
Individual and Combined Effects of Booting and Flowering High-Temperature Stress on Rice Biomass Accumulation

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Effect of High Temperature at Booting, Flowering, and Combined Stages on Photosynthetic Property Parameters
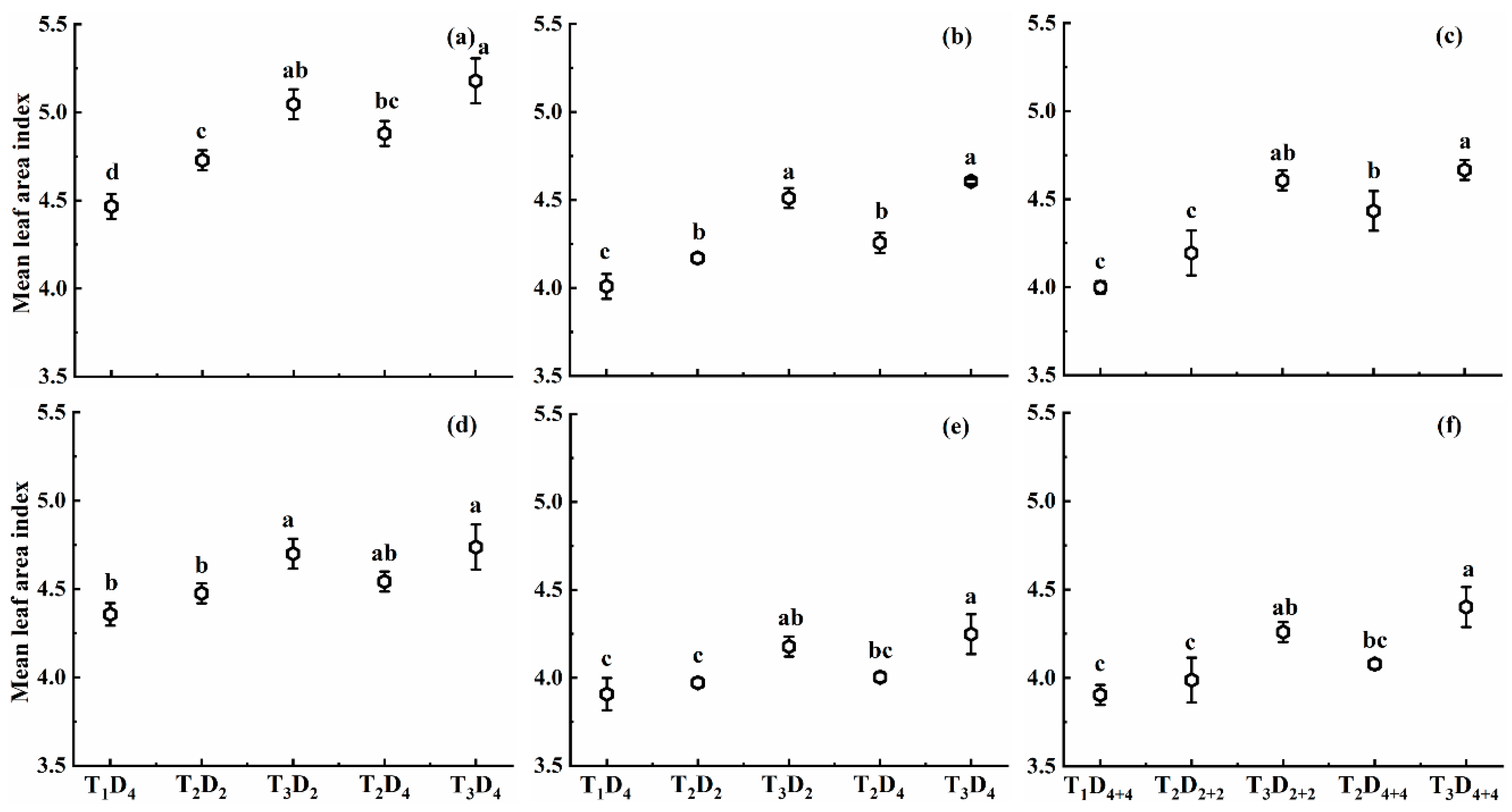
2.1.1. Effect of High Temperature on Mean Leaf Area Index
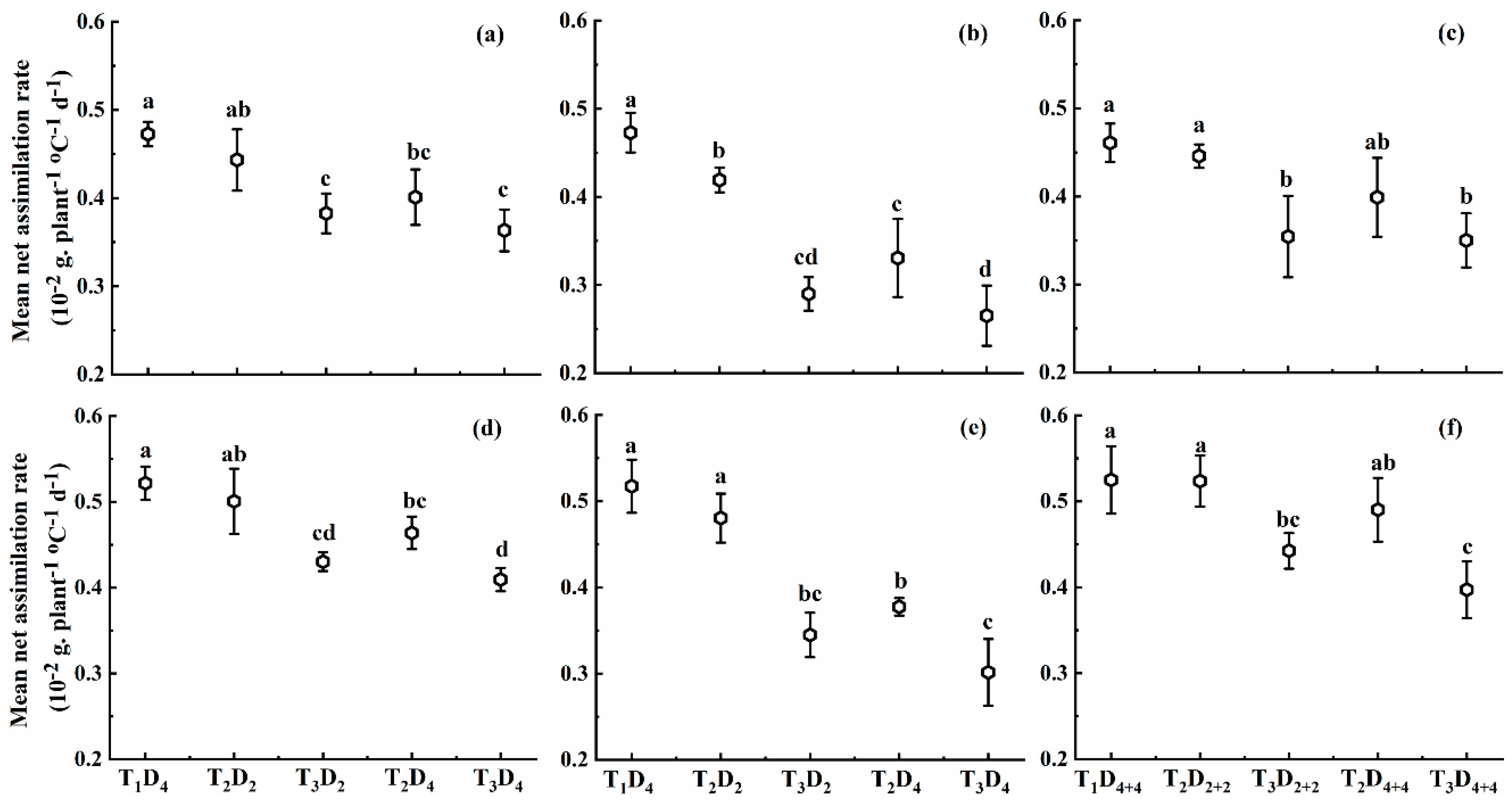
2.1.2. Effect of High Temperature on Mean Net Assimilation Rate
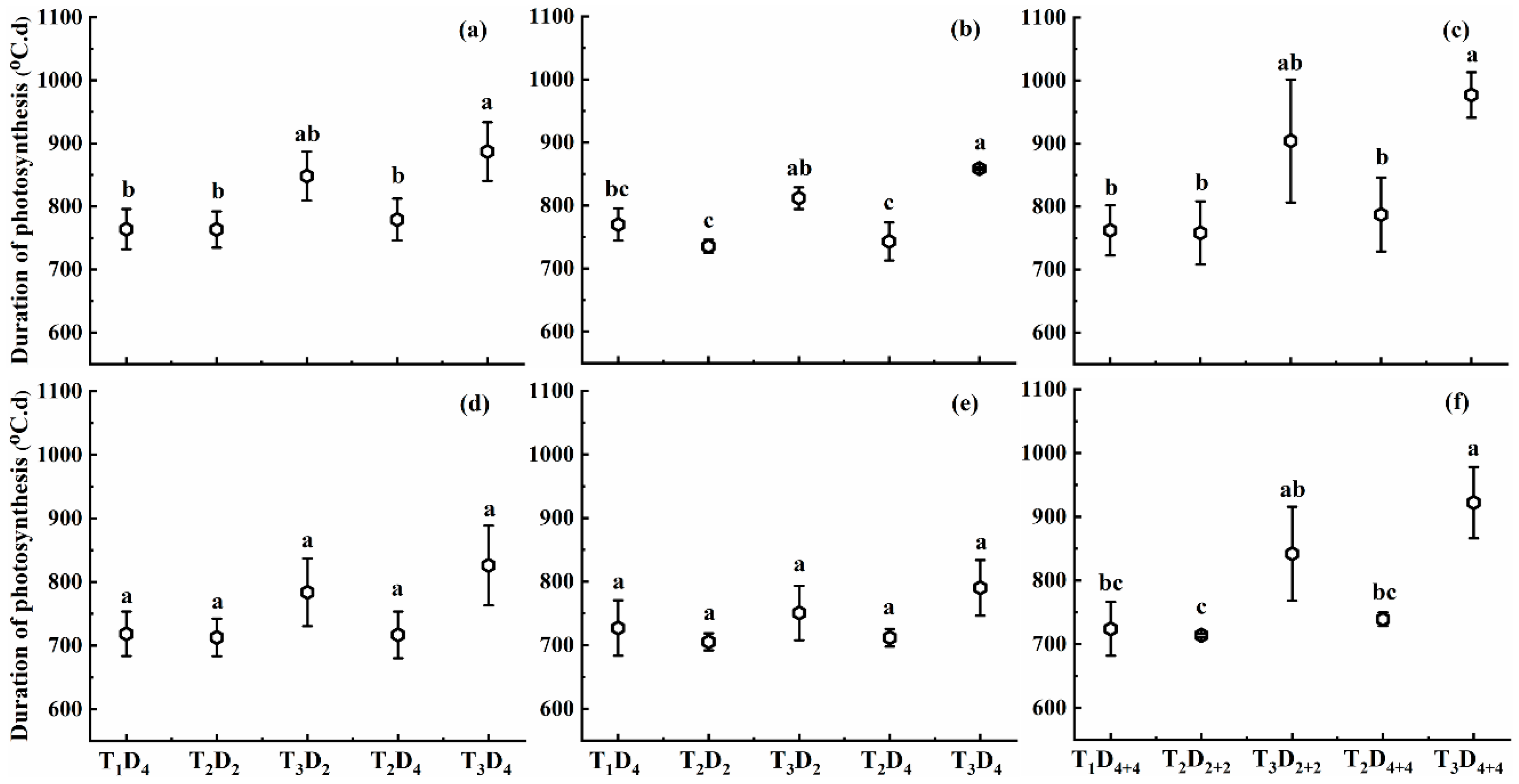
2.1.3. Effect of High Temperature on Duration of Photosynthesis
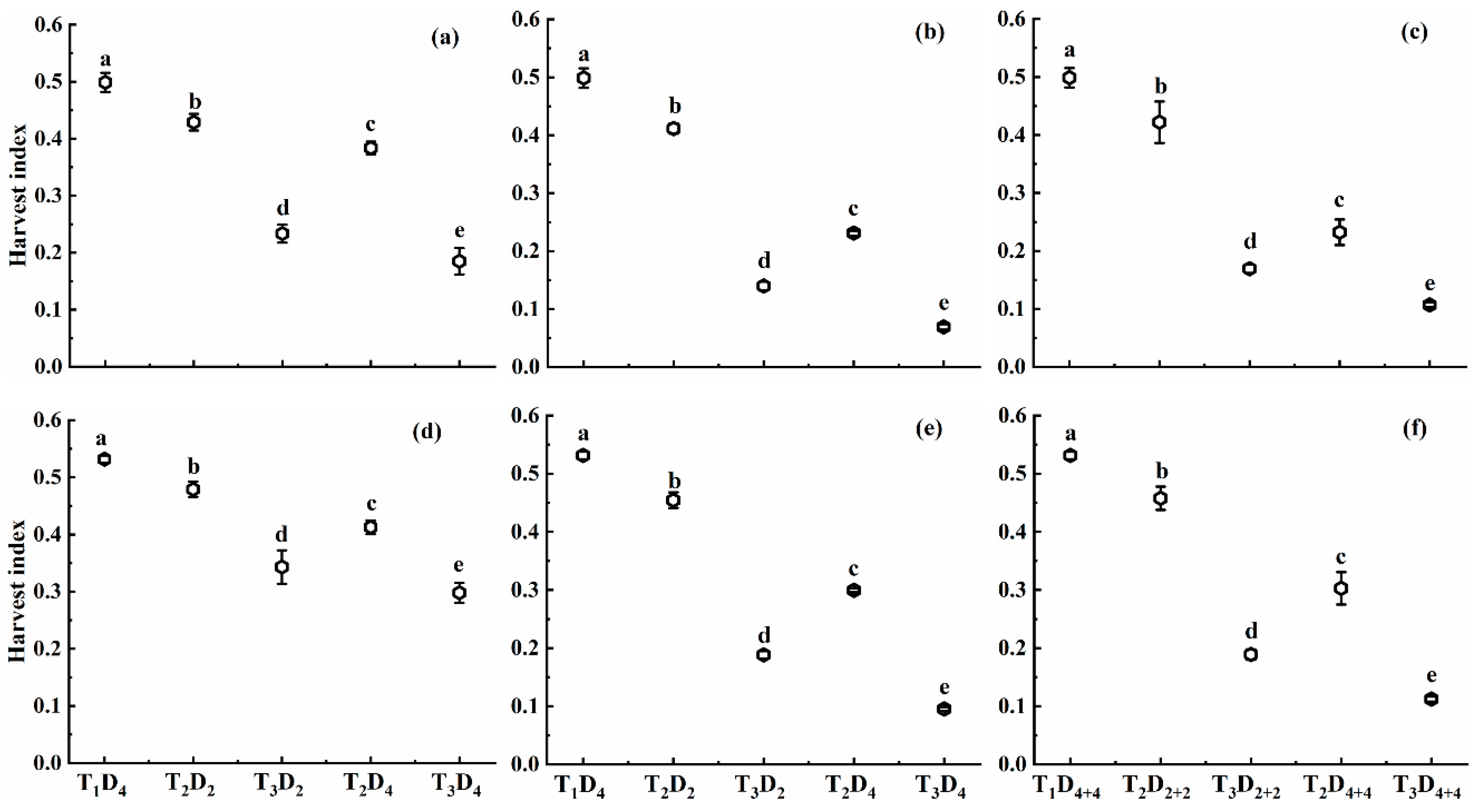
2.1.4. Effect of High Temperature on Harvest Index
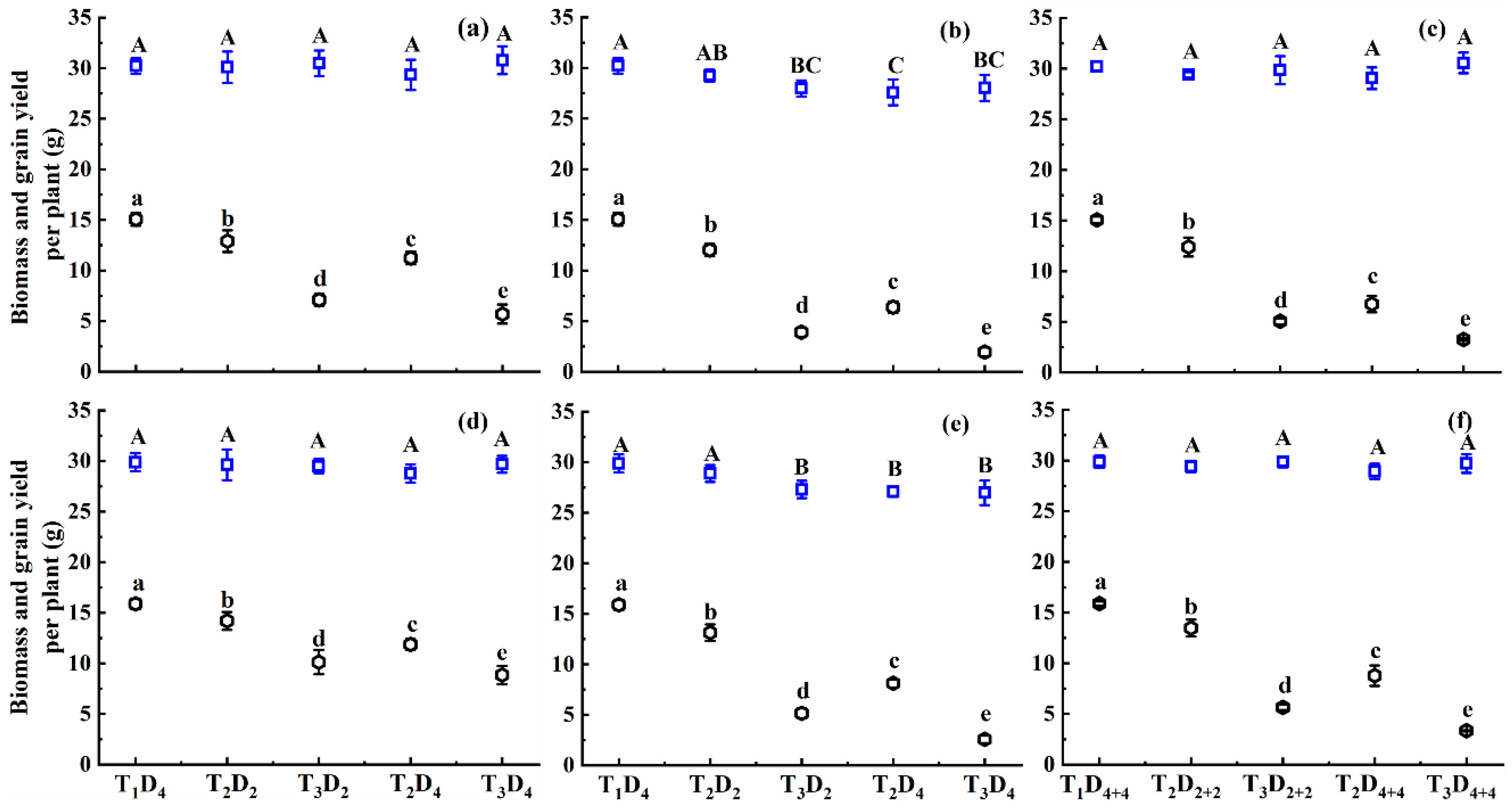
2.1.5. Effect of High Temperature on Biomass Accumulation and Grain Yield
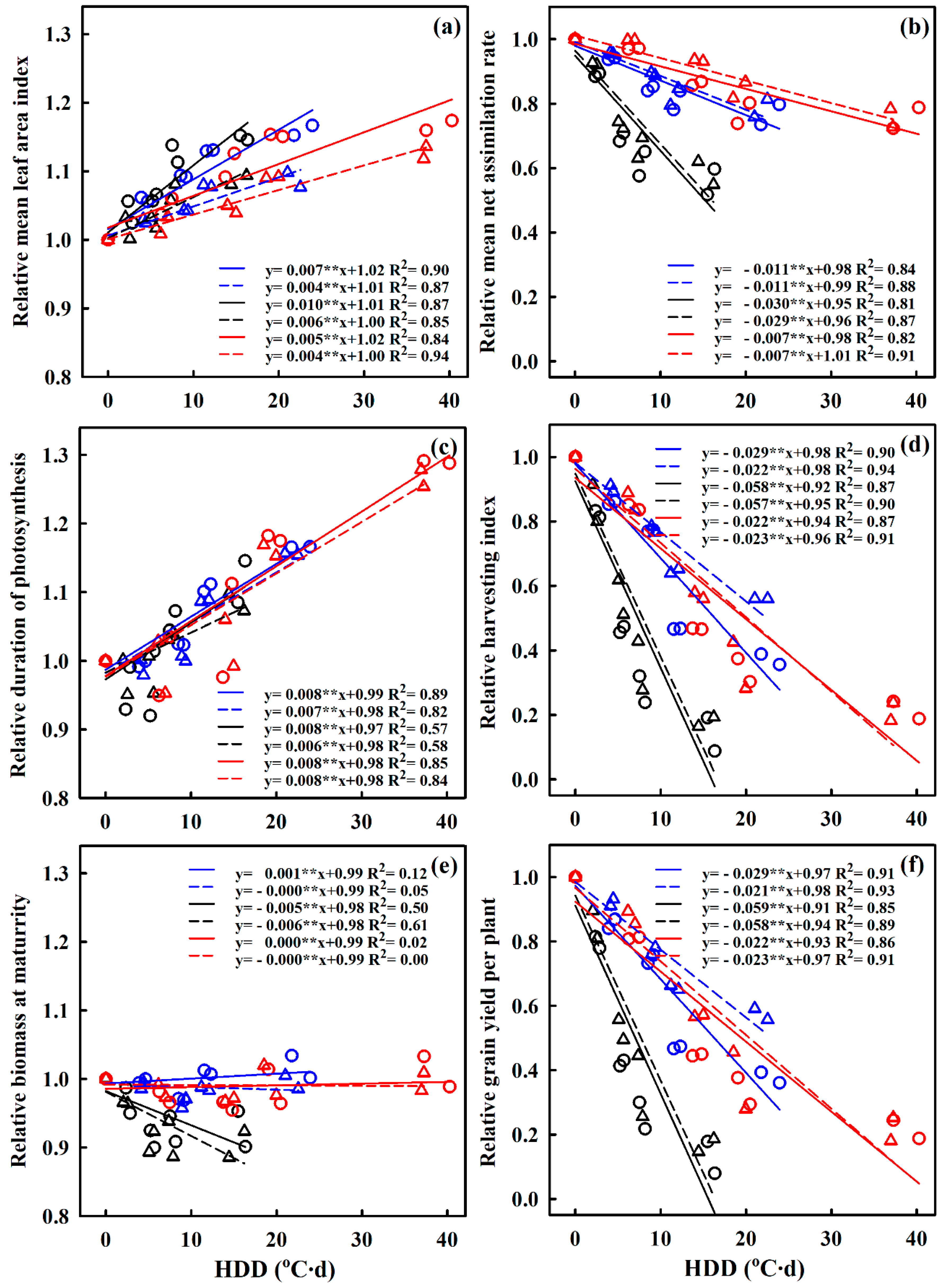
2.2. Quantifying the Effects of Extreme High-Temperature Stress on Photosynthetic Property Parameters, Biomass Accumulation, and Grain Yield in Response to HDD
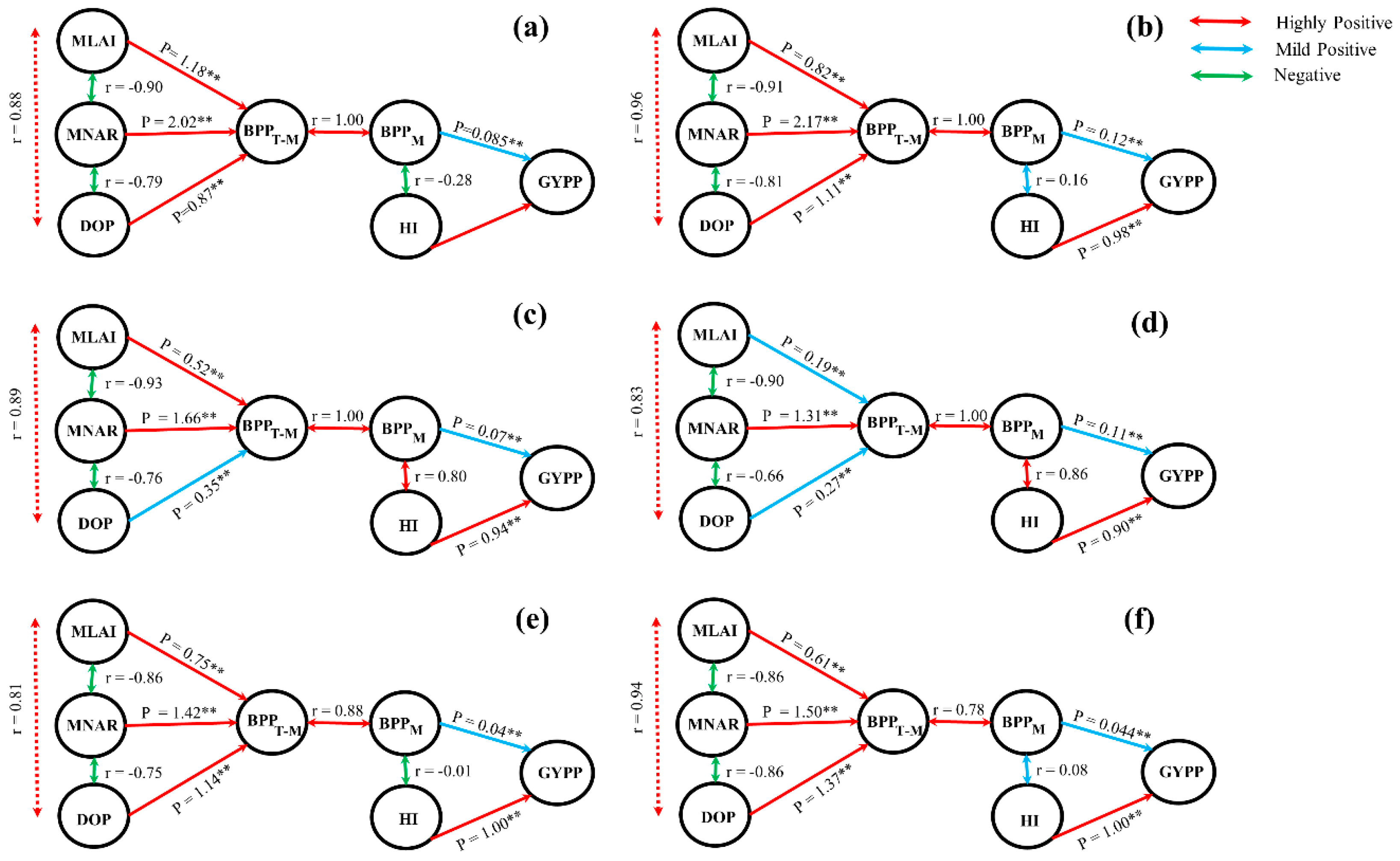
2.3. Relationships Between Grain Yield, Biomass Accumulation and Photosynthetic Property Parameters under High-Temperature Condition
3. Discussion
3.1. Biomass Production and Grain Yield Responses to High Temperatures
3.2. Relationships of HDD with Relative Biomass Production and Grain Yield
3.3. Limitation of this Study
4. Materials and Methods
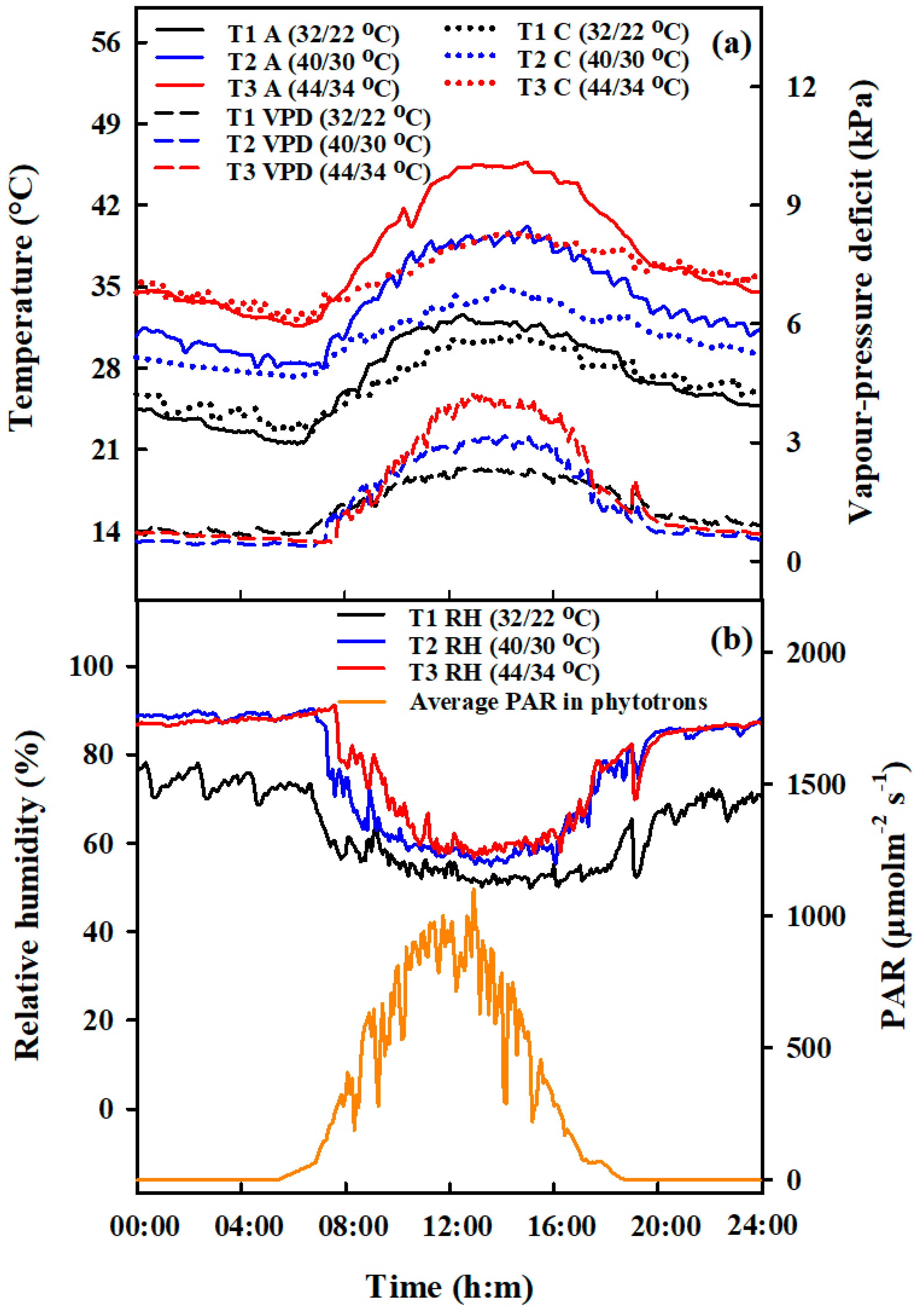
4.1. Experimental Design
4.2. Leaf Area and Biomass Measurements
4.3. Measurement of Grain Yield
4.4. Calculation of Grain Yield Formation
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V. High-Temperature Effects on Rice Growth, Yield, and Grain Quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar]
