
Topical Application of Double-Stranded RNA Targeting 2b and CP Genes of Cucumber mosaic virus Protects Plants against Local and Systemic Viral Infection


Abstract
:1. Introduction
2. Results
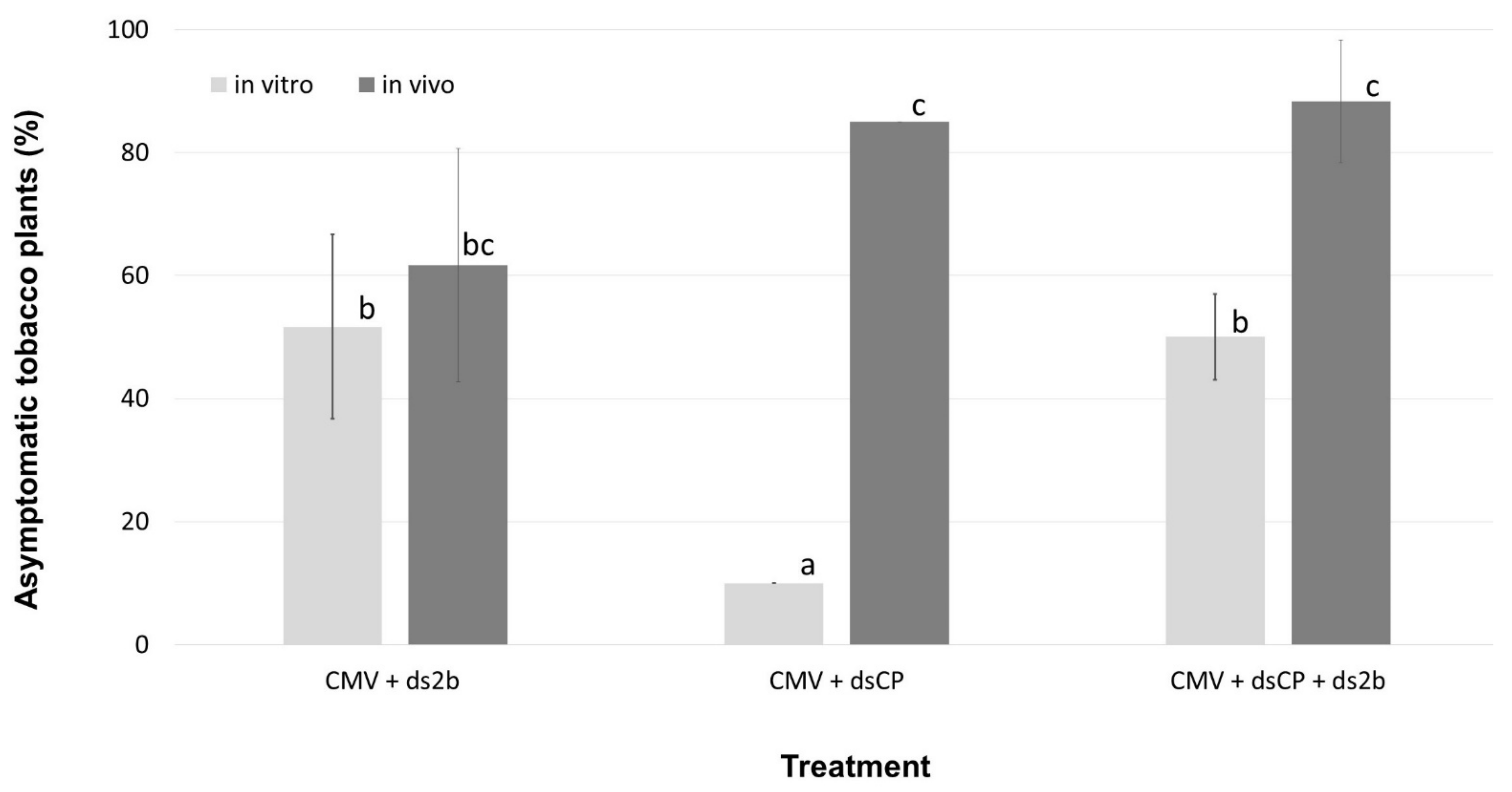
2.1. dsRNA Treatments of Tobacco Plants Resulted in Variable Levels of Protection against CMV Infection, Which Depended on the dsRNA Construct Used and the Timing of Virus Inoculation
2.2. dsRNA Treatment Protects from Local Infection in Chenopodium Quinoa Plants
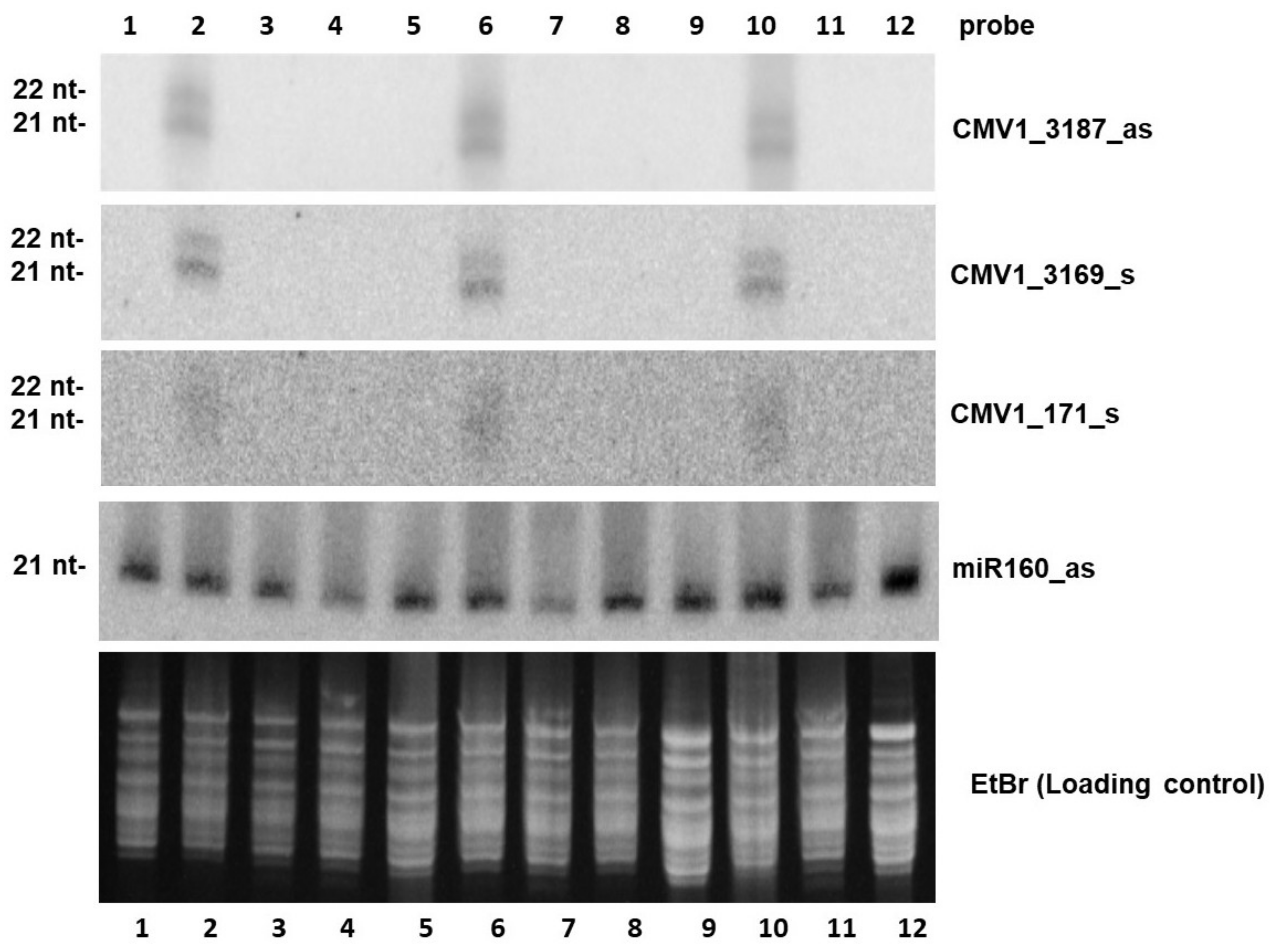
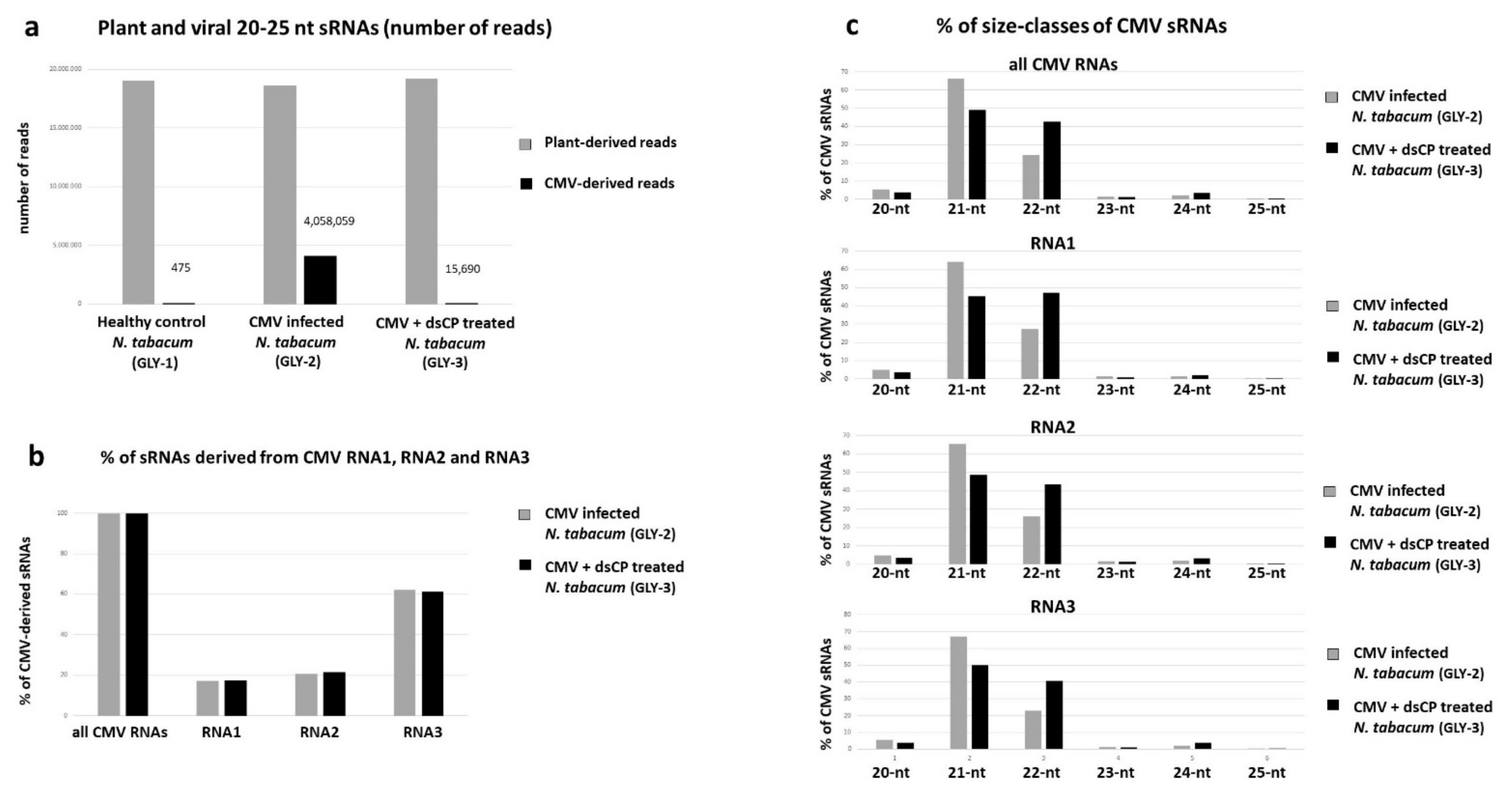
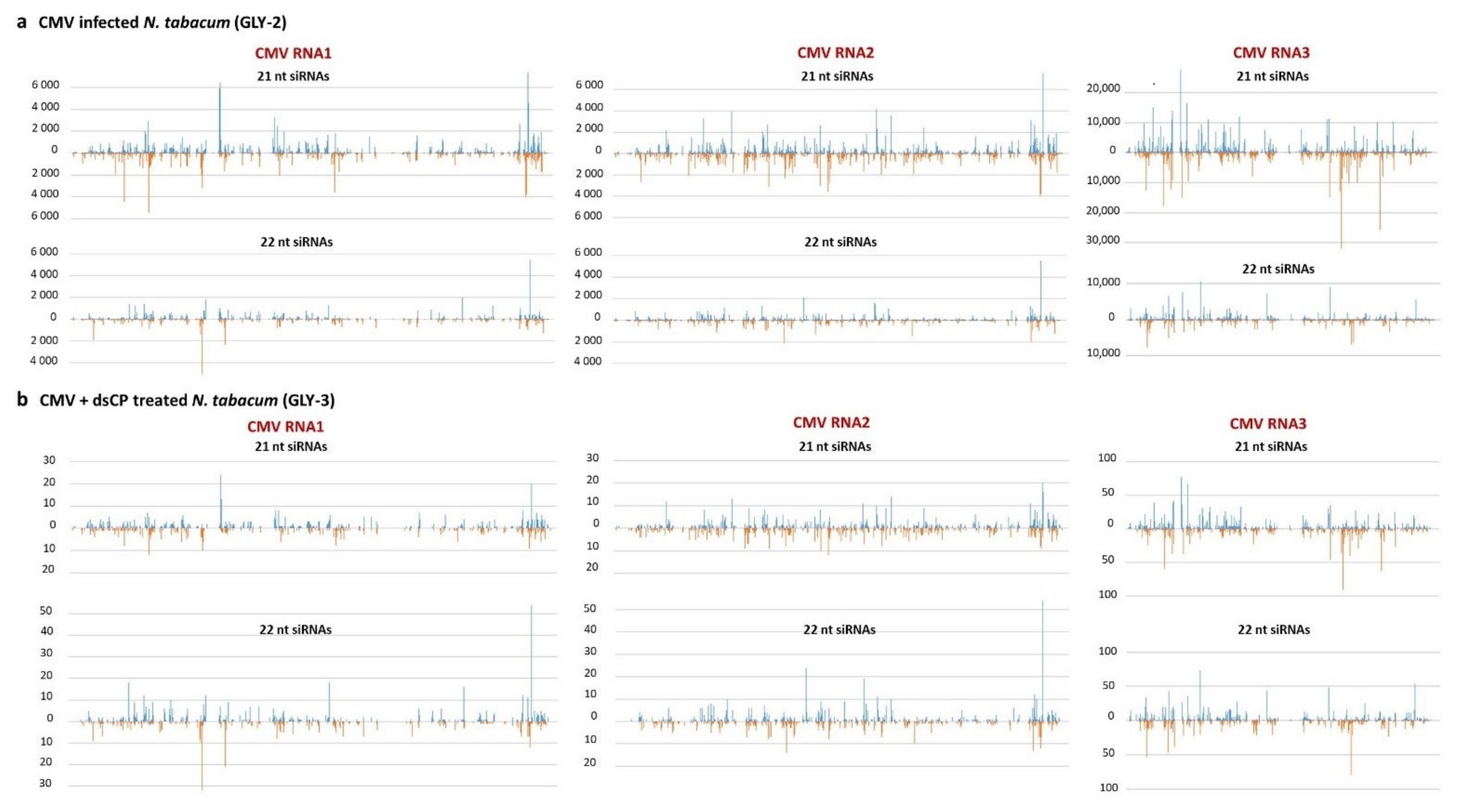
2.3. dsRNA Treatment Leads to Reduced Accumulation of Viral siRNAs and Alteration in Their Size Profile in Systemic Leaves
3. Discussion
4. Materials and Methods
4.1. Virus Isolate, Bacterial Strains, and Plant Material
4.2. Construction of the Target DNA Templates for Transcription
4.3. Production of dsRNA Molecules
4.3.1. In Vivo dsRNA Production
4.3.2. In Vitro dsRNA Production
4.4. Plant Bioassays
4.4.1. Bioassay on Tobacco
4.4.2. Bioassay on Chenopodium Quinoa Plants
4.4.3. Small RNA Blot Hybridization Analysis
4.4.4. Illumina Sequencing and Bioinformatics Analysis of Viral siRNAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zotti, M.; Dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA interference technology in crop protection against arthropod pests, pathogens and nematodes. Pest Manag. Sci. 2018, 74, 1239–1250. [Google Scholar] [CrossRef]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Bronkhorst, A.W.; van Rij, R.P. The long and short of antiviral defense: Small RNA-based immunity in insects. Curr. Opin. Virol. 2014, 7, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Pantaleo, V.; Burgyán, J. RNA silencing: An antiviral mechanism. Adv. Virus Res. 2009, 75, 35–71. [Google Scholar] [PubMed]
- Li, Y.; Lu, J.; Han, Y.; Fan, X.; Ding, S.W. RNA interference functions as an antiviral immunity mechanism in mammals. Science 2013, 342, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.B.; Metzlaff, M. RNA silencing and antiviral defense in plants. Curr. Opin. Plant Biol. 2005, 8, 216–222. [Google Scholar] [CrossRef]
- Wang, M.B.; Masuta, C.; Smith, N.A.; Shimura, H. RNA silencing and plant viral diseases. Mol. Plant Microbe Interact. 2012, 25, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, A. RNA-targeted antiviral immunity: More than just RNA silencing. Trends Microbiol. 2019, 27, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Valkonen, J.P. Utilization of engineered resistance to viruses in crops of the developing world, with emphasis on sub-Saharan Africa. Curr. Opin. Virol. 2017, 26, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Pooggin, M.M. RNAi-mediated resistance to viruses: A critical assessment of methodologies. Curr. Opin. Virol. 2017, 26, 28–35. [Google Scholar] [CrossRef]
