
The (Bio)chemical Base of Flower Colour in Bidens ferulifolia

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
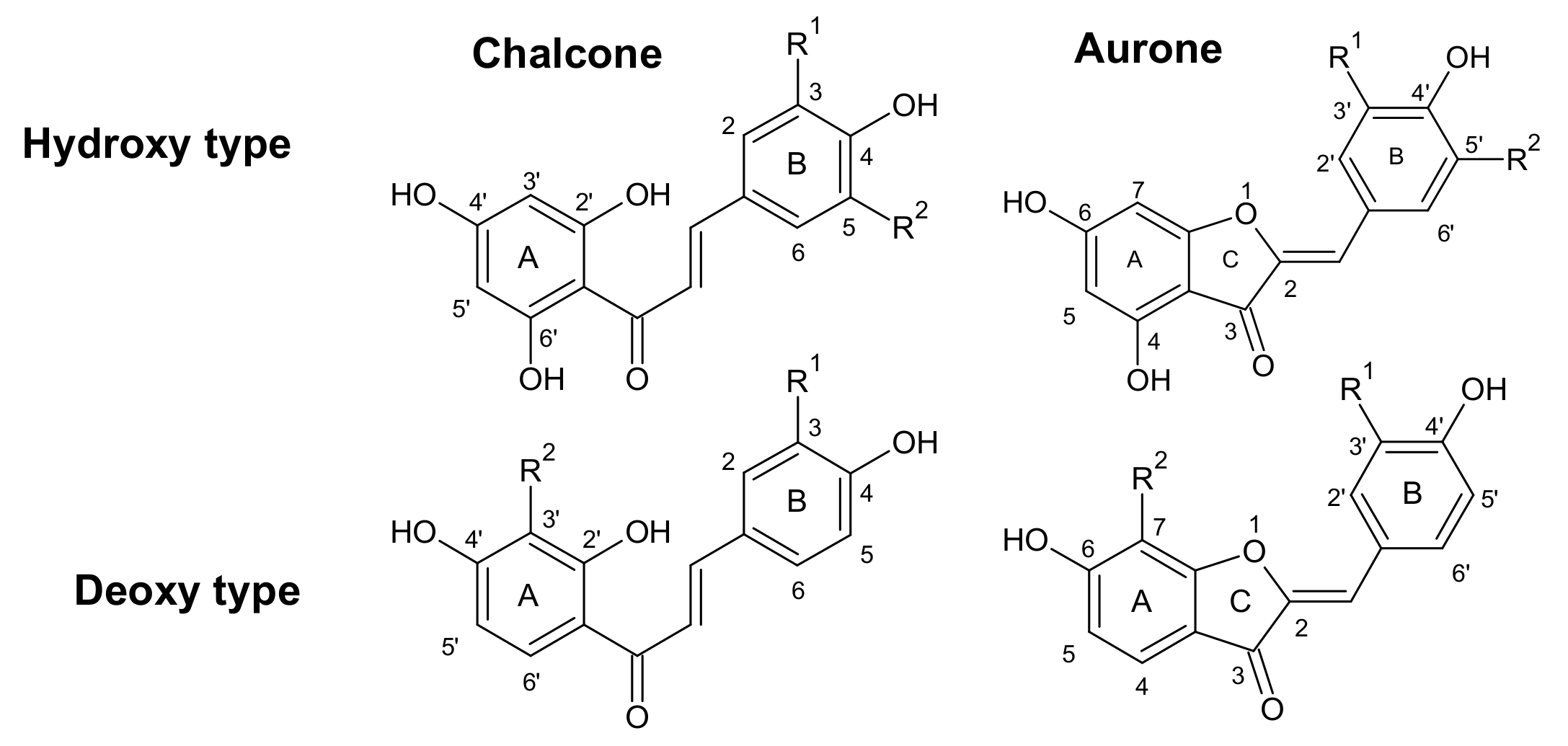
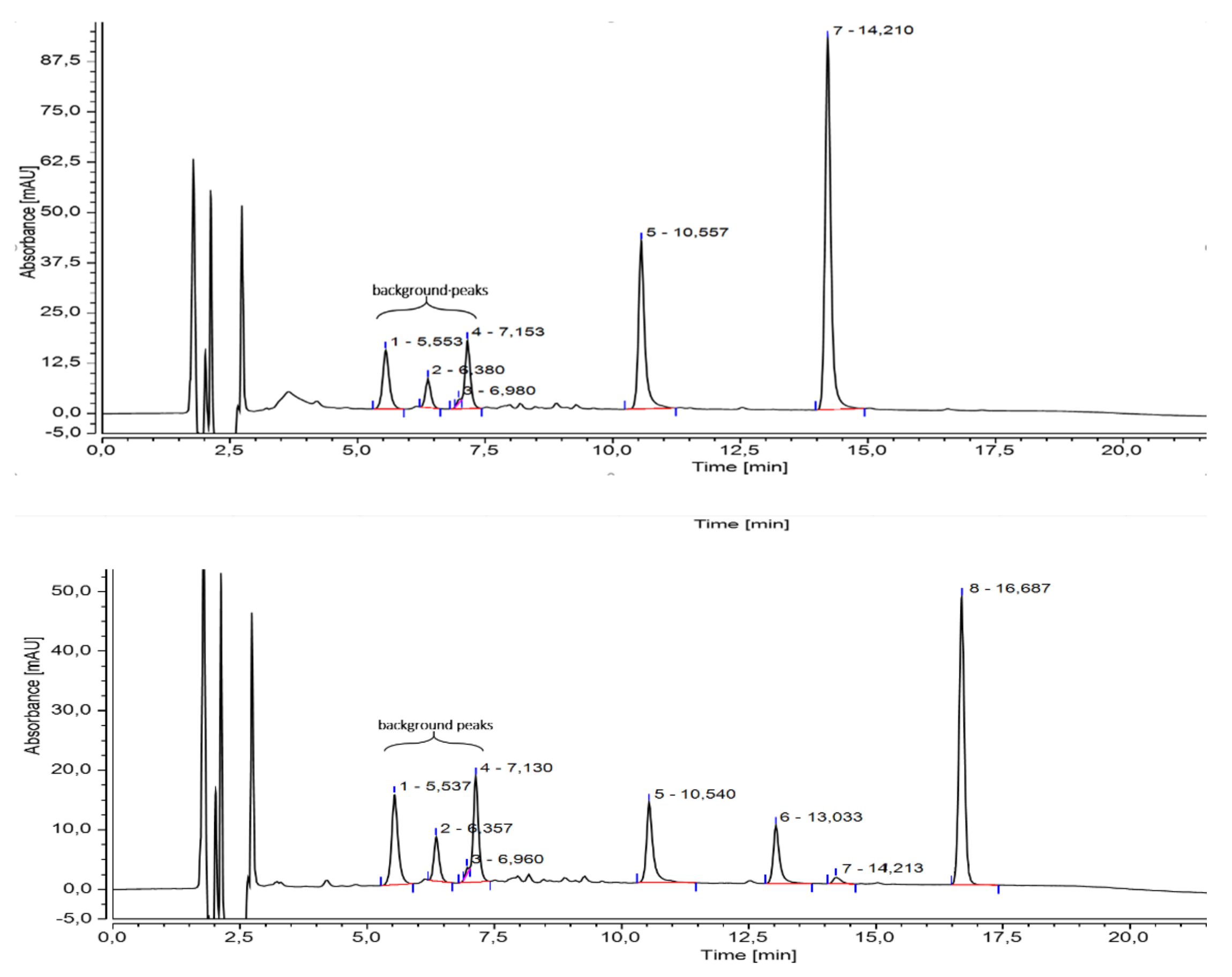
2.1. Anthochlor and Flavonoid Pigment Composition
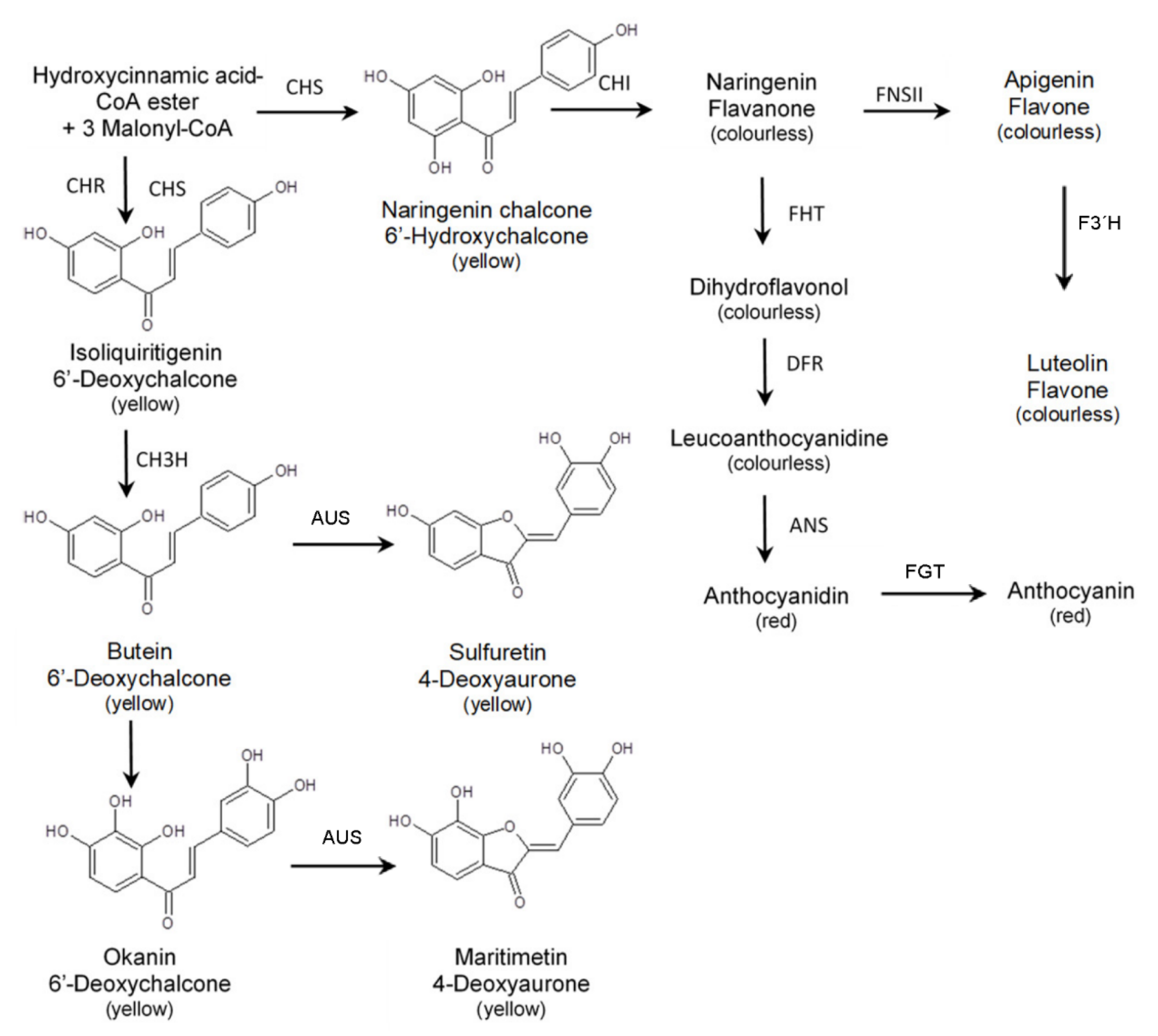
2.2. Enzyme Activities of the Flavonoid and Anthochlor Pathway
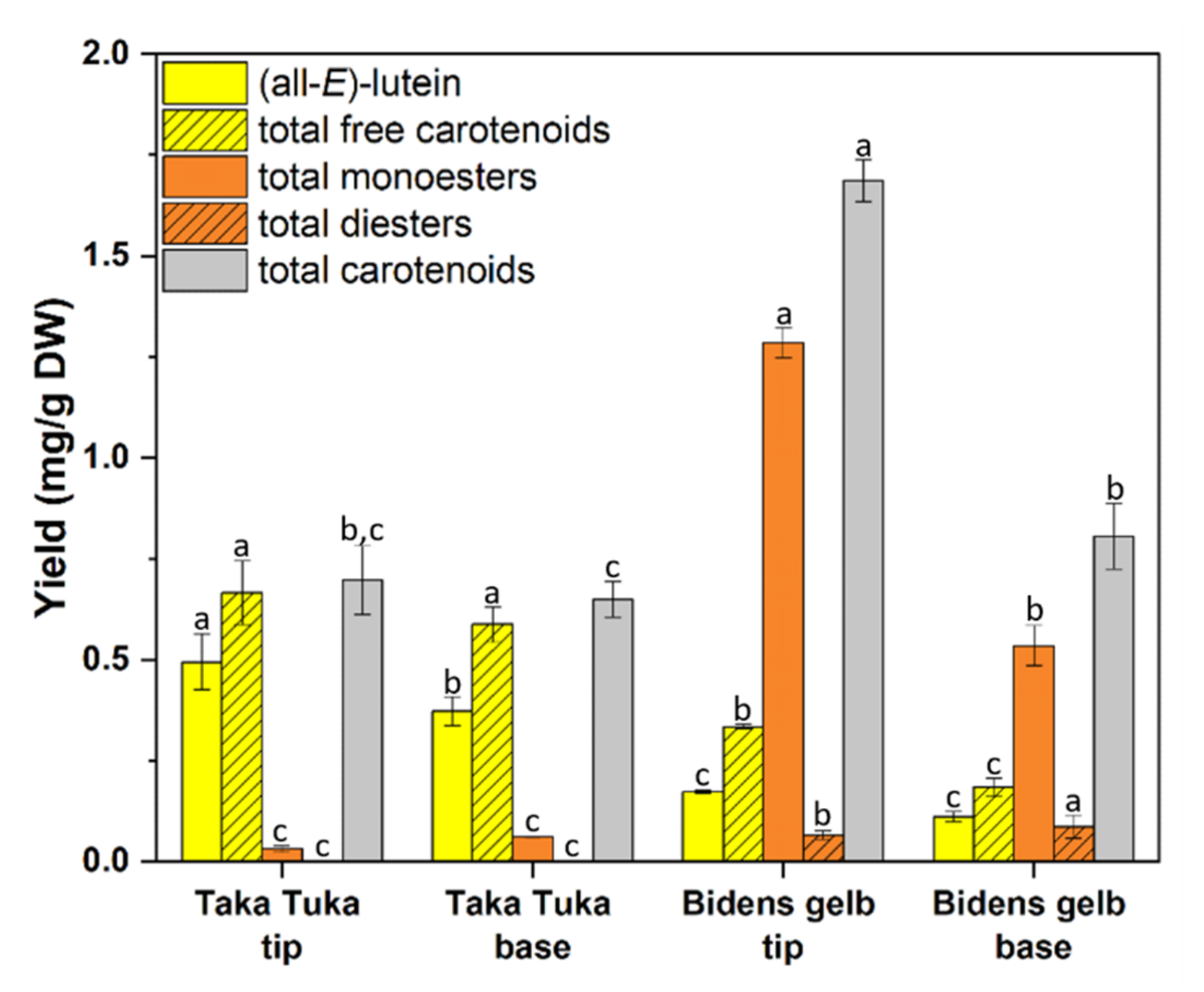
2.3. Carotenoids
3. Discussion
3.1. Anthocyanins
3.2. Anthochlors
3.3. Carotenoids
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals
4.3. Anthochlor and Flavonoid Analysis
4.4. Carotenoid Analysis
4.5. Enzyme Assays
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, D.J.; Mort, M.E. Phylogeny of Eastern North American Coreopsis (Asteraceae-Coreopsideae): Insights from nuclear and plastid sequences, and comments on character evolution. Am. J. Botany 2005, 92, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-J.; Choi, K.-S.; Jansen, R.K. Two chloroplast DNA inversions originated simultaneously during the early evolution of the sunflower family (Asteraceae). Mol. Biol. Evol. 2005, 22, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Crowe, D.R.; Parker, W.H. Hybridization and agamospermy of Bidens in northwestern Ontario. Taxon 1981, 30, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, M. Bidens are biding their time no longer. Available online: https://rotarybotanicalgardens.org/bidens-are-biding-their-time-no-longer/ (accessed on 11 November 2021).
- García-Mendoza, A.; del Castillo, J.M. Diversidad Florística de Oaxaca: De Musgos a Angispermas; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2011. [Google Scholar]
- First registered non-yellow Bidens available in Europe. Available online: http://www.newplantsandflowers.com/first-registered-non-yellow-bidens-available-in-europe/ (accessed on 11 November 2021).