- Fischer, E.M.; Schär, C.; Ban, N.; Bresch, D.N.; Casanueva, A.; Feigenwinter, I.; Graf, M.A.; Hirschi, M.; Kotlarski, S.; Liniger, M. Climate Extremes and Climate Indices; National Centre for Climate Services NCCS: Zürich, Switzerland, 2018; pp. 103–140. [Google Scholar]
- Lobell, D.B.; Sibley, A.; Ortiz-Monasterio, J.I. Extreme heat effects on wheat senescence in India. Nat. Clim. Chang. 2012, 2, 186. [Google Scholar] [CrossRef]
- Prasad, P.; Boote, K.; Allen, L., Jr.; Sheehy, J.; Thomas, J. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z.; Zhang, T.; Fahad, S.; Cui, K.; Nie, L.; Peng, S.; Huang, J. The effect of season-long temperature increases on rice cultivars grown in the central and southern regions of China. Front. Plant Sci. 2017, 8, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buu, B.C.; Chan, C.Y.; Lang, N.T. Molecular Breeding for Improving Heat Stress Tolerance in Rice: Recent Progress and Future Perspectives. In Molecular Breeding for Rice Abiotic Stress Tolerance and Nutritional Quality; Hossain, M.A., Hassan, L., Ifterkharuddaula, K.M., Kumar, A., Henry, R., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; pp. 92–119. [Google Scholar] [CrossRef]
- Jagadish, S.K. Heat stress during flowering in cereals–effects and adaptation strategies. New Phytol. 2020, 226, 1567–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahad, S.; Adnan, M.; Hassan, S.; Saud, S.; Hussain, S.; Wu, C.; Wang, D.; Hakeem, K.R.; Alharby, H.F.; Turan, V. Rice responses and tolerance to high temperature. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publ. Ltd.: Cambridge, UK, 2019; pp. 201–224. [Google Scholar] [CrossRef]
- Wahid, A. Physiological implications of metabolite biosynthesis for net assimilation and heat-stress tolerance of sugarcane (Saccharum officinarum) sprouts. J. Plant Res. 2007, 120, 219–228. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Change Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef]