- Dong, O.X.; Ronald, P.C. Genetic engineering for disease resistance in plants: Recent progress and future perspectives. Plant Physiol. 2019, 180, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Tenllado, F.; Diaz-Ruiz, J.R. Double-stranded RNA-mediated interference with plant virus infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [Green Version]
- Tenllado, F.; Martínez-García, B.; Vargas, M. Díaz-Ruíz, J.R. Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC Biotechnol. 2003, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Xu, Z.P.; Carroll, B.J. Induction of virus resistance by exogenous application of double-stranded RNA. Curr. Opin. Virol. 2017, 26, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Voloudakis, A.E.; Holeva, M.C.; Kaldis, A.; Kim, D. Tools and techniques for production of double stranded RNA and its application for management of plant viral diseases. In Genes, Genetics and Transgenics for Virus Resistance in Plants; Basavaprabhu, P.L., Ed.; Caister Academic Press: Poole, UK, 2018; pp. 119–140. [Google Scholar]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.; Papadopoulou, K.K. Genetically Modified Organism-Free RNA interference: Exogenous application of RNA molecules in plants. Plant Physiol. 2020, 182, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pooggin, M.M. Small RNA-omics for plant virus identification, virome reconstruction, and antiviral defense characterization. Front Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Roossinck, M.J. Evolutionary history of Cucumber mosaic virus deduced by phylogenetic analyses. J. Virol. 2002, 76, 3382–3387. [Google Scholar] [CrossRef] [Green Version]
- Jacquemond, M. Cucumber Mosaic Virus. Adv. Virus Res. 2012, 84, 439–504. [Google Scholar]
- Sclavounos, A.P.; Voloudakis, A.E.; Arabatzis, C.; Kyriakopoulou, P.E. A severe Hellenic CMV tomato isolate: Symptom variability in tobacco, characterization and discrimination of variants. Eur. J. Plant Pathol. 2006, 115, 163–172. [Google Scholar] [CrossRef]