- Brouillard, R.; Dangles, O. Flavonoids and flower colour. In The Flavonoids—Advances in Research; Chapman & Hall: London, UK, 1994; pp. 565–586. [Google Scholar]
- Scogin, R.; Zakar, K. Anthochlor pigments and floral UV patterns in the genus Bidens. Biochem. Syst. Ecol. 1976, 4, 165–167. [Google Scholar] [CrossRef]
- Harborne, J.B.; Smith, D.M. Anthochlors and other flavonoids as honey guides in the Compositae. Biochem. Syst. Ecol. 1978, 6, 287–291. [Google Scholar] [CrossRef]
- Miosic, S.; Knop, K.; Hölscher, D.; Greiner, J.; Gosch, C.; Thill, J.; Kai, M.; Shrestha, B.K.; Schneider, B.; Crecelius, A.C. 4-Deoxyaurone formation in Bidens ferulifolia (Jacq.) DC. PLoS ONE 2013, 8, e61766. [Google Scholar] [CrossRef] [Green Version]
- McCrea, K.D.; Levy, M. Photographic visualization of floral colors as perceived by honeybee pollinators. Am. J. Bot. 1983, 70, 369–375. [Google Scholar] [CrossRef]
- Briscoe, A.D.; Chittka, L. The Evolution of Color Vision in Insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B. Comparative Biochemistry of Flavonoids; Academic Press: London, UK, 1967. [Google Scholar]
- Bomati, A.K.; Austin, M.B.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structural elucidation of chalcone reductase and implications for deoxychalcone biosynthesis. J. Biol. Chem. 2005, 280, 30496–30503. [Google Scholar] [CrossRef] [Green Version]
- Halbwirth, H.; Muster, G.; Stich, K. Unraveling the biochemical base of dahlia flower coloration. Nat. Prod. Commun. 2008, 3, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Bohm, B.A. The minor flavonoids. In The Flavonoids: Advances in Research Since 1980; Chapman & Hall: London, UK, 1988. [Google Scholar]
- Bohm, B.A. The minor flavonoids. In The Flavonoids: Advances in Research Since 1986; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Boucherle, B.; Peuchmaur, M.; Boumendjel, A.; Haudecoeur, R. Occurrences, biosynthesis and properties of aurones as high-end evolutionary products. Phytochemistry 2017, 142, 92–111. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Sato, T.; Fukui, Y.; Yonekura-Sakakibara, K.; Hayashi, K.; Tanaka, Y.; Kusumi, T.; Nishino, T. Aureusidin synthase: A polyphenol oxidase homolog responsible for flower coloration. Science 2000, 290, 1163–1165. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Sato, M.; Fukui, Y.; Yonekura-Sakakibara, K.; Hayashi, H.; Tanaka, Y.; Kusumi, T.; Nishino, T. Specifity analysis and mechanism of aurone synthasis catalyzed by aureusidin synthase, a polyphenol oxidase homolog responsible for flower coloration. FEBS Lett. 2001, 499, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Nakayama, T.; Kikuchi, S.; Fukui, Y.; Yonekura-Sakakibara, K.; Ueda, T.; Nishino, T.; Tanaka, Y.; Kusumi, T. Enzymatic formation of aurones in the extracts of yellow snapdragon flowers. Plant Sci. 2001, 160, 229–236. [Google Scholar] [CrossRef]
- Ono, E.; Hatayama, M.; Isono, Y.; Sato, T.; Watanabe, R.; Yonekura-Sakakibara, K.; Fukuchi-Mizutani, M.; Tanaka, Y.; Kusumi, T.; Nishino, T.; et al. Localization of a flavonoid biosynthetic polyphenol oxidase in vacuoles. Plant J. 2006, 45, 133–143. [Google Scholar] [CrossRef]
- Ono, E.; Fukuchi-Mizutani, M.; Nakamura, N.; Fukui, Y.; Yonekura-Sakakibara, K.; Yamaguchi, M.; Nakayama, T.; Tanaka, T.; Kusumi, T.; Tanaka, Y. Yellow flower generated by expression of the aurone biosynthetic pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 11075–11080. [Google Scholar] [CrossRef] [Green Version]
- Molitor, C.; Mauracher, S.G.; Pargan, S.; Mayer, R.L.; Halbwirth, H.; Rompel, A. Latent and active aurone synthase from petals of C. grandiflora: A polyphenol oxidase with unique characteristics. Planta 2015, 242, 519–537. [Google Scholar] [CrossRef] [Green Version]
- Kaintz, C.; Molitor, C.; Thill, J.; Kampatsikas, I.; Michael, C.; Halbwirth, H.; Rompel, A. Cloning and functional expression in E. coli of a polyphenol oxidase transcript from C. grandiflora involved in aurone formation. FEBS Lett. 2014, 588, 3417–3426. [Google Scholar] [CrossRef] [Green Version]
- Schlangen, K.; Miosic, S.; Thill, J.; Halbwirth, H. Cloning, functional expression, and characterization of a chalcone-3-hydroxylase from Cosmos sulphureus. J. Exp. Bot. 2010, 61, 3451–3459. [Google Scholar] [CrossRef] [Green Version]