- Shi, P.; Zhu, Y.; Tang, L.; Chen, J.; Sun, T.; Cao, W.; Tian, Y. Differential effects of temperature and duration of heat stress during anthesis and grain filling stages in rice. Environ. Exp. Bot. 2016, 132, 28–41. [Google Scholar] [CrossRef]
- Zhen, F.; Zhou, J.; Mahmood, A.; Wang, W.; Chang, X.; Liu, B.; Liu, L.; Cao, W.; Zhu, Y.; Tang, L. Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice. Crop J. 2020, 8, 194–212. [Google Scholar] [CrossRef]
- Impa, S.M.; Raju, B.; Hein, N.T.; Sandhu, J.; Prasad, P.V.; Walia, H.; Jagadish, S.K. High night temperature effects on wheat and rice: Current status and way forward. Plant Cell Environ. 2021. [Google Scholar] [CrossRef]
- Espe, M.B.; Hill, J.E.; Hijmans, R.J.; McKenzie, K.; Mutters, R.; Espino, L.A.; Leinfelder-Miles, M.; van Kessel, C.; Linquist, B.A. Point stresses during reproductive stage rather than warming seasonal temperature determine yield in temperate rice. Glob. Chang. Biol. 2017, 23, 4386–4395. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Hasegawa, T.; Tang, L.; Wang, W.; Zhou, J.; Liu, L.; Liu, B.; Cao, W.; Zhu, Y. Stage-dependent temperature sensitivity function predicts seed-setting rates under short-term extreme heat stress in rice. Agric. For. Meteorol. 2018, 256, 196–206. [Google Scholar] [CrossRef]
- Shi, P.; Tang, L.; Lin, C.; Liu, L.; Wang, H.; Cao, W.; Zhu, Y. Modeling the effects of post-anthesis heat stress on rice phenology. Field Crops Res. 2015, 177, 26–36. [Google Scholar] [CrossRef]
- Kato, M.; Kobayashi, K.; Ogiso, E.; Yokoo, M. Photosynthesis and dry-matter production during ripening stage in a female-sterile line of rice. Plant Prod. Sci. 2004, 7, 184–188. [Google Scholar] [CrossRef]
- Zhang, H.; Jing, W.; Zhao, B.; Wang, W.; Xu, Y.; Zhang, W.; Gu, J.; Liu, L.; Wang, Z.; Yang, J. Alternative fertilizer and irrigation practices improve rice yield and resource use efficiency by regulating source-sink relationships. Field Crops Res. 2021, 265, 108124. [Google Scholar] [CrossRef]