- Ye, J.; Qu, J.; Zhang, J.F.; Geng, Y.F.; Fang, R.X. A critical domain of the Cucumber mosaic virus 2b protein for RNA silencing suppressor activity. FEBS Lett. 2009, 583, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Masuta, C.; Inaba, J.; Shimura, H. The 2b proteins of Cucumber mosaic virus generally have the potential to differentially induce necrosis on Arabidopsis. Plant Signal Behav. 2012, 7, 43–45. [Google Scholar] [CrossRef] [Green Version]
- Gallitelli, D. The ecology of Cucumber mosaic virus and sustainable agriculture. Virus Res. 2000, 71, 9–21. [Google Scholar] [CrossRef]
- Kyriakopoulou, P.E.; Bem, F.; Varveri, C. Tomato shrinkage and tomato fruit toughness, two new diseases in Greece, probably related to cucumber mosaic virus. Ann. Inst. Phytopathol. Benaki 1991, 16, 151–156. [Google Scholar]
- Kyriakopoulou, P.E.; Perdikis, D.C.; Sclavounos, A.P.; Girgis, S.M.; Lycouressis, D.P.; Tsitsipis, J.A.; Christakis, P.A. Cucumber mosaic cucumovirus incidence in open-field tomato in the Olympia area and trap captures of alate aphids. Bull. OEPP 2000, 30, 305–315. [Google Scholar] [CrossRef]
- Kyriakopoulou, P.E.; Vassilakos, N.; Bem, F.P.; Varveri, C.; Girkis, M.S.; Skopelitis, N.G. Cucumber mosaic virus in industrial tomato of Eleia county and the new threat from tomato necrosis. Phytopathol. Mediterr. 2000, 39, 330. [Google Scholar]
- Jorda, C.; Alfaro, A.; Arranda, M.A.; Moriones, E.; Garcia-Arenal, F. An epidemic of Cucumber mosaic virus plus satellite RNA in tomatoes in eastern Spain. Plant Dis. 1992, 76, 363–366. [Google Scholar] [CrossRef]
- Shintaku, M. Coat protein gene sequences of two Cucumber mosaic virus strains reveal a single amino acid change correlating with chlorosis induction. J. Gen. Virol. 1991, 72, 2587–2589. [Google Scholar] [CrossRef] [PubMed]
- Shintaku, M.; Zhang, L.; Palukaitis, P. A single amino acid substitution in the coat protein of Cucumber mosaic virus induces chlorosis in tobacco. Plant Cell 1992, 4, 751–757. [Google Scholar] [PubMed]
- Palukaitis, P.; Garcia-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar]
- Morroni, M.; Thompson, J.R.; Tepfer, M. Twenty Years of transgenic plants resistant to Cucumber mosaic virus. Mol. Plant Microbe Interact. 2008, 21, 675–684. [Google Scholar] [CrossRef]
- Beachy, R.N.; Loesch-Fries, S.; Tumer, N.E. Coat protein-mediated resistance against virus infection. Ann. Rev. Phytopath. 1990, 28, 451–472. [Google Scholar] [CrossRef]
- Fuchs, M.; Provvidenti, R.; Slightom, J.L.; Gonsalves, D. Evaluation of transgenic tomato plants expressing the coat protein gene of Cucumber mosaic virus strain WL under field conditions. Plant Dis. 1996, 80, 270–275. [Google Scholar] [CrossRef]
