
Recent Advancement in mRNA Vaccine Development and Applications
 , , ,
, , ,
Abstract
:1. Introduction
2. Development of mRNA Vaccines
2.1. Types of mRNA Vaccines
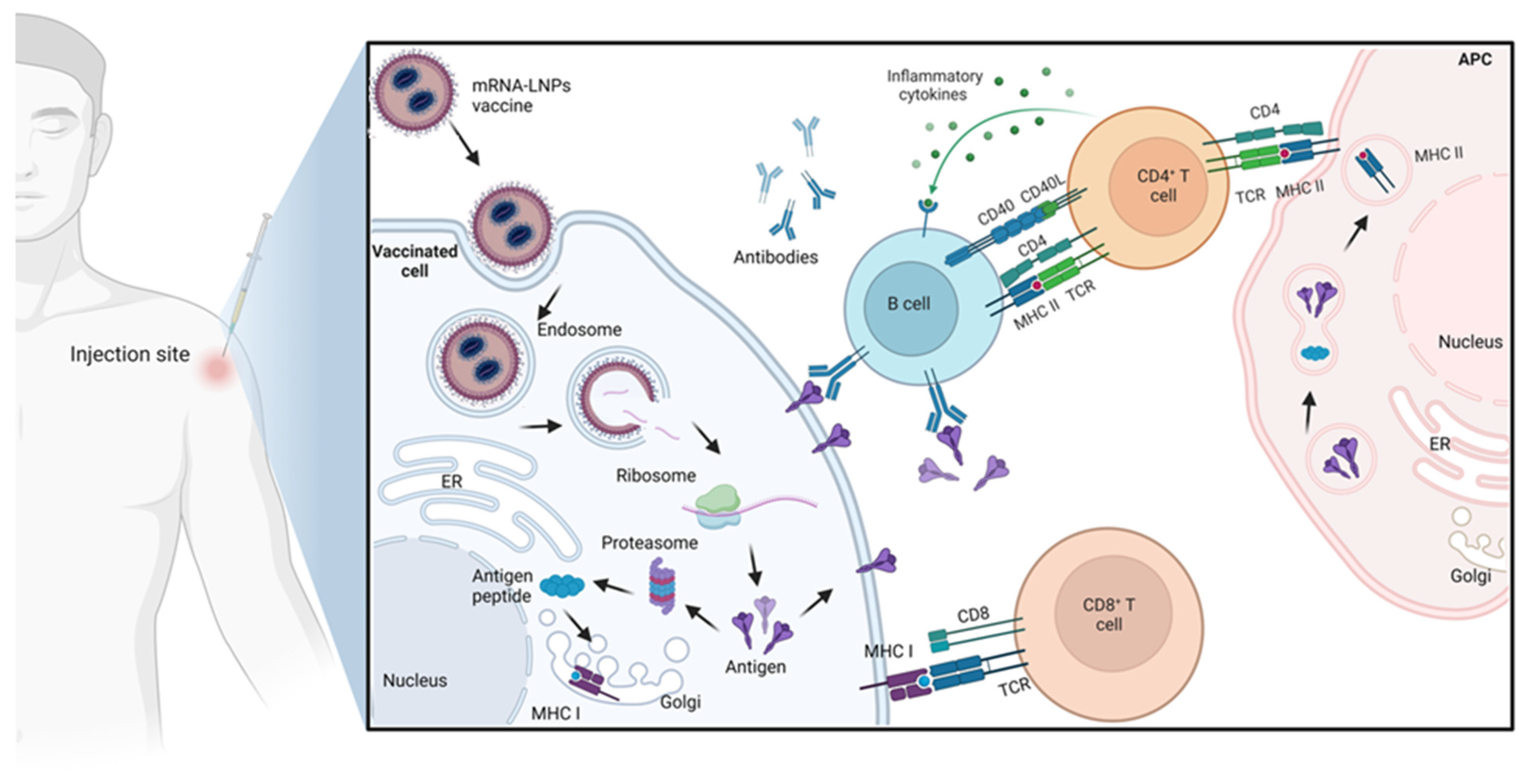
2.2. Mechanism of Action of mRNA Vaccines
2.3. Optimizing mRNA Vaccines
3. The mRNA Vaccine Delivery Systems
3.1. Polymeric Nanoparticles
3.2. Peptides and Proteins Nanoparticles
3.3. Protamine Nanoparticles
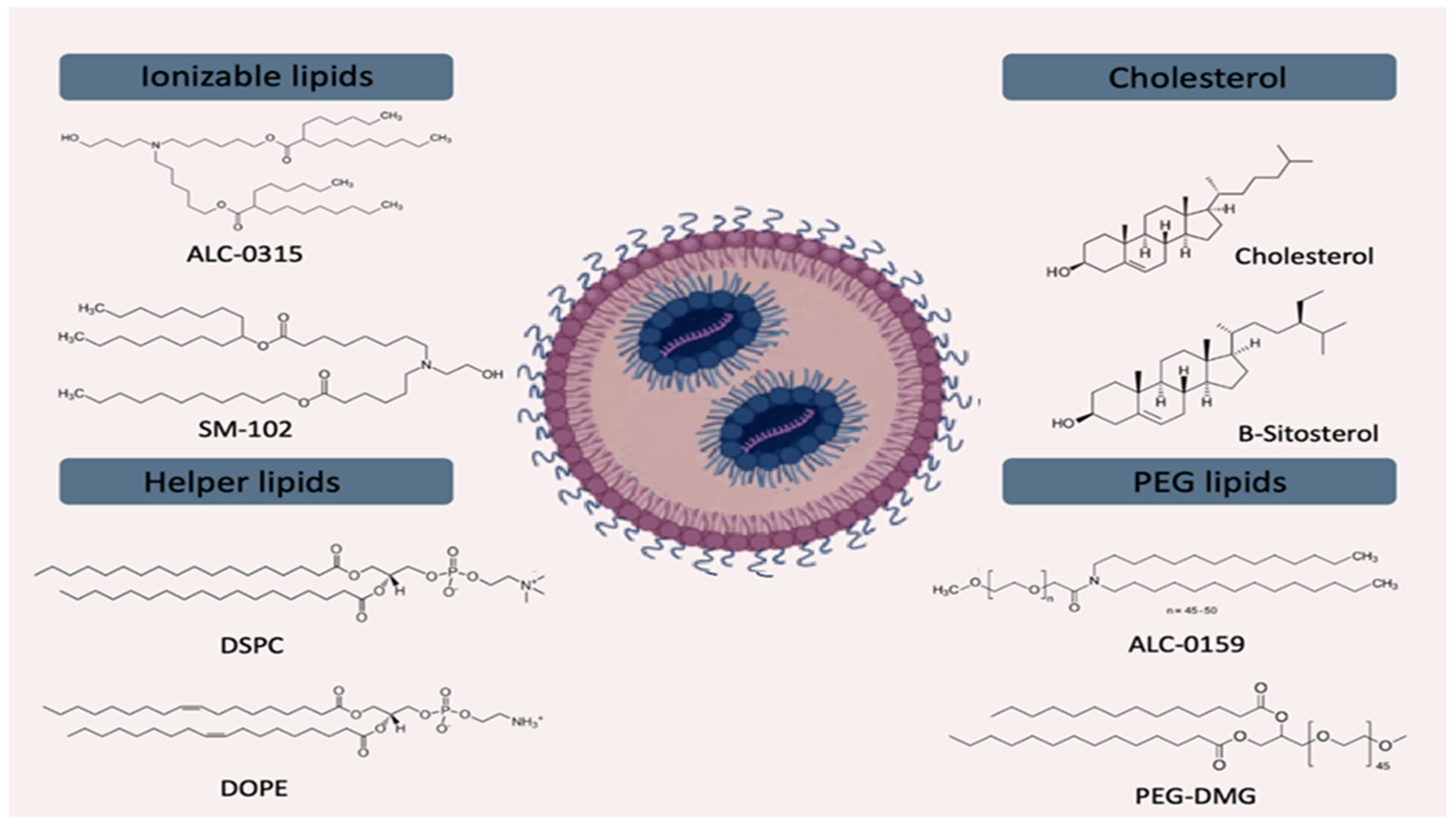
3.4. Lipid Nanoparticles (LNPs)
4. The mRNA Vaccine Applications
4.1. The mRNA Vaccines against Infectious Diseases
4.2. The mRNA Vaccines against Cancers
5. Clinical Trials of mRNA Vaccines

{kind=link}
{kind=link}
| Name of Product | ClinicalTrials.Gov Number | Payload | Disease | Phase | Route of Administration | Trial Status | Sponsor | Reference |
|---|---|---|---|---|---|---|---|---|
| Infectious diseases | ||||||||
| mRNA-1893 | NCT04917861 | Structural proteins of the Zika virus | Zika virus | 2 | I.M. | Active, recruiting | Moderna | [124] |
| mRNA-1647 | NCT04232280 | Six mRNA codings for pentamer viral antigen and gB protein of Cytomegalovirus | Cytomegalovirus infection | 2 | I.M. | Active, recruiting | Moderna | [125] |
| mRNA-1345 | NCT05127434 | The stabilized prefusion F protein | Respiratory syncytial virus | 2–3 | I.M. | Active, recruiting | Moderna | [126] |