- Schlangen, K.; Miosic, S.; Halbwirth, H. Allelic variants from Dahlia variabilis encode flavonoid 3′-hydroxylases with functional differences in chalcone 3-hydroxylase activity. Arch. Biochem. Biophys. 2010, 494, 40–45. [Google Scholar] [CrossRef]
- Molitor, C.; Mauracher, S.G.; Rompel, A. Aurone synthase is a catechol oxidase with hydroxylase activity and provides insights into the mechanism of plant polyphenol oxidases. Proc. Natl. Acad. Sci. USA 2016, 113, 1806–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Hao, Z.; Tao, J. Effects of shade on plant growth and flower quality in the herbaceous peony (Paeonia lactiflora Pall.). Plant Physiol. Biochem. 2012, 61, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D. Regulation of flower pigmentation and growth: Multiple signaling pathways control anthocyanin synthesis in expanding petals. Physiol. Plant. 2000, 110, 152–157. [Google Scholar] [CrossRef]
- Walliser, B.; Lucaciu, C.R.; Molitor, C.; Marinovic, S.; Nitarska, D.A.; Aktaş, D.; Rattei, T.; Kampatsikas, I.; Stich, K.; Haselmair-Gosch, C. Dahlia variabilis cultivar ‘Seattle’as a model plant for anthochlor biosynthesis. Plant Physiol. Biochem. 2021, 159, 193–201. [Google Scholar] [CrossRef]
- Berim, A.; Gang, D.R. The roles of a flavone-6-hydroxylase and 7-O-demethylation in the flavone biosynthetic network of sweet basil. J. Biol. Chem. 2013, 288, 1795–1805. [Google Scholar] [CrossRef] [Green Version]
- Halbwirth, H.; Stich, K. An NADPH and FAD dependent enzyme catalyzes hydroxylation of flavonoids in position 8. Phytochemistry 2006, 67, 1080–1087. [Google Scholar] [CrossRef]
- Ohmiya, A. Diversity of carotenoid composition in flower petals. Jpn. Agric. Res. Q.: JARQ 2011, 45, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Valadon, L.; Mummery, R.S. Carotenoids of Compositae flowers. Phytochemistry 1971, 10, 2349–2353. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus Handbook for Carotenoid Analysis; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2004; Volume 2. [Google Scholar]
- Rodrigues, D.B.; Mercadante, A.Z.; Mariutti, L.R.B. Marigold carotenoids: Much more than lutein esters. Food Res. Int. 2019, 119, 653–664. [Google Scholar] [CrossRef]
- Haselmair-Gosch, C.; Miosic, S.; Nitarska, D.; Walliser, B.; Roth, B.L.; Paltram, R.; Lucaciu, R.C.; Eidenberger, L.; Rattei, T.; Olbricht, K.; et al. Great Cause—Small Effect: Undeclared genetically engineered orange petunias harbor an inefficient dihydroflavonol 4-reductase. Front. Plant Sci. 2018, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thill, J.; Miosic, S.; Ahmed, R.; Schlangen, K.; Muster, G.; Stich, K.; Halbwirth, H. ‘Le Rouge et le Noir’: A decline in flavone formation correlates with the rare color of black dahlia (Dahlia variabilis hort.) flowers. BMC Plant Biol. 2012, 12, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doppler, P.; Kornpointner, C.; Halbwirth, H.; Remias, D.; Spadiut, O. Tetraedron minimum, First reported member of Hydrodictyaceae to accumulate secondary carotenoids. Life 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.C.; Mercadante, A.Z. New method for carotenoid extraction and analysis by HPLC-DAD-MS/MS in freeze-dried citrus and mango pulps. J. Braz. Chem. Soc. 2018, 29, 205–215. [Google Scholar] [CrossRef]
- Petry, F.C.; Mercadante, A.Z. Composition by LC-MS/MS of new carotenoid esters in mango and citrus. J. Agric. Food Chem. 2016, 64, 8207–8224. [Google Scholar] [CrossRef]
- Mercadante, A.Z.; Rodrigues, D.B.; Petry, F.C.; Mariutti, L.R.B. Carotenoid esters in foods—A review and practical directions on analysis and occurrence. Food Res. Int. 2017, 99, 830–850. [Google Scholar] [CrossRef]
- Bonaccorsi, I.; Cacciola, F.; Utczas, M.; Inferrera, V.; Giuffrida, D.; Donato, P.; Dugo, P.; Mondello, L. Characterization of the pigment fraction in sweet bell peppers (Capsicum annuum L.) harvested at green and overripe yellow and red stages by offline multidimensional convergence chromatography/liquid chromatography–mass spectrometry. J. Sep. Sci. 2016, 39, 3281–3291. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical procedures for agricultural research. R Package Version 2014, 1, 1–4. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 1 April 2022).
