
The Potential Use of Peptides in the Fight against Chagas Disease and Leishmaniasis

The Azrieli Faculty of Medicine in the Galilee, Bar-Ilan University, Safed 1311502, Israel
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pharmaceutics 2024, 16(2), 227; https://doi.org/10.3390/pharmaceutics16020227
Submission received: 12 November 2023
/
Revised: 28 December 2023
/
Accepted: 30 January 2024
/
Published: 4 February 2024
(This article belongs to the Special Issue Drug Delivery Systems for Treatment of Leishmaniasis)
Abstract
:Chagas disease and leishmaniasis are both neglected tropical diseases that affect millions of people around the world. Leishmaniasis is currently the second most widespread vector-borne parasitic disease after malaria. The World Health Organization records approximately 0.7–1 million newly diagnosed leishmaniasis cases each year, resulting in approximately 20,000–30,000 deaths. Also, 25 million people worldwide are at risk of Chagas disease and an estimated 6 million people are infected with Trypanosoma cruzi. Pentavalent antimonials, amphotericin B, miltefosine, paromomycin, and pentamidine are currently used to treat leishmaniasis. Also, nifurtimox and benznidazole are two drugs currently used to treat Chagas disease. These drugs are associated with toxicity problems such as nephrotoxicity and cardiotoxicity, in addition to resistance problems. As a result, the discovery of novel therapeutic agents has emerged as a top priority and a promising alternative. Overall, there is a need for new and effective treatments for Chagas disease and leishmaniasis, as the current drugs have significant limitations. Peptide-based drugs are attractive due to their high selectiveness, effectiveness, low toxicity, and ease of production. This paper reviews the potential use of peptides in the treatment of Chagas disease and leishmaniasis. Several studies have demonstrated that peptides are effective against Chagas disease and leishmaniasis, suggesting their use in drug therapy for these diseases. Overall, peptides have the potential to be effective therapeutic agents against Chagas disease and leishmaniasis, but more research is needed to fully investigate their potential.
1. Introduction
Chagas disease (CD), leishmaniasis, and human African trypanosomiasis (HAT) are neglected tropical diseases (NTDs) caused by insect vector-borne protozoan parasites. Over 100,000 people die each year from NTDs worldwide, most living on less than USD 2 per day [1]. Chagas disease (aka American trypanosomiasis) is an infectious disease caused by the parasite Trypanosoma cruzi (T. cruzi), African Trypanosomiasis (“sleeping sickness”) is caused by Trypanosoma brucei (T. brucei), and leishmaniasis is caused by a protozoa parasite from over 20 Leishmania species [2,3]. Leishmaniasis is ranked second among all protozoan diseases for mortality after malaria [4]. The current drugs used against NTDs are suboptimal. For example, the only Food and Drug Administration (FDA)-approved medications for leishmaniasis are intravenous liposomal amphotericin B (L-AmB) for visceral leishmaniasis (VL) and oral miltefosine for cutaneous leishmaniasis (CL), mucosal leishmaniasis (ML), and VL caused by particular species [5]. The cost of treatment with amphotericin B deoxycholate varies widely between countries, ranging from less than USD 1 to USD 171 per day. Liposomal amphotericin B is a newer formulation that is less toxic but more expensive than deoxycholate amphotericin B. The cost of a single dose of 50 mg amphotericin B deoxycholate varies widely across countries. In Zambia, the Netherlands, Russia, and Chile, it is less than USD 1 per day. In India, prices range from USD 1 to USD 10 per dose, while in the United States (US), a single dose costs approximately USD 50 [6,7]. Amphotericin B deoxycholate is well known for its severe and potentially lethal side effects (e.g., fever, shaking, chills, flushing, loss of appetite, and many more). As a result, the recommended course of treatment is frequently not followed, resulting in the development of resistance [8,9,10]. Moreover, there are no effective vaccines for leishmaniasis [10]. Similar challenges are also associated with drugs for Chagas disease. The safety profile of the currently available drugs is far from ideal, with frequent adverse events and high drug discontinuation rates. There are only two drugs available for treating Chagas disease: benznidazole (BZN) and nifurtimox (NFX). Although these drugs are effective in the acute phase, they have limitations for chronic infections, and both drugs are restricted due to their toxic effects. For example, Nifurtimox is no longer used in many places since it causes neurological disorders or psychiatric episodes. Therefore, there is an urgent need to develop effective, safe, and affordable drugs [11].
Drug discovery aims to identify novel compounds that specifically and significantly change the course of the diseases they are intended to treat. There is a pressing need to identify new therapeutics for NTDs and novel approaches to drug design and development are needed. However, due to the lack of commercial benefits, pharmaceutical companies are discouraged from investing in NTD product development [12,13]. Recently, virtual screening, a convenient and cost-effective solution in the early stages of drug discovery, has been used for the discovery of anti-trypanosomal and anti-leishmanial drugs with some promising results [14]. Nevertheless, computational methods should be validated experimentally [15]. Other examples are target-based or phenotype-based drug discovery, which are probably the most effective strategies for drug discovery for NTDs [16]. Another appealing approach is repurposing existing drugs. Drug repurposing discovers new disease indications for previously approved drugs that benefit from accessible compound libraries. This has yielded some promising compounds for various NTDs. Yet, repurposing an existing drug poses a major challenge in identifying an appropriate therapeutic indication for it. It involves multiple factors, such as technology, patents, investment, and clinical trials. Recent advances in production, modification, and analytical technologies have led to an increased interest in peptides as therapeutics for NTDs [17,18].
2. Leishmaniasis and Chagas Disease
2.1. Life Cycle of Trypanosomatidae
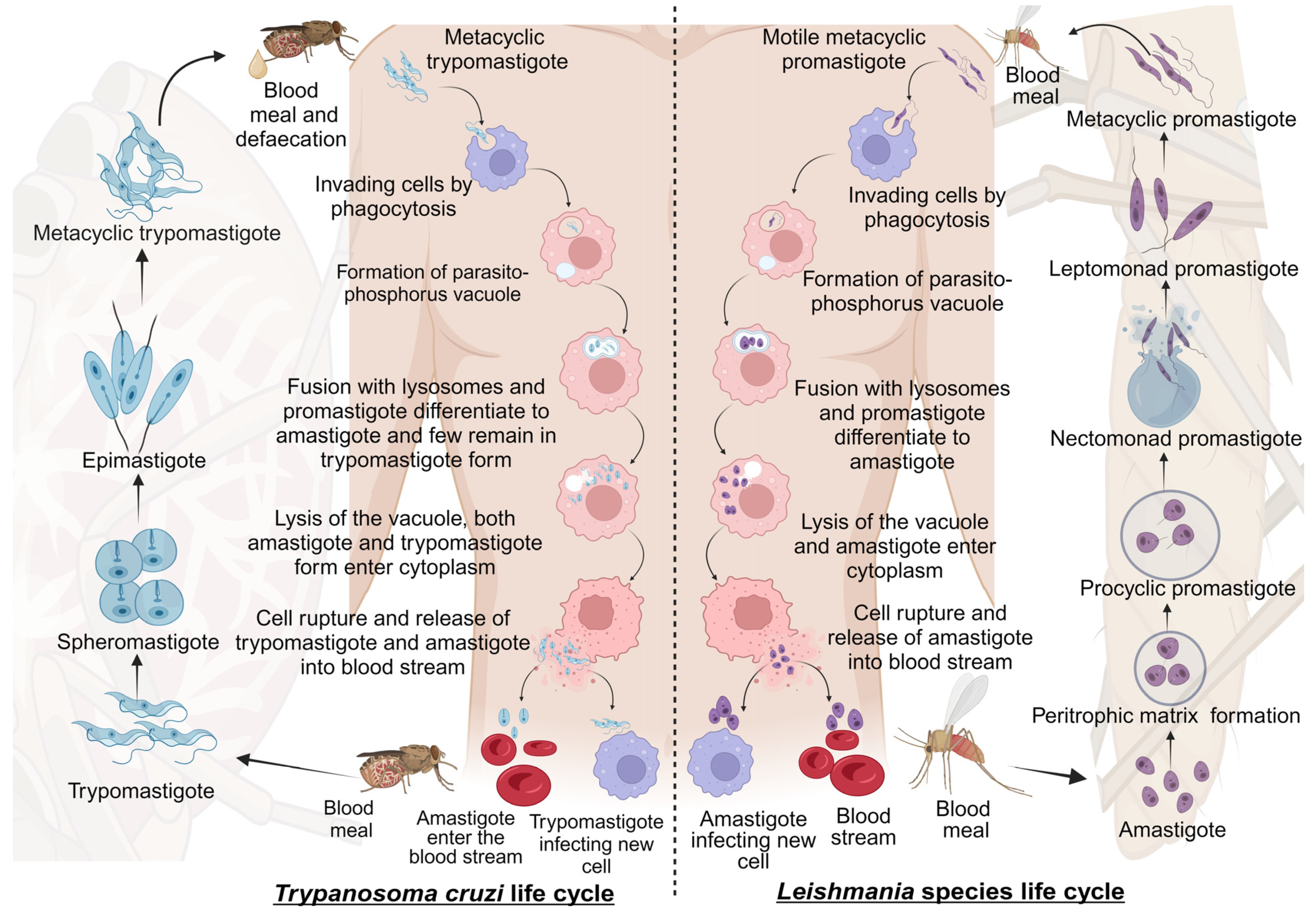
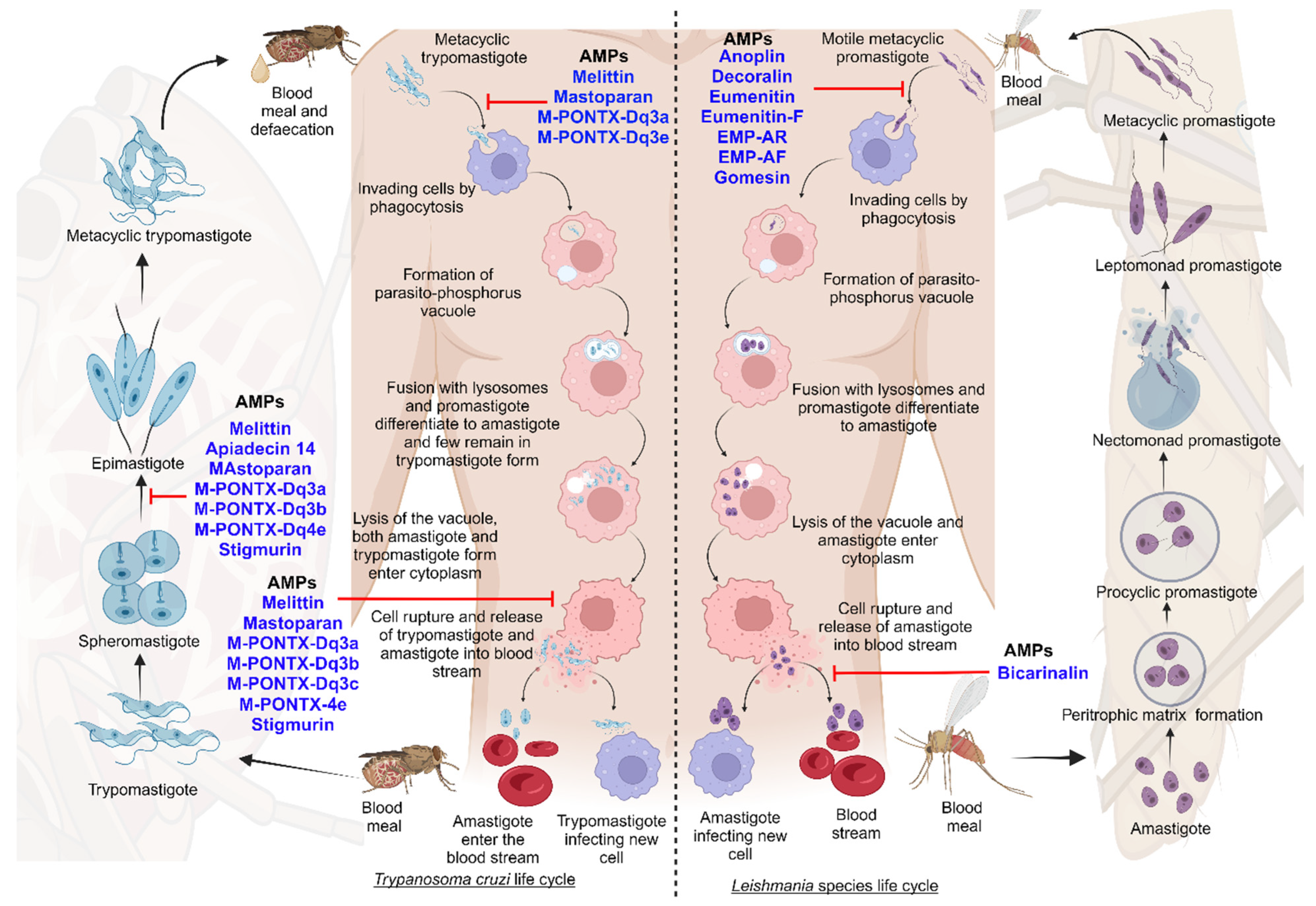
Leishmaniasis and Chagas disease are caused by trypanosomatid parasites. In mammals, leishmaniasis and Chagas disease are transmitted through the sandfly and the triatomine bug, respectively. The differentiation of these trypanosomatids occurs partially in the vector and the host. This is accomplished with different developmental stages giving rise to the infective stage of the parasite that infects mammals [19]. We discuss the stages of development and differentiation of the parasites in the vector and host involved in the life cycles of both leishmaniasis and Chagas disease (Figure 1).
2.1.1. Life Cycle of Leishmania Species
Leishmaniasis is caused by the protozoan parasite of the genus Leishmania belonging to the Trypanosomatidae family. It is spread through female sandflies of the genera Phlebotomus and Lutzomyia [20]. The parasite’s life cycle is complex as it includes vector and mammalian reservoirs for its morphogenesis; hence, it resembles a digenetic life cycle [21]. The parasite’s entry is during the blood meal event, where motile metacyclic promastigotes are transferred into the blood through sandfly saliva [22]. Next, the metacyclic promastigotes migrate to healthy macrophages. Proteins on the parasite flagella interact with the macrophagic cell membrane and gain entry through phagocytosis by forming pseudopods infecting macrophages [23,24]. In the macrophage, promastigote forms differentiate from motile forms, which have elongated cell shapes and long flagellums, to amastigote forms, which have short flagellums [1]. After fusing with the parasitophorous vacuole, the host lysosomes create an environment for parasite replication. Finally, the macrophages rupture, and the amastigotes are released into the bloodstream [22].
The blood meal is digested in the sandfly midgut where the amastigotes cluster together forming nest cells. The nest cells are surrounded by an enclosed structure called the peritrophic matrix, which protects the amastigotes from the insect’s gut environment [22,25]. The amastigotes transform into elongated flagellated procyclic promastigotes inside the peritrophic matrix. It is their first replicative stage. After 48–72 h, the procyclic promastigotes differentiate into long motile nectomonad promastigotes that break open the peritrophic matrix and enter the midgut lumen. Nectomonad promastigotes transform into short motile nectomonad promastigotes called leptomonad promastigotes and adhere to the microvilli of the midgut epithelium of the insect [26,27]. Here, the leptomonad promastigotes migrate and concentrate at the anterior stomodeal valve. Here, they differentiate into metacyclic promastigotes that infect mammalian hosts. During the transformation to metacyclic stages, they secrete a gel called promastigote secretory gel (PSG), creating a “block fly” that forces the sandfly to feed on the blood and transfer metacyclic promastigotes from the vector to the mammalian host, thereby starting the cycle all over [28].
2.1.2. Life Cycle of Trypanosoma cruzi
Chagas disease is a protozoan disease caused by T. cruzi of the genus Trypanosoma belonging to the Trypanosomatidae family. It is caused by the parasitic defecation of a blood-sucking vector called the triatomine bug (aka the kissing bug). As a result of the undetermined spread of feces on the wound by human actions, the parasite enters the host [29]. The first step upon entry into the host is adhesion. This is where metacyclic trypomastigotes attach to host cellular receptors with their flagella. Parasite flagella consists of molecules belonging to the Apicomplexa family that adhere to the host cell membrane. By binding glycoproteins to the cell membrane, metacyclic trypomastigotes mobilize calcium necessary for entry into the cytoplasm [30]. Next, the adhered parasite recognizes and alters ligands such as lectin-like molecules on the surface for safe internalization via endocytosis mediated by clathrin and caveolar lipids [30]. Also, internalization by phagocytosis, micropinocytosis, and circular dorsal ruffle formation has been reported [31]. The parasite resides safely within the parasite phosphorus vacuole formed by the plasma membrane and then fusses with the lysosome. When lysosomes fuse, parasites can differentiate from trypomastigotes to short globular organisms with small flagella, known as amastigotes, whereas some remain trypomastigotes [32]. After or during differentiation, the amastigotes secrete hemolysin, also called Tc-Tox, and trypomastigotes secrete trans-sialidase/neuraminidase, required for fragmentation of the parasite phosphorus vacuole, and enter the cytoplasm. As this process occurs during the differentiation of metacyclic trypomastigotes into amastigotes, both forms are observed in the cytoplasm. The intracellular amastigote form further differentiates to form bloodstream trypomastigotes. Bloodstream trypomastigotes rupture the host cell and enter the bloodstream, infecting other cells [32,33,34].
During the blood meal of an infected individual, trypomastigotes enter the triatomine bug’s gut. Here, most bloodstream trypomastigotes are digested. Surviving parasites differentiate and transform into small spherical structures called spheromastigotes with a small extended flagellum. In the midgut, the spheromastigotes elongate and further extend their flagella, differentiating them into epimastigotes [35]. The long-flagellated epimastigotes migrate to the hindgut and anchor to the perimicrovillar membranes of the hindgut intestine. This anchorage initiates the non-infective epimastigotes differentiation to infective metacyclic trypomastigotes which detach from the intestinal membranes and are migrated to the rectum where they are excreted in the feces and urine of the triatomine bug, infecting mammalian hosts [36].
2.2. Epidemiology of Trypanosomatids
Trypanosomatids are a family of parasites causing infectious diseases categorized into trypanosomiasis and leishmaniasis. Trypanosomiasis is further subdivided into American and African diseases, also known as Chagas disease and African sleeping sickness, respectively. Leishmaniasis is also further classified based on its prevalent geographical location. Trypanosomiasis and leishmaniasis are neglected diseases widely observed in underdeveloped countries, mainly in Southern Asian countries, African countries, and South American countries [36]. Various factors influence trypanosomiasis and leishmaniasis species and their development, including carrier vectors, vertebrate hosts, biochemical enzyme patterns, parasite phylogeny, and geographical distribution [37,38]. Table 1 lists the main Leishmania and Trypanosoma species.
2.3. Clinical Complications and Conditions of Leishmaniasis and Chagas Disease
Leishmaniasis and Chagas disease cover a broad disease spectrum that affects humans and other mammals. Clinical manifestations and treatment options for leishmaniasis and Chagas disease are limited due to various factors, including the complexity of the parasitic species and subtypes. This makes it very difficult for doctors to determine the disease’s type and choose the optimal treatment plan [55].
2.3.1. Leishmaniasis
Cutaneous Leishmaniasis (CL)
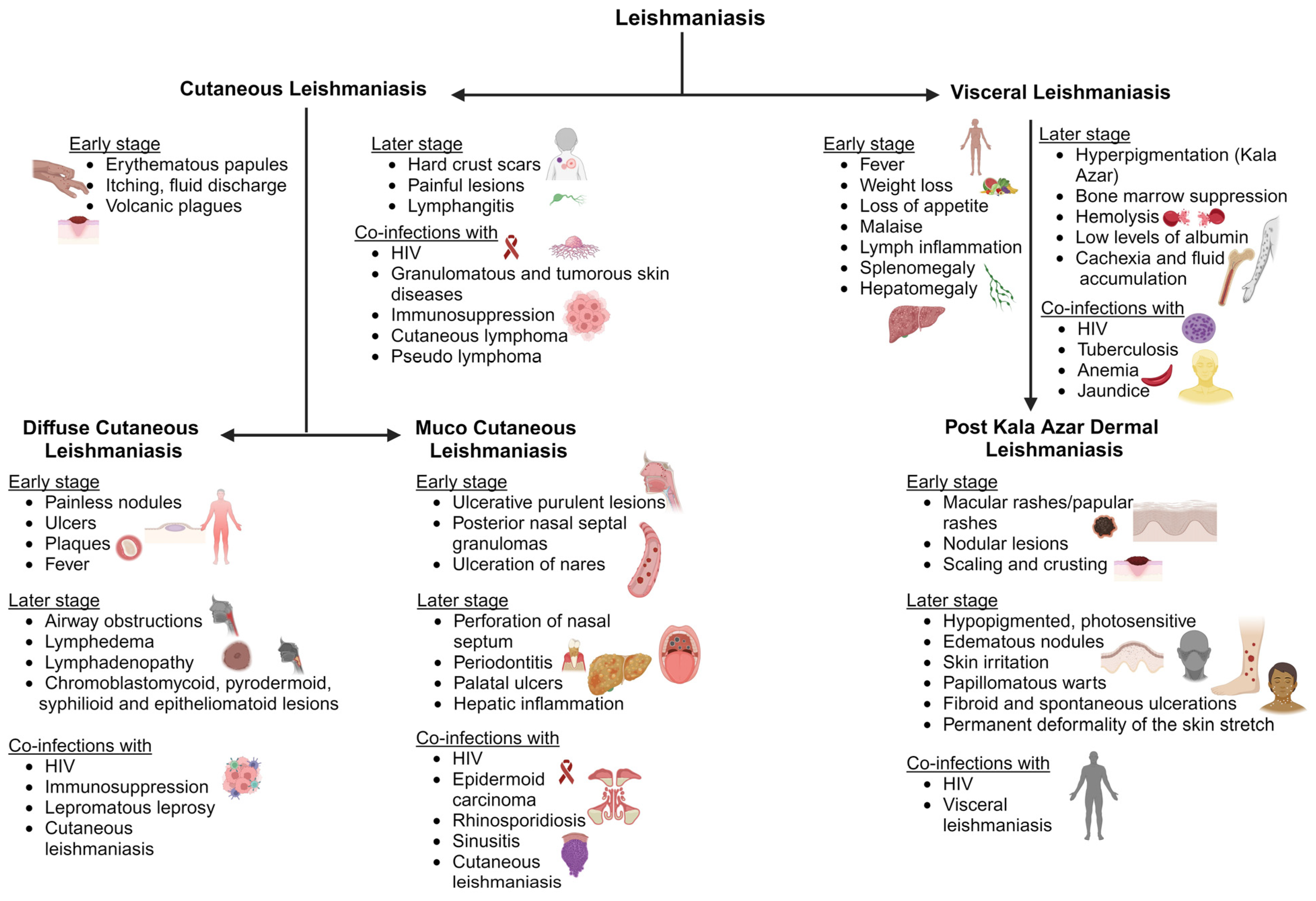
Cutaneous leishmaniasis (CL) is the most typical clinical symptom of Leishmania infection. Initial signs include single or multiple skin lesions localized to exposed body parts such as the face and may result in social stigma [56]. Numerous painless brownish erythematous papules with multiple nodules that resemble plague are seen after an incubation period of between two and seven months. It is followed by itchy and painful fluid discharge, warmth, swelling, and fever, which further develop into volcanic plagues associated with lymphangitis that differentiate from other ulcerative plagues (Figure 2). Bacterial and fungal infections are common on exposed cutaneous lesions, causing secondary infections [57]. There are findings of co-infection with diseases like helminthiasis, leprosy, Human African trypanosomiasis, and Chagas disease [58]. A few reports of leishmaniasis co-infection with immunosuppressed patients under Human immunodeficiency virus (HIV) treatment have also been observed among patients. There have also been granulomatous and tumorous skin diseases, including subcutaneous mycosis, deep mycosis, cutaneous lymphoma, pseudolymphoma, basal cell carcinoma, and squamous cell carcinoma, among patients (Figure 2) [59]. Due to these clinical complications, doctors cannot use a specific treatment regimen to treat the disease.
Diffuse Cutaneous Leishmaniasis (DCL)
Diffuse cutaneous leishmaniasis (DCL) is widely observed in patients with a suppressed cell-mediated immune response against the invading parasite, also termed anergy [60]. DCL is characterized in patients with a low count of CD4 T-cells and immediately after antiretroviral drug therapy [61]. Several Leishmania (L.) species such as L. aethiopica, L. major, L. amazonensis, L. mexicana, and L. braziliensis cause DCL [62]. An initial symptom is erythematous lesions with a well-defined periphery that extend to mucocutaneous junctions including nodules, ulcers, and plaques [63]. As the lesions progress, they spread to the face, buttocks, and extremities and ulcerate the entire surface. The nodules across the nasopharynx and oropharynx cause airway obstructions and may be associated with lymphedema and lymphadenopathy, where the lesions are observed as sporotrichoid in morphology [64,65]. The lesions also resemble other chronic lesions [66]. DCL is associated with immunocompromised diseases such as acquired immunodeficiency syndrome (AIDS) and lepromatous leprosy [67]. These findings are a major clinical observation in defining Leishmania infection.