- Fuchs, M.; Chirco, E.M.; Gonsalves, D. Movement of coat protein genes from a commercial virus-resistant transgenic squash into a wild relative. Environ. Biosafety Res. 2004, 3, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gielen, J.; Ultzen, T.; Bontems, S.; Loots, W.; van Schepen, A. Coat protein-mediated protection to Cucumber mosaic virus infections in cultivated tomato. Euphytica 1996, 88, 139–149. [Google Scholar] [CrossRef]
- Jacquemond, M.; Teycheney, P.-Y.; Carrère, I.; Navas-Castillo, J.; Tepfer, M. Resistance phenotypes of transgenic tobacco plants expressing different Cucumber mosaic virus (CMV) coat protein genes. Mol. Breed. 2001, 8, 85–94. [Google Scholar] [CrossRef]
- Kalantidis, K.; Psaradakis, S.; Tabler, M.; Tsagris, M. The occurrence of CMV-specific short RNAs in transgenic tobacco expressing virus-derived double-stranded RNA is indicative of resistance to the virus. Mol. Plant Microbe Interact. 2002, 15, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-K.; Lohuis, D.; Goldbach, R.; Prins, M. High frequency induction of RNA-mediated resistance against Cucumber mosaic virus using inverted repeat constructs. Mol. Breed. 2004, 14, 215–226. [Google Scholar] [CrossRef]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA interference: A natural immune system of plants to counteract biotic stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Ye, J.; Fang, R. Artificial microRNA-mediated virus resistance in plants. J. Virol. 2007, 81, 6690–6699. [Google Scholar] [CrossRef] [Green Version]
- Voloudakis, A.E.; Holeva, M.C.; Sarin, L.P.; Bamford, D.H.; Vargas, M.; Poranen, M.M.; Tenllado, F. Efficient double-stranded RNA production methods for utilization in plant virus control. Methods Mol. Biol. 2015, 1236, 255–274. [Google Scholar]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.B.; Jovel, J.; Udomporn, P.; Wang, Y.; Wu, Q.; Li, W.X.; Gasciolli, V.; Vaucheret, H.; Ding, S.W. The 21-nucleotide, but not 22-nucleotide, viral secondary small interfering RNAs direct potent antiviral defense by two cooperative argonautes in Arabidopsis thaliana. Plant Cell 2011, 23, 1625–1638. [Google Scholar] [CrossRef] [Green Version]
- Kaldis, A.; Berbati, M.; Melita, O.; Reppa, C.; Holeva, M.; Otten, P.; Voloudakis, A. Exogenously applied dsRNA molecules deriving from the Zucchini yellow mosaic virus (ZYMV) genome move systemically and protect cucurbits against ZYMV. Mol. Plant Pathol. 2018, 19, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Namgial, T.; Kaldis, A.; Chakraborty, S.; Voloudakis, A. Topical application of double-stranded RNA molecules containing sequences of Tomato leaf curl virus and Cucumber mosaic virus confers protection against the cognate viruses. Physiol. Mol. Plant Pathol. 2019, 101432. [Google Scholar] [CrossRef]