- Kim, J.; Shon, J.; Lee, C.-K.; Yang, W.; Yoon, Y.; Yang, W.-H.; Kim, Y.-G.; Lee, B.-W. Relationship between grain filling duration and leaf senescence of temperate rice under high temperature. Field Crops Res. 2011, 122, 207–213. [Google Scholar] [CrossRef]
- Oh-e, I.; Saitoh, K.; Kuroda, T. Effects of high temperature on growth, yield and dry-matter production of rice grown in the paddy field. Plant Prod. Sci. 2007, 10, 412–422. [Google Scholar] [CrossRef]
- Morales, F.; Ancín, M.; Fakhet, D.; González-Torralba, J.; Gámez, A.L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic metabolism under stressful growth conditions as a bases for crop breeding and yield improvement. Plants 2020, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Cai, C.; He, J.; Gu, J.; Zhu, G.; Zhang, W.; Zhu, J.; Liu, G. Yield, dry matter distribution and photosynthetic characteristics of rice under elevated CO2 and increased temperature conditions. Field Crops Res. 2020, 248, 107605. [Google Scholar] [CrossRef]
- Heggenstaller, A.H.; Liebman, M.; Anex, R.P. Growth analysis of biomass production in sole-crop and double-crop corn systems. Crop Sci. 2009, 49, 2215–2224. [Google Scholar] [CrossRef]
- Hunt, R. Plant Growth Curves. The Functional Approach to Plant Growth Analysis; Edward Arnold Ltd.: London, UK, 1982. [Google Scholar]
- Liu, L.; Ji, H.; An, J.; Shi, K.; Ma, J.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y. Response of biomass accumulation in wheat to low-temperature stress at jointing and booting stages. Environ. Exp. Bot. 2019, 157, 46–57. [Google Scholar] [CrossRef]