| CVnCOV | NCT04652102 | SARS-CoV-2 | SARS-CoV-2 | 2–3 | I.M. | Active, not recruiting | CureVac | [127] |
| ARCT-021 | NCT04668339 | SARS-CoV-2 | SARS-CoV-2 | 2 | I.M. | Active, not recruiting | Arcturus | [128] |
| BNT162b2 | NCT04380701 | SARS-CoV-2 | SARS-CoV-2 | 1–2 | I.M. | Active, recruiting | BioNTech–Pfizer | [129] |
| mRNA-1273 | NCT04785144 | Codes for the full-length prefusion stabilized S protein of the SARS-CoV-2 B.1.351 variant. | SARS-CoV-2 B.1.351 variant | 2 | I.M. | Active, not recruiting | Moderna | [130] |
| Cancer | ||||||||
| BNT122 | NCT03815058 | 20 patient-specific neoantigens | Melanoma | 2 | I.V. | Active, recruiting | BioNTech– Genentech | [115] |
| mRNA-4157 | NCT03897881 | Encode up to 34 neoantigens | Melanoma | 2 | I.M. | Active, recruiting | Moderna–Merck | [51] |
| BNT111 | NCT04526899 | A mix of four melanoma-associated antigens | Melanoma | 2 | I.V. | Active, recruiting | BioNTech | [115] |
| CV9202 | NCT03164772 | NY-ESO-1, MAGE C1, MAGE C2, TPBG (5T4), survivin, MUC1 | Non-small cell lung cancer | 1–2 | I.D. | Active, not recruiting | CureVac, Ludwig Institute | [131] |
| BNT141 | NCT04683939 | Secreted IgG antibody | Cancer | 1–2 | I.V. | Not yet recruiting | BioNTech | [100] |
| BNT311/GEN1046 | NCT03917381 | Bispecific antibody PD-L1 × 4-1BB | Cancer | 1–2 | I.V. | Active, recruiting | Genmab–BioNTech | [121] |