- Tabein, S.; Jansen, M.; Noris, E.; Vaira, A.M.; Marian, D.; Behjatnia, S.A.A.; Accotto, G.P.; Miozzi, L. The induction of an effective dsRNA-mediated resistance against Tomato spotted wilt virus by exogenous application of double-stranded RNA largely depends on the selection of the viral RNA target region. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Melita, O.; Kaldis, A.; Berbati, M.; Reppa, C.; Holeva, M.; Lapidot, M.; Gelbart, D.; Otten, P.; Voloudakis, A. Topical application of double-stranded RNA molecules deriving from Tomato yellow leaf curl virus reduces cognate virus infection in tomato. Biol. Plant. 2021, in press. [Google Scholar]
- Holeva, M.C.; Sclavounos, A.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. In vitro produced dsRNA induces resistance against a severe Hellenic CMV isolate in tobacco and tomato. In Book of Abstracts, Proceedings of the 8th International Congress of Plant Molecular Biology, Adelaide, South Australia, 20–25 August 2006; POS-THU-580; Springer: Berlin/Heidelberg, Germany, 2006; p. 221. [Google Scholar]
- Holeva, M.C.; Sclavounos, A.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. External application of dsRNA of the capsid protein (CP) or 2b gene of CMV reduces the severity of CMV-infection in tobacco. In Book of Abstracts, Proceedings of the XIII International Congress of IS-MPMI, Sorrento, Italy, 21–27 July 2007; PS 12-574; Springer: Berlin/Heidelberg, Germany, 2007; p. 286. [Google Scholar]
- Borah, M.; Berbati, M.; Reppa, C.; Holeva, M.; Nath, P.D.; Voloudakis, A. RNA-based vaccination of Bhut Jolokia pepper (Capsicum chinense Jacq.) against Cucumber mosaic virus. Virusdisease 2018, 29, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Gago-Zachert, S.; Schuck, J.; Weinholdt, C.; Knoblich, M.; Pantaleo, V.; Grosse, I.; Gursinsky, T.; Behrens, S.-E. Highly efficacious antiviral protection of plants by small interfering RNAs identified in vitro. Nucleic Acids Res. 2019, 47, 9343–9357. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V. Exogenous RNAs for gene regulation and plant resistance. Int. J. Mol. Sci. 2019, 20, 2282. [Google Scholar] [CrossRef] [Green Version]