- Nagai, T.; Makino, A. Differences between rice and wheat in temperature responses of photosynthesis and plant growth. Plant Cell Physiol. 2009, 50, 744–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, K.; Bhattacharyya, P.; Neogi, S.; Rao, K.; Adhya, T. Combined effect of elevated CO2 and temperature on dry matter production, net assimilation rate, C and N allocations in tropical rice (Oryza sativa L.). Field Crops Res. 2012, 139, 71–79. [Google Scholar] [CrossRef]
- Richards, R. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Li, G.; Yang, H.; Yang, J.; Liu, H.; Struik, P.C.; Luo, W.; Yin, X.; Di, L.; Guo, X. Do all leaf photosynthesis parameters of rice acclimate to elevated CO2, elevated temperature, and their combination, in FACE environments? Glob. Chang. Biol. 2018, 24, 1685–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, A.; Tarpley, L. High nighttime temperatures affect rice productivity through altered pollen germination and spikelet fertility. Agric. For. Meteorol. 2009, 149, 999–1008. [Google Scholar] [CrossRef]
- Li, Z.-G.; Min, X.; Zhou, Z.-H. Hydrogen sulfide: A signal molecule in plant cross-adaptation. Front. Plant Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [Green Version]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat priming during early reproductive stages enhances thermo-tolerance to post-anthesis heat stress via improving photosynthesis and plant productivity in winter wheat (Triticum aestivum L.). Front. Plant. Sci. 2018, 9. [Google Scholar] [CrossRef]
- Shi, W.; Lawas, L.; Raju, B.; Jagadish, S. Acquired thermo-tolerance and trans-generational heat stress response at flowering in rice. J. Agron. Crop Sci. 2016, 202, 309–319. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Jin, M.; Yu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J. Cereal Sci. 2012, 55, 331–336. [Google Scholar] [CrossRef]