| BNT312/GEN1042 | NCT04083599 | Bispecific antibody CD40 × 4-1BB | Solid tumors | 1–2 | I.V. | Active, recruiting | Genmab–BioNTech | [100] |
| BNT211 | NCT04503278 | CAR T for CLDN6 + tumor + mRNA for CLDN6 | CLDN6+ tumors | 1–2 | I.V. | Active, recruiting | BioNTech | [100] |
6. Future Applications of mRNA Vaccines
6.1. Future Consideration in mRNA Vaccines Development
6.1.1. Utilizing Different Types of mRNA
6.1.2. Ideal Delivery System for mRNA
6.1.3. Route of Administration and Storage Considerations of mRNA
6.2. Novel and Versatile Future Therapy of the mRNA Vaccines
6.2.1. The mRNA Vaccines for Infectious Diseases Prevention
6.2.2. The mRNA Vaccines for Cancer Therapy
6.2.3. The mRNA Vaccines for Genetic Disorders Therapy
6.2.4. The mRNA Vaccines in Regenerative Medicine
6.3. Advantages and Acceptance Criteria of mRNA Vaccines
6.3.1. Speed of Production
6.3.2. Scalability
6.3.3. Safety
6.3.4. Versatility
6.3.5. Effectiveness
6.4. Disadvantages and Challenges of mRNA Vaccines
6.4.1. The mRNA Stability
6.4.2. The mRNA Delivery
6.4.3. Accessibility and Acceptance
6.4.4. Variability
6.4.5. Immunogenicity
6.4.6. Variation among the Populations
6.4.7. Adverse Reactions
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Immunization Agenda 2030. Available online: https://www.who.int/teams/immunization-vaccines-and-biologicals/strategies/ia2030 (accessed on 23 May 2023).