Mucocutaneous Leishmaniasis (ML)
The majority of mucosal/mucocutaneous leishmaniasis (ML) is transmitted by L. braziliensis, L. amazonensis, L. panamensis, L. infantum, and L. guyanensis [68]. It is initially painless, but ulcerative and purulent lesions develop. They continue as erythema, edema, posterior nasal septal granulomas, and ulceration of the nares [69,70]. It spreads to the oral cavity of the oropharynx and larynx. This causes perforation of the nasal septum, periodontitis, palatal ulcers, and gingivitis, damaging the vocal cartilage and resulting in speech abnormalities (Figure 2) [71]. Lymphedema leads to systemic infection causing fever and liver inflammation. Secondary complications include airway obstruction and fungal and bacterial infections [72]. The associated infections include epidermoid carcinoma, rhinosporidiosis or sinusitis, and AIDS, where a differential diagnosis is required (Figure 2) [69]. ML is associated with cross- and co-infections, which can worsen physical and psychological conditions.
Visceral Leishmaniasis (VL)
Visceral leishmaniasis (VL) is usually caused by L. donovani and L. infantum. These species are commonly found across North and East Africa, the Indian subcontinent, and Southern American nations. Infection is prevalent in all age groups but most prevalent in immune-suppressed patients and children [11]. The parasite incubates for about two to six months and leads to chronic infection lasting for several years. The initial clinical observations include fever, weight loss, loss of appetite, and a malaise that progresses to a few months and is followed by enlarged lymph nodes, splenomegaly, and hepatomegaly [72]. The disease is also called Kala Azar due to hyperpigmentation of the skin and visceral regions. These hyperpigmentation patches are observed as dark patches stretching the skin surface associated with abdominal congestion and pain as the infection continues. The patient also suffers from bone marrow suppression, hemolysis, low albumin levels, jaundice, thrombocytopenia, cachexia, fluid accumulation, and bleeding from the mucosal cavities [73]. This may be followed by hemophagocytic lymph cytosis and intravascular coagulation [74].
Infections of the oral mucosa, nasal, and gastrointestinal tracts are prone to bacterial and fungal contamination. This leads to sepsis and pneumonia which is fatal in children and geriatrics. It could cause abortion or congenital leishmaniasis in pregnant patients [75]. HIV infection, tuberculosis, anemia, and jaundice cases are frequently reported as co-infections with VL and increase VL mortality in southern Asian and African countries [76,77]. Post kala azar dermal leishmaniasis is also common after recovery from VL, which may lead to other medical complications.
Post Kala Azar Dermal Leishmaniasis (PKDL)
Post kala azar dermal leishmaniasis (PKDL) is mostly observed post VL treatment. It could be defined as the final stage of VL infection before complete recovery from the disease. It is mostly observed in Asian and African countries as VL is a concern in these regions [78]. African and Asian PKDL can be differentiated by macular rashes in Asian countries and papular rashes in African countries [79]. The onset of PKDL symptoms usually starts a few months post VL treatment. This mimics a chronic skin infection that lasts for three to four years if untreated. The initial clinical presentations include dispersed hyperpigmented maculopapular and nodular lesions on the face which spread to the chest and arms. These lesions cover the entire length of the skin with scaly skin and dense nodular skin rashes (Figure 2). The lesions or rashes may be ulcerative, erythematous, or keloidal [80]. The nodules and papules are hypopigmented, photosensitive, and edematous, with a succulent nature, which irritates the skin. They appear warty, papillomatous, and fibroid, with spontaneous ulcerations in the peritoneal area, chin, and parts of the face [81]. As the infection spreads to sensitive parts such as the tongue, groin, genitalia, and axilla, it can cause permanent deformity in the patient. Treatment approaches should be enhanced against infection and active surveillance should be carried out for a better understanding of how diseases spread.
2.3.2. Chagas Disease
Clinical outcomes of T. cruzi infection are divided into two categories: acute and chronic stages. Before the infection progresses to the chronic stage, it incubates in the intermediate stage, which mediates chronic infection. We discuss each clinical complication and condition of Chagas disease in detail below.
Acute Chagas Disease
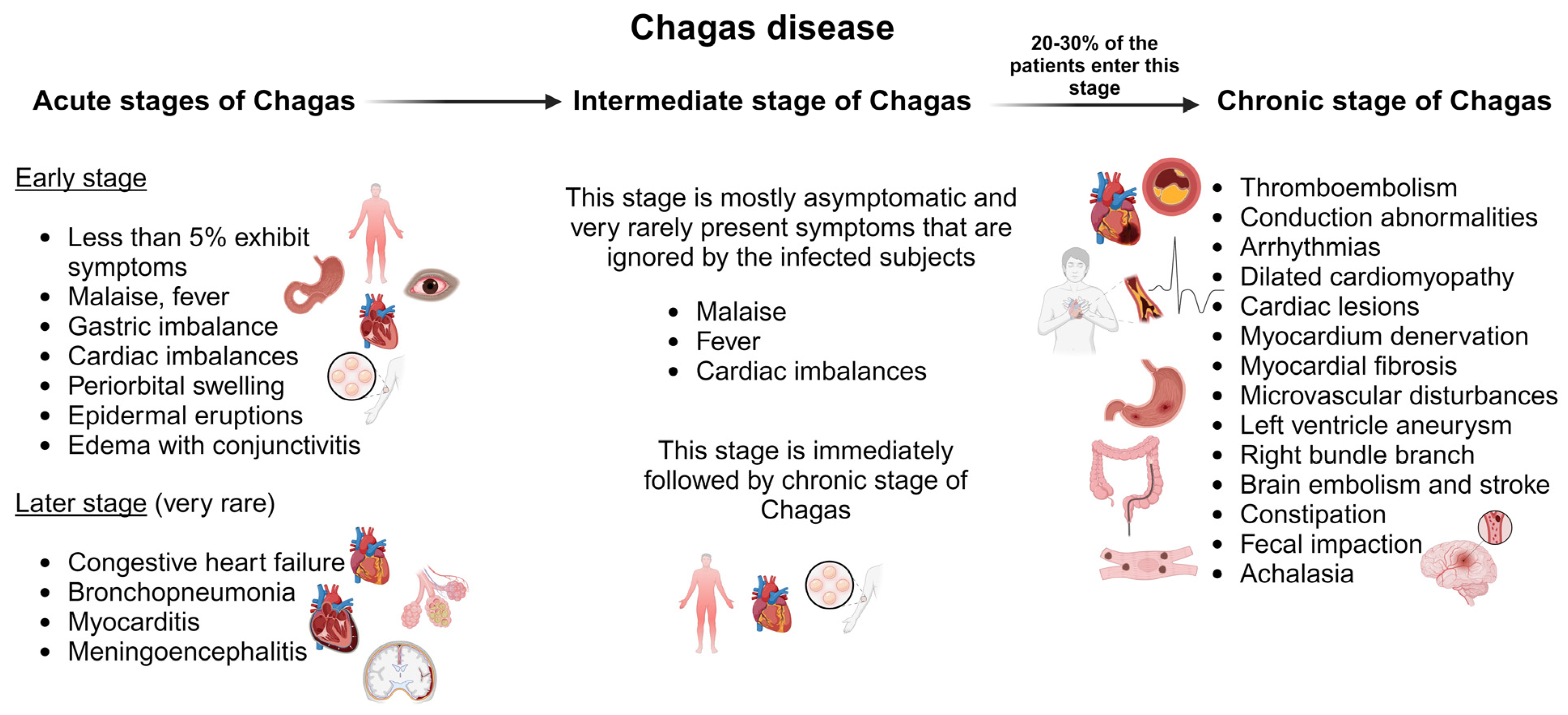
The acute stage of Chagas disease is mostly asymptomatic and unnoticed by the patient. Symptoms are rarely observed after 30 days of incubation. During this stage, invasive trypomastigotes can be detected in fresh blood smears [82]. Less than 5% of patients in their acute stage of infection display symptoms such as malaise, fever, gastric imbalance, cardiac imbalances, inflammation at the inoculation site, periorbital swelling, epidermal eruptions, and edema with conjunctivitis indicating cutaneous infection (Figure 3) [83]. It might be accompanied by anemia, thrombocytopenia, hepatomegaly, and splenomegaly [84]. Death during this stage of infection is very rare and is primarily due to congestive heart failure, bronchopneumonia, myocarditis, or meningoencephalitis [85]. Children are more prone to death and are affected by myocarditis [86]. The acute stage persists for three–four months and then the infection enters the intermediate stage, which lasts for six–eight weeks.
Chronic Chagas Disease
The immediate phase after the acute stage is asymptomatic. Clinically, the patient has a normal electrocardiogram (ECG), gastrointestinal findings, and healthy physical condition but positive serological findings for infection. The intermediate phase of infection is followed by the chronic phase of infection, which is fatal [87]. Around 20–30% of patients with acute and intermediate infections progress to the chronic phase of Chagas disease, which can have highly symptomatic symptoms. This includes thromboembolism, conduction abnormalities, arrhythmias, and heart failure due to dilated cardiomyopathy (Figure 3) [82]. The severity of the disease depends on the duration, location, and nature of the cardiac lesions. The parasite resides in the cardiac tissues and causes immune activation in the myocardium. This causes denervation, myocardial fibrosis, microvascular disturbances, and myocardial injury [88,89]. Cardiomyopathies in Chagas disease manifest as ventricular remodeling, dysautonomia, and imbalances in cardiac perfusion [90,91]. Chronic Chagas cardiomyopathy is defined as a left ventricle aneurysm. This leads to an increase in left atrial pressure resulting in systolic dysfunction and reduced left ventricular filling [92]. The right bundle branch block is a common ECG found in chronic Chagas disease and is mostly found in association with an anterior fascicular block [84]. The chances of sudden death under this condition are very high due to ventricular tachyarrhythmia [93].
Damage to the brain and stroke are very common in Chagas disease due to hypoxia and reduced blood supply to the brain [84]. Brain embolism is a clinically recognized event associated with stroke in asymptomatic patients [82]. Other clinical manifestations include mega syndrome, which is a result of the denervation of tubular structures of the enteric nerves that modulate normal gastrointestinal function directed towards acute infection in the rectum, colon, and esophagus (aka oesophagus). In the colon and rectum, it leads to bowel dysmotility, constipation, and fecal impaction. In the esophagus, it causes achalasia. Also, it causes dysphagia, odynophagia, weight loss, idiopathic achalasia, regurgitation, cough, and chronic aspiration [83].
2.4. Pathways Involved in Leishmaniasis and Chagas Disease
Trypanosomatids survival and differentiation involve various pathways. The parasite cannot synthesize all the metabolites needed for its survival, so it interferes with the host’s biochemical mechanisms to fulfill its nutritional needs. There are enzymes that resemble host intermediates and are salvaged into the parasite to create a safe environment, escape the immune system, and develop for their own purposes [94]. Some of the biochemical pathways affected by trypanosomatids infection are discussed below.
Sterols serve as a key metabolic element for Leishmania and T. curzi survival. Both these parasites increase ergosterol production by interfering with host sterol biosynthesis [94]. Leishmania and T. curzi parasites infest macrophagic sterol levels for parasitic differentiation, cellular shape, and division. This results in lowering cholesterol levels in macrophage plasma membranes [94,95]. In Chagas disease, cholesterol is employed in the endocytic vesicle and cytostome of amastigotes [96]. It leads to CD40 signaling pathway disturbances and alters Interleukin 12 (IL-12) production. It also increases arginase I expression, which inhibits NO synthesis and favors parasite survival and differentiation [97,98].
Both Leishmania and T. curzi are incapable of synthesizing the purines required for their differentiation and survival; hence, they salvage the purine molecules used by the host for their living [99]. Purine phosphoribosyl transferase assists Leishmania and T. curzi salvage purines from the phosphorus vacuole. It helps in catalyzing the formation of guanosine monophosphate and inosine monophosphate, which is required for parasite survival [100,101]. Typically, hypoxanthine–Guanine Phosphoribosyltransferase (HGPRT) and xanthine Phosphoribosyltransferase (XPRT) enzymes are involved in purine salvage [102]. Other enzymes such as adenine phosphoribosyl transferase (APRT), hypoxanthine–guanine–xanthine phosphoribosyl transferase (HGXPRT), adenosine kinase (AK), and nucleoside hydrolase (NH) were also identified to play a major role in acquiring purines from the host in cases of both Leishmania and T. curzi infections [103,104].
Cell surface proteins are anchored by glycosylphosphatidylinositol (GPI) molecules. GPI molecules, including Lipophosphoglycan (LPG) and Gylcoinositol phospholipids (GIPL), are synthesized by Leishmania and T. curzi, which help them anchor to macrophages and internalize them [105]. Most GPI anchors require trimannose backbones and dolichol–phosphate–mannose (DPM), which act as mannose donors [106]. These GPI molecules help host and parasite communication and immune invasion [107]. Mucins also play a role in anchoring parasite development. Trans-sialidase is one such mucin responsible for active anchorage and infectivity [108]. Both Leishmania and T. curzi contain mucin, but T. curzi contains more than Leishmania [109]. Molecules such as phosphatidylinositol, inositol phosphorylceramide, glycoproteins 63, glycoprotein 82, and glycoprotein 30 also assist in parasite anchorage and internalization of both Leishmania and T. curzi parasites. Also, these molecules facilitate the mobilization of intracellular calcium and the exocytosis of lysosomes, leading to a cascade of signaling between the host and the parasite for cellular invasion [110].
Folic acid and pteridines play a major role in Leishmania and T. curzi metabolic interventions [111]. Tetrahydrofolate is essential for purine, thymidylate, and pantothenate biosynthesis. Also, it is required for RNA protein formation and is involved in various signaling cascades [112]. Dihydrofolate reductase (DHFR) and Thymidine synthase (TS) act as bifunctional enzymes as they are both connected to N- and C-terminals, respectively, and separated by a linker peptide [113]. DHFR-TS helps protect dihydrofolate and drives cellular colocalization and parasite survival [114]. Pteridine reductase I (PTR-1) also plays a role in the conversion of biopterin to tetrahydrobiopterin when DHFR-TS inhibitors are used. PTR-I is activated when DHFR-TS is inhibited or malfunctioning and is responsible for parasite survival and a proper survival environment for their growth [115]. Trypanothione (TSH2) is a principal metabolite that protects Leishmania and T. curzi parasites from oxidative stress and is responsible for parasite differentiation. The formation of TSH2 is mediated by glutathionyl spermidine synthase (GSPS) and trypanothine synthase (TRYS). TSH2 is involved in the reduction in host glutathione and decreases macrophage oxidative stress during infection [116]. TSH2 is maintained at reduced levels as required by the parasites by trypanothione reductase (TR) [117]. TR shares 67% similarities among Leishmania species and 80% similarities among trypanosome species [118]. In Leishmania, TSH2 catalyzes the hydroperoxide reduction with the assistance of two proteins, namely tryparedoxin (TXN) and tryparedoxin-dependent peroxidase (TDPX). This reduces oxidative stress and enhances parasite survival rates [119]. TSH2 also possesses the properties of xenobiotics and endobiotic neutralization and enhances the reduction in ascorbate and iron–sulfur complexes [120].
N-(2-amino2-hyroxymethyl) lysine, also called hypusine, is found in two eukaryotic proteins, eukaryotic translation initiation factor (eIF)-5A1 and eIF5A2 [121]. eIF5A is responsible for cell cycle regulation, apoptosis, translation, elongation, and termination [122]. The post-translational modification leads to the binding of hypusine with lysine residues of eIF5A proteins by the transfer of 4-aminobutyl moiety from spermidine to the lysine chain, which is aided by the enzymes deoxyhypusine synthase (DHPS) and deoxyhypusine hydroxylase (DOHH), also known as hypusination [123]. Arginine serves as a precursor for eIF5A, which is essential in hypusine synthesis [124] and determines Leishmania and T. curzi infection rates. Arginine is translocated to parasites using amino acid permease 3 (AAP3) [125]. Arginine plays a role in the polyamine pathway. In Leishmania, the promastigotes accumulate arginine, utilize it in polyamine biosynthesis, and regulate arginine transport under conditions of arginine deprivation [126].
3. Drug Discovery Strategies and Insights into Current Therapeutics
3.1. Approaches to Drug Development against Leishmaniasis and Chagas Disease
Drug discovery strategies for leishmaniasis and Chagas disease have evolved based on various approaches, such as structural biology, genetic engineering, molecular modeling, and high-content screening platforms [127,128]. Identification of functional molecular targets is crucial to target-based drug discovery [129]. Subsequently, molecules are crafted to disrupt specific targets crucial for parasite survival [16]. High-throughput, targeting-based drug screening assays were developed based on target identification from T. cruzi genome data [130]. In drug development, two different mechanisms are assumed for compound identification such as target-based drug discovery (TDD) or phenotype-based drug discovery (PDD) [16]. Identifying new drugs for inadequately treated medical conditions relies on TDD [129]. However, PDD involves evaluating diverse chemicals in contradiction to the pathogen’s phenotype in a biological system and animal model. For example, benzothiophene analogs emerged as novel agents from a phenotypic screen against GSK’s kinetoboxes [131,132]. However, the increased cost and high rates of failure of traditional drug discovery and development approaches have provoked the community to explore alternative approaches, such as computer-aided drug discovery which comprises structure-based, ligand-based, and system-based approaches to drug discovery [133,134].
Structure-based drug discovery (SBDD) is the use of 3D structures of molecular targets to increase ligand–receptor complementarity. It is mainly obtained by biophysical techniques and used in research for the determination of pharmacological targets [135]. For example, virtual screening approaches have recently been undertaken using imidazole–pyridine as a reference structure [14]. In addition, the identification of oxidosqualene cyclase as a novel molecular target in Leishmania could lead to promising programs to discover new agents for leishmaniasis treatment [38]. Finally, the ligand-based drug design (LBDD) approach also identifies key characteristics that contribute to biological activity while improving or identifying new chemotypes [136,137,138].
3.2. Current Drugs for Leishmaniasis and Chagas Disease
The chemotherapy currently used for leishmaniasis and Chagas disease includes a limited range of drugs that are used in combination or as monotherapy [55,139]. Currently, there is no vaccine available to prevent leishmaniasis or Chagas disease. Treatment plans vary depending on factors such as disease type, parasite species, and geographic location. Each year, thousands of compounds are tested for leishmaniasis and Chagas disease drug discovery. For the discovery of new treatment leads, high-throughput screening (HTS) campaigns have been conducted [140]. To identify new drug candidates for diseases like leishmaniasis and Chagas, the Drugs for Neglected Diseases Initiative (DNDi) is actively incorporating natural compounds and synthetic compounds into its portfolio. Several pharmaceutical companies collaborated with the DNDi in 2015 to launch the NTD Drug Discovery Booster [141]. As part of its ongoing efforts, the DNDi is initiating the ‘Chagas Hit-to-Lead’ project, which aims to identify promising leads in animal models of the disease, as well as develop innovative approach to discover possible drugs for treatment [141,142].
The majority of the mechanisms of action of drugs used today to treat leishmaniasis and Chagas disease are unknown. Further, many of them are ineffective, and their overuse is associated with a variety of emerging resistance patterns, as well as severe side effects [143,144]. Therefore, the development of novel drug discovery programs for anti-leishmanial and anti-trypanosomal compounds is essential [145,146]. The NTD Drug Discovery Booster (DDB) was launched in 2015 to avoid early-stage commercial obstacles between pharmaceutical participants, allowing the DNDi to search millions of unique compounds and use computational methods to refine the search iteratively in the hunt for new treatment leads [147]. Since its creation in 2015, the NTD-DDB has launched 45 iterations around 22 seed compounds, with 13 hit series released, of which 6 have progressed to in vivo proof-of-concept studies for Chagas disease, leishmaniasis, or both. Two compounds from one hit series have demonstrated efficacy in a leishmaniasis infection model: one was transitioned into lead optimization and is progressing to the pre-clinical stage. In 2017, more than 20 novel series were identified and are currently being processed [147].
3.2.1. Leishmaniasis
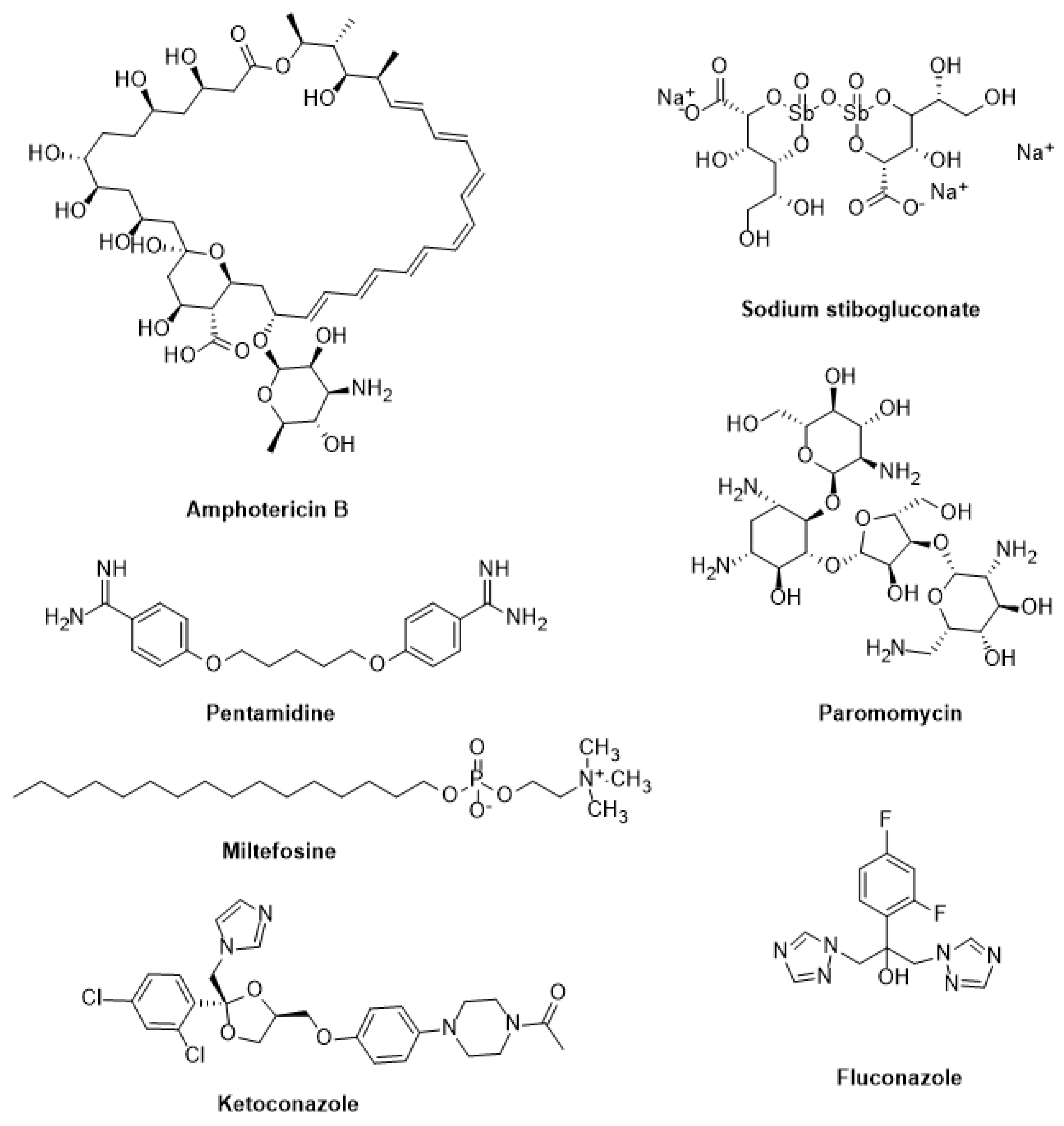
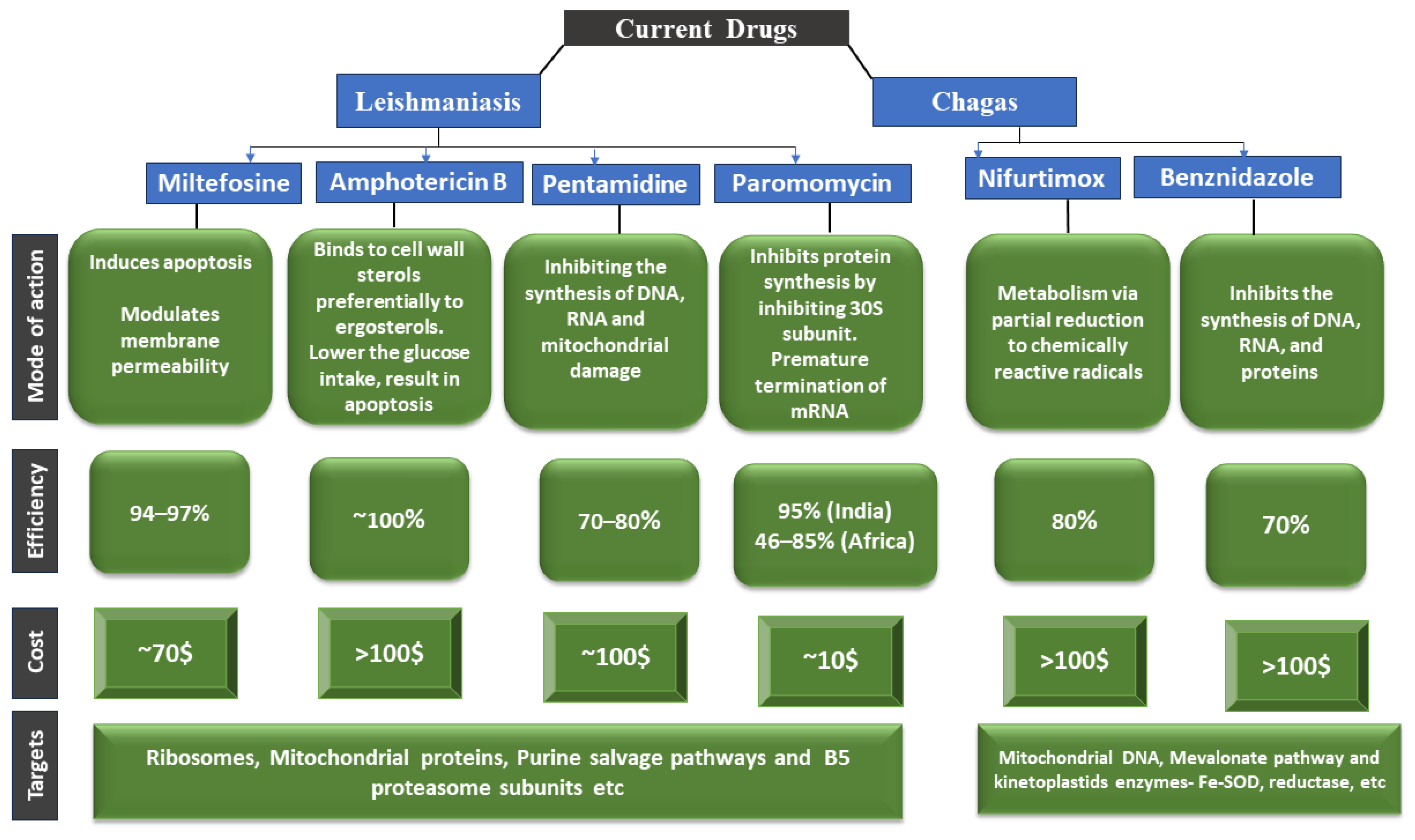
Sodium stibogluconate and meglumine antimoniate have been used extensively to treat visceral, cutaneous, and mucocutaneous leishmaniasis for decades [148]. Yet, their parenteral therapy is long, the drugs are toxic, and resistance is on the rise. Therefore, other medications such as pentamidine, paromomycin, fluconazole, ketoconazole, miltefosine, and amphotericin B have been repurposed (Figure 4) [149]. Additionally, it has been noted that amphotericin B therapy does not result in a sterile cure and is linked to rising kala azar leishmaniasis rates, as well as the emergence of resistant parasites in clinical settings [150]. Although repositioning efforts have shown promise in discovering new treatments for this disease, the identification of novel therapeutic targets and the development of effective leishmanicidal drugs should be prioritized [151].