- Vogel, E.; Santos, D.; Mingels, L.; Verdonckt, T.-W.; Broeck, J.V. RNA Interference in Insects: Protecting beneficials and controlling pests. Front. Physiol. 2019, 9, 1912. [Google Scholar] [CrossRef] [Green Version]
- Christiaens, O.; Dzhambazova, T.; Kostov, K.; Arpaia, S.; Joga, M.R.; Urru, I.; Sweet, J.; Smagghe, G. Literature review of baseline information on RNAi to support the environmental risk assessment of RNAi-based GM plants. EFSA Supp. Publ. 2018, 173. [Google Scholar] [CrossRef]
- Roberts, A.F.; Devos, Y.; Lemgo, G.N.Y.; Zhou, X. Biosafety research for non-target organism risk assessment of RNAi-based GE plants. Front. Plant Sci. 2015, 6, 958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casacuberta, J.M.; Devos, Y.; du Jardin, P.; Ramon, M.; Vaucheret, H.; Nogué, F. Biotechnological uses of RNA interference in plants: Risk assessment considerations. Trends Biotechnol. 2015, 33, 145–147. [Google Scholar]
- Ramon, M.; Devos, Y.; Lanzoni, A.; Liu, Y.; Gomes, A.; Gennaro, A.; Waigmann, E. RNAi-based GM plants: Food for thought for risk assessors. Plant Biotechnol. J. 2014, 12, 1271–1273. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, J.G.; Duan, J.J. RNAi-based insecticidal crops: Potential effects on nontarget species. Bioscience 2013, 63, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Ossowski, S.; Schwab, R.; Weigel, D. Gene silencing in plants using artificial microRNAs and other small RNAs. Plant J. 2008, 53, 674–690. [Google Scholar]
- Schwab, R.; Ossowski, S.; Riester, M.; Warthmann, N.; Weigel, D. Highly specific gene silencing by artificial microRNAs in Arabidopsis. Plant Cell 2006, 18, 1121–1133. [Google Scholar] [CrossRef] [Green Version]
- Konakalla, N.C.; Kaldis, A.; Berbati, M.; Masarapu, H.; Voloudakis, A.E. Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco. Planta 2016, 244, 961–969. [Google Scholar]
- Numata, K.; Ohtani, M.; Yoshizumi, T.; Demura, T.; Kodama, Y. Local gene silencing in plants via synthetic dsRNA and carrier peptide. Plant Biotechnol. J. 2014, 12, 1027–1034. [Google Scholar] [CrossRef]
- Jiang, L.; Ding, L.; He, B.; Shen, J.; Xu, Z.; Yin, M.; Zhang, X. Systemic gene silencing in plants triggered by fluorescent nanoparticle-delivered double stranded RNA. Nanoscale 2014, 6, 9965–9969. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Demirer, G.S.; Zhang, H.; Ye, T.; Goh, N.S.; Aditham, A.J.; Cunningham, F.J.; Fan, C.; Landry, M.P. DNA nanostructures coordinate gene silencing in mature plants. Proc. Natl. Acad. Sci. USA 2019, 116, 7543–7548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, B.L.; Raghu, R.; Dangwal, M.; Byregowda, M.; Voloudakis, A. Exogenous dsRNA-mediated field protection against Pigeonpea sterility mosaic emaravirus. J. Plant Biochem. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Worrall, E.A.; Bravo-Cazar, A.; Nilon, A.T.; Fletcher, S.J.; Robinson, K.E.; Carr, J.P.; Mitter, N. Exogenous application of RNAi-inducing double-stranded RNA inhibits aphid-mediated transmission of a plant virus. Front. Plant Sci. 2019, 10, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmons, L.; Court, D.L.; Fire, A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene 2001, 263, 103–112. [Google Scholar] [CrossRef]
- Rizos, H.; Gunn, L.V.; Pares, R.D.; Gillings, M.R. Differentiation of Cucumber mosaic virus isolates using the polymerase chain reaction. J. Gen. Virol. 1992, 73, 2099–2103. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; p. 2100. [Google Scholar]
- Akbergenov, R.; Si-Ammour, A.; Blevins, T.; Amin, I.; Kutter, C.; Vanderschuren, H.; Zhang, P.; Gruissem, W.; Meins, F., Jr.; Hohn, T.; et al. Molecular characterization of geminivirus-derived small RNAs in different plant species. Nucleic Acids Res. 2006, 34, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.S.; Vazquez, F.; Robertson, D.; Meins, F., Jr.; Hohn, T.; et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing. Nucleic Acids Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef] [Green Version]
- Blevins, T.; Rajeswaran, R.; Aregger, M.; Borah, B.K.; Schepetilnikov, M.; Baerlocher, L.; Farinelli, L.; Meins, F., Jr.; Hohn, T.; Pooggin, M.M. Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense. Nucleic Acids Res. 2011, 39, 5003–5014. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer | Sequence 5′ to 3′ | PCR Thermal Profile | Reference |
|---|---|---|---|
| 5′CP | CTCGAATTCGGATCCGCTTCTCCGCGAG | (94 °C for 3 min) × 1 cycle, (94 °C for 1 min, 50 °C for 1 min, 72 °C for 1 min) × 35 cycles, (72 °C for 10 min) × 1 cycle | [69] |
| 3′CP | GGCGAATTCGAGCTCGCCGTAAGCTGGATGGAC | [69] | |
| CMV-2b-For2 | AGGGTTGAGCGTGTAAATTCC | As with 5′CP/3′CP oligonucleotides | This work |
| CMV-2b-Rev | CCGT(AT) AGCTGGATGGACAACC | This work | |
| CMV-CP-LIT-For * | GCGGAATTCTCATGGACAAATC | (94 °C for 5 min) × 1 cycle, (94 °C for 1 min, 65 °C for 1 min, 72 °C for 1 min) × 35 cycles, (72 °C for 10 min) × 1 cycle | This work |
| CMV-CP-LIT-Rev * | GCGGGATCCGTTCAAACTGG | This work | |
| T7 | TAATACGACTCACTATAGG | (94 °C for 3 min) × 1 cycle, (94 °C for 30 sec, 50 °C for 30 sec, 72 °C for 30 sec) × 25 cycles, (72 °C for 5 min) × 1 cycle | New England Biolabs |