- Plotkin, S.A. Vaccines: The Fourth Century. Clin. Vaccine Immunol. 2009, 16, 1709–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A. A Comprehensive Review of MRNA Vaccines. Int. J. Mol. Sci. 2023, 24, 2700. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Szuster-Ciesielska, A.; Dzieciątkowski, T.; Gwenzi, W.; Fal, A. MRNA Vaccines: The Future of Prevention of Viral Infections? J. Med. Virol. 2023, 95, e28572. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Publ. Group 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogan, M.J.; Pardi, N. MRNA Vaccines in the COVID-19 Pandemic and Beyond. Annu. Rev. Med. 2022, 73, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Hogan, M.J.; Weissman, D. ScienceDirect Recent Advances in MRNA Vaccine Technology. Curr. Opin. Immunol. 2020, 65, 14–20. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. New Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. New Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. MRNA-Based Therapeutics—Developing a New Class of Drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Wojtczak, B.A.; Sikorski, P.J.; Fac-Dabrowska, K.; Nowicka, A.; Warminski, M.; Kubacka, D.; Nowak, E.; Nowotny, M.; Kowalska, J.; Jemielity, J. 5′-Phosphorothiolate Dinucleotide Cap Analogues: Reagents for Messenger RNA Modification and Potent Small-Molecular Inhibitors of Decapping Enzymes. J. Am. Chem. Soc. 2018, 140, 5987–5999. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Luo, X.; Dong, Y. Effects of Chemically Modified Messenger RNA on Protein Expression. Available online: https://pubs.acs.org/doi/pdf/10.1021/acs.bioconjchem.6b00090 (accessed on 23 May 2023).
- Svitkin, Y.V.; Cheng, Y.M.; Chakraborty, T.; Presnyak, V.; John, M.; Sonenberg, N. N1-Methyl-Pseudouridine in MRNA Enhances Translation through EIF2α-Dependent and Independent Mechanisms by Increasing Ribosome Density. Nucleic Acids Res. 2017, 45, 6023–6036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the Delivery of RNA Therapeutics: From Concept to Clinical Reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsui, N.B.; Ng, E.K.; Lo, Y.D. Stability of Endogenous and Added RNA in Blood Specimens, Serum, and Plasma. Clin. Chem. 2002, 48, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-Viral Vectors for Gene-Based Therapy. Nat. Rev. Genet. 2014, 15, 541. [Google Scholar] [CrossRef] [PubMed]
- Cullis, P.R.; Hope, M.J. Lipid Nanoparticle Systems for Enabling Gene Therapies. Mol. Ther. 2017, 25, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, S.; Ranjan, S.; Tathagata, S. Delivery Strategies for MRNA Vaccines. Pharm. Med. 2022, 36, 11–20. [Google Scholar] [CrossRef]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic MRNA Delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika Virus Protection by a Single Low-Dose Nucleoside-Modified MRNA Vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Secreto, A.J.; Shan, X.; Debonera, F.; Glover, J.; Yi, Y.; Muramatsu, H.; Ni, H.; Mui, B.L.; Tam, Y.K.; et al. Administration of Nucleoside-Modified MRNA Encoding Broadly Neutralizing Antibody Protects Humanized Mice from HIV-1 Challenge. Nat. Commun. 2017, 8, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Ishihara, H. Difference in the Lipid Nanoparticle Technology Employed in Three Approved SiRNA (Patisiran) and MRNA (COVID-19 Vaccine) Drugs. Drug Metab. Pharmacokinet. 2021, 41, 100424. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Mixich, L.; Boonstra, E.; Cabral, H. Polymer-Based MRNA Delivery Strategies for Advanced Therapies. Adv. Healthc. Mater. 2023, 12, 2202688. [Google Scholar] [CrossRef] [PubMed]