Miltefosine: Miltefosine (hexadecyl-phosphocholine) is the “only oral drug” available for leishmaniasis chemotherapy [152]. It is an amphipathic alkyl phosphocholine drug with a polar phosphocholine group and a long alkyl chain. Originally used to treat breast cancer, it has shown great promise as an anti-leishmanial agent since the 1980s, particularly in antimonials-resistant cases [153]. Miltefosine works primarily by (a) changing the composition of plasma membranes by inhibiting phospholipid and alkyl lipid metabolism [154], (b) causing programmed cell death by damaging mitochondria (depolarization) and obstructing cytochrome c oxidase [155], and (c) increasing nitric oxide synthetase 2 (iNOS2) expression in host macrophages, which produces nitric oxide (NO), which is toxic to parasite survival [154].
Amphotericin B: Also known as Amp B, this antifungal antibiotic exhibits a high affinity for the fungal membrane ergosterol and a low affinity for the host membrane cholesterol [156]. Amp B was identified in 1960 as a promising anti-leishmanial candidate due to its groundbreaking in vitro activity [157]. Following resistance to antimonials in India, Amp B became the first medication of choice to treat VL. By changing the membrane fluidity and creating macropores and micropores that leak essential components into the cells, it binds 24-substituted sterol (ergosterol) to the biosynthesis pathway and causes cell death [10]. Other processes that prevent the parasite from surviving include auto-oxidation and reactive oxygen species (ROS) [158].
Paromomycin: Paromomycin is an antibiotic isolated from Streptomyces rimosus and has a broad range of clinical applicability. It is very helpful against many Gram-positive and Gram-negative bacteria, as well as infections like giardiasis, amoebiasis, and cryptosporidiosis [159]. It is a member of the aminoglycoside class. Its qualities as an anti-leishmanial agent were identified in the 1960s, and it has since been demonstrated to be a successful treatment for both CL and VL [160]. As a result of binding to its 30S smaller subunit, paromomycin hinders protein synthesis machinery. Consequently, the ribosome cannot be recycled due to this binding, which further halts protein synthesis. The membrane potential can also be disrupted by lipid metabolism and membrane fluidity changes [161].
Pentamidine: Pentamidine has been used as a second-line treatment for VL, CL, and DCL for over 40 years. Despite its reintroduction in the 1990s after clinical trials for New World CL [162], pentamidine is barely used as an anti-leishmanial drug. Pentamidine’s anti-leishmanial mechanisms of action remain unclear. However, they may affect the mitochondrial inner membrane potential, inhibit polyamine biosynthesis, and bind to minor grooves in DNA. Clones of L. donovani and L. amazonensis that are resistant to pentamidine exhibit increased efflux and reduced uptake by 18 and 75 folds, respectively [163]. Additional information suggests that pentamidine accumulation in the leishmania mitochondria is significant. However, specific transporters for pentamidine uptake have been identified and may play a role in resistance. In comparison to resistant cells, wild-type promastigotes accumulate more pentamidine in the mitochondria [163,164].
3.2.2. Chagas Disease
Nifurtimox (NFX) and Benznidazole (BZN) are the two main drugs used to treat Chagas disease, and both were developed more than 50 years ago (Figure 5). The treatment is lengthy (60–90 days) and ineffective for chronic patients, in addition to having a toxic effect and the potential for evolving resistance [165,166,167]. Although the exact mechanism underlying Benznidazole’s activity is unknown, it is activated by NADH-dependent trypanosomal reductases and produces reductive metabolites that are thought to have several negative consequences, including DNA damage and protein synthesis inhibition [168]. The FDA approved benznidazole for the treatment of Chagas disease in children aged 2 to 12 years old in 2017. It was the first medicine licensed in the United States for Chagas disease. A new drug that is both safe and effective for Chagas disease’s acute and chronic stages is required. However, several factors impediment the development of new candidate drugs, including the lack of biomarkers for the two stages of the disease and for evaluating treatment success, as well as the genetic diversity of T. cruzi strains [169,170].
Nifurtimox and Benznidazole: Nifurtimox and benznidazole are the current drugs used to treat Chagas disease. Benznidazole’s mechanism of action is a prodrug that is metabolized by the parasite to form free radicals that cause oxidative damage to the parasite’s macromolecules, ultimately leading to death. These free radicals are thought to harm the parasite’s DNA, proteins, and lipids [171]. Nifurtimox’s mechanism of action is unknown, but it is thought to be related to its metabolism via partial reduction to chemically reactive radicals that produce oxygen radicals such as superoxide, hydroperoxide, and hydroxyl. These radicals cause oxidative stress in the parasite, which ultimately leads to death [172]. Table 2 describes the summarized mechanism of action.
Current treatments for leishmaniasis and Chagas disease have several problems. Most of them have side effects, treatment failures, and relapses caused by drug resistance. These anti-trypanosomal and anti-leishmanial medications also have drawbacks that exclude their use in specific population cohorts or under specific conditions. For example, several of them are teratogenic and inappropriate for pregnant women and newborns. Another concern is the poor oral bioavailability of most of these drugs [173]. Chagas disease and leishmaniasis treatments also differ depending on the endemic location. Table 2 summarizes the WHO-recommended treatment regimens for major Chagas disease and leishmaniasis endemic sites. Treatment alternatives are generally insufficient, and new medications are desperately needed. Most drugs used to treat Chagas disease and leishmaniasis are on the 19th edition of the WHO Model List of Essential Medicines, including pentavalent antimonials (SbV), miltefosine, amphotericin B deoxycolate or formulated in liposomal formulations, paromomycin, and pentamidine (Figure 4 and Figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Current pharmacological treatments against Chagas disease and leishmaniasis.
| Disease | Drug | Route | Mechanism of Action | Disadvantages | Toxicity | References |
|---|---|---|---|---|---|---|
| Leishmaniasis | Miltefosine | Oral 5–100 mg/kg/day for 28 days | An anti-apoptotic agent targeting lipid metabolism, mitochondria, and immune function | Need allometric administration in children | Gastrointestinal complications, teratogenic | [174,175,176] |
| Leishmaniasis | Paromomycin | Parenteral (im) 15 mg/kg/day for 21 days | There is little knowledge about this. Several studies show that cationic paromomycin binds negatively charged glycocalyxes and lipophosphoglycans found on the surfaces of leishmania promastigotes, a major component of leishmania promastigotes | Poor results against African VL as monotherapy | Pain in the injection site, hepatotoxicity | [177,178,179] |
| Leishmaniasis | Pentamidine | Slow infusion (iv) 4 mg/kg monthly for 12 months | Not known; drug entry through polyamine/arginine transporters | Multiple adverse effects | Insulin-dependent diabetes, myocarditis, nephrotoxicity | [180] |
| Leishmaniasis | SbV—paromomycin combination | Parenteral (im) SbV 20 mg/kg/day + paromomycin for 17 days | Currently, there are two models of Sb(V) activity: the prodrug model of conversion to toxic Sb(III) and the intrinsic Sb(V) activity based on complex formation with ribose/inhibition of type I DNA topoisomerase. | Require hospitalization | Problems regarding SbV administration | [174,181] |
| Leishmaniasis | Amphotericine B deoxycholate | Slow infusion (iv) 1 mg/kg/day for 30 days | Channel/pore formation on interaction with membrane sterol | Require hospitalization | Nephrotoxicity | [182,183] |
| Leishmaniasis | SbV—based drugs | Parenteral (im) 20 mg/kg/day for 28–30 days | Prodrug model conversion of Sb(V) to toxic Sb(III) and intrinsic Sb(V) activity: by inhibiting type I DNA topoisomerase/complex formation with ribose | Drug resistance in Bihar (India), PKDL | Pain in the injection site, cardiotoxicity, pancreatitis | [184,185] |
| Leishmaniasis | AmBisome | Slow infusion (iv) 10 mg/kg single dose | Channel/pore formation on interaction with membrane sterol | Costly, chemically unstable | Fever during infusion, back pain, nephrotoxicity | [186,187] |
| Chagas disease | Benznidazole (BZL) | Given for 60 days on daily basis at 5–7 mg/kg, and 10 mg/kg for adults and children, respectively | Inhibits the synthesis of DNA, RNA, and proteins within the T. cruzi parasite | Low solubility, toxic, and several side effects | Low bioavailability and drug effectiveness, chronic effects | [86,188,189,190] |
| Chagas disease | Nifurtimox (NFX) | 8–10 mg/kg daily in three divided doses for adults, and 15–20 mg/kg daily in four divided doses for children during 60 to 90 days | Metabolism via partial reduction to chemically reactive radicals that cause production of toxic reduced products of oxygen | Toxic and have side effect, causes gastrointestinal, maladies (nausea, vomiting, abdominal pain) effects | Have higher toxicity and adverse effect than BZL, and it affects the pancreases and heart via increasing of oxidative stress | [191,192,193] |
Abbreviations: im—intramuscular, iv—intravenous.
The current drugs for leishmaniasis and Chagas disease treatment have limitations and challenges. Drugs currently available to treat Chagas disease and leishmaniasis differ in their mode of action, their efficacy, their cost, and their targets [194]. Paromomycin (injectable, long treatment, region-dependent efficacy), miltefosine (cost, teratogenicity, long treatment), and liposomal amphotericin B (cost, hospitalization, region-dependent efficacy) are three examples of drugs. There are only a few validated drug targets known against these parasitic diseases, and many drugs used to treat them have serious side effects and limited efficacy [195]. A severe lack of drug targets that have been rigorously validated genetically and chemically has hampered the development of better and safer drugs to treat these diseases [196,197]. The mode of action of some drugs used to treat these diseases is known [197]. Figure 6 depicts the mode of action, biochemical characterization, and potency correlation of the current drugs reported in the literature to treat Chagas disease and leishmaniasis.
4. Novel Strategies for Leishmaniasis and Chagas Disease Treatments
There is an urgent need to discover and develop compounds effective against leishmaniasis and Chagas disease, which demonstrate high bioactivity, low toxicity, and an acceptable cost and administration profile [16,198]. In the past two decades, numerous assays have been designed for use in screening programs targeting the various lifecycle stages of Leishmania and Trypanosoma parasites. As soon as active compounds are developed against the intracellular amastigote form of the parasite, they are tested in vivo, following a pharmacokinetic and pharmacodynamic assessment [140]. Barriers to the development of new drugs include the lack of well-characterized and validated targets, the absence of diagnostic biomarkers resulting in diagnostic failures, the variety of parasite strains, issues with the standardization of methodologies, such as various in vitro assays, different host cell culture lines, and issues with the translation process [141]. Further, the treatment of these deadly diseases is still challenging due to limited drug regimens, resistance, toxicities, co-infection cases, and low investments in new drug discovery/development [199,200]. Various strategies must be explored and implemented to combat these NTDs and overcome conventional drugs’ drawbacks. The following section summarizes recent developments and upcoming insights into traditional anti-trypanosomal and anti-leishmanial therapies.
4.1. Host-Directed Therapy (HDT)
Host-directed therapy (HDT) aims to achieve a clinical cure by manipulating the host immune system rather than using drugs to treat the parasite [201]. HDT uses various molecules (e.g., repurposed drugs, immuno-modulators, synthetic nucleic acids, cytokines, cellular therapy, and micronutrients) that can boost the immune function of the host body or control the inflammatory response. Both have a long-lasting healing effect and lower mortality and morbidity [202]. A better understanding of the immune response to Leishmania has revealed the crucial fact that infection depends on the successful downregulation of the T-helper cell type 1 (Th1) immune response and the development of the Th2 responses [203]. Since the immune response profile in Chagas disease is similar to that in Leishmania [204], host-directed therapies relevant to Leishmania might also benefit patients with Trypanosome infection [204,205].
For example, simvastatin inhibits the pathway that produces cholesterol and causes macrophages to mature their phagosomes, increasing parasite clearance [206]. Similarly, lovastatin, which is chemically similar to simvastatin, is essential for lowering parasite internalization and infectivity in the J774A.1 macrophage cell line due to cholesterol depletion [207]. Eugenol (Syzygium aromaticum) and its derivatives have been hailed as promising treatments for inducing immunostimulatory responses that benefit the host. Eugenol derivatives inhibit leishmania promastigotes and amastigotes [208]. In conclusion, immuno-modulatory compounds may prove valuable candidates for treating leishmaniasis and Chagas disease. Some drugs/compounds identified through HDT are listed in Table 3.
4.2. Multi-Drug or Combination Therapy
The primary goals of the combinational or multi-drug approach are to shorten the treatment period, lower the relapse rate, reduce dosage, avoid fatal side effects, and combat drug resistance problems [174]. HIV, malaria, and tuberculosis have all been successfully treated using this approach [219,220]. Itraconazole, ezetimibe, and miltefosine were some of the drugs used in a multi-drug therapy approach to evaluate anti-leishmanial efficacy against the VL model of infection in BALB/c mice [203]. Due to its capacity to maximize the efficacy of currently available medications while lowering dosage requirements and treatment times, the combination therapy approach has recently seen significant use in the treatment of leishmaniasis and Chagas disease. A wide range of trustworthy combinations to treat leishmaniasis is possible. In the search for the most effective leishmaniasis treatment regimen, novel drug combinations can now be further investigated.
4.3. Drug Repurposing
Drug repurposing (aka reprofiling, drug repositioning, or re-tasking) is an approach that identifies novel applications for clinically approved drugs originally developed for another medical treatment [221]. This method is associated with a shorter drug development timeline and lower investment costs because the drugs have already undergone clinical studies [222]. Numerous medications have been repurposed for leishmaniasis and have shown promising results, such as Amp B, miltefosine, and azoles (fluconazole, itraconazole, and posaconazole) [223]. Numerous antidepressants, including Ketanserin, imipramine, clomipramine, nitroimipramine, sertraline, etc., have shown promising results when used as repurposed anti-leishmanial medications [223,224]. A recent study using transmission electron microscopy and metabolomics platforms identified the mechanism of sertraline’s leishmanicidal effect on Leishmania parasites [225]. Some of the drugs/compounds identified through drug repurposing are presented in Table 4.
4.4. Promising Natural Products
The Drug Discovery Research Program and the Tropical Diseases Program of the TDR (Tropical Disease Research) WHO intend to make plant pharmacology a priority. An article with a complete list of plants and natural products that demonstrated anti-leishmanial and anti-trypanosomal activities was published in 2015 [234]. Several studies have been conducted since then that demonstrate the pursuit of novel products derived from microbial or marine sources. For example, a glycoprotein isolated from the sponge Pachymatisma johnstonii showed strong in vitro activity against L. donovani, L. braziliensis, and L. Mexicana [235]. Essential oil’s anti-leishmanial activity has also been studied [236]. Advanced research has also evaluated additional potential substances isolated from natural sources that showed anti-leishmanial and anti-trypanosomal activities [196].
Valli et al. report on Cytochalasine A-D-based compounds that are similar to benznidazole or miltefosine in treating intracellular T. cruzi and leishmania that are comparable to the standard drugs of benznidazole and miltefosine [237]. Several recent studies concluded that natural products have better or comparable potency to conventional drugs in treating leishmaniasis and Chagas disease [238,239]. For example, Cortes et al. list natural products extracted from Old World flora that are warranted for leishmanial treatments [240]. Another study demonstrated that natural products could serve as effective chemotherapeutic agents against parasites by targeting mitochondrial homeostasis. The research revealed that these natural compounds can induce parasite death by impacting their mitochondrial function [241]. In conclusion, natural products have a bright future in drug design and research.
4.5. Nanotechnology
Drug delivery and drug design can use nanomedicine, a branch of science that deals with particles at the nanoscale [242]. Low permeability, insolubility, painful injections, prolonged hospitalization, and unfavorable side effects are just a few of the limitations of the conventional delivery system. In the search for the most effective leishmaniasis treatment, researchers have also used nanoparticles (NPs). NPs are employed in drug delivery due to their biocompatibility, improved drug solubility, on-target drug delivery to the target organ, immuno-compatibility, and increased potential for a variety of administration routes [243]. Target-mediated drug delivery systems use metallic, inorganic, organic, and polymeric nanomaterials, such as carbon nanotubes, dendrimers, liposomes, and micelles [244]. Since phagocytic cells, particularly macrophages, are the target organs in leishmaniasis, liposomal derivatives and polymeric NPs are used for drug delivery. Higher ROS production, an increase in immuno-modulatory response, DNA damage, disruption of mitochondrial membrane potential and electron transport chain, and inhibition of trypanothione reductase enzyme vital for the Leishmania parasite’s anti-oxidation process are just a few of the mechanisms that NP-encapsulated drugs/molecules use to kill Leishmania parasites [245]. Although there are countless benefits, nanoscience’s use in practical life is limited. This is because it can sometimes have a significant adverse effect on the body due to its biological sensitivity [246]. Preparation and manipulation require specialized and engineered products, making it a cost-intensive task unsuitable for NTDs like leishmaniasis [247]. Some of the drugs/compounds identified through nanotechnology are listed in Table 5.
4.6. Nano Vaccines
As a novel approach to vaccination, nanovaccines cause both humoral and cell-mediated immune responses, providing better treatment for several diseases, including leishmaniasis [255]. A chitosan nanoparticle-loaded recombinant superoxide dismutase (SODB1) vaccine was developed in 2011 using the ionotropic gelation method and evaluated on BALB/c mice. In both the single and triple doses of SODB1 nanoparticles, IgG2a and IgG2a/IgG1 were significantly higher than in the other groups [256]. A second study examined the efficacy of chimeric peptides containing HLA-restricted epitopes from three immunogenic L. infantum proteins in poly (lactic coglycolic acid) acid nanoparticles with or without monophosphoryl lipid A (MPLA) or surface modification. The nanovaccine activated non-producing CD8+ T cells specific for peptides and induced dendritic cell maturation [257]. Additional studies used synthetic peptide-based nanovaccines along with an MPLA adjuvant co-encapsulated in PLGA nanoparticles. The results demonstrated a strong spleen lymphoproliferative response and high levels of IL-2, interferon gamma (IFN-γ), and tumor necrosis factor α (TNF-α) versus low IL-4 and IL-10 secretion [258,259]. A 2019 study developed a process to prepare lipidic NPs loaded with plasmid pVAX1-NH36 for application as a leishmaniasis nanovaccine [236].
Developing novel therapeutics and substitutes for existing ones is one approach to treating leishmaniasis and Chagas disease [195]. As a means of treating these illnesses, researchers are creating innovative therapeutics and screening techniques to find new compounds/chemical diversity [195,260]. We described above some of the new treatment approaches for Chagas disease and leishmaniasis. As possible hits and leads, a large variety of compounds from several families have been found; some are even undergoing clinical testing [10]. Table 6 lists several candidate drugs that are being considered, including inhibitors that affect the metabolism of thiols, sterols, glycolytic processes, folate, and trypanothione.
5. Peptide Targeting for Leishmaniasis and Chagas Disease Therapies
5.1. Peptide Therapies
Peptides are short chains of amino acids that play critical roles in human physiology. They were first used in medicine in the first half of the 20th century. Therapeutic peptides are exceptional pharmaceutical tools with a molecular weight of 500–5000 Da [270]. As technology advances, peptide drug discovery has become established. This includes drug design, peptide drug discovery, peptide synthesis, structural modification, and the association between these developments and peptide bioactivities [271,272]. Peptides have unique intrinsic characteristics which are produced and modified through chemical and biochemical methods associated with novel design and delivery strategies. Currently, in addition to many preclinical studies, more than 170 peptides are actively in clinical development [270,273] and sales total more than USD 70 billion [274]. Compared with biologics, therapeutic peptides show less immunogenicity and have lower production costs [275,276]. Compared to small molecules, peptides typically demonstrate higher potency and selectivity for their targets [277,278].
Therapeutic peptides have two basic limitations: low membrane permeability and poor in vivo stability. Yet, peptides as drugs have many advantages including high potency, high selectivity, and low toxicity. Thus, efforts using several models confirmed the effects of several peptides on various parasites (Table 7) [272,279,280].
5.2. Anti-Microbial Peptides
An anti-microbial peptide (AMP) is a small cationic molecule, amphipathic in structure, which has a variety of activities against viruses, bacteria, fungi, and parasites [289,290]. AMPs are more effective than conventional antibiotics, with a low bactericidal concentration and a quick ability to kill germs. They can even effectively combat bacteria strains resistant to conventional antibiotics [291]. Furthermore, AMPs can also overcome drug resistance in bacteria. Some of them have biological activities, such as anti-parasitic, anti-virus, and anti-fungal properties [291,292]. Current research efforts are focused on developing AMPs for therapeutic applications against NTDs, including leprosy, trachoma, African trypanosomiasis, Chagas disease, and leishmaniasis [292,293]. AMPs are effective against a variety of NTDs, including African trypanosomes, leishmaniasis, and Chagas disease. Research into the development of new anti-microbial agents has been driven by the increase in resistance to traditional antibiotics and emerging infectious diseases. Due to their direct anti-microbial killing activity and significant role in innate immunity, AMPs represent a promising alternative to existing antibiotics in the treatment and prevention of microbial infections [292]. To date, more than 2000 AMPs have been discovered, and many exhibit broad-spectrum anti-bacterial, anti-viral, and anti-parasitic activity [294]. Research on the potential application of AMPs’ structural and natural analogs in the fight against NTDs is ongoing [292].