- Jagadish, S.; Craufurd, P.; Wheeler, T. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, L.; Zhou, J.; Hu, S.; Chen, H.; Xiang, J.; Zhang, Y.; Zeng, Y.; Shi, Q.; Zhu, D. Research progress on heat stress of rice at flowering stage. Rice Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Peng, S. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice. Sci. Rep. 2016, 6, 34978. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schmid, B.; Wang, F.; Paine, C.T. Net assimilation rate determines the growth rates of 14 species of subtropical forest trees. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Shipley, B. Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis. Funct. Ecol. 2006, 20, 565–574. [Google Scholar] [CrossRef]
- Gordon, A.; Hesketh, J.; Peters, D. Soybean leaf photosynthesis in relation to maturity classification and stage of growth. Photosynth. Res. 1982, 3, 81–93. [Google Scholar] [CrossRef]
- Dias, A.; Lidon, F. Evaluation of grain filling rate and duration in bread and durum wheat, under heat stress after anthesis. J. Agron. Crop Sci. 2009, 195, 137–147. [Google Scholar] [CrossRef]
- Wang, X.; Xin, C.; Cai, J.; Zhou, Q.; Dai, T.; Cao, W.; Jiang, D. Heat priming induces trans-generational tolerance to high temperature stress in wheat. Front. Plant Sci. 2016, 7, 501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Z.; Li, L.; Zhou, Q.; Xiao, Y.; Wei, X.; Zhou, M. Short-term complete submergence of rice at the tillering stage increases yield. PLoS ONE 2015, 10, e0127982. [Google Scholar] [CrossRef]
- Liu, B.; Asseng, S.; Wang, A.; Wang, S.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Modelling the effects of post-heading heat stress on biomass growth of winter wheat. Agric. For. Meteorol. 2017, 247, 476–490. [Google Scholar] [CrossRef]
- Hossain, M.A.; Li, Z.-G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and possible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Bhagsari, A.S.; Brown, R. Leaf Photosynthesis and its Correlation with Leaf Area 1. Crop Sci. 1986, 26, 127–132. [Google Scholar] [CrossRef]