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cv./Line | Petal Part | Colour | Anthochlors | Flavones | Anthocyanins | Total |
|---|---|---|---|---|---|---|
| µg/g fresh weight | ||||||
| 9157 | entirety | Purple | n.d.l | 2944b | 345d | 4632 |
| Firewheel | base | Red | 6184efgh | 787fg | 249e | 7381 |
| tip | Red | 5272ghi | 1090e | 1806a | 8753 | |
| Painted Red | base | Red- Yellow | 9223abcd | 594hi | 19f | 9841 |
| tip | Red | 8334bcde | 559hi | 1597b | 10560 | |
| Bidens gelb | entirety | Yellow | 1987kl | 152lm | n.d.f | 2139 |
| 3176 | base | Yellow | 3556ijk | 338jk | n.d.f | 3894 |
| tip | Red | 11095a | 1287d | n.d.f | 12382 | |
| 3277 (Star type) | edge | Yellow | 6269efgh | 193kl | n.d.f | 6461 |
| star | Red | 8315bcde | 794fg | n.d.f | 9109 | |
| Blazing Embers | base | Yellow | 7202cdefg | 685gh | n.d.f | 7886 |
| tip | Red | 3253ijk | 176klm | n.d.f | 3428 | |
| Painted Yellow | base | Red | 2015kl | n.d.m | 517c | 2717 |
| tip | Yellow | 1577kl | n.d.m | 250e | 1843 | |
| Giant | base | Yellow | 8110bcdef | 533hi | 5f | 8651 |
| tip | Yellow | 1593kl | 39lm | 28f | 1665 | |
| Mega Charm | base | Yellow | 6829defgh | 764fg | n.d.f | 7593 |
| tip | Yellow | 2003kl | 170lm | n.d.f | 2173 | |
| Eldoro Red Nails | base | Yellow | 4514hij | 458ij | n.d.f | 4972 |
| tip | Red | 4788ghij | 421ij | 1627b | 7300 | |
| Eldoro Y. Red Star | star | Red | 10000ab | 583hi | 353d | 10966 |
| edge | Yellow | 1545kl | 46lm | n.d.f | 1591 | |
| Taka Tuka | base | Yellow | 9513abc | 901f | n.d.f | 35998 |
| tip | Cream | 2543jkl | 112lm | n.d.f | 11088 | |
| 3267 | base | Yellow | 9291abcd | 1371d | n.d.f | 41143 |
| tip | Cream | 5627fghi | 416ij | n.d.f | 18807 | |
| Beedance White | entirety | White | n.d. l | 3252a | 7f | 3484 |
| 9163 | entirety | White | n.d. l | 1806c | n.d.f | 1852 |
| Cv./Line | Petal | Colour | CHS | FHT | DFR | FNSII | CH3H | CH3′H |
|---|---|---|---|---|---|---|---|---|
| Section | Specific Activity (nmols−1kg−1) | |||||||
| 9157 | entirety | Purple | 852 bcd | 321 b | 1286 a | 183 de | 2121 a | 1642 ghi |
| Firewheel | base | Red | 383 ghi | 90 ef | 473 cd | 417 ab | 3663 a | 7472 a |
| tip | Red | 253 i | 145 de | 892 b | 469 a | 4463 a | 3367 defgh | |
| Painted Red | base | Red- Yellow | 774 cde | n.d.g | 605 c | 435 a | 3568 a | 4701 bcd |
| tip | Red | 658 cdefg | 546 a | 975 b | 391 abc | 3704 a | 5989 ab | |
| Bidens gelb | entirety | Yellow | 465 fghi | 69 fg | n.d. h | 162 de | 2988 a | 3414 defgh |
| 3176 | base | Yellow | 219 i | 219 c | 11 h | 400 abc | 3519 a | 4000 cdef |
| tip | Red | 250 i | 249 c | 188 fg | 418 ab | 5589 a | 3980 cdef | |