The primary mechanism by Ih AIPs inhibit parasItic infection is by binding to and rupturing the plasma membrane of the parasites [294,295]. Most of these peptides are cationic–amphipathic, exhibiting two main mechanisms of action: direct lysis and modulating the immune system [289]. In addition to depolarizing membranes, disrupting plasma membrane permeability, and causing programmed cell death, these peptides also have major microbicidal effects. As a result, AMPs play a crucial role in the production of cytokines which modulate immune responses in the host [296,297]. Researchers working on parasites pay a lot of attention to AMPs, which are also called host-defense peptides [293]. Murine cathelicidin or cathelin-derived AMP was evaluated against L. major infection. Compared to control cells, J774A.1 cells expressed more cathelin-derived AMP in vitro. Furthermore, cathelin-derived AMP expression was upregulated at dermal inoculation sites in L. major-infected BALB/c mice, unlike the control group [203,297]. Additionally, protein assays showed that cathelin-related AMP levels increased in infected macrophages and mice models, suggesting that the immune response against L. major may be mediated by cathelin-related AMPs. A study conducted by Zahedifard et al. evaluated the leishmanicidal potential of Brevinin 2R (belonging to the class of defensins) alone or in combination with lauric acid against L. major parasites and found that mice treated with Brevinin 2R exhibited decreased parasite load in the lymph nodes [203,298].
Since AMPs have excellent leishmanicidal and immunomodulatory properties, they can serve as leads during drug discovery pipelines and vaccine design for leishmaniasis and Chagas disease. In general, AMPs receive more attention due to different factors such as antibiotic-resistant microorganisms [299]. While AMP mechanisms remain unclear, many AMPs cause swelling in the promastigote membrane, which leads to loss of integrity and substantial cell death [208]. The development of AMPs for therapeutic applications against NTDs faces several challenges, such as natural AMP lability, susceptibility to protease, pH changes, and potential toxicity [289,294]. The advantages of AMPs over conventional antibiotics include a slower emergence of resistance, broad-spectrum activity, and efficacy against drug-resistant Gram-positive organisms [291,300]. Despite these obstacles, AMPs have demonstrated promise for treating drug-resistant bacteria and infectious diseases, including non-therapeutic diseases like leishmaniasis and Chagas disease. Even though there are challenges to overcome, there are potential advantages in treating NTDs and AMPs represent a promising field for research and development.
5.2.1. Anti-Microbial Peptides against Chagas Disease and Leishmaniasis
K777, a vinyl sulfone cysteine protease inhibitor, is considered a highly powerful and well-recognized cysteine peptidase inhibitor. It inhibits cruzain (aka Cruzipain), a key protease required for T. cruzi survival. It has not promoted a parasitological cure but did significantly reduce parasite-induced heart damage in vivo [301]. Over the past few years, several AMPs have been evaluated for their effects on T. cruzi, and apidaecin, magainin II, melittin, and cecropin A have been identified as potential candidates for Chagas disease as they kill T. cruzi in low concentrations [302].
AMPs are an essential protection mechanism and part of vertebrates’ native immunity. These peptides damage the protozoan’s cell membrane, affecting membrane integrity and producing lethal pores [303]. Over 2,000 AMPs have been identified from a variety of organisms, including bacteria, insects, plants, amphibians, birds, reptiles, and mammals, including humans. Some of these AMPs demonstrate leishmanicidal activity, such as halictine-2 from the poison of eusocial honey-bees [304]. Attacin, cecropin, and defensins from lutzomyia longipalpis respond to Leishmania infection [305], and Dragomide E., a linear lipopeptide isolated from the cyanobacteria Lyngbya majuscula, demonstrates anti-leishmanial activity against L. donovani promastigotes. The LZ1 peptide, derived from Snake cathelicidin, is an artificially designed and synthesized active polypeptide demonstrating high anti-microbial activity. It inhibits ATP production in parasite-infected erythrocytes [306]. Finally, Phylloseptin-1, a cationic peptide from the skin secretion of Phyllomedusa azurea, demonstrates high anti-parasitic activity and prevents cross-resistance because of its distinctive chemical structure [307].
The pharmaceutical industry recognizes that peptide-based drugs are a significant class of therapeutic agents. Anti-protozoal therapeutics can be derived from insects, bacteria, marine organisms, and amphibians [296]. In terms of parasite propagation, there are two stages in which the parasites can be investigated: the diagnostic stage (amastigotes multiply in cells) and the infectious stage (the promastigotes are injected into the blood) [157]. Currently, there are no specific details or clear evidence regarding the stages of clinical trials of AMPs against leishmaniasis and Chagas disease. Additionally, the pipelines for drugs to treat Chagas disease, leishmaniasis of the skin, mucocutaneous leishmaniasis, and PKDL are poor. New drugs are urgently needed to treat these diseases [308]. To achieve the WHO road map for neglected tropical diseases, new drugs are needed for leishmaniasis and Chagas disease [309]. In the case of tropical diseases, there are some studies of clinical trials investigating anti-microbial activities against bacteria [310,311]. The next figure describes some of the AMPs that have been identified and their functions against the different developmental forms of parasites (Figure 7) [303].
In general, several AMPs have shown promising activity against Chagas disease and leishmaniasis. Oxaborole DNDI-6148 has been nominated as a potential candidate for Chagas disease and is currently undergoing phase I clinical trials [139]. However, there is little progress in the pipeline for Chagas disease treatment. Other reports indicate that the acylated synthetic anti-microbial peptide Oct-CA (1-7)M (2-9) was used to treat leishmaniasis in dogs, but it is unclear whether it is currently in clinical trials [289]. Overall, several AMPs have demonstrated anti-trypanosomal and anti-leishmanial activity, and they are promising candidates for clinical trials.
5.2.2. Structural Analysis of Selected Anti-Microbial Peptide Profiles against Leishmaniasis and Chagas Disease
AMPs are significant and essential defense mechanisms and highly active against various pathogens. AMPs are a component of the immune systems of vertebrates and invertebrates [303]. Most natural AMPs are short (10 to 50 amino acids) and have positive net charges (+2 to +11), with a significant portion of hydrophobic residues [312]. Due to their anti-parasitic properties, AMPs are used as pharmaceutical leads [313]. Positively charged residues on the polar face and net charge become essential for both anti-microbial and hemolytic activities [314,315,316,317]. Table 8 below summarizes the net charge and activities of selected AMPs for leishmaniasis and Chagas disease.
AMPs’ amino acid composition determines their charging properties and selective actions [329,330]. The cationic charge in their structures has a strong correlation with anti-microbial activity, and it is responsible for the initial electrostatic interaction between peptides and the anionic microbial surface through electrostatic interactions. The hydrophobic residues enable the AMPs to further insert into the membrane bilayer [331]. Positive-charge peptides are usually highly alkali at the N-terminal ends and rich in basic amino acids, while they are neutral hydrophobicity at their C-terminal ends [332]. Modification of AMPs is intended to increase their stability and efficacy while decreasing their cytotoxicity and untargeted side effects [333]. Peptides with higher net positive charges can be considered anti-microbial potential inhibitors. On the other hand, the hydrophobicity of those selected AMPs usually does not increase or decrease with net positive charges. There is an optimum hydrophobicity window for high anti-microbial activity. The decrease in anti-microbial activities at high peptide hydrophobicity can be explained by the strong peptide self-association, which prevents the peptide from passing through the cell wall in prokaryotic cells [334].
5.3. Protein–Protein Interactions as Drug Targets in Leishmaniasis and Chagas Disease
The human proteome contains approximately 30,000 proteins and significantly more protein–protein interactions (PPIs) that play critical roles in biological processes. Their dysregulation results in the onset and progression of a variety of diseases. PPIs thus represent a treasure trove of disease-modifying drug targets [335]. However, targeting these is difficult when converting drug-like small molecules into therapeutics. When targeting PPIs, it is critical to strike a balance between the interacting proteins to elicit a therapeutic effect while avoiding a significant adverse effect [336,337]. Researchers have reported that PPIs as a drug discovery strategy target a variety of diseases [337]. However, there is limited information about PPIs for leishmaniasis and Chagas disease. The proteins tryparedoxin peroxidase (PDB ID: 3TUE) [338], trypanothione reductase: (PDB ID: 5EBK) [339], and pyruvate kinase (PDB ID: 3PP7) [340] have recently been identified as structural macromolecules that play a variety of roles in the Leishmania parasite [4]. It is worth considering the importance of these proteins in the context of Leishmania, as well as their potential implications for future research into the parasite’s metabolism and pathogenicity. Therefore, PPIs offer the potential for further research into drug discovery for leishmaniasis and Chagas disease.
6. Conclusions and Future Directions
Leishmaniasis and Chagas disease are NTDs and global burdens. As a result of recurrent failures in leishmaniasis and Chagas disease diagnosis, chemotherapy is delayed and, eventually, this results in death. Despite the efforts of many research groups, leishmaniasis and Chagas disease remain without an effective solution. Challenges regarding leishmaniasis and Chagas disease include accurate and timely disease diagnosis, which is crucial in identifying asymptomatic patients and co-infected cases. However, recent diagnostic procedures such as advanced molecular techniques, proteomics, and nano-diagnosis may improve the prognosis of these diseases. Other key challenges are related to currently available treatments, including drug resistance and toxicities. These may be overcome by less traditional approaches, such as multi-drug and host-directed therapies, which have yielded promising results in recent studies. In addition, clinical trials with innovative treatment strategies should be performed to explore the impact of novel approaches on improving treatment efficacy, decreasing side effects, and lowering treatment costs. The review discusses the advancement of novel treatment strategies, featuring peptides as potential therapeutics against NTDs. Since peptides have specific actions and reduced toxicity, they are promising as new drug discovery targets and for development. It is evident that these targeted compounds are becoming increasingly popular within the pharmaceutical industry and hopefully they will greatly improve the health of many people suffering in the world’s poorest countries.
Existing anti-parasitic drugs for leishmaniasis and Chagas disease, in use for decades, are effective but pose significant concerns due to adverse effects and drug resistance. Drugs like pentamidine and amphotericin can lead to hospitalization and face resistance issues. High costs limit amphotericin’s accessibility. Miltefosine, that was originally developed as an anti-cancer drug and later was found to be useful for leishmaniasis, may cause teratogenicity and resistance. Chagas treatments, benznidazole and nifurtimox, exhibit adverse effects. Combination therapy faces limitations due to resistance and toxicities. This has prompted exploration of new strategies like host-directed therapy, nanovaccines, and natural extracts, showing promise with better stability and reduced toxicity. Vaccines against these diseases prove effective. Peptide-based therapies, despite stability and permeability challenges, offer low toxicity and resistance, making them potential candidates for leishmaniasis and Chagas treatment, exhibiting high efficacy at the infection site. Overall, peptides, vaccines, and novel approaches present alternatives with lower resistance, toxicity, and higher efficacy against parasitic infections.
Author Contributions
M.Z., M.K.C.S., H.B. and N.Q. conceptualized and wrote the review and prepared the tables and figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Israel Science Foundation (ISF)-CIHR-IDRC through a research grant (no. 2993/22) awarded to N.Q.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data can be found within the article.
Acknowledgments
We thank those who were involved in the proofreading and editing of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stuart, K.; Brun, R.; Croft, S.; Fairlamb, A.; Gürtler, R.E.; McKerrow, J.; Reed, S.; Tarleton, R. Kinetoplastids: Related protozoan pathogens, different diseases. J. Clin. Investig. 2008, 118, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Naula, C.; Parsons, M.; Mottram, J.C. Protein kinases as drug targets in trypanosomes and Leishmania. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2005, 1754, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Abadías-Granado, I.; Diago, A.; Cerro, P.; Palma-Ruiz, A.; Gilaberte, Y. Cutaneous and mucocutaneous leishmaniasis. Actas Dermo-Sifiliográficas 2021, 112, 601–618. [Google Scholar] [CrossRef]
- Singh, R.; Kashif, M.; Srivastava, P.; Manna, P.P. Recent Advances in Chemotherapeutics for Leishmaniasis: Importance of the Cellular Biochemistry of the Parasite and Its Molecular Interaction with the Host. Pathogens 2023, 12, 706. [Google Scholar] [CrossRef] [PubMed]
- Assis, T.S.M.d.; Rosa, D.C.P.; Teixeira, E.d.M.; Cota, G.; Azeredo-da-Silva, A.L.F.; Werneck, G.; Rabello, A. The direct costs of treating human visceral leishmaniasis in Brazil. Rev. Soc. Bras. Med. Trop. 2017, 50, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.S.; Muthoga, C.; Meya, D.B.; Tugume, L.; Williams, D.; Rajasingham, R.; Boulware, D.R.; Mwandumba, H.C.; Moyo, M.; Dziwani, E.N. Cost-effectiveness of single, high-dose, liposomal amphotericin regimen for HIV-associated cryptococcal meningitis in five countries in sub-Saharan Africa: An economic analysis of the AMBITION-cm trial. Lancet Glob. Health 2022, 10, e1845–e1854. [Google Scholar] [CrossRef] [PubMed]
- Boettiger, D.C.; Lin, T.K. Cost-effectiveness of liposomal amphotericin B for HIV-associated cryptococcal meningitis. Lancet Glob. Health 2022, 10, e1705–e1706. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Arana, B.I. Appraisal of Leishmaniasis Chemotherapy, Current Status and Pipeline Strategies Chapter 1 Leishmaniasis, Impact and Therapeutic Needs 3. Drug Discov. Leishmaniasis 2018, 10, 9781788010177-00001. [Google Scholar]
- Rajao, M.A.; Furtado, C.; Alves, C.L.; Passos-Silva, D.G.; De Moura, M.B.; Schamber-Reis, B.L.; Kunrath-Lima, M.; Zuma, A.A.; Vieira-da-Rocha, J.P.; Garcia, J.B.F. Unveiling benznidazole’s mechanism of action through overexpression of DNA repair proteins in Trypanosoma cruzi. Environ. Mol. Mutagen. 2014, 55, 309–321. [Google Scholar] [CrossRef]
- Sangshetti, J.N.; Khan, F.A.K.; Kulkarni, A.A.; Arote, R.; Patil, R.H. Antileishmanial drug discovery: Comprehensive review of the last 10 years. RSC Adv. 2015, 5, 32376–32415. [Google Scholar] [CrossRef]
- Chappuis, F.; Sundar, S.; Hailu, A.; Ghalib, H.; Rijal, S.; Peeling, R.W.; Alvar, J.; Boelaert, M. Visceral leishmaniasis: What are the needs for diagnosis, treatment and control? Nat. Rev. Microbiol. 2007, 5, 873–882. [Google Scholar] [CrossRef]
- Sundberg, S.A. High-throughput and ultra-high-throughput screening: Solution-and cell-based approaches. Curr. Opin. Biotechnol. 2000, 11, 47–53. [Google Scholar] [CrossRef]
- Mayr, L.M.; Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 2009, 9, 580–588. [Google Scholar] [CrossRef]
- Akao, Y.; Canan, S.; Cao, Y.; Condroski, K.; Engkvist, O.; Itono, S.; Kaki, R.; Kimura, C.; Kogej, T.; Nagaoka, K. Collaborative virtual screening to elaborate an imidazo [1, 2-a] pyridine hit series for visceral leishmaniasis. RSC Med. Chem. 2021, 12, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Schaduangrat, N.; Lampa, S.; Simeon, S.; Gleeson, M.P.; Spjuth, O.; Nantasenamat, C. Towards reproducible computational drug discovery. J. Cheminformatics 2020, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, E.; Ioset, J.-R. Phenotypic screening approaches for Chagas disease drug discovery. Expert. Opin. Drug Discov. 2018, 13, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Zahedifard, F.; Rafati, S. Prospects for antimicrobial peptide-based immunotherapy approaches in Leishmania control. Expert. Rev. Anti-Infect. Ther. 2018, 16, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Rafferty, J.; Nagaraj, H.; McCloskey, A.P.; Huwaitat, R.; Porter, S.; Albadr, A.; Laverty, G. Peptide therapeutics and the pharmaceutical industry: Barriers encountered translating from the laboratory to patients. Curr. Med. Chem. 2016, 23, 4231–4259. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Garrido, P.; Cárdenas-Guerra, R.E.; Martínez, I.; Poggio, S.; Rodríguez-Hernández, K.; Rivera-Santiago, L.; Ortega-López, J.; Sánchez-Esquivel, S.; Espinoza, B. Differential activity on trypanosomatid parasites of a novel recombinant defensin type 1 from the insect Triatoma (Meccus) pallidipennis. Insect Biochem. Mol. Biol. 2021, 139, 103673. [Google Scholar] [CrossRef] [PubMed]
- Dostálová, A.; Volf, P. Leishmania development in sand flies: Parasite-vector interactions overview. Parasites Vectors 2012, 5, 276. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Teixeira, D.E.; Benchimol, M.; Rodrigues, J.C.; Crepaldi, P.H.; Pimenta, P.F.; de Souza, W. The cell biology of Leishmania: How to teach using animations. PLoS Pathog. 2013, 9, e1003594. [Google Scholar] [CrossRef]
- Sunter, J.; Gull, K. Shape, form, function and Leishmania pathogenicity: From textbook descriptions to biological understanding. Open Biol. 2017, 7, 170165. [Google Scholar] [CrossRef]
- Saada, E.A.; Kabututu, Z.P.; Lopez, M.; Shimogawa, M.M.; Langousis, G.; Oberholzer, M.; Riestra, A.; Jonsson, Z.O.; Wohlschlegel, J.A.; Hill, K.L. Insect stage-specific receptor adenylate cyclases are localized to distinct subdomains of the Trypanosoma brucei flagellar membrane. Eukaryot. Cell 2014, 13, 1064–1076. [Google Scholar] [CrossRef]
- Secundino, N.; Eger-Mangrich, I.; Braga, E.; Santoro, M.; Pimenta, P. Lutzomyia longipalpis peritrophic matrix: Formation, structure, and chemical composition. J. Med. Entomol. 2005, 42, 928–938. [Google Scholar] [CrossRef]
- Rogers, M.E.; Chance, M.; Bates, P. The role of promastigote secretory gel in the origin and transmission of the infective stage of Leishmania mexicana by the sandfly Lutzomyia longipalpis. Parasitology 2002, 124, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Gossage, S.M.; Rogers, M.E.; Bates, P.A. Two separate growth phases during the development of Leishmania in sand flies: Implications for understanding the life cycle. Int. J. Parasitol. 2003, 33, 1027–1034. [Google Scholar] [CrossRef]
- Bates, P.A. Revising Leishmania’s life cycle. Nat. Microbiol. 2018, 3, 529–530. [Google Scholar] [CrossRef] [PubMed]
- Martín-Escolano, J.; Marín, C.; Rosales, M.J.; Tsaousis, A.D.; Medina-Carmona, E.; Martín-Escolano, R. An updated view of the Trypanosoma cruzi life cycle: Intervention points for an effective treatment. ACS Infect. Dis. 2022, 8, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Onyekwelu, K.C. Life Cycle of Trypanosoma cruzi in the Invertebrate and the Vertebrate Hosts. In Biology of Trypanosoma cruzi; BoD—Books on Demand: Norderstedt, Germany, 2019; pp. 1–19. [Google Scholar]
- Rodríguez-Bejarano, O.H.; Avendaño, C.; Patarroyo, M.A. Mechanisms associated with Trypanosoma cruzi host target cell adhesion, recognition and internalization. Life 2021, 11, 534. [Google Scholar] [CrossRef]
- Teixeira, D.E.; Benchimol, M.; Crepaldi, P.H.; de Souza, W. Interactive multimedia to teach the life cycle of Trypanosoma cruzi, the causative agent of Chagas disease. PLoS Neglected Trop. Dis. 2012, 6, e1749. [Google Scholar] [CrossRef]
- Andreoli, W.; Taniwaki, N.; Mortara, R. Survival of Trypanosoma cruzi metacyclic trypomastigotes within Coxiella burnetii vacuoles: Differentiation and replication within an acidic milieu. Microbes Infect. 2006, 8, 172–182. [Google Scholar] [CrossRef]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Alves, C.R.; Albuquerque-Cunha, J.M.; Mello, C.; Garcia, E.d.S.; Nogueira, N.; Bourguingnon, S.C.; De Souza, W.; Azambuja, P.; Gonzalez, M.S. Trypanosoma cruzi: Attachment to perimicrovillar membrane glycoproteins of Rhodnius prolixus. Exp. Parasitol. 2007, 116, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.S.; Ratcliffe, N.A.; Whitten, M.M.; Gonzalez, M.S.; Azambuja, P. Exploring the role of insect host factors in the dynamics of Trypanosoma cruzi–Rhodnius prolixus interactions. J. Insect Physiol. 2007, 53, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Roldan, J.A.; Votýpka, J.; Bandi, C.; Epis, S.; Modrý, D.; Tichá, L.; Volf, P.; Otranto, D. Leishmania tarentolae: A new frontier in the epidemiology and control of the leishmaniases. Transbound. Emerg. Dis. 2022, 69, e1326–e1337. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.C.; Quintella, L.P.; Schubach, A.d.O.; Miranda, L.d.F.C.; Madeira, M.d.F.; Pimentel, M.I.F.; Vasconcellos, É.d.C.F.e.; Lyra, M.R.; Oliveira, R.d.V.C.d.; Menezes, R.C. Comparison between Colorimetric In Situ Hybridization, Histopathology, and Immunohistochemistry for the Diagnosis of New World Cutaneous Leishmaniasis in Human Skin Samples. Trop. Med. Infect. Dis. 2022, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Hadermann, A.; Heeren, S.; Maes, I.; Dujardin, J.-C.; Domagalska, M.A.; Van den Broeck, F. Genome diversity of Leishmania aethiopica. Front. Cell. Infect. Microbiol. 2023, 13, 406. [Google Scholar] [CrossRef] [PubMed]
- Bezemer, J.M.; Freire-Paspuel, B.P.; Schallig, H.D.; de Vries, H.J.; Calvopiña, M. Leishmania species and clinical characteristics of Pacific and Amazon cutaneous leishmaniasis in Ecuador and determinants of health-seeking delay: A cross-sectional study. BMC Infect. Dis. 2023, 23, 395. [Google Scholar] [CrossRef] [PubMed]
- Preativatanyou, K.; Chinwirunsirisup, K.; Phumee, A.; Khositharattanakool, P.; Sunantaraporn, S.; Depaquit, J.; Siriyasatien, P. Species diversity of phlebotomine sand flies and sympatric occurrence of Leishmania (Mundinia) martiniquensis, Leishmania (Leishmania) donovani complex, and Trypanosoma spp. in the visceral leishmaniasis focus of southern Thailand. Acta Trop. 2023, 244, 106949. [Google Scholar] [CrossRef] [PubMed]
