomato | 94.08 |
| 13 | KC811208 | Orobanche aegyptiaca | China | Tomato | 94.08 |
| 14 | KC811183 | Orobanche aegyptiaca | China | Tomato | 94.08 |
| 15 | KC811180 | Orobanche aegyptiaca | China | Chili | 94.08 |
| 16 | EU581766 | Phelipanche sp. | Spain | - | 93.81 |
| 17 | KC811177 | Orobanche aegyptiaca | China | Watermelon | 93.91 |
| 18 | EU581745 | Phelipanche sp. | Spain | - | 93.65 |
| 19 | KC811166 | Orobanche aegyptiaca | China | Tomato | 93.77 |
| 20 | EU581721 | Phelipanche sp. | Spain | - | 93.34 |
| 21 | KF359504 | Orobanche krylowii | Russia | - | 77.37 |
| 22 | EU817103 | Orobanche lycoctoni | Slovenia | - | 77.31 |
| 23 | AY209260 | Orobanche bartlingii | Croatia | - | 77.07 |
| 24 | AY209240 | Orobanche gracilis | Morocco | - | 76.42 |
| 25 | AY209282 | Orobanche anatolica | Turkey | - | 93.65 |
| 26 | EU581738 | Orobanche rosmarina | France | - | 76.97 |
| 27 | AY209257 | Orobanche raddeana | Georgia | - | 76.26 |
| 28 | AY209234 | Orobanche cernua | Jordan | - | 88.47 |
| 29 | AY209309 | Orobanche purpurea | Germany | - | 76.19 |
| 30 | EU655623 | Orobanche elatior | France | - | 77.13 |
| A | T | C | G | |
|---|---|---|---|---|
| A | - | 3.51 | 4.05 | 14.26 |
| T | 3.16 | - | 24.09 | 3.87 |
| C | 3.16 | 20.87 | - | 3.87 |
| G | 11.63 | 3.51 | 4.05 | - |
| m | n | S | ps | θ | π | D | |
|---|---|---|---|---|---|---|---|
| Group I | 8 | 579 | 126 | 0.218 | 0.084 | 0.069 | −0.945 |
| Group II | 22 | 584 | 147 | 0.252 | 0.069 | 0.026 | −2.556 |
| Overall | 30 | 560 | 212 | 0.3791 | 0.096 | 0.088 | −0.304 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousefi, A.R.; Ahmadikhah, A.; Fotovat, R.; Rohani, L.; Soheily, F.; Uberti, D.L.; Mastinu, A. Molecular Characterization of a New Ecotype of Holoparasitic Plant Orobanche L. on Host Weed Xanthium spinosum L. Plants 2022, 11, 1406. https://doi.org/10.3390/plants11111406
Yousefi AR, Ahmadikhah A, Fotovat R, Rohani L, Soheily F, Uberti DL, Mastinu A. Molecular Characterization of a New Ecotype of Holoparasitic Plant Orobanche L. on Host Weed Xanthium spinosum L. Plants. 2022; 11(11):1406. https://doi.org/10.3390/plants11111406
Chicago/Turabian StyleYousefi, Ali Reza, Asadollah Ahmadikhah, Reza Fotovat, Leila Rohani, Foad Soheily, Daniela Letizia Uberti, and Andrea Mastinu. 2022. "Molecular Characterization of a New Ecotype of Holoparasitic Plant Orobanche L. on Host Weed Xanthium spinosum L." Plants 11, no. 11: 1406. https://doi.org/10.3390/plants11111406