- Udhayakumar, V.K.; De Beuckelaer, A.; McCaffrey, J.; McCrudden, C.M.; Kirschman, J.L.; Vanover, D.; Van Hoecke, L.; Roose, K.; Deswarte, K.; De Geest, B.G.; et al. Arginine-Rich Peptide-Based MRNA Nanocomplexes Efficiently Instigate Cytotoxic T Cell Immunity Dependent on the Amphipathic Organization of the Peptide. Adv. Healthc. Mater. 2017, 6, 1601412. [Google Scholar] [CrossRef] [PubMed]
- Farshi, E. Peptide-Based MRNA Vaccines. J. Gastro. Hepato. 2023, 9, 1–6. [Google Scholar]
- Jarzebska, N.T.; Mellett, M.; Frei, J.; Kündig, T.M.; Pascolo, S. Protamine-Based Strategies for RNA Transfection. Pharmaceutics 2021, 13, 877. [Google Scholar] [CrossRef]
- Ruseska, I.; Fresacher, K.; Petschacher, C.; Zimmer, A. Use of Protamine in Nanopharmaceuticals—A Review. Nanomaterials 2021, 11, 1508. [Google Scholar] [CrossRef]
- Oberli, M.A.; Reichmuth, A.M.; Dorkin, J.R.; Mitchell, M.J.; Fenton, O.S.; Jaklenec, A.; Anderson, D.G.; Langer, R.; Blankschtein, D. Lipid Nanoparticle Assisted MRNA Delivery for Potent Cancer Immunotherapy. Nano Lett. 2017, 17, 1326–1335. [Google Scholar] [CrossRef]
- Chen, J.; Ye, Z.; Huang, C.; Qiu, M.; Song, D.; Li, Y.; Xu, Q. Lipid Nanoparticle-Mediated Lymph Node–Targeting Delivery of MRNA Cancer Vaccine Elicits Robust CD8+ T Cell Response. Proc. Natl. Acad. Sci. 2022, 119, e2207841119. [Google Scholar] [CrossRef]
- Sheridan, C. MRNA Printers Kick-Start Personalized Medicines for All. Nat. Biotechnol. 2022, 40, 1160–1162. [Google Scholar] [CrossRef]
- Semple, S.C.; Leone, R.; Barbosa, C.J.; Tam, Y.K.; Lin, P.J.C. Lipid Nanoparticle Delivery Systems to Enable MRNA-Based Therapeutics. Pharmaceutics 2022, 14, 398. [Google Scholar] [CrossRef]
- Pardi, N.; Weissman, D. Chapter 6 Nucleoside Modified MRNA Vaccines for Infectious Diseases. RNA Vaccines Methods Protoc. 2017, 1499, 109–121. [Google Scholar]
- Kutzler, M.A.; Weiner, D.B. DNA Vaccines: Ready for Prime Time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; van den Berg, F.; Arbuthnot, P. Self-Amplifying RNA Vaccines for Infectious Diseases. Gene Ther. 2021, 28, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Self-Replicating RNA Viruses for RNA Therapeutics. Molecules 2018, 23, 3310. [Google Scholar] [CrossRef] [Green Version]
- Tews, B.A.; Meyers, G. Self-Replicating RNA. Methods Mol. Biol. 2017, 1499, 15–35. [Google Scholar]
- George, J.; Raju, R. Alphavirus RNA Genome Repair and Evolution: Molecular Characterization of Infectious Sindbis Virus Isolates Lacking a Known Conserved Motif at the 3′ End of the Genome. J. Virol. 2000, 74, 9776–9785. [Google Scholar] [CrossRef] [Green Version]
- Papukashvili, D.; Rcheulishvili, N.; Liu, C.; Ji, Y.; He, Y.; Wang, P.G. Self-Amplifying RNA Approach for Protein Replacement Therapy. Int. J. Mol. Sci. 2022, 23, 12884. [Google Scholar] [CrossRef]
- Iavarone, C.; O’hagan, D.T.; Yu, D.; Delahaye, N.F.; Ulmer, J.B. Mechanism of Action of MRNA-Based Vaccines. Expert Rev. Vaccines 2017, 16, 871–881. [Google Scholar] [CrossRef]
- Desterro, J.; Bak-Gordon, P.; Carmo-Fonseca, M. Targeting MRNA Processing as an Anticancer Strategy. Nat. Rev. Drug Discov. 2020, 19, 112–129. [Google Scholar] [CrossRef]
- Fabbri, L.; Chakraborty, A.; Robert, C.; Vagner, S. The Plasticity of MRNA Translation during Cancer Progression and Therapy Resistance. Nat. Rev. Cancer 2021, 21, 558–577. [Google Scholar] [CrossRef]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. MRNA-Based Therapeutics: Powerful and Versatile Tools to Combat Diseases. Sig. Transduct. Target Ther. 2022, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Kormann, M.S.D.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A.; et al. Expression of Therapeutic Proteins after Delivery of Chemically Modified MRNA in Mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef]