- Dinç, M.; Yalçın, T.; Çavuş, İ.; Özbilgin, A. Comparative proteomic analysis of Leishmania parasites isolated from visceral and cutaneous leishmaniasis patients. Parasitology 2022, 149, 298–305. [Google Scholar] [CrossRef]
- Gramiccia, M.; Gradoni, L. The current status of zoonotic leishmaniases and approaches to disease control. Int. J. Parasitol. 2005, 35, 1169–1180. [Google Scholar] [CrossRef]
- Schriefer, A.; Guimarães, L.H.; Machado, P.R.; Lessa, M.; Lessa, H.A.; Lago, E.; Ritt, G.; Góes-Neto, A.; Schriefer, A.L.; Riley, L.W. Geographic clustering of leishmaniasis in northeastern Brazil. Emerg. Infect. Dis. 2009, 15, 871. [Google Scholar] [CrossRef]
- Montalvo Alvarez, A.M.; Nodarse, J.F.; Goodridge, I.M.; Fidalgo, L.M.; Marin, M.; Van Der Auwera, G.; Dujardin, J.-C.; Velez Bernal, I.D.; Muskus, C. Differentiation of Leishmania (Viannia) panamensis and Leishmania (V.) guyanensis using Bcc I for hsp 70 PCR-RFLP. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 364–367. [Google Scholar] [CrossRef]
- Ducharme, O.; Simon, S.; Ginouves, M.; Prévot, G.; Couppie, P.; Demar, M.; Blaizot, R. Leishmania naiffi and lainsoni in French Guiana: Clinical features and phylogenetic variability. PLoS Neglected Trop. Dis. 2020, 14, e0008380. [Google Scholar] [CrossRef]
- Espada, C.R.; Ferreira, B.A.; Ortiz, P.A.; Uliana, S.R.; Coelho, A.C. Full nucleotide sequencing of ribosomal DNA internal transcribed spacer of Leishmania species causing cutaneous leishmaniasis in Brazil and its potential for species typing. Acta Trop. 2021, 223, 106093. [Google Scholar] [CrossRef]
- Espinoza-Morales, D.; Rodríguez, A.L.; Silva-Caso, W.; Suarez-Ognio, L.; Pons, M.J.; del Valle Mendoza, J. An atypical case of disseminated cutaneous leishmaniasis due to Leishmania peruviana in the valleys of Ancash-Peru. Asian Pac. J. Trop. Med. 2017, 10, 1101–1103. [Google Scholar] [CrossRef] [PubMed]
- Passero, L.F.D.; Carvalho, A.K.; Bordon, M.L.; Bonfim-Melo, A.; Carvalho, K.; Kallás, E.G.; Santos, B.B.; Toyama, M.H.; Paes-Leme, A.; Corbett, C.E. Proteins of Leishmania (Viannia) shawi confer protection associated with Th1 immune response and memory generation. Parasites Vectors 2012, 5, 64. [Google Scholar] [CrossRef] [PubMed]
- Sukhumavasi, W.; Kaewamatawong, T.; Somboonpoonpol, N.; Jiratanh, M.; Wattanamethanont, J.; Kaewthamasorn, M.; Leelayoova, S.; Tiwananthagorn, S. Liver-and spleen-specific immune responses in experimental leishmania martiniquensis infection in BALB/c mice. Front. Vet. Sci. 2021, 8, 794024. [Google Scholar] [CrossRef] [PubMed]
- Supsrisunjai, C.; Kootiratrakarn, T.; Puangpet, P.; Bunnag, T.; Chaowalit, P.; Wessagowit, V. Case report: Disseminated autochthonous dermal leishmaniasis caused by Leishmania siamensis (PCM2 Trang) in a patient from central Thailand infected with human immunodeficiency virus. Am. J. Trop. Med. Hyg. 2017, 96, 1160. [Google Scholar] [PubMed]
- Ramírez, J.D.; Hernández, C.; León, C.M.; Ayala, M.S.; Flórez, C.; González, C. Taxonomy, diversity, temporal and geographical distribution of Cutaneous Leishmaniasis in Colombia: A retrospective study. Sci. Rep. 2016, 6, 28266. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.R.; Simarro, P.P.; Diarra, A.; Jannin, J.G. Epidemiology of human African trypanosomiasis. Clin. Epidemiol. 2014, 6, 257–275. [Google Scholar]
- Schmunis, G.A.; Yadon, Z.E. Chagas disease: A Latin American health problem becoming a world health problem. Acta Trop. 2010, 115, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An overview on target-based drug design against kinetoplastid protozoan infections: Human African trypanosomiasis, Chagas disease and leishmaniases. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef] [PubMed]
- Bilgic-Temel, A.; Murrell, D.F.; Uzun, S. Cutaneous leishmaniasis: A neglected disfiguring disease for women. Int. J. Women’s Dermatol. 2019, 5, 158–165. [Google Scholar] [CrossRef]
- Kaluarachchi, T.J.; Campbell, P.M.; Wickremasinghe, R.; Ranasinghe, S.; Yasewardene, S.; De Silva, H.; McBain, A.J.; Weerasekera, M. Possible clinical implications and future directions of managing bacterial biofilms in cutaneous leishmaniasis wounds. Trop. Med. Health 2022, 50, 58. [Google Scholar] [CrossRef]
- Martínez, D.Y.; Verdonck, K.; Kaye, P.M.; Adaui, V.; Polman, K.; Llanos-Cuentas, A.; Dujardin, J.-C.; Boelaert, M. Tegumentary leishmaniasis and coinfections other than HIV. PLoS Neglected Trop. Dis. 2018, 12, e0006125. [Google Scholar] [CrossRef]
- Meireles, C.B.; Maia, L.C.; Soares, G.C.; Teodoro, I.P.P.; Gadelha, M.d.S.V.; da Silva, C.G.L.; de Lima, M.A.P. Atypical presentations of cutaneous leishmaniasis: A systematic review. Acta Trop. 2017, 172, 240–254. [Google Scholar] [CrossRef]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef]
- Sinha, S.; Fernández, G.; Kapila, R.; Lambert, W.C.; Schwartz, R.A. Diffuse cutaneous leishmaniasis associated with the immune reconstitution inflammatory syndrome. Int. J. Dermatol. 2008, 47, 1263–1270. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Gomez, E.L.; Kato, H.; Martini, L.R.; Velez, L.N.; Uezato, H. Diffuse and disseminated cutaneous leishmaniasis: Clinical cases experienced in Ecuador and a brief review. Trop. Med. Health 2016, 44, 2. [Google Scholar] [CrossRef] [PubMed]
- Goihman-Yahr, M. American mucocutaneous leishmaniasis. Dermatol. Clin. 1994, 12, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Vera-Izaguirre, D.S.; Vega-Memije, E.; Quintanilla-Cedillo, M.R.; Arenas, R. Leishmaniasis. A review. Dermatol. Cosmética Médica Y Quirúrgica 2006, 4, 252–260. [Google Scholar]
- Murray, H.W.; Berman, J.D.; Davies, C.R.; Saravia, N.G. Advances in leishmaniasis. Lancet 2005, 366, 1561–1577. [Google Scholar] [CrossRef] [PubMed]
- CONVIT, J.; REYES, O.; KERDEL, F. Disseminated Anergic American Leishmaniasis: Report of Three Cases of a Type Clinically Resembling Lepromatous Leprosy. AMA Arch. Dermatol. 1957, 76, 213–217. [Google Scholar] [CrossRef]
- Chaudhary, R.G.; Bilimoria, F.E.; Katare, S. Diffuse cutaneous leishmaniasis: Co-infection with human immunodeficiency virus (HIV). Indian J. Dermatol. Venereol. Leprol. 2008, 74, 641. [Google Scholar] [CrossRef]
- David, C.V.; Craft, N. Cutaneous and mucocutaneous leishmaniasis. Dermatol. Ther. 2009, 22, 491–502. [Google Scholar] [CrossRef]
- Garrido-Jareño, M.; Sahuquillo-Torralba, A.; Chouman-Arcas, R.; Castro-Hernández, I.; Molina-Moreno, J.M.; Llavador-Ros, M.; Gómez-Ruiz, M.D.; López-Hontangas, J.L.; Botella-Estrada, R.; Salavert-Lleti, M. Cutaneous and mucocutaneous leishmaniasis: Experience of a Mediterranean hospital. Parasites Vectors 2020, 13, 24. [Google Scholar] [CrossRef]
- Ahluwalia, S.; Lawn, S.D.; Kanagalingam, J.; Grant, H.; Lockwood, D.N. Mucocutaneous leishmaniasis: An imported infection among travellers to central and South America. BMJ 2004, 329, 842–844. [Google Scholar] [CrossRef]
- Marra, F.; Chiappetta, M.C.; Vincenti, V. Ear, nose and throat manifestations of mucocutaneous Leishmaniasis: A literature review. Acta Biomed. 2014, 85, 3–7. [Google Scholar] [PubMed]
- Bern, C. Visceral Leishmaniasis: Clinical Manifestations and Diagnosis. In U: UpToDate, Post TW ur. UpToDate; UpToDate: Waltham, MA, USA, 2021. [Google Scholar]
- Baba, C.S.; Makharia, G.K.; Mathur, P.; Ray, R.; Gupta, S.D.; Samantaray, J. Chronic diarrhea and malabsorption caused by Leishmania donovani. Indian J. Gastroenterol. 2006, 25, 309. [Google Scholar] [PubMed]
- Boukhris, I.; Azzabi, S.; Chérif, E.; Kéchaou, I.; Mahjoub, S.; Kooli, C.; Aoun, K.; Khalfallah, N. Hemophagocytosis and disseminated intravascular coagulation in visceral leishmaniasis in adults: Three new cases. Pan Afr. Med. J. 2015, 22, 96. [Google Scholar] [PubMed]
- Pagliano, P.; Carannante, N.; Rossi, M.; Gramiccia, M.; Gradoni, L.; Faella, F.S.; Gaeta, G.B. Visceral leishmaniasis in pregnancy: A case series and a systematic review of the literature. J. Antimicrob. Chemother. 2005, 55, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Orfanos, G.; Argaw, D.; Mulugeta, A.; Aparicio, P.; Parreño, F.; Bernal, O.; Rubens, D.; Pedraza, J.; Lima, M.A. Natural history of a visceral leishmaniasis outbreak in highland Ethiopia. Am. J. Trop. Med. Hyg. 2009, 81, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Mueller, Y.; Mbulamberi, D.B.; Odermatt, P.; Hoffmann, A.; Loutan, L.; Chappuis, F. Risk factors for in-hospital mortality of visceral leishmaniasis patients in eastern Uganda. Trop. Med. Int. Health 2009, 14, 910–917. [Google Scholar] [CrossRef]
- Zijlstra, E.E. Biomarkers in post-kala-azar dermal leishmaniasis. Front. Cell. Infect. Microbiol. 2019, 9, 228. [Google Scholar] [CrossRef]
- Zijlstra, E.E. The immunology of post-kala-azar dermal leishmaniasis (PKDL). Parasites Vectors 2016, 9, 464. [Google Scholar] [CrossRef]
- Zijlstra, E.; Musa, A.; Khalil, E.; El Hassan, I.; El-Hassan, A. Post-kala-azar dermal leishmaniasis. Lancet Infect. Dis. 2003, 3, 87–98. [Google Scholar] [CrossRef]
- Kumar, P.; Chatterjee, M.; Das, N.K. Post kala-azar dermal leishmaniasis: Clinical features and differential diagnosis. Indian J. Dermatol. 2021, 66, 24. [Google Scholar]
- Suárez, C.; Nolder, D.; García-Mingo, A.; Moore, D.A.; Chiodini, P.L. Diagnosis and clinical management of Chagas disease: An increasing challenge in non-endemic areas. Res. Rep. Trop. Med. 2022, 2022, 25–40. [Google Scholar] [CrossRef]
- Nunes, M.C.P.; Dones, W.; Morillo, C.A.; Encina, J.J.; Ribeiro, A.L.; Council on Chagas Disease of the Interamerican Society of Cardiology. Chagas disease: An overview of clinical and epidemiological aspects. J. Am. Coll. Cardiol. 2013, 62, 767–776. [Google Scholar] [CrossRef]
- Hemmige, V.; Tanowitz, H.; Sethi, A. Trypanosoma cruzi infection: A review with emphasis on cutaneous manifestations. Int. J. Dermatol. 2012, 51, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; Nitz, N.; Guimaro, M.; Gomes, C.; Santos-Buch, C. Chagas disease. Postgrad. Med. J. 2006, 82, 788–798. [Google Scholar] [CrossRef]
- World Health Organization. Control of Chagas Disease: Second Report of the WHO Expert Committee; World Health Organization: Geneva, Switzerland, 2002; Volume 2. [Google Scholar]
- Malik, L.H.; Singh, G.D.; Amsterdam, E.A. The epidemiology, clinical manifestations, and management of chagas heart disease. Clin. Cardiol. 2015, 38, 565–569. [Google Scholar] [CrossRef]
- de Lourdes Higuchi, M.; Fukasawa, S.; De Brito, T.; Parzianello, L.C.; Bellotti, G.; Ramires, J.A.F. Different microcirculatory and interstitial matrix patterns in idiopathic dilated cardiomyopathy and Chagas’ disease: A three dimensional confocal microscopy study. Heart 1999, 82, 279–285. [Google Scholar] [CrossRef]
- Benvenuti, L.; Roggério, A.; Freitas, H.; Mansur, A.; Fiorelli, A.; Higuchi, M. Chronic American trypanosomiasis: Parasite persistence in endomyocardial biopsies is associated with high-grade myocarditis. Ann. Trop. Med. Parasitol. 2008, 102, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Marin-Neto, J.A.; Cunha-Neto, E.; Maciel, B.C.; Simões, M.V. Pathogenesis of chronic Chagas heart disease. Circulation 2007, 115, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.A.; Tanowitz, H.B.; Malvestio, L.M.; Celes, M.R.; Campos, E.C.; Blefari, V.; Prado, C.M. Coronary microvascular disease in chronic Chagas cardiomyopathy including an overview on history, pathology, and other proposed pathogenic mechanisms. PLoS Neglected Trop. Dis. 2010, 4, e674. [Google Scholar] [CrossRef]
- Rocha, M.O.C.; Nunes, M.C.P.; Ribeiro, A.L. Morbidity and prognostic factors in chronic chagasic cardiopathy. Memórias Do Inst. Oswaldo Cruz 2009, 104, 159–166. [Google Scholar] [CrossRef]
- Rassi, A., Jr.; Rassi, S.G.; Rassi, A. Sudden death in Chagas’ disease. Arq. Bras. Cardiol. 2001, 76, 86–96. [Google Scholar] [CrossRef]
- Tripodi, K.E.; Menendez Bravo, S.M.; Cricco, J.A. Role of heme and heme-proteins in trypanosomatid essential metabolic pathways. Enzym. Res. 2011, 2011, 873230. [Google Scholar] [CrossRef]
- Friggeri, L.; Hargrove, T.Y.; Rachakonda, G.; Blobaum, A.L.; Fisher, P.; De Oliveira, G.M.; Da Silva, C.F.; Soeiro, M.d.N.C.; Nes, W.D.; Lindsley, C.W. Sterol 14α-demethylase structure-based optimization of drug candidates for human infections with the protozoan Trypanosomatidae. J. Med. Chem. 2018, 61, 10910–10921. [Google Scholar] [CrossRef] [PubMed]
- de Souza, W.; Rodrigues, J.C.F. Sterol biosynthesis pathway as target for anti-trypanosomatid drugs. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 642502. [Google Scholar] [CrossRef] [PubMed]
- Marathe, C.; Bradley, M.N.; Hong, C.; Lopez, F.; de Galarreta, C.M.R.; Tontonoz, P.; Castrillo, A. The arginase II gene is an anti-inflammatory target of liver X receptor in macrophages. J. Biol. Chem. 2006, 281, 32197–32206. [Google Scholar] [CrossRef]
- Gallardo-Soler, A.; Gómez-Nieto, C.; Campo, M.L.; Marathe, C.; Tontonoz, P.; Castrillo, A.; Corraliza, I. Arginase I induction by modified lipoproteins in macrophages: A peroxisome proliferator-activated receptor-γ/δ-mediated effect that links lipid metabolism and immunity. Mol. Endocrinol. 2008, 22, 1394–1402. [Google Scholar] [CrossRef]
- Silva-Jardim, I.; Thiemann, O.H.; Anibal, F.F. Leishmaniasis and Chagas disease chemotherapy: A critical review. J. Braz. Chem. Soc. 2014, 25, 1810–1823. [Google Scholar] [CrossRef]
- McConville, M.J.; Naderer, T. Metabolic pathways required for the intracellular survival of Leishmania. Annu. Rev. Microbiol. 2011, 6, 543–561. [Google Scholar] [CrossRef]
- Ortiz, D.; Sanchez, M.A.; Pierce, S.; Herrmann, T.; Kimblin, N.; Archie Bouwer, H.; Landfear, S.M. Molecular genetic analysis of purine nucleobase transport in Leishmania major. Mol. Microbiol. 2007, 64, 1228–1243. [Google Scholar] [CrossRef]
- Figueroa-Villar, J.D.; Sales, E.M. The importance of nucleoside hydrolase enzyme (NH) in studies to treatment of Leishmania: A review. Chem-Biol. Interact. 2017, 263, 18–27. [Google Scholar] [CrossRef]
- Majumder, H.K. Drug Targets in Kinetoplastid Parasites; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; Volume 625. [Google Scholar]
- Berg, M.; Van der Veken, P.; Goeminne, A.; Haemers, A.; Augustyns, K. Inhibitors of the purine salvage pathway: A valuable approach for antiprotozoal chemotherapy? Curr. Med. Chem. 2010, 17, 2456–2481. [Google Scholar] [CrossRef]
- Ilgoutz, S.C.; Zawadzki, J.L.; Ralton, J.E.; McConville, M.J. Evidence that free GPI glycolipids are essential for growth of Leishmania mexicana. EMBO J. 1999, 18, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.K.; Mayor, S.; Schwarz, R.T. Biosynthesis of glycosyl-phosphatidylinositol lipids in Trypanosoma brucei: Involvement of mannosyl-phosphoryldolichol as the mannose donor. EMBO J. 1990, 9, 4249–4258. [Google Scholar] [CrossRef] [PubMed]
- Horn, D. Antigenic variation in African trypanosomes. Mol. Biochem. Parasitol. 2014, 195, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.M.; Colli, W. Role of the gp85/trans-sialidase superfamily of glycoproteins in the interaction of Trypanosoma cruzi with host structures. Mol. Mech. Parasite Invasion Subcell. Biochem. 2008, 47, 58–69. [Google Scholar]
- Giorgi, M.E.; Lederkremer, R.M.d. The glycan structure of T. cruzi mucins depends on the host. Insights on the chameleonic galactose. Molecules 2020, 25, 3913. [Google Scholar] [CrossRef]
- Barreto de Albuquerque, J.; Silva dos Santos, D.; Stein, J.V.; de Meis, J. Oral versus intragastric inoculation: Similar Pathways of Trypanosoma cruzi Experimental infection? From target tissues, parasite evasion, and immune response. Front. Immunol. 2018, 9, 1734. [Google Scholar] [CrossRef]
- Vickers, T.J.; Beverley, S.M. Folate metabolic pathways in Leishmania. Essays Biochem. 2011, 51, 63–80. [Google Scholar]
- Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The concept of folic acid in health and disease. Molecules 2021, 26, 3731. [Google Scholar] [CrossRef]
- Beltran-Hortelano, I.; Alcolea, V.; Font, M.; Pérez-Silanes, S. Examination of multiple Trypanosoma cruzi targets in a new drug discovery approach for Chagas disease. Bioorganic Med. Chem. 2022, 58, 116577. [Google Scholar] [CrossRef]
- Robello, C.; Navarro, P.; Castanys, S.; Gamarro, F. A pteridine reductase gene ptr1 contiguous to a P-glycoprotein confers resistance to antifolates in Trypanosoma cruzi. Mol. Biochem. Parasitol. 1997, 90, 525–535. [Google Scholar] [CrossRef]
- Bello, A.R.; Nare, B.; Freedman, D.; Hardy, L.; Beverley, S.M. PTR1: A reductase mediating salvage of oxidized pteridines and methotrexate resistance in the protozoan parasite Leishmania major. Proc. Natl. Acad. Sci. USA 1994, 91, 11442–11446. [Google Scholar] [CrossRef]
- Krauth-Siegel, R.L.; Comini, M.A. Redox control in trypanosomatids, parasitic protozoa with trypanothione-based thiol metabolism. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2008, 1780, 1236–1248. [Google Scholar] [CrossRef]
- Colotti, G.; Ilari, A. Polyamine metabolism in Leishmania: From arginine to trypanothione. Amino Acids 2011, 40, 269–285. [Google Scholar] [CrossRef]
- Bailey, S.; Smith, K.; Fairlamb, A.H.; Hunter, W.N. Substrate interactions between trypanothione reductase and N1-glutathionylspermidine disulphide at 0.28-nm resolution. Eur. J. Biochem. 1993, 213, 67–75. [Google Scholar] [CrossRef]
- Saitoh, M.; Nishitoh, H.; Fujii, M.; Takeda, K.; Tobiume, K.; Sawada, Y.; Kawabata, M.; Miyazono, K.; Ichijo, H. Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1. EMBO J. 1998, 17, 2596–2606. [Google Scholar] [CrossRef]
- Manta, B.; Comini, M.; Medeiros, A.; Hugo, M.; Trujillo, M.; Radi, R. Trypanothione: A unique bis-glutathionyl derivative in trypanosomatids. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 3199–3216. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Sahu, U.; Kumar, A.; Khare, P. Metabolic pathways of Leishmania parasite: Source of pertinent drug targets and potent drug candidates. Pharmaceutics 2022, 14, 1590. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.d.S.; Duarte, A.P.; Pais, F.S.M.; Silva-Pereira, R.A.d.; Romanha, A.J.; Schenkman, S.; Murta, S.M.F. Overexpression of eukaryotic initiation factor 5A (eIF5A) affects susceptibility to benznidazole in Trypanosoma cruzi populations. Memórias Inst. Oswaldo Cruz 2018, 113, e180162. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Cleveland, J.L. Targeting the polyamine-hypusine circuit for the prevention and treatment of cancer. Amino Acids 2016, 48, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Pinkovich, A.; Balno, C.; Strasser, R.; Zeituni-Molad, M.; Bendelak, K.; Rentsch, D.; Ephros, M.; Wiese, M.; Jardim, A.; Myler, P.J. An arginine deprivation response pathway is induced in Leishmania during macrophage invasion. PLoS Pathog. 2016, 12, e1005494. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.F.L.; Floeter-Winter, L.M. Arginase in leishmania. Proteins Proteom. Leishmania Trypanos. 2013, 74, 103–117. [Google Scholar]
- Darlyuk, I.; Goldman, A.; Roberts, S.C.; Ullman, B.; Rentsch, D.; Zilberstein, D. Arginine homeostasis and transport in the human pathogen Leishmania donovani. J. Biol. Chem. 2009, 284, 19800–19807. [Google Scholar] [CrossRef] [PubMed]
- Maddison, J.E.; Page, S.W.; Church, D.B. Small Animal Clinical Pharmacology; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008; Volume 5. [Google Scholar]
- Marchal, B.; Van Dormael, M.; Pirard, M.; Cavalli, A.; Kegels, G.; Polman, K. Neglected tropical disease (NTD) control in health systems: The interface between programmes and general health services. Acta Trop. 2011, 120, S177–S185. [Google Scholar] [CrossRef] [PubMed]
- Moffat, J.G.; Vincent, F.; Lee, J.A.; Eder, J.; Prunotto, M. Opportunities and challenges in phenotypic drug discovery: An industry perspective. Nat. Rev. Drug Discov. 2017, 16, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Myler, P.J. Searching the Tritryp genomes for drug targets. Drug Targets Kinetoplastid Parasites 2008, 625, 133–140. [Google Scholar]
- Paradela, L.S.; Wall, R.J.; Carvalho, S.; Chemi, G.; Corpas-Lopez, V.; Moynihan, E.; Bello, D.; Patterson, S.; Güther, M.L.S.; Fairlamb, A.H. Multiple unbiased approaches identify oxidosqualene cyclase as the molecular target of a promising anti-leishmanial. Cell Chem. Biol. 2021, 28, 711–721.e8. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.; Prema, K.; Balaji, S. Machine learning models for drug–target interactions: Current knowledge and future directions. Drug Discov. Today 2020, 25, 748–756. [Google Scholar] [CrossRef] [PubMed]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [PubMed]
- Thomas, D.W. Clinical development success rates 2006–2015. BIO Ind. Anal. 2016, 1, 16. [Google Scholar]
- Surade, S.; Blundell, T.L. Structural biology and drug discovery of difficult targets: The limits of ligandability. Chem. Biol. 2012, 19, 42–50. [Google Scholar] [CrossRef]
- Gamo, F.-J.; Sanz, L.M.; Vidal, J.; De Cozar, C.; Alvarez, E.; Lavandera, J.-L.; Vanderwall, D.E.; Green, D.V.; Kumar, V.; Hasan, S. Thousands of chemical starting points for antimalarial lead identification. Nature 2010, 465, 305–310. [Google Scholar] [CrossRef]
- Don, R.; Ioset, J.-R. Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infections. Parasitology 2014, 141, 140–146. [Google Scholar] [CrossRef]
- Guiguemde, W.A.; Shelat, A.A.; Bouck, D.; Duffy, S.; Crowther, G.J.; Davis, P.H.; Smithson, D.C.; Connelly, M.; Clark, J.; Zhu, F. Chemical genetics of Plasmodium falciparum. Nature 2010, 465, 311–315. [Google Scholar] [CrossRef]
- De Rycker, M.; Wyllie, S.; Horn, D.; Read, K.D.; Gilbert, I.H. Anti-trypanosomatid drug discovery: Progress and challenges. Nat. Rev. Microbiol. 2023, 21, 35–50. [Google Scholar] [CrossRef]
- Martínez-Peinado, N.; Cortes-Serra, N.; Losada-Galvan, I.; Alonso-Vega, C.; Urbina, J.A.; Rodríguez, A.; VandeBerg, J.L.; Pinazo, M.-J.; Gascon, J.; Alonso-Padilla, J. Emerging agents for the treatment of Chagas disease: What is in the preclinical and clinical development pipeline? Expert. Opin. Investig. Drugs 2020, 29, 947–959. [Google Scholar] [CrossRef]
- Mansoldo, F.R.P.; Carta, F.; Angeli, A.; Cardoso, V.d.S.; Supuran, C.T.; Vermelho, A.B. Chagas disease: Perspectives on the past and present and challenges in drug discovery. Molecules 2020, 25, 5483. [Google Scholar] [CrossRef] [PubMed]
- Francisco, E.C.; de Almeida Junior, J.N.; Queiroz-Telles, F.; Aquino, V.R.; Mendes, A.V.A.; de Oliveira Silva, M.; Castro, P.d.T.O.e.; Guimarães, T.; Ponzio, V.; Hahn, R.C. Correlation of Trichosporon asahii genotypes with anatomical sites and antifungal susceptibility profiles: Data analyses from 284 isolates collected in the last 22 years across 24 medical centers. Antimicrob. Agents Chemother. 2021, 65, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Schiavuzzo, J.; Teixeira, J.; Melo, B.; Dos Santos, D.d.S.; Jorge, C.; Oliveira-Fusaro, M.; Parada, C. Muscle hyperalgesia induced by peripheral P2X3 receptors is modulated by inflammatory mediators. Neuroscience 2015, 285, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Mackey, T.K.; Liang, B.A.; Cuomo, R.; Hafen, R.; Brouwer, K.C.; Lee, D.E. Emerging and reemerging neglected tropical diseases: A review of key characteristics, risk factors, and the policy and innovation environment. Clin. Microbiol. Rev. 2014, 27, 949–979. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, P.A.; Santos, M.M. Recent progress in the development of indole-based compounds active against malaria, trypanosomiasis and leishmaniasis. Molecules 2022, 27, 319. [Google Scholar] [CrossRef] [PubMed]