- Lin, M.-Y.; Chai, K.-H.; Ko, S.-S.; Kuang, L.-Y.; Lur, H.-S.; Charng, Y.-Y. A positive feedback loop between HEAT SHOCK PROTEIN101 and HEAT STRESS-ASSOCIATED 32-KD PROTEIN modulates long-term acquired thermotolerance illustrating diverse heat stress responses in rice varieties. Plant Physiol. 2014, 164, 2045–2053. [Google Scholar] [CrossRef]
- Van Herwaarden, A.; Richards, R.; Farquhar, G.; Angus, J. ‘Haying-off’, the negative grain yield response of dryland wheat to nitrogen fertiliser III. The influence of water deficit and heat shock. Aust. J. Agric. Res. 1998, 49, 1095–1110. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. FAO Rome 1998, 300, D05109. [Google Scholar]
- Tang, L.; Zhu, Y.; Hannaway, D.; Meng, Y.; Liu, L.; Chen, L.; Cao, W. RiceGrow: A rice growth and productivity model. NJAS Wagening. J. Life Sci. 2009, 57, 83–92. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Cultivar | Photosynthetic Properties | T | D | T × D |
|---|---|---|---|---|---|
| Booting | Huaidao-5 | MLAI | <0.0001 | ns | ns |
| MNAR | <0.0001 | ns | ns | ||
| DOP | 0.011 | ns | ns | ||
| HI | <0.0001 | 0.002 | ns | ||
| BBPM | ns | ns | ns | ||
| GYPP | <0.0001 | 0.001 | 0.034 | ||
| Wuyunjing-24 | MLAI | 0.002 | ns | ns | |
| MNAR | <0.0001 | ns | ns | ||
| DOP | 0.045 | ns | ns | ||
| HI | <0.0001 | <0.0001 | 0.012 | ||
| BBPM | ns | ns | ns | ||
| GYPP | <0.0001 | <0.0001 | 0.009 | ||
| Flowering | Huaidao-5 | MLAI | <0.0001 | ns | ns |
| MNAR | <0.0001 | 0.019 | 0.046 | ||
| DOP | 0.002 | ns | ns | ||
| HI | <0.0001 | <0.0001 | <0.0001 | ||
| BBPM | <0.0001 | ns | ns | ||
| GYPP | <0.0001 | <0.0001 | <0.0001 | ||
| Wuyunjing-24 | MLAI | 0.003 | ns | ns | |
| MNAR | <0.0001 | 0.004 | 0.048 | ||
| DOP | ns | ns | ns | ||