| 3277 (Star type) | edge | Yellow | 489 efghi | n.d.g | 55 gh | 383 abc | 4386 a | 4762 bcd |
| star | Red | 734 cdef | 381 b | 240 ef | 315 abcd | 3781 a | 5619 bc | |
| Blazing Embers | base | Yellow | 226 i | n.d.g | 20 g | 118 de | 2310 a | 3159 defghi |
| tip | Red | 276 hi | 242 c | 153 fgh | 179 de | 1996 a | 2184 fghi | |
| Painted Yellow | base | Red | 446 fghi | 180 cd | 377 de | 231 bcde | 2619 a | 2454 efghi |
| tip | Yellow | 483 cde | n.d.g | 18 h | 131 de | 2870 a | 3101 cdef | |
| Giant | base | Yellow | 480 fghi | n.d.g | 255 ef | 172 de | 3640 a | 3958 cdef |
| tip | Yellow | 403 ghi | n.d.g | 559 c | 104 e | 1769 a | 3505 defgh | |
| Mega Charm | base | Yellow | 430 ghi | n.d.g | n.d. h | 46 e | 2795 a | 4135 cde |
| tip | Yellow | 217 i | n.d.g | n.d. h | 70 e | 1730 a | 1366 i | |
| Eldoro Red Nails | base | Yellow | 412 ghi | n.d.g | n.d. h | 218 cde | 3880 a | 3126 defghi |
| tip | Red | 436 ghi | 375 b | 1028 b | 221 cde | 3617 a | 3400 defgh | |
| Eldoro Yellow Red Star | star | Red | 561 efghi | 187 cd | 435 d | 184 de | 2878 a | 2437 efghi |
| edge | Yellow | 490 defgh | n.d.g | n.d. h | 45 e | 1363 a | 1751 ghi | |
| Taka Tuka | base | Yellow | 827 bcd | 30 fg | n.d. h | 137 de | 2683 a | 2736 efghi |
| tip | Cream | 786 bcd | n.d.g | n.d. h | 60 e | 1732 a | 3376 defgh | |
| 3267 | base | Yellow | 1663 a | 43 fg | n.d. h | 131 de | 3371 a | 4167 cde |
| tip | Cream | 923 bcd | n.d.g | n.d. h | 55 e | 1578 a | 3347 defgh | |
| Beedance W. | entirety | White | 1707 a | 56 fg | n.d. h | 69 e | 1609 a | 2157 fghi |
| 9163 | entirety | White | 1064 b | 101 ef | n.d. h | 159 de | 3273 a | 1543 hi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walliser, B.; Marinovic, S.; Kornpointner, C.; Schlosser, C.; Abouelnasr, M.; Hutabarat, O.S.; Haselmair-Gosch, C.; Molitor, C.; Stich, K.; Halbwirth, H. The (Bio)chemical Base of Flower Colour in Bidens ferulifolia . Plants 2022, 11, 1289. https://doi.org/10.3390/plants11101289
Walliser B, Marinovic S, Kornpointner C, Schlosser C, Abouelnasr M, Hutabarat OS, Haselmair-Gosch C, Molitor C, Stich K, Halbwirth H. The (Bio)chemical Base of Flower Colour in Bidens ferulifolia . Plants. 2022; 11(10):1289. https://doi.org/10.3390/plants11101289
Chicago/Turabian StyleWalliser, Benjamin, Silvija Marinovic, Christoph Kornpointner, Christopher Schlosser, Mustafa Abouelnasr, Olly Sanny Hutabarat, Christian Haselmair-Gosch, Christian Molitor, Karl Stich, and Heidi Halbwirth. 2022. "The (Bio)chemical Base of Flower Colour in Bidens ferulifolia " Plants 11, no. 10: 1289. https://doi.org/10.3390/plants11101289