- Reimão, J.Q.; Scotti, M.T.; Tempone, A.G. Anti-leishmanial and anti-trypanosomal activities of 1, 4-dihydropyridines: In vitro evaluation and structure–activity relationship study. Bioorganic Med. Chem. 2010, 18, 8044–8053. [Google Scholar] [CrossRef]
- Seo, Y.J.; Shin, H.; Lee, H.W.; Jung, H.R. Causes of necrotic features in fine-needle aspirates from cervical lymph nodes. J. Pathol. Transl. Med. 2021, 55, 60–67. [Google Scholar] [CrossRef]
- Yadav, C.L.; Manar, K.K.; Yadav, M.K.; Tiwari, N.; Singh, R.K.; Drew, M.G.; Singh, N. Synthesis, crystal structures and properties of new homoleptic Ni (II)/Pd (II) β-oxodithioester chelates. J. Mol. Struct. 2018, 1160, 488–496. [Google Scholar] [CrossRef]
- Sahu, A.; Kumar, D.; Agrawal, R.K. Antileishmanial drug discovery: Synthetic methods, chemical characteristics, and biological potential of quinazolines and its derivatives. Anti-Inflamm. Anti-Allergy Agents Med. Chem. (Former. Curr. Med. Chem-Anti-Inflamm. Anti-Allergy Agents) 2017, 16, 3–32. [Google Scholar] [CrossRef]
- Charlton, R.L.; Rossi-Bergmann, B.; Denny, P.W.; Steel, P.G. Repurposing as a strategy for the discovery of new anti-leishmanials: The-state-of-the-art. Parasitology 2018, 145, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Reguera, R.M.; Pérez-Pertejo, Y.; Gutiérrez-Corbo, C.; Domínguez-Asenjo, B.; Ordóñez, C.; García-Estrada, C.; Martínez-Valladares, M.; Balaña-Fouce, R. Current and promising novel drug candidates against visceral leishmaniasis. Pure Appl. Chem. 2019, 91, 1385–1404. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Investigational drugs for visceral leishmaniasis. Expert. Opin. Investig. Drugs 2015, 24, 43–59. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Nogueira, F.; Avino, V.C.; Galvis-Ovallos, F.; Pereira-Chioccola, V.L.; Moreira, M.A.B.; Romariz, A.P.P.L.; Molla, L.M.; Menz, I. Use of miltefosine to treat canine visceral leishmaniasis caused by Leishmania infantum in Brazil. Parasites Vectors 2019, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Martinez, A.K.; Rodriguez-Durán, J.; Serrano-Martin, X.; Hernandez-Rodriguez, V.; Benaim, G. Mechanism of action of miltefosine on Leishmania donovani involves the impairment of acidocalcisome function and the activation of the sphingosine-dependent plasma membrane Ca2+ channel. Antimicrob. Agents Chemother. 2018, 62, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.-C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.W.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Neglected Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef]
- Autmizguine, J.; Guptill, J.T.; Cohen-Wolkowiez, M.; Benjamin Jr, D.K.; Capparelli, E.V. Pharmacokinetics and pharmacodynamics of antifungals in children: Clinical implications. Drugs 2014, 74, 891–909. [Google Scholar] [CrossRef]
- Nagle, A.S.; Khare, S.; Kumar, A.B.; Supek, F.; Buchynskyy, A.; Mathison, C.J.; Chennamaneni, N.K.; Pendem, N.; Buckner, F.S.; Gelb, M.H. Recent developments in drug discovery for leishmaniasis and human African trypanosomiasis. Chem. Rev. 2014, 114, 11305–11347. [Google Scholar] [CrossRef]
- Singh, S.K.; Patwa, D.K.; Tilak, R.; Das, A.; Singh, T.B. In vitro susceptibility of dermatophytes to oral antifungal drugs and amphotericin B in Uttar Pradesh, India. Indian J. Dermatol. Venereol. Leprol. 2019, 85, 388. [Google Scholar] [CrossRef]
- Taslimi, Y.; Zahedifard, F.; Rafati, S. Leishmaniasis and various immunotherapeutic approaches. Parasitology 2018, 145, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Lindoso, J.A.L. Current diagnosis and treatment of cutaneous and mucocutaneous leishmaniasis. Expert. Rev. Anti-Infect. Ther. 2010, 8, 419–433. [Google Scholar] [CrossRef]
- Mansuri, R.; Singh, J.; Diwan, A. An insight into the current perspective and potential drug targets for visceral leishmaniasis (VL). Curr. Drug Targets 2020, 21, 1105–1129. [Google Scholar] [CrossRef]
- Jha, S.; Singh, N.; Jha, T. Changing response to diamidine compounds in cases of kala-azar unresponsive to antimonial. J. Assoc. Physicians India 1991, 39, 314–316. [Google Scholar] [PubMed]
- Bray, P.G.; Barrett, M.P.; Ward, S.A.; de Koning, H.P. Pentamidine uptake and resistance in pathogenic protozoa: Past, present and future. Trends Parasitol. 2003, 19, 232–239. [Google Scholar] [CrossRef]
- Coelho, A.C.; Beverley, S.M.; Cotrim, P.C. Functional genetic identification of PRP1, an ABC transporter superfamily member conferring pentamidine resistance in Leishmania major. Mol. Biochem. Parasitol. 2003, 130, 83–90. [Google Scholar] [CrossRef]
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi Jr, A.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C. Randomized trial of benznidazole for chronic Chagas’ cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar] [CrossRef] [PubMed]
- Junior, P.A.S.; Molina, I.; Murta, S.M.F.; Sánchez-Montalvá, A.; Salvador, F.; Corrêa-Oliveira, R.; Carneiro, C.M. Experimental and clinical treatment of Chagas disease: A review. Am. J. Trop. Med. Hyg. 2017, 97, 1289. [Google Scholar] [CrossRef]
- Caldas, I.S.; Santos, E.G.; Novaes, R.D. An evaluation of benznidazole as a Chagas disease therapeutic. Expert. Opin. Pharmacother. 2019, 20, 1797–1807. [Google Scholar] [CrossRef]
- Patterson, S.; Wyllie, S. Nitro drugs for the treatment of trypanosomatid diseases: Past, present, and future prospects. Trends Parasitol. 2014, 30, 289–298. [Google Scholar] [CrossRef]
- Chatelain, E.; Scandale, I. Animal models of Chagas disease and their translational value to drug development. Expert Opin. Drug Discov. 2020, 15, 1381–1402. [Google Scholar] [CrossRef]
- Vermelho, A.B.; Rodrigues, G.C.; Supuran, C.T. Why hasn’t there been more progress in new Chagas disease drug discovery? Expert Opin. Drug Discov. 2020, 15, 145–158. [Google Scholar] [CrossRef]
- Wilkinson, S.R.; Taylor, M.C.; Horn, D.; Kelly, J.M.; Cheeseman, I. A mechanism for cross-resistance to nifurtimox and benznidazole in trypanosomes. Proc. Natl. Acad. Sci. USA 2008, 105, 5022–5027. [Google Scholar] [CrossRef] [PubMed]
- Woster, P. Antiprotozoal Agents (African Trypanosomiasis, Chagas Disease, and Leishmaniasis); Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Bhattacharya, S.; Sur, D.; Karbwang, J. Childhood visceral leishmaniasis. Indian J. Med. Res. 2006, 123, 353. [Google Scholar] [PubMed]
- Sundar, S.; Singh, A. Chemotherapeutics of visceral leishmaniasis: Present and future developments. Parasitology 2018, 145, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Dorlo, T.P.; Kip, A.E.; Younis, B.M.; Ellis, S.J.; Alves, F.; Beijnen, J.H.; Njenga, S.; Kirigi, G.; Hailu, A.; Olobo, J. Visceral leishmaniasis relapse hazard is linked to reduced miltefosine exposure in patients from Eastern Africa: A population pharmacokinetic/pharmacodynamic study. J. Antimicrob. Chemother. 2017, 72, 3131–3140. [Google Scholar] [CrossRef] [PubMed]
- Mbui, J.; Olobo, J.; Omollo, R.; Solomos, A.; Kip, A.E.; Kirigi, G.; Sagaki, P.; Kimutai, R.; Were, L.; Omollo, T. Pharmacokinetics, safety, and efficacy of an allometric miltefosine regimen for the treatment of visceral leishmaniasis in Eastern African children: An open-label, phase II clinical trial. Clin. Infect. Dis. 2019, 68, 1530–1538. [Google Scholar] [CrossRef] [PubMed]
- Musa, A.M.; Younis, B.; Fadlalla, A.; Royce, C.; Balasegaram, M.; Wasunna, M.; Hailu, A.; Edwards, T.; Omollo, R.; Mudawi, M. Paromomycin for the treatment of visceral leishmaniasis in Sudan: A randomized, open-label, dose-finding study. PLoS Neglected Trop. Dis. 2010, 4, e855. [Google Scholar] [CrossRef] [PubMed]
- Jamil, K.M.; Haque, R.; Rahman, R.; Faiz, M.A.; Bhuiyan, A.T.M.R.H.; Kumar, A.; Hassan, S.M.; Kelly, H.; Dhalaria, P.; Kochhar, S. Effectiveness study of paromomycin IM injection (PMIM) for the treatment of visceral leishmaniasis (VL) in Bangladesh. PLoS Neglected Trop. Dis. 2015, 9, e0004118. [Google Scholar] [CrossRef] [PubMed]
- Jhingran, A.; Chawla, B.; Saxena, S.; Barrett, M.P.; Madhubala, R. Paromomycin: Uptake and resistance in Leishmania donovani. Mol. Biochem. Parasitol. 2009, 164, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Diro, E.; Ritmeijer, K.; Boelaert, M.; Alves, F.; Mohammed, R.; Abongomera, C.; Ravinetto, R.; De Crop, M.; Fikre, H.; Adera, C. Long-term clinical outcomes in visceral leishmaniasis-HIV co-infected patients during and after pentamidine secondary prophylaxis in Ethiopia: A single-arm clinical trial Authors and affiliations. Clin. Infect. Dis. 2017, 66, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.-E.; Mabrouk, R. The Impact of First-line Nurse Managers Leadership Development Training Program on Workgroup Climate and Performance. Doctoral Dissertation, University of Alexandria, Alexandria, Egypt, 2017. [Google Scholar]
- Balasegaram, M.; Ritmeijer, K.; Lima, M.A.; Burza, S.; Ortiz Genovese, G.; Milani, B.; Gaspani, S.; Potet, J.; Chappuis, F. Liposomal amphotericin B as a treatment for human leishmaniasis. Expert Opin. Emerg. Drugs 2012, 17, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Deray, G. Amphotericin B nephrotoxicity. J. Antimicrob. Chemother. 2002, 49, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Burza, S.; Croft, S.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Antimony toxicity. Int. J. Environ. Res. Public Health 2010, 7, 4267–4277. [Google Scholar] [CrossRef]
- Arce, A.; Estirado, A.; Ordobas, M.; Sevilla, S.; García, N.; Moratilla, L.; De La Fuente, S.; Martínez, A.; Pérez, A.; Aránguez, E. Re-emergence of leishmaniasis in Spain: Community outbreak in Madrid, Spain, 2009 to 2012. Eurosurveillance 2013, 18, 20546. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J.; Agarwal, D.; Rai, M.; Murray, H.W. Single-dose liposomal amphotericin B for visceral leishmaniasis in India. N. Engl. J. Med. 2010, 362, 504–512. [Google Scholar] [CrossRef]
- Pereira, I.R.; Vilar-Pereira, G.; Silva, A.A.d.; Lannes-Vieira, J. Severity of chronic experimental Chagas’ heart disease parallels tumour necrosis factor and nitric oxide levels in the serum: Models of mild and severe disease. Memórias Inst. Oswaldo Cruz 2014, 109, 289–298. [Google Scholar] [CrossRef]
- Molina, I.; Gómez i Prat, J.; Salvador, F.; Treviño, B.; Sulleiro, E.; Serre, N.; Pou, D.; Roure, S.; Cabezos, J.; Valerio, L. Randomized trial of posaconazole and benznidazole for chronic Chagas’ disease. N. Engl. J. Med. 2014, 370, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Lima, Á.A.; Soares-Sobrinho, J.L.; Silva, J.L.; Corrêa-Júnior, R.A.; Lyra, M.A.; Santos, F.L.; Oliveira, B.G.; Hernandes, M.Z.; Rolim, L.A.; Rolim-Neto, P.J. The use of solid dispersion systems in hydrophilic carriers to increase benznidazole solubility. J. Pharm. Sci. 2011, 100, 2443–2451. [Google Scholar] [CrossRef]
- World Health Organization. Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases: Second WHO Report on Neglected Diseases; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- de Mecca, M.M.; Fanelli, S.L.; Bartel, L.C.; de Castro, C.R.; Díaz, E.G.; Castro, J.A. Nifurtimox nitroreductase activity in different cellular fractions from male rat pancreas. Biochemical and ultrastructural alterations. Life Sci. 2007, 81, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Mecca, M.M.d.; Bartel, L.C.; Castro, C.R.d.; Castro, J.A. Benznidazole biotransformation in rat heart microsomal fraction without observable ultrastructural alterations: Comparison to Nifurtimox-induced cardiac effects. Mem. Inst. Oswaldo Cruz 2008, 103, 549–553. [Google Scholar] [CrossRef]
- Altamura, F.; Rajesh, R.; Catta-Preta, C.M.; Moretti, N.S.; Cestari, I. The current drug discovery landscape for trypanosomiasis and leishmaniasis: Challenges and strategies to identify drug targets. Drug Dev. Res. 2022, 83, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Chanda, K. An overview on the therapeutics of neglected infectious diseases—Leishmaniasis and Chagas diseases. Front. Chem. 2021, 9, 622286. [Google Scholar]
- Santos, S.S.; de Araújo, R.V.; Giarolla, J.; El Seoud, O.; Ferreira, E.I. Searching for drugs for Chagas disease, leishmaniasis and schistosomiasis: A review. Int. J. Antimicrob. Agents 2020, 55, 105906. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Wyllie, S. The critical role of mode of action studies in kinetoplastid drug discovery. Front. Drug Discov. 2023, 3, 1185679. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Singh, R. Developments in diagnosis and antileishmanial drugs. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 626838. [Google Scholar] [CrossRef]
- Alcântara, L.M.; Ferreira, T.C.; Gadelha, F.R.; Miguel, D.C. Challenges in drug discovery targeting TriTryp diseases with an emphasis on leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Roatt, B.M.; de Oliveira Cardoso, J.M.; De Brito, R.C.F.; Coura-Vital, W.; de Oliveira Aguiar-Soares, R.D.; Reis, A.B. Recent advances and new strategies on leishmaniasis treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.S.H.; Johnson, M.M.; Tokarski II, R.J.; Satoskar, A.R.; Fuchs, J.R.; Bachelder, E.M.; Ainslie, K.M. Evaluation of synergy between host and pathogen-directed therapies against intracellular Leishmania donovani. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Novais, F.O.; Amorim, C.F.; Scott, P. Host-Directed Therapies for Cutaneous Leishmaniasis. Front. Immunol. 2021, 12, 660183. [Google Scholar] [CrossRef]
- Kumari, D.; Perveen, S.; Sharma, R.; Singh, K. Advancement in leishmaniasis diagnosis and therapeutics: An update. Eur. J. Pharmacol. 2021, 910, 174436. [Google Scholar] [CrossRef]
- Rassi, A., Jr.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Parihar, S.P.; Hartley, M.-A.; Hurdayal, R.; Guler, R.; Brombacher, F. Topical simvastatin as host-directed therapy against severity of cutaneous leishmaniasis in mice. Sci. Rep. 2016, 6, 33458. [Google Scholar] [CrossRef]
- Kumar, P.; Shivam, P.; Mandal, S.; Prasanna, P.; Kumar, S.; Prasad, S.R.; Kumar, A.; Das, P.; Ali, V.; Singh, S.K. Synthesis, characterization, and mechanistic studies of a gold nanoparticle–amphotericin B covalent conjugate with enhanced antileishmanial efficacy and reduced cytotoxicity. Int. J. Nanomed. 2019, 14, 6073. [Google Scholar] [CrossRef]
- Raja, M.R.C.; Velappan, A.B.; Chellappan, D.; Debnath, J.; Mahapatra, S.K. Eugenol derived immunomodulatory molecules against visceral leishmaniasis. Eur. J. Med. Chem. 2017, 139, 503–518. [Google Scholar] [CrossRef]
- Fernández, O.L.; Rosales-Chilama, M.; Quintero, N.; Travi, B.L.; Wetzel, D.M.; Gómez, M.A.; Saravia, N.G. Potency and preclinical evidence of synergy of oral azole drugs and miltefosine in an ex vivo model of Leishmania (Viannia) panamensis infection. Antimicrob. Agents Chemother. 2022, 66, e01425-21. [Google Scholar] [CrossRef]
- Vishwakarma, P.; Parmar, N.; Chandrakar, P.; Sharma, T.; Kathuria, M.; Agnihotri, P.K.; Siddiqi, M.I.; Mitra, K.; Kar, S. Ammonium trichloro [1, 2-ethanediolato-O, O′]-tellurate cures experimental visceral leishmaniasis by redox modulation of Leishmania donovani trypanothione reductase and inhibiting host integrin linked PI3K/Akt pathway. Cell. Mol. Life Sci. 2018, 75, 563–588. [Google Scholar] [CrossRef]
- Kyriazis, I.D.; Koutsoni, O.S.; Aligiannis, N.; Karampetsou, K.; Skaltsounis, A.-L.; Dotsika, E. The leishmanicidal activity of oleuropein is selectively regulated through inflammation-and oxidative stress-related genes. Parasites Vectors 2016, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, I.; Smirlis, D.; Papadaki, A.; Koutsoni, O.; Aligiannis, N.; Skaltsounis, A.; Dotsika, E. Leishmanicidal activity of oleuropein: Leishmania donovani promastigote cell death through a possibly ROS-independent mechanism. J. Pharmacogn. Nat. Prod. 2017, 3, 141. [Google Scholar] [CrossRef]
- Dayakar, A.; Chandrasekaran, S.; Veronica, J.; Maurya, R. Leptin induces the phagocytosis and protective immune response in Leishmania donovani infected THP-1 cell line and human PBMCs. Exp. Parasitol. 2016, 160, 54–59. [Google Scholar] [CrossRef]
- Roy, A. A review on the alkaloids an important therapeutic compound from plants. IJPB 2017, 3, 1–9. [Google Scholar]
- Roy, S.; Saha, S.; Gupta, P.; Ukil, A.; Das, P.K. Crosstalk of PD-1 signaling with the SIRT1/FOXO-1 axis during the progression of visceral leishmaniasis. J. Cell Sci. 2019, 132, jcs226274. [Google Scholar] [CrossRef] [PubMed]
- Raja, M.R.C.; Kar, A.; Srinivasan, S.; Chellappan, D.; Debnath, J.; Mahapatra, S.K. Oral administration of eugenol oleate cures experimental visceral leishmaniasis through cytokines abundance. Cytokine 2021, 145, 155301. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Saha, G.; Sasidharan, S.; Dubey, V.K.; Saudagar, P. Biochemical characterization and chemical validation of Leishmania MAP Kinase-3 as a potential drug target. Sci. Rep. 2019, 9, 16209. [Google Scholar] [CrossRef] [PubMed]
- Merida-de-Barros, D.A.; Chaves, S.P.; Belmiro, C.L.R.; Wanderley, J.L.M. Leishmaniasis and glycosaminoglycans: A future therapeutic strategy? Parasites Vectors 2018, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Tindana, P.; de Haan, F.; Amaratunga, C.; Dhorda, M.; van der Pluijm, R.W.; Dondorp, A.M.; Cheah, P.Y. Deploying triple artemisinin-based combination therapy (TACT) for malaria treatment in Africa: Ethical and practical considerations. Malar. J. 2021, 20, 119. [Google Scholar] [CrossRef]
- Larkins-Ford, J.; Greenstein, T.; Van, N.; Degefu, Y.N.; Olson, M.C.; Sokolov, A.; Aldridge, B.B. Systematic measurement of combination-drug landscapes to predict in vivo treatment outcomes for tuberculosis. Cell Syst. 2021, 12, 1046–1063.e1047. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Breckenridge, A.; Jacob, R. Overcoming the legal and regulatory barriers to drug repurposing. Nat. Rev. Drug Discov. 2019, 18, 1–2. [Google Scholar] [CrossRef]
- Braga, S.S. Multi-target drugs active against leishmaniasis: A paradigm of drug repurposing. Eur. J. Med. Chem. 2019, 183, 111660. [Google Scholar] [CrossRef]
- Andrade, C.H.; Neves, B.J.; Melo-Filho, C.C.; Rodrigues, J.; Silva, D.C.; Braga, R.C.; Cravo, P.V. In silico chemogenomics drug repositioning strategies for neglected tropical diseases. Curr. Med. Chem. 2019, 26, 4355–4379. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.A.M.; Lang, K.L. Drug repositioning: Concept, classification, methodology, and importance in rare/orphans and neglected diseases. J. Appl. Pharm. Sci. 2018, 8, 157–165. [Google Scholar]
- Lima, I.D.; Lima, A.L.; Mendes-Aguiar, C.d.O.; Coutinho, J.F.; Wilson, M.E.; Pearson, R.D.; Queiroz, J.W.; Jeronimo, S.M. Changing demographics of visceral leishmaniasis in northeast Brazil: Lessons for the future. PLoS Neglected Trop. Dis. 2018, 12, e0006164. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.P.; Costa, L.E.; Lage, D.P.; Dias, D.S.; Ribeiro, P.A.; Machado, A.S.; Ramos, F.F.; Salles, B.C.; Fagundes, M.I.; Carvalho, G.B. Diagnostic application of recombinant Leishmania proteins and evaluation of their in vitro immunogenicity after stimulation of immune cells collected from tegumentary leishmaniasis patients and healthy individuals. Cell. Immunol. 2018, 334, 61–69. [Google Scholar] [CrossRef]
- Bustamante, C.; Ochoa, R.; Asela, C.; Muskus, C. Repurposing of known drugs for leishmaniasis treatment using bioinformatic predictions, in vitro validations and pharmacokinetic simulations. J. Comput-Aided Mol. Des. 2019, 33, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Tunes, L.G.; Morato, R.E.; Garcia, A.; Schmitz, V.; Steindel, M.; Correa-Junior, J.D.; Dos Santos, H.l.F.; Frezard, F.; de Almeida, M.V.; Silva, H. Preclinical gold complexes as oral drug candidates to treat leishmaniasis are potent trypanothione reductase inhibitors. ACS Infect. Dis. 2020, 6, 1121–1139. [Google Scholar] [CrossRef] [PubMed]
- Pedra-Rezende, Y.; Fernandes, M.C.; Mesquita-Rodrigues, C.; Stiebler, R.; Bombaça, A.C.S.; Pinho, N.; Cuervo, P.; De Castro, S.L.; Menna-Barreto, R.F. Starvation and pH stress conditions induced mitochondrial dysfunction, ROS production and autophagy in Trypanosoma cruzi epimastigotes. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2021, 1867, 166028. [Google Scholar] [CrossRef] [PubMed]
- Tabrez, S.; Rahman, F.; Ali, R.; Muhammad, F.; Alshehri, B.M.; Alaidarous, M.A.; Banawas, S.; Dukhyil, A.A.B.; Rub, A. Repurposing of FDA-approved drugs as inhibitors of sterol C-24 methyltransferase of Leishmania donovani to fight against leishmaniasis. Drug Dev. Res. 2021, 82, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Khadir, F.; Shaler, C.R.; Oryan, A.; Rudak, P.T.; Mazzuca, D.M.; Taheri, T.; Dikeakos, J.D.; Haeryfar, S.M.; Rafati, S. Therapeutic control of leishmaniasis by inhibitors of the mammalian target of rapamycin. PLoS Neglected Trop. Dis. 2018, 12, e0006701. [Google Scholar] [CrossRef]
- Dominguez-Asenjo, B.; Gutierrez-Corbo, C.; Perez-Pertejo, Y.; Iborra, S.; Balana-Fouce, R.; Reguera, R.M. Bioluminescent imaging identifies thymus, as overlooked colonized organ, in a chronic model of Leishmania donovani mouse visceral leishmaniasis. ACS Infect. Dis. 2021, 7, 871–883. [Google Scholar] [CrossRef]
- Jantan, I.; Bukhari, S.N.A.; Mohamed, M.A.S.; Wai, L.K.; Mesaik, M.A. The evolving role of natural products from the tropical rainforests as a replenishable source of new drug leads. In Drug Discovery and Development—From Molecules to Medicine; IntechOpen: London, UK, 2015; pp. 3–38. [Google Scholar]
- García, M.; Monzote, L. Marine products with anti-protozoal activity: A review. Curr. Clin. Pharmacol. 2014, 9, 258–270. [Google Scholar] [CrossRef]
- Chaurasia, R.; Jain, N. Current Strategies And Recent Advances In Nanostructured Delivery Systems For Managing Leishmaniasis—A Review. J. Adv. Sci. Res. 2022, 13, 43–55. [Google Scholar] [CrossRef]
- Valli, M.; Souza, J.M.; Chelucci, R.C.; Biasetto, C.R.; Araujo, A.R.; Bolzani, V.D.S.; Andricopulo, A.D. Identification of natural cytochalasins as leads for neglected tropical diseases drug discovery. PLoS ONE 2022, 17, e0275002. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.M.; Yang, L.Y.; Huang, S.S.; Chigor, V.N.; Eze, E.A.; Pan, L.X.; Zhang, T.; Yang, D.F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty 2021, 10, 9. [Google Scholar] [CrossRef]
- Cortes, S.; Bruno de Sousa, C.; Morais, T.; Lago, J.; Campino, L. Potential of the natural products against leishmaniasis in Old World—A review of in-vitro studies. Pathog. Glob. Health 2020, 114, 170–182. [Google Scholar] [CrossRef]
- Lazarin-Bidóia, D.; Garcia, F.P.; Ueda-Nakamura, T.; Silva, S.O.; Nakamura, C.V. Natural compounds based chemotherapeutic against Chagas disease and leishmaniasis: Mitochondrion as a strategic target. Mem. Inst. Oswaldo Cruz 2022, 117, e220396. [Google Scholar] [CrossRef] [PubMed]
- Gharpure, S.; Ankamwar, B. Use of nanotechnology in combating coronavirus. 3 Biotech 2021, 11, 358. [Google Scholar] [CrossRef] [PubMed]
- Karuppusamy, C.; Venkatesan, P. Role of nanoparticles in drug delivery system: A comprehensive review. J. Pharm. Sci. Res. 2017, 9, 318. [Google Scholar]
- Kim, D.Y.; Kadam, A.; Shinde, S.; Saratale, R.G.; Patra, J.; Ghodake, G. Recent developments in nanotechnology transforming the agricultural sector: A transition replete with opportunities. J. Sci. Food Agric. 2018, 98, 849–864. [Google Scholar] [CrossRef] [PubMed]