| HI | <0.0001 | <0.0001 | <0.0001 | ||
| BBPM | <0.0001 | 0.029 | ns | ||
| GYPP | <0.0001 | <0.0001 | <0.0001 | ||
| Combined | Huaidao-5 | MLAI | <0.0001 | ns | ns |
| MNAR | <0.0001 | ns | ns | ||
| DOP | 0.010 | ns | ns | ||
| HI | <0.0001 | <0.0001 | <0.0001 | ||
| BBPM | ns | ns | ns | ||
| GYPP | <0.0001 | <0.0001 | <0.0001 | ||
| Wuyunjing-24 | MLAI | 0.001 | ns | ns | |
| MNAR | <0.0001 | ns | ns | ||
| DOP | 0.004 | ns | ns | ||
| HI | <0.0001 | <0.0001 | <0.0001 | ||
| BBPM | ns | ns | ns | ||
| GYPP | <0.0001 | <0.0001 | <0.0001 |
| Cultivar | Temperature Levels | Stage | Durations |
|---|---|---|---|
| Huaidao-5 and Wuyunjing-24 | T1(32/22 °C) T2(40/30 °C) T3(44/34 °C) | Booting | D2 (two days booting) and D4 (four days booting) |
| Flowering | D2 (two days flowering) and D4 (four days flowering) | ||
| Combined stages | D2+2 (four days = 2 days booting + 2 days flowering) and D4+4 (eight days = 4 days booting + 4 days flowering) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, A.; Wang, W.; Ali, I.; Zhen, F.; Osman, R.; Liu, B.; Liu, L.; Zhu, Y.; Cao, W.; Tang, L. Individual and Combined Effects of Booting and Flowering High-Temperature Stress on Rice Biomass Accumulation. Plants 2021, 10, 1021. https://doi.org/10.3390/plants10051021
Mahmood A, Wang W, Ali I, Zhen F, Osman R, Liu B, Liu L, Zhu Y, Cao W, Tang L. Individual and Combined Effects of Booting and Flowering High-Temperature Stress on Rice Biomass Accumulation. Plants. 2021; 10(5):1021. https://doi.org/10.3390/plants10051021
Chicago/Turabian StyleMahmood, Aqib, Wei Wang, Iftikhar Ali, Fengxian Zhen, Raheel Osman, Bing Liu, Leilei Liu, Yan Zhu, Weixing Cao, and Liang Tang. 2021. "Individual and Combined Effects of Booting and Flowering High-Temperature Stress on Rice Biomass Accumulation" Plants 10, no. 5: 1021. https://doi.org/10.3390/plants10051021