- Jarallah, S.J.; Aldossary, A.M.; Tawfik, E.A.; Altamimi, R.M.; Alsharif, W.K.; Alzahrani, N.M.; As Sobeai, H.M.; Qamar, W.; Alfahad, A.J.; Alshabibi, M.A.; et al. GL67 Lipid-Based Liposomal Formulation for Efficient SiRNA Delivery into Human Lung Cancer Cells. Saudi Pharm. J. 2023, 31, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Pijlman, G.P. Alphavirus Infection: Host Cell Shut-Off and Inhibition of Antiviral Responses. Viruses 2016, 8, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götte, B.; Liu, L.; McInerney, G.M. The Enigmatic Alphavirus Non-Structural Protein 3 (NsP3) Revealing Its Secrets at Last. Viruses 2018, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-Assembled MRNA Vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112. [Google Scholar] [CrossRef]
- Fessel, J. A Vaccine to Prevent Initial Loss of Cognition and Eventual Alzheimer’s Disease in Elderly Persons. Alzheimers Dement. 2021, 7, e12126. [Google Scholar] [CrossRef]
- Kwon, H.; Kim, M.; Seo, Y.; Moon, Y.S.; Lee, H.J.; Lee, K.; Lee, H. Emergence of Synthetic MRNA: In Vitro Synthesis of MRNA and Its Applications in Regenerative Medicine. Biomaterials 2018, 156, 172–193. [Google Scholar] [CrossRef]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of MRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Gallie, D.R. The Cap and Poly(A) Tail Function Synergistically to Regulate MRNA Translational Efficiency. Genes Dev. 1991, 5, 2108–2116. [Google Scholar] [CrossRef] [Green Version]
- Jemielity, J.; Fowler, T.; Zuberek, J.; Stepinski, J.; Lewdorowicz, M.; Niedzwiecka, A.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Novel “Anti-Reverse” Cap Analogs with Superior Translational Properties. RNA 2003, 9, 1108–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, A.N.; Diken, M.; Kreiter, S.; Selmi, A.; Kowalska, J.; Jemielity, J.; Darzynkiewicz, E.; Huber, C.; Türeci, Ö.; Sahin, U. Phosphorothioate Cap Analogs Increase Stability and Translational Efficiency of RNA Vaccines in Immature Dendritic Cells and Induce Superior Immune Responses in Vivo. Gene Ther. 2010, 17, 961–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine into MRNA Yields Superior Nonimmunogenic Vector with Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Poliskey, J.A.; Crowley, S.T.; Ramanathan, R.; White, C.W.; Mathew, B.; Rice, K.G. Metabolically Stabilized Double Stranded MRNA Polyplexes. Gene Ther. 2018, 25, 473–484. [Google Scholar] [CrossRef]
- Tan, L.; Zheng, T.; Li, M.; Zhong, X.; Tang, Y.; Qin, M.; Sun, X. Optimization of an MRNA Vaccine Assisted with Cyclodextrin-Polyethyleneimine Conjugates. Drug Deliv. Transl. Res. 2020, 10, 678–689. [Google Scholar] [CrossRef]
- Kaczmarek, J.C.; Kauffman, K.J.; Fenton, O.S.; Sadtler, K.; Patel, A.K.; Heartlein, M.W.; DeRosa, F.; Anderson, D.G. Optimization of a Degradable Polymer-Lipid Nanoparticle for Potent Systemic Delivery of MRNA to the Lung Endothelium and Immune Cells. Nano Lett. 2018, 18, 6449–6454. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Nguyen, L.H.; Miller, J.B.; Yan, Y.; Kos, P.; Xiong, H.; Li, L.; Hao, J.; Minnig, J.T.; Zhu, H.; et al. Modular Degradable Dendrimers Enable Small RNAs to Extend Survival in an Aggressive Liver Cancer Model. Proc. Natl. Acad. Sci. USA 2016, 113, 520–525. [Google Scholar] [CrossRef]
- Ulkoski, D.; Bak, A.; Wilson, J.T.; Krishnamurthy, V.R. Recent Advances in Polymeric Materials for the Delivery of RNA Therapeutics. Expert Opin. Drug Deliv. 2019, 16, 1149–1167. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.; Kim, E.H.; Jang, H.; Jang, Y.; Chi, S.-G.; Yang, Y.; Kim, S.H. The Potential of Cell-Penetrating Peptides for MRNA Delivery to