- Celis-Giraldo, C.T.; López-Abán, J.; Muro, A.; Patarroyo, M.A.; Manzano-Román, R. Nanovaccines against animal pathogens: The latest findings. Vaccines 2021, 9, 988. [Google Scholar] [CrossRef] [PubMed]
- Javed, R.; Ghonaim, R.; Shathili, A.; Khalifa, S.A.; El-Seedi, H.R. Phytonanotechnology: A greener approach for biomedical applications. In Biogenic Nanoparticles for Cancer Theranostics; Elsevier: Amsterdam, The Netherlands, 2021; pp. 43–86. [Google Scholar]
- Sangar, O.S.; Patil, A.C.; Payghan, S.A. Nanoparticles: As a Nano based Drug Delivery System. Asian J. Pharm. Clin. Res. 2022, 11, 30–35. [Google Scholar]
- Almayouf, M.A.; El-Khadragy, M.; Awad, M.A.; Alolayan, E.M. The effects of silver nanoparticles biosynthesized using fig and olive extracts on cutaneous leishmaniasis-induced inflammation in female balb/c mice. Biosci. Rep. 2020, 40, BSR20202672. [Google Scholar] [CrossRef]
- Parvez, S.; Yadagiri, G.; Singh, A.; Karole, A.; Singh, O.P.; Sundar, S.; Mudavath, S.L. Improvising anti-leishmanial activity of amphotericin B and paromomycin using co-delivery in d-α-tocopheryl polyethylene glycol 1000 succinate (TPGS) tailored nano-lipid carrier system. Chem. Phys. Lipids 2020, 231, 104946. [Google Scholar] [CrossRef]
- Bahraminejad, S.; Pardakhty, A.; Sharifi, I.; Ranjbar, M.; Karami-Mohajeri, S.; Sharifi, F. Preparation and evaluation of physicochemical properties and anti-leishmanial activity of zirconium/tioxolone niosomes against Leishmania major. Arab. J. Chem. 2022, 15, 104156. [Google Scholar]
- Singh, K.; Ahlawat, S.; Kumari, D.; Matlani, U.; Meenakshi; Kaur, T.; Rao, A. Safety of Nanoparticles: Emphasis on Antimicrobial Properties. In Biomedical Applications and Toxicity of Nanomaterials; Springer: Singapore, 2023; pp. 425–458. [Google Scholar]
- Parvez, S.; Abbas, G.; Shahid, M.; Amjad, M.; Hussain, M.; Asad, S.A.; Imran, M.; Naeem, M.A. Effect of salinity on physiological, biochemical and photostabilizing attributes of two genotypes of quinoa (Chenopodium quinoa Willd.) exposed to arsenic stress. Ecotoxicol. Environ. Saf. 2020, 187, 109814. [Google Scholar] [CrossRef]
- Gedda, M.R.; Singh, B.; Kumar, D.; Singh, A.K.; Madhukar, P.; Upadhyay, S.; Singh, O.P.; Sundar, S. Post kala-azar dermal leishmaniasis: A threat to elimination program. PLoS Neglected Trop. Dis. 2020, 14, e0008221. [Google Scholar] [CrossRef]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L. Two phase 3 trials of inclisiran in patients with elevated LDL cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef]
- Nafari, A.; Cheraghipour, K.; Sepahvand, M.; Shahrokhi, G.; Gabal, E.; Mahmoudvand, H. Nanoparticles: New agents toward treatment of leishmaniasis. Parasite Epidemiol. Control 2020, 10, e00156. [Google Scholar] [CrossRef]
- Prasanna, P.; Kumar, P.; Kumar, S.; Rajana, V.K.; Kant, V.; Prasad, S.R.; Mohan, U.; Ravichandiran, V.; Mandal, D. Current status of nanoscale drug delivery and the future of nano-vaccine development for leishmaniasis–A review. Biomed. Pharmacother. 2021, 141, 111920. [Google Scholar] [CrossRef]
- Athanasiou, E.; Agallou, M.; Tastsoglou, S.; Kammona, O.; Hatzigeorgiou, A.; Kiparissides, C.; Karagouni, E. A poly (lactic-co-glycolic) acid nanovaccine based on chimeric peptides from different Leishmania infantum proteins induces dendritic cells maturation and promotes peptide-specific IFNγ-producing CD8+ T cells essential for the protection against experimental visceral leishmaniasis. Front. Immunol. 2017, 8, 684. [Google Scholar]
- Agallou, M.; Margaroni, M.; Athanasiou, E.; Toubanaki, D.K.; Kontonikola, K.; Karidi, K.; Kammona, O.; Kiparissides, C.; Karagouni, E. Identification of BALB/c immune markers correlated with a partial protection to Leishmania infantum after vaccination with a rationally designed multi-epitope cysteine protease a peptide-based nanovaccine. PLoS Neglected Trop. Dis. 2017, 11, e0005311. [Google Scholar] [CrossRef]
- Assolini, J.P.; Carloto, A.C.M.; da Silva Bortoleti, B.T.; Gonçalves, M.D.; Pellissier, F.T.; Feuser, P.E.; Cordeiro, A.P.; de Araújo, P.H.H.; Sayer, C.; Sapla, M.M.M. Nanomedicine in leishmaniasis: A promising tool for diagnosis, treatment and prevention of disease-An update overview. Eur. J. Pharmacol. 2022, 923, 174934. [Google Scholar] [CrossRef] [PubMed]
- Roquero, I.; Cantizani, J.; Cotillo, I.; Manzano, M.P.; Kessler, A.; Martín, J.J.; McNamara, C.W. Novel chemical starting points for drug discovery in leishmaniasis and Chagas disease. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Yousef, B.A.; Elwaseela, T.H.; Ali, T.A.; Mohammed, F.E.; Mohammed, W.O.; Alobaid, M.; Dirar, A.I. Anti-malarial drugs as potential inhibitors of leishmania glycolytic enzymes: Development of new anti-leishmanial agents. Pharmacol. Clin. Pharm. Res. 2020, 5, 77–88. [Google Scholar] [CrossRef]
- Verma, A.; Ghosh, S.; Salotra, P.; Singh, R. Artemisinin-resistant Leishmania parasite modulates host cell defense mechanism and exhibits altered expression of unfolded protein response genes. Parasitol. Res. 2019, 118, 2705–2713. [Google Scholar] [CrossRef]
- Hendrickx, S.; Caljon, G.; Maes, L. Need for sustainable approaches in antileishmanial drug discovery. Parasitol. Res. 2019, 118, 2743–2752. [Google Scholar] [CrossRef] [PubMed]
- Wróbel, A.; Arciszewska, K.; Maliszewski, D.; Drozdowska, D. Trimethoprim and other nonclassical antifolates an excellent template for searching modifications of dihydrofolate reductase enzyme inhibitors. J. Antibiot. 2020, 73, 5–27. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Bharatam, P.V. Identification of selective inhibitors of Ld DHFR enzyme using pharmacoinformatic methods. J. Comput. Biol. 2021, 28, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Singh, P.; Kashyap, A.; Silakari, O. Structure based designing of benzimidazole/benzoxazole derivatives as anti-leishmanial agents. SAR QSAR Environ. Res. 2019, 30, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, P.K.; Chakraborti, S.; Bagchi, A.; Chakraborti, T. Bioassay-based Corchorus capsularis L. leaf-derived β-sitosterol exerts antileishmanial effects against Leishmania donovani by targeting trypanothione reductase. Sci. Rep. 2020, 10, 20440. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumari, E.; Bhardwaj, R.; Kumar, R.; Dubey, V.K. Molecular events leading to death of Leishmania donovani under spermidine starvation after hypericin treatment. Chem. Biol. Drug Des. 2017, 90, 962–971. [Google Scholar] [CrossRef]
- Singh, S.; Sarma, S.; Katiyar, S.P.; Das, M.; Bhardwaj, R.; Sundar, D.; Dubey, V.K. Probing the molecular mechanism of hypericin-induced parasite death provides insight into the role of spermidine beyond redox metabolism in Leishmania donovani. Antimicrob. Agents Chemother. 2015, 59, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Henninot, A.; Collins, J.C.; Nuss, J.M. The current state of peptide drug discovery: Back to the future? J. Med. Chem. 2018, 61, 1382–1414. [Google Scholar] [CrossRef]
- Qvit, N.; Kornfeld, O.S. Development of a Backbone Cyclic Peptide Library as Potential Antiparasitic Therapeutics Using Microwave Irradiation. J. Vis. Exp. 2016, 107, e53589. [Google Scholar] [CrossRef]
- Qvit, N.; Schechtman, D.; Pena, D.A.; Berti, D.A.; Soares, C.O.; Miao, Q.; Liang, L.A.; Baron, L.A.; Teh-Poot, C.; Martínez-Vega, P.; et al. Scaffold proteins LACK and TRACK as potential drug targets in kinetoplastid parasites: Development of inhibitors. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Derda, R.; Jafari, M.R. Synthetic cross-linking of peptides: Molecular linchpins for peptide cyclization. Protein Pept. Lett. 2018, 25, 1051–1075. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Jin, H.-S.; Lee, S.-E.; Kim, H.J.; Koh, H.; Lee, D.-W. New approaches towards the discovery and evaluation of bioactive peptides from natural resources. Crit. Rev. Environ. Sci. Technol. 2020, 50, 72–103. [Google Scholar] [CrossRef]
- Davda, J.; Declerck, P.; Hu-Lieskovan, S.; Hickling, T.P.; Jacobs, I.A.; Chou, J.; Salek-Ardakani, S.; Kraynov, E. Immunogenicity of immunomodulatory, antibody-based, oncology therapeutics. J. Immunother. Cancer 2019, 7, 105. [Google Scholar] [CrossRef]
- Nishioka, K.; Hayashi, S.; Farrer, M.J.; Singleton, A.B.; Yoshino, H.; Imai, H.; Kitami, T.; Sato, K.; Kuroda, R.; Tomiyama, H. Clinical heterogeneity of α-synuclein gene duplication in Parkinson’s disease. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child. Neurol. Soc. 2006, 59, 298–309. [Google Scholar] [CrossRef]
- Qvit, N.; Rubin, S.J.S.; Urban, T.J.; Mochly-Rosen, D.; Gross, E.R. Peptidomimetic therapeutics: Scientific approaches and opportunities. Drug Discov. Today 2017, 22, 454–462. [Google Scholar] [CrossRef]
- Weng, H.-B.; Chen, H.-X.; Wang, M.-W. Innovation in neglected tropical disease drug discovery and development. Infect. Dis. Poverty 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Qvit, N.; Crapster, J.A. Peptides that target protein-protein interactions as an anti-parasite strategy. Chim. Oggi/CHEMISTRY Today 2014, 32, 62–66. [Google Scholar]
- Jacobs, T.; Bruhn, H.; Gaworski, I.; Fleischer, B.; Leippe, M. NK-lysin and its shortened analog NK-2 exhibit potent activities against Trypanosoma cruzi. Antimicrob. Agents Chemother. 2003, 47, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.L.; Faria, R.X.; Calabrese, K.S.; Hardoim, D.J.; Taniwaki, N.; Alves, L.A.; De Simone, S.G. Temporizin and temporizin-1 peptides as novel candidates for eliminating Trypanosoma cruzi. PLoS ONE 2016, 11, e0157673. [Google Scholar] [CrossRef]
- Kleschenko, Y.E.; Karpenko, L.; Villalta, F. Effects of human defensin-α1 on Trypanosoma cruzi trypomastigotes in vitro. Bull. Exp. Biol. Med. 2010, 149, 731. [Google Scholar] [CrossRef]
- Pinto, E.G.; Pimenta, D.C.; Antoniazzi, M.M.; Jared, C.; Tempone, A.G. Antimicrobial peptides isolated from Phyllomedusa nordestina (Amphibia) alter the permeability of plasma membrane of Leishmania and Trypanosoma cruzi. Exp. Parasitol. 2013, 135, 655–660. [Google Scholar] [CrossRef]
- Brand, G.D.; Leite, J.R.S.; Silva, L.P.; Albuquerque, S.; Prates, M.V.; Azevedo, R.B.; Carregaro, V.; Silva, J.S.; Sá, V.C.; Brandao, R.A. Dermaseptins from Phyllomedusa oreades and Phyllomedusa distincta: Anti-Trypanosoma cruzi Activity without Cytotoxicity To Mammalian Cells. J. Biol. Chem. 2002, 277, 49332–49340. [Google Scholar] [CrossRef]
- Adade, C.M.; Oliveira, I.R.; Pais, J.A.; Souto-Padrón, T. Melittin peptide kills Trypanosoma cruzi parasites by inducing different cell death pathways. Toxicon 2013, 69, 227–239. [Google Scholar] [CrossRef]
- Freire, K.A.; Torres, M.D.T.; Lima, D.B.; Monteiro, M.L.; Martins, A.M.C.; Oliveira Jr, V.X. Wasp venom peptide as a new antichagasic agent. Toxicon 2020, 181, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.A.; Cruz, G.S.; Vieira, F.A.; Queiroz, B.R.; Freitas, C.D.; Mesquita, F.P.; Souza, P.F. Systematic review of antiprotozoal potential of antimicrobial peptides. Acta Trop. 2022, 236, 106675. [Google Scholar] [CrossRef] [PubMed]
- El-Dirany, R.; Shahrour, H.; Dirany, Z.; Abdel-Sater, F.; Gonzalez-Gaitano, G.; Brandenburg, K.; Martinez de Tejada, G.; Nguewa, P.A. Activity of anti-microbial peptides (AMPs) against Leishmania and other parasites: An overview. Biomolecules 2021, 11, 984. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.E.; Crowe, S.E. Celiac Disease. Ann. Intern. Med. 2020, 172, Itc1–Itc16. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919. [Google Scholar]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Lewies, A.; Wentzel, J.F.; Jacobs, G.; Du Plessis, L.H. The potential use of natural and structural analogues of antimicrobial peptides in the fight against neglected tropical diseases. Molecules 2015, 20, 15392–15433. [Google Scholar] [CrossRef] [PubMed]
- Robledo, S.M.; Pérez-Silanes, S.; Fernández-Rubio, C.; Poveda, A.; Monzote, L.; González, V.M.; Alonso-Collado, P.; Carrión, J. Neglected Zoonotic Diseases: Advances in the Development of Cell-Penetrating and Antimicrobial Peptides against Leishmaniosis and Chagas Disease. Pathogens 2023, 12, 939. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Pirela, M.; Kemmerling, U.; Quiñones, W.; Michels, P.A.; Rojas, V. Antimicrobial Peptides (AMPs): Potential Therapeutic Strategy against Trypanosomiases? Biomolecules 2023, 13, 599. [Google Scholar] [CrossRef] [PubMed]
- Robles-Loaiza, A.A.; Pinos-Tamayo, E.A.; Mendes, B.; Teixeira, C.; Alves, C.; Gomes, P.; Almeida, J.R. Peptides to tackle leishmaniasis: Current status and future directions. Int. J. Mol. Sci. 2021, 22, 4400. [Google Scholar] [CrossRef] [PubMed]
- Price, R.L. Determining the Effect of Treatment with an Exogenous Host Defence Peptide on Mycobacterium marinum-Zebrafish Infection. Ph.D. Thesis, Imperial College London, London, UK, 2017. [Google Scholar]
- Zahedifard, F.; Lee, H.; No, J.H.; Salimi, M.; Seyed, N.; Asoodeh, A.; Rafati, S. Anti-leishmanial activity of Brevinin 2R and its Lauric acid conjugate type against L. major: In vitro mechanism of actions and in vivo treatment potentials. PLoS Neglected Trop. Dis. 2019, 13, e0007217. [Google Scholar]
- Rivas, L.; Luque-Ortega, J.R.; Andreu, D. Amphibian antimicrobial peptides and Protozoa: Lessons from parasites. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 1570–1581. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Salas-Sarduy, E.; Niemirowicz, G.T.; José Cazzulo, J.; Alvarez, V.E. Target-based screening of the Chagas box: Setting up enzymatic assays to discover specific inhibitors across bioactive compounds. Curr. Med. Chem. 2019, 26, 6672–6686. [Google Scholar] [CrossRef]
- Fieck, A.; Hurwitz, I.; Kang, A.S.; Durvasula, R. Trypanosoma cruzi: Synergistic cytotoxicity of multiple amphipathic anti-microbial peptides to T. cruzi and potential bacterial hosts. Exp. Parasitol. 2010, 125, 342–347. [Google Scholar] [CrossRef]
- Sabia, E.F., Jr.; Menezes, L.F.S.; de Araújo, I.F.S.; Schwartz, E.F. Natural occurrence in venomous arthropods of antimicrobial peptides active against protozoan parasites. Toxins 2019, 11, 563. [Google Scholar] [CrossRef]
- Richter, A.; Sutherland, D.; Ebrahimikondori, H.; Babcock, A.; Louie, N.; Li, C.; Coombe, L.; Lin, D.; Warren, R.L.; Yanai, A. Associating Biological Activity and Predicted Structure of Antimicrobial Peptides from Amphibians and Insects. Antibiotics 2022, 11, 1710. [Google Scholar] [CrossRef]
- Telleria, E.L.; Tinoco-Nunes, B.; Leštinová, T.; de Avellar, L.M.; Tempone, A.J.; Pitaluga, A.N.; Volf, P.; Traub-Csekö, Y.M. Lutzomyia longipalpis antimicrobial peptides: Differential expression during development and potential involvement in vector interaction with microbiota and leishmania. Microorganisms 2021, 9, 1271. [Google Scholar] [CrossRef]
- Fang, Y.; He, X.; Zhang, P.; Shen, C.; Mwangi, J.; Xu, C.; Mo, G.; Lai, R.; Zhang, Z. In vitro and in vivo antimalarial activity of LZ1, a peptide derived from snake cathelicidin. Toxins 2019, 11, 379. [Google Scholar] [CrossRef]
- Adade, C.M.; Souto-Padrón, T. Venoms as sources of novel anti-parasitic agents. Toxins Drug Discov. 2015, 1–31. [Google Scholar] [CrossRef]
- De Rycker, M.; Baragaña, B.; Duce, S.L.; Gilbert, I.H. Challenges and recent progress in drug discovery for tropical diseases. Nature 2018, 559, 498–506. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef] [PubMed]
- Vargas Buonfiglio, L.G.; Vanegas Calderon, O.G.; Cano, M.; Simmering, J.E.; Polgreen, P.M.; Zabner, J.; Gerke, A.K.; Comellas, A.P. Seasonal Antimicrobial Activity of the Airway: Post-Hoc Analysis of a Randomized Placebo-Controlled Double-Blind Trial. Nutrients 2020, 12, 2602. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef] [PubMed]
- Giovati, L.; Ciociola, T.; Magliani, W.; Conti, S. Antimicrobial peptides with antiprotozoal activity: Current state and future perspectives. Future Med. Chem. 2018, 10, 2569–2572. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.L.G.; de Moraes, J.; Andricopulo, A.D. Approaches to advance drug discovery for neglected tropical diseases. Drug Discov. Today 2022, 27, 2278–2287. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.B.; Witt, G.; Kuzikov, M.; Ellinger, B.; Calogeropoulou, T.; Prousis, K.C.; Mangani, S.; Di Pisa, F.; Landi, G.; Iacono, L.D.; et al. Accelerating Drug Discovery Efforts for Trypanosomatidic Infections Using an Integrated Transnational Academic Drug Discovery Platform. SLAS Discov. 2019, 24, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Kalesh, K. Defeating the trypanosomatid trio: Proteomics of the protozoan parasites causing neglected tropical diseases. RSC Med. Chem. 2020, 11, 625–645. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic alpha-helical cationic antimicrobial peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef]
- Bittencourt, C.R.; de Oliveira Farias, E.A.; Bezerra, K.C.; Véras, L.M.C.; Silva, V.C.; Costa, C.H.N.; Bemquerer, M.P.; Silva, L.P.; Souza de Almeida Leite, J.R.; Eiras, C. Immobilization of cationic antimicrobial peptides and natural cashew gum in nanosheet systems for the investigation of anti-leishmanial activity. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 59, 549–555. [Google Scholar] [CrossRef]
- Mendes, B.; Almeida, J.R.; Vale, N.; Gomes, P.; Gadelha, F.R.; Da Silva, S.L.; Miguel, D.C. Potential use of 13-mer peptides based on phospholipase and oligoarginine as leishmanicidal agents. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 226, 108612. [Google Scholar] [CrossRef]
- Rangel, M.; Cabrera, M.P.; Kazuma, K.; Ando, K.; Wang, X.; Kato, M.; Nihei, K.; Hirata, I.Y.; Cross, T.J.; Garcia, A.N.; et al. Chemical and biological characterization of four new linear cationic α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 2011, 57, 1081–1092. [Google Scholar] [CrossRef]
- Brand, G.D.; Leite, J.R.; de Sá Mandel, S.M.; Mesquita, D.A.; Silva, L.P.; Prates, M.V.; Barbosa, E.A.; Vinecky, F.; Martins, G.R.; Galasso, J.H.; et al. Novel dermaseptins from Phyllomedusa hypochondrialis (Amphibia). Biochem. Biophys. Res. Commun. 2006, 347, 739–746. [Google Scholar] [CrossRef]
- Díaz-Achirica, P.; Ubach, J.; Guinea, A.; Andreu, D.; Rivas, L. The plasma membrane of Leishmania donovani promastigotes is the main target for CA(1-8)M(1-18), a synthetic cecropin A-melittin hybrid peptide. Biochem. J. 1998, 330 Pt 1, 453–460. [Google Scholar] [CrossRef]
- Pérez-Cordero, J.J.; Lozano, J.M.; Cortés, J.; Delgado, G. Leishmanicidal activity of synthetic antimicrobial peptides in an infection model with human dendritic cells. Peptides 2011, 32, 683–690. [Google Scholar] [CrossRef]
- Kückelhaus, S.A.; Leite, J.R.; Muniz-Junqueira, M.I.; Sampaio, R.N.; Bloch, C., Jr.; Tosta, C.E. Antiplasmodial and antileishmanial activities of phylloseptin-1, an antimicrobial peptide from the skin secretion of Phyllomedusa azurea (Amphibia). Exp. Parasitol. 2009, 123, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Papo, N.; Saugar, J.M.; Barra, D.; Shai, Y.; Simmaco, M.; Rivas, L. Effect of natural L- to D-amino acid conversion on the organization, membrane binding, and biological function of the antimicrobial peptides bombinins H. Biochemistry 2006, 45, 4266–4276. [Google Scholar] [CrossRef] [PubMed]
- Marr, A.; Cen, S.; Hancock, R.; McMaster, W. Identification of synthetic and natural host defense peptides with leishmanicidal activity. Antimicrob. Agents Chemother. 2016, 60, 2484–2491. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Márquez, I.A.; Patiño-González, E.; Hernández-Villa, L.; Ortíz-Reyes, B.; Manrique-Moreno, M. Identification and evaluation of Galleria mellonella peptides with antileishmanial activity. Anal. Biochem. 2018, 546, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rhaiem, R.B.; Houimel, M. Targeting Leishmania major parasite with peptides derived from a combinatorial phage display library. Acta Trop. 2016, 159, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Zanetti, M. Cathelicidins, multifunctional peptides of the innate immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef]
- Li, J.; Koh, J.J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef]
- Oñate-Garzón, J.; Manrique-Moreno, M.; Trier, S.; Leidy, C.; Torres, R.; Patiño, E. Antimicrobial activity and interactions of cationic peptides derived from Galleria mellonella cecropin D-like peptide with model membranes. J. Antibiot. 2017, 70, 238–245. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M. Antimicrobial Peptides: From Design to Clinical Application. Antibiotics 2022, 11, 349. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.D.; Qvit, N.; Mochly-Rosen, D. Peptides and peptidomimetics as regulators of protein-protein interactions. Curr. Opin. Struct. Biol. 2017, 44, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Rubin, S.; Qvit, N. Cyclic Peptides for Protein-Protein Interaction Targets: Applications to Human Disease. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Rapposelli, S.; Gaudio, E.; Bertozzi, F.; Gul, S. Editorial: Protein-Protein Interactions: Drug Discovery for the Future. Front. Chem. 2021, 9, 811190. [Google Scholar] [CrossRef]
- Gundampati, R.K.; Sahu, S.; Shukla, A.; Pandey, R.K.; Patel, M.; Banik, R.M.; Jagannadham, M.V. Tryparedoxin peroxidase of Leishmania braziliensis: Homology modeling and inhibitory effects of flavonoids for anti-leishmanial activity. Bioinformation 2014, 10, 353. [Google Scholar] [CrossRef]
- Venkatesan, S.K.; Saudagar, P.; Shukla, A.K.; Dubey, V.K. Screening natural products database for identification of potential antileishmanial chemotherapeutic agents. Interdiscip. Sci. Comput. Life Sci. 2011, 3, 217–231. [Google Scholar] [CrossRef]
- Amiri-Dashatan, N.; Rezaei-Tavirani, M.; Ranjbar, M.M.; Koushki, M.; Nasab, S.D.M.; Ahmadi, N. Discovery of Novel Pyruvate Kinase Inhibitors Against Leishmania major among FDA Approved Drugs Through System Biology and Molecular Docking Approach. Turk. J. Pharm. Sci. 2021, 18, 710. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The life cycle of Leishmania species and Trypanosoma cruzi. The figure elaborates the similarities and differences between the two parasitic infestations involving the vector, host, and stages of parasitic differentiation. Though there are few similarities in the parasitic differentiation of Chagas and leishmaniasis in the host cells, the figure emphasizes parasitic differentiation in the vector. During a blood meal, it is also transmitted to the host. When insects feed on blood, they release feces that contain metacyclic trypomastigotes. A bite wound or conjunctiva allows the parasite to enter the bloodstream. When infiltrating the bloodstream, T. cruzi can infect any nucleated cell. Once the parasite enters a host cell and transforms into an amastigote, it reproduces in the cytoplasm. An intracellular parasite matures into a trypomastigote after several replications. When a mammalian host is disrupted, trypomastigotes are released into the bloodstream and ingested by triatomines during a blood meal. Epimastigotes and metacyclic trypomastigotes are present in the insect gut at various developmental stages. The figure also distinguishes between various steps involved in parasite development inside the cellular compartments and provides a complete life cycle illustration with a comparison of the life cycle of the parasites inside the vector and the host. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.