| CMV-CP-F-Linker ** | GGGGATCCATGGACAAATCTGAATC | (98 °C for 30 sec) × 1 cycle, (98 °C for 10 sec, 60 °C for 30 sec, 72 °C for 30 sec) × 35 cycles, (72 °C for 10 min) × 1 cycle | This work |
| CMV-CP-R- Linker ** | GGGGATCCTCAAACTGGGAGCAC | This work | |
| CMV-2b-F-Linker ** | GGGGATCCATGGAATCGAACGAAG | (98 °C for 30 sec) × 1 cycle, (98 °C for 10 sec, 62 °C for 30 sec, 72 °C for 1 min) × 35 cycles, (72 °C for 10 min) × 1 cycle | This work |
| CMV-2b-R-Linker ** | GGGGATCCTCAAAACGCACCTTC | This work | |
| EcoRI-T7-Linker- BamHI *,# | GAGAATTCTAATACGACTCACTATAGGGGATCC | (98 °C for 30 sec) × 1 cycle, (98 °C for 10 sec, 55 °C for 30 sec, 72 °C for 30 sec × 35 cycles, (72 °C for 10 min) 1 cycle | This work |
| pUC/M13 Forward | CGCCAGGGTTTTCCCAGTCACGAC | (94 °C for 5 min) × 1 cycle, (94 °C for 1 min, 55 °C for 1 min, 72 °C for 1 min) × 35 cycles, (72 °C for 10 min) × 1 cycle | General primer |
| pUC/M13 Reverse | AGCGGATAACAATTTCACACAGGA | General primer |
| DNA Template for In Vitro Transcription | Primer Pair Used in 1st Step PCR | DNA Template Used in the 1st PCR * |
|---|---|---|
| CP fragment of 657 bp | CMV-CP-F-Linker CMV-CP-R-Linker | pCRIITopo::CP657 |
| 2b (336 bp) | CMV-2b-F-Linker CMV-2b-R-Linker | pCRIITopo::2b500 |
| Name of the Probe | Sequence (5′ to 3′) | Number of Nucleotides |
|---|---|---|
| Cmv1_3169_s * | TCCATCCAGCTTACGGCTAAAATG | 24 |
| Cmv1_3187_as * | GATTTCTCCACGACTGACCATTTT | 24 |
| Cmv1_171_s | GTTGATAAGACAGCTCATGAGCAGC | 25 |
| miR160a_as | TGGCATACAGGGAGCCAGGCA | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holeva, M.C.; Sklavounos, A.; Rajeswaran, R.; Pooggin, M.M.; Voloudakis, A.E. Topical Application of Double-Stranded RNA Targeting 2b and CP Genes of Cucumber mosaic virus Protects Plants against Local and Systemic Viral Infection. Plants 2021, 10, 963. https://doi.org/10.3390/plants10050963
Holeva MC, Sklavounos A, Rajeswaran R, Pooggin MM, Voloudakis AE. Topical Application of Double-Stranded RNA Targeting 2b and CP Genes of Cucumber mosaic virus Protects Plants against Local and Systemic Viral Infection. Plants. 2021; 10(5):963. https://doi.org/10.3390/plants10050963
Chicago/Turabian StyleHoleva, Maria C., Athanasios Sklavounos, Rajendran Rajeswaran, Mikhail M. Pooggin, and Andreas E. Voloudakis. 2021. "Topical Application of Double-Stranded RNA Targeting 2b and CP Genes of Cucumber mosaic virus Protects Plants against Local and Systemic Viral Infection" Plants 10, no. 5: 963. https://doi.org/10.3390/plants10050963