Figure 1.
The life cycle of Leishmania species and Trypanosoma cruzi. The figure elaborates the similarities and differences between the two parasitic infestations involving the vector, host, and stages of parasitic differentiation. Though there are few similarities in the parasitic differentiation of Chagas and leishmaniasis in the host cells, the figure emphasizes parasitic differentiation in the vector. During a blood meal, it is also transmitted to the host. When insects feed on blood, they release feces that contain metacyclic trypomastigotes. A bite wound or conjunctiva allows the parasite to enter the bloodstream. When infiltrating the bloodstream, T. cruzi can infect any nucleated cell. Once the parasite enters a host cell and transforms into an amastigote, it reproduces in the cytoplasm. An intracellular parasite matures into a trypomastigote after several replications. When a mammalian host is disrupted, trypomastigotes are released into the bloodstream and ingested by triatomines during a blood meal. Epimastigotes and metacyclic trypomastigotes are present in the insect gut at various developmental stages. The figure also distinguishes between various steps involved in parasite development inside the cellular compartments and provides a complete life cycle illustration with a comparison of the life cycle of the parasites inside the vector and the host. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.

Figure 2.
Clinical presentation of leishmaniasis. Clinical manifestations of individual leishmaniasis types during infection. It is characterized by early and later stages of infection and possible conditions. Infection is associated with other types of leishmania infections, complicating the patient’s condition. The figure also elaborates on the possible co-infections with leishmaniasis to provide the broader aspects of the infection. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.
Figure 2.
Clinical presentation of leishmaniasis. Clinical manifestations of individual leishmaniasis types during infection. It is characterized by early and later stages of infection and possible conditions. Infection is associated with other types of leishmania infections, complicating the patient’s condition. The figure also elaborates on the possible co-infections with leishmaniasis to provide the broader aspects of the infection. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.

Figure 3.
Diagram of a clinical representation of Chagas disease. Chagas disease has two stages or clinical phases: an acute and a chronic phase. Most infected people (between 70 and 80%) remain asymptomatic their entire lives, but in 20 to 30% of cases, the disease progresses to cause chronic symptoms. The infection is also associated with other diseases and infections, thus complicating the patient’s condition. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.
Figure 3.
Diagram of a clinical representation of Chagas disease. Chagas disease has two stages or clinical phases: an acute and a chronic phase. Most infected people (between 70 and 80%) remain asymptomatic their entire lives, but in 20 to 30% of cases, the disease progresses to cause chronic symptoms. The infection is also associated with other diseases and infections, thus complicating the patient’s condition. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.

Figure 4.
Chemical structures of commonly available leishmaniasis drugs.

Figure 5.
The chemical structures of currently used drugs against Chagas disease.

Figure 6.
The mode of action, biochemical characterization, and potency of current drugs in leishmania and Chagas infection against the parasites.
Figure 6.
The mode of action, biochemical characterization, and potency of current drugs in leishmania and Chagas infection against the parasites.

Figure 7.
Schematic representations of T. cruzi and Leishmania life cycle-associated anti-microbial peptides (AMPs) against each specific developmental form of the parasite. The figure elaborates on different AMPs acting on parasitic differentiation stages in both host and vector. This provides a detailed view of the AMPs’ treatment regimen. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.
Figure 7.
Schematic representations of T. cruzi and Leishmania life cycle-associated anti-microbial peptides (AMPs) against each specific developmental form of the parasite. The figure elaborates on different AMPs acting on parasitic differentiation stages in both host and vector. This provides a detailed view of the AMPs’ treatment regimen. BioRender.com (accessed on 25 September 2023) was used to generate this Figure.

Table 1.
Geographical and species distribution of leishmaniasis and Chagas disease.
| No | Diseases | Species | Types | Geographic Distributions | References |
|---|---|---|---|---|---|
| 1 | Leishmaniasis | Leishmania (L.) Aethiopica | Cutaneous and mucocutaneous leishmaniasis | Ethiopia, Kenya | [39] |
| 2 | L. Tropica | Visceral and cutaneous leishmaniasis | Eastern and Northern India, Central Asia, East and South Africa, Middle East | [39] | |
| 3 | L. Amazonensis | Cutaneous and mucocutaneous leishmaniasis | Brazil, Bolivia, Venezuela | [40] | |
| 4 | L. Infantum | Visceral and cutaneous leishmaniasis | Mexico, Brazil, Bolivia, Venezuela, Northern Africa, Middle East, Mediterranean regions, Southern Europe, Central Asia | [41] | |
| 5 | L. Donovani | Post Kala azar dermal leishmaniasis, visceral and cutaneous leishmaniasis | India, Myanmar, Nepal, Sri Lanka, Middle East, China, Ethiopia, Kenya, Sudan | [41] | |
| 6 | L. Major | Cutaneous and mucocutaneous leishmaniasis | Central Asia, Middle East, Central, Northern and West Africa | [42] | |
| 7 | L. Mexicana | Cutaneous and visceral leishmaniasis | United States of America, Venezuela, Ecuador, Peru, Brazil | [40] | |
| 8 | L. Venezuelensis | Cutaneous leishmaniasis | Northern and Southern America, Venezuela | [43] | |
| 9 | L. Braziliensis | Cutaneous and mucocutaneous leishmaniasis | Amazon stretch, Brazil, Southern America, Bolivia, Peru, Venezuela | [44] | |
| 10 | L. Guyanensis | Cutaneous and mucocutaneous leishmaniasis | Southern America, French Guiana, Suriname, Brazil | [45] | |
| 11 | L. Panamensis | Cutaneous and mucocutaneous leishmaniasis | Panama, South and Northern America, Brazil, Ecuador, Columbia, Venezuela | [45] | |
| 12 | L. Lainsoni | Cutaneous leishmaniasis | French Guiana, Peru, Bolivia, Brazil | [46] | |
| 13 | L. Naffi | Cutaneous leishmaniasis | French Guiana, Brazil | [46] | |
| 14 | L. Lindenberg | Cutaneous leishmaniasis | Brazil | [47] | |
| 15 | L. Peruviana | Cutaneous and mucocutaneous leishmaniasis | Peru, Bolivia, Amazon | [48] | |
| 16 | L. Shawi | Cutaneous leishmaniasis | Brazil | [49] | |
| 17 | L. Martiniquensis | Visceral and cutaneous leishmaniasis | Martinique, Thailand, France, Germany, Switzerland, Myanmar | [50] | |
| 18 | L. Siamensis | Visceral and cutaneous leishmaniasis | Central Europe, Thailand, United States of America | [51] | |
| 19 | L. Colombiensis | Visceral and cutaneous leishmaniasis | Columbia | [52] | |
| 20 | African Trypanosomiasis (aka Sleeping Sickness) | T. brucei | Acute and chronic infections | Eastern, Western, Southern and Central Africa | [53] |
| 21 | Chagas disease (aka American trypanosomiasis) | T. cruzi | Acute and chronic infections | Bolivia, Argentina, Paraguay, Ecuador, El Salvador, Guatemala | [54] |
Table 3.
Drugs/compounds with anti-leishmanial effects identified through a host-directed therapy nanotechnology approach.
Table 3.
Drugs/compounds with anti-leishmanial effects identified through a host-directed therapy nanotechnology approach.
| Drug/ Compounds | Disease | Target Organism | Model | Outcomes | References |
|---|---|---|---|---|---|
| Imatinib | CL | L. amazonensis | C57BL/6 mice | A reduction in abrasion in mice and an increase in phagocytosis | [209] |
| AS101 (tellurium-based compound) | VL | L. donovani | BALB/c mice | An increase in the production of ROS and NO. Nuclear factor kappa B (NF-κB) and mitogen-activated protein kinase pathways are activated | [210] |
| Oleuropein | VL | L. donovani | J774A.1 cell line and BALB/c mice | Inflammatory response accompanied by increased levels of interferon-gamma (IFN-γ) and IL-12 | [211,212] |
| Leptin | VL | L. donovani | THP-1 cell line | Increase NO production and promote Th1 response to kill parasites | [213] |
| Mahanine | VL | L. donovani | J774A.1 cell line and BALB/c mice | Inhibits the production of Th2 cytokines (IL10) while modulating Th1 cytokines | [214,215] |
| Simvastatin | CL | L. major | BALB/c mice | The drug inhibits cholesterol biosynthesis and reduces promastigotes’ attachment to the host | [206] |
| Eugenol | CL | L. amazonensis | BALB/c mice | The Th1 immune response is triggered by the release of IL-12 and IFN-γ | [216,217] |
| Phospholipase A2 | CL | L. amazonensis | BALB/c mice | The stimulation of NF-κB in macrophages and tumor necrosis factor-alpha (TNF-alpha) production (increased Th1 immune response) and NO production is also enhanced | [218] |
Reproduced with permission from [203].
Table 4.
Drugs/compounds with anti-leishmanial effects that were identified through drug repurposing.
Table 4.
Drugs/compounds with anti-leishmanial effects that were identified through drug repurposing.
| Drug/Compounds | Disease | Target Organism | Model | Outcomes | References |
|---|---|---|---|---|---|
| Sertraline | VL | L. infantum | BALB/c mice | Reduced the parasite’s growth significantly. In addition to oxidative damage, essential metabolic pathways were altered | [226,227] |
| Cladribine, Lamivudine, Metformin, Perphenazine, Rifabutin, and Tenofovir | CL | L. braziliensis and L. panamensis | U-937 cell line | According to in vitro studies, rifabutin and perphenazine inhibited parasites better than the other drugs | [228] |
| Gold (I) triphenyl- phosphine and triethyl- phosphine based complexes | CL | L. amazonensis and L. braziliensis | BALB/c mice | It is reported that the IC50 for the anti-leishmanial activity is 0.5–5.5 μM. ROS-mediated cell death is caused by the inhibition of trypanothione reductase activity | [229] |
| Triclosan | CL | L. amazonensis | BALB/c mice | The parasite was inhibited in its growth, as well as mitochondria were damaged and membrane permeability was reduced | [230] |
| Simeprevir | VL | L. donovani | THP-1 cell line | ROS-mediated cell death combined with leishmanicidal properties against Promastigotes and Amastigotes | [231] |
| Rapamycin | CL | L. major and L. tropica | BALB/c mice | Proliferation of the parasite was markedly inhibited. A reduction in parasite load in mice along with the polarization of the immune system towards Th1 was observed | [232] |
| Nifuratel | CL and VL | L. major, L. infantum, and L. donovani | BALB/c mice | In a mouse model, both oral and intralesional administration cured cutaneous and visceral infections | [233] |
Reproduced with permission from [203].
Table 5.
Drugs/compounds with anti-leishmanial effects identified through nanotechnology approach.
| Drug/Compounds | Disease | Target Organism | Model | Outcomes | References |
|---|---|---|---|---|---|
| Silver NPs containing Fig and Olive extracts | CL | L. major | BALB/c mice | Reduction in skin lesions and improved antioxidative capacity | [248] |
| Nano-hydrogels loaded with buparvaquone | CL | L. amazonensis | BALB/c mice | Infected BALB/c mice showed a significant decrease in parasitic burden of 95% | [249] |
| Chitosan/CdO core shell nanodots | CL | L. major | THP-1 cell line | The compound inhibited promastigotes proliferation and induced Th1 immunity at IC50 0.6 μL/mL | [250] |
| Cyclodextrin NPs containing Amp B and paromomycin | VL | L. donovani | J774A.1 cell line and Swiss albino mice | By inhibiting parasite growth with an IC50 0.013 μg/mL, it reduced the parasitic burden by 70–90% | [251] |
| Chitosan NPs containing Amp B and paromomycin | VL | L. donovani | J774A.1 cell line | NPs inhibit parasite growth better than Amp B alone, and >90% parasite clearance was reported | [252] |
| Nanotube appended with Amp B | VL | L. donovani | J774A.1 cell line and golden hamster | Composite graphene–carbon nanotubes significantly reduced parasitic proliferation by >90% when compared to AmB alone, proving that it is a safe cure for parasites | [253] |
| Guar gum NPs containing and Amp B | VL | L. donovani | Golden hamster | It inhibits parasites by 2–3 fold compared to drug alone and reduces parasitic burden by 95% | [254] |
Reproduced with permission from [203].
Table 6.
Lists of novel anti-leishmanial drugs that target specific biochemical pathways.
| Drug Candidate | Drug Target | Mode of Action | References |
|---|---|---|---|
| Artesunate Quinine Mefloquine | GAPDH | Inhibits the parasites’ glycolytic enzymes GAPDH | [261,262] |
| Cycloguanil | DHFR | Inhibits DHFR | [263,264,265] |
| Trimethoprim (TMP, 2) | |||
| ZINC57774418 (Z18) | |||
| ZINC69844431 (Z31) | |||
| ZINC71746025 (Z25) | |||
| D11596 (DB96) | Inhibits DHFR activity | [265] | |
| 2-(4-((2,4- dichlorobenzyl)oxy)phenyl)-1Hbenzo[d]imidazole | DHFR and PTR1 | DHFR-TS/PTR1 inhibitors | [266] |
| 2-(4-((2,4- dichlorobenzyl)oxy)phenyl)-1Hbenzo[d]imidazole-1Hbenzo[d]oxazole | DHFR and PTR1 | ||
| Trichloro[1,2-ethanediolato-O,O’]tellurate (AS101) | TR | Induces ROS-mediated apoptosis by binding to TR cysteine residues | [210] |
| β-sitosterol CCL | Inhibit TR activity | [267] | |
| Hypericin | Spermidine synthase | ROS and spermidine reduction | [268,269] |
Reproduced with permission from [4].
Table 7.
Studies involving advanced peptides against leishmaniasis and Chagas disease.
| Peptide | Source | Sequence | IC50 µg/mL | Study Model | Reference |
|---|---|---|---|---|---|
| NK-2 | Synthetic peptide | KILRGVCKKIMRTFLRRISKDILTGKK | - | in vitro | [281] |
| Temporizin-1 | Synthetic peptide | FLPLWLWLWLWLWKLK | - | in vitro | [282] |
| Defensin-α1 | Homo sapiens (Human) | ACYCRIPACIAGERRYGTCIYQGRLWAFCC | - | in vitro | [283] |
| Phylloseptin 7 | Phyllomedusa nordestina (Frog) | FLSLIPHAINAVSAIAKHF | 0.34 | in vitro | [284] |
| DS 01 | Phyllomedusa oreades (Frog) | GLWSTIKQKGKEAAIAAAKAAGQAALGAL | - | in vitro | [285] |
| Melittin | Apis mellifera (Bee) | - | 2.44 | in vitro | [286] |
| Polybia-CP | Polybia paulista (Wasp) | ILGTILGLLSKL | - | in vitro | [287] |
| Hmc 364–382 | Penaeus monodon (Shrimp) | NVQYYGALHNTAHIVLGRQ | 4.79 | in vitro | [287] |
Reproduced with permission from [288].
Table 8.
Structural analysis of selected peptide profiles for leishmaniasis and Chagas disease.
| Species (Diseases) | Peptide Name | Sources | Sequences | Study Model | Net Charge | Hydrophobicity (kcal/mol) | PI | Length | References |
|---|---|---|---|---|---|---|---|---|---|
| Leishmania | DRS 01 | Phyllomedusa | GLWSTIKQKGKEAAIAAAKAAGQAALGAL | in vitro | +3 | +26.50 | 10.60 | 29 | [318] |
| Leishmania | Dermaseptin 4 | Phyllomedusa | GLWSTIKQKGKEAAIAAAKAAGKAALNAASEAL | in vitro | +3 | +33.32 | 10.43 | 33 | [284] |
| Leishmania | Dermaseptin 1 | Nordestina | GLWSTIKNVGKEAAIAAGKAALGAL | in vitro | +2 | +21.05 | 10.37 | 25 | [284] |
| Leishmania | p-Acl | Agkistrodon contortrix | KKYKAYFKFKCKK | in vitro | +7 | +23.14 | 10.45 | 13 | [319] |
| Leishmania | p-AclR7 | Laticinctus | RRYRAYFRFRCRR | in vitro | +7 | +16.21 | 12.36 | 13 | [319] |
| Leishmania | Eumenitin-R | Eumenes rubrofemoratus | LNLKGLIKKVASLLN | in vitro | +3 | +12.28 | 10.89 | 15 | [320] |
| Leishmania | Dermaseptin-01 | Amphibian | GLWSTIKNVGKEAAIAAGKAALGAL | - | +2 | +21.05 | 10.35 | 25 | [321] |
| Leishmania | Dermaseptin-H3 | Amphibian | GLWSTIKNVGEAAIAAGKAALGAL | - | +1 | +18.25 | 9.95 | 24 | [321] |
| Leishmania | Melittin | Insect | GIGAVLKVLTTGLPALISWIKRKQQ | - | +4 | +13.83 | 11.79 | 24 | [322] |
| Leishmania | Melittin | Insect | GIGAVLTTGLPALISWIKRKRQQ | - | +4 | +14.55 | 12.51 | 23 | [322,323] |
| Leishmania | Phylloseptin-1 | Amphibian | FLSLIPHAINAVSAIAKHN | - | +1 | +12.09 | 9.93 | 19 | [324] |
| Leishmania | Bombinin H | Amphibian | IIGPVLGLVGSALGGLLKKI | - | +2 | +9.82 | 10.65 | 20 | [325] |
| Leishmania | LL-37 | Cathelicidin | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | in vitro | +6 | +41.03 | 11.15 | 37 | [326] |
| Leishmania | E6 | Synthetic | RRWRIVVIRVRR | in vitro | +6 | +13.05 | 13.18 | 12 | [326] |
| Leishmania | cecropin-A | Hemolymph of the giant silkworm Hyalophora cecropia | KWKLFKKIEKVGQNIRDGIIKAGPAVAWVGQATQIAK | in vitro | +6 | +33.11 | 10.94 | 37 | [327] |
| Leishmania | Cecropin-D | Galleria mellonella hemolymph | ENFFKEIERAGQRIRDAIISAAPAVETLAQAQKIIKGGD | in vitro | 0 | +42.64 | 7.07 | 39 | [323] |
| Leishmania | Eumenitin-F | LNLKGIFKKVASLLT | in vitro | +3 | +11.22 | 10.89 | 15 | [303] | |
| Leishmania | Eumenitin-R | LNLKGLIKKVASLLN | in vitro | +3 | +12.28 | 10.89 | 15 | [303] | |
| Leishmania | P1 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | MASKPQR | in vitro and in vivo | +2 | +13.71 | 11.53 | 7 | [328] |
| Leishmania | P2 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | MAAKYN | in vitro and in vivo | +1 | +11.17 | 9.58 | 6 | [328] |
| Leishmania | P3 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | MAHYSG | in vitro and in vivo | 0 | +10.96 | 7.63 | 6 | [328] |
| Leishmania | P4 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | MYVIRG | in vitro and in vivo | +1 | +7.90 | 9.68 | 6 | [328] |
| Leishmania | P5 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | SLSWVC | in vitro and in vivo | 0 | +5.00 | 4.93 | 6 | [328] |
| Leishmania | P6 | Direct screening of a linear hexa-peptide library on L. major metacyclic parasite | QRKMAS | in vitro and in vivo | +2 | +13.57 | 11.52 | 6 | [328] |
| Chagas disease | Mucin-associated surface proteins (MASP) | SLLSDAENPGGEVFNDNK | in vitro | −3 | +26.98 | 3.53 | 18 | [328] | |
| Chagas disease | Mucin-associated surface proteins (MASP) | DAENPGGEVFNDNKKGLSRV | in vitro | −1 | +33.07 | 4.48 | 20 | [328] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Berhe, H.; Kumar Cinthakunta Sridhar, M.; Zerihun, M.; Qvit, N. The Potential Use of Peptides in the Fight against Chagas Disease and Leishmaniasis. Pharmaceutics 2024, 16, 227. https://doi.org/10.3390/pharmaceutics16020227
AMA Style
Berhe H, Kumar Cinthakunta Sridhar M, Zerihun M, Qvit N. The Potential Use of Peptides in the Fight against Chagas Disease and Leishmaniasis. Pharmaceutics. 2024; 16(2):227. https://doi.org/10.3390/pharmaceutics16020227
Chicago/Turabian StyleBerhe, Hayelom, Mahesh Kumar Cinthakunta Sridhar, Mulate Zerihun, and Nir Qvit. 2024. "The Potential Use of Peptides in the Fight against Chagas Disease and Leishmaniasis" Pharmaceutics 16, no. 2: 227. https://doi.org/10.3390/pharmaceutics16020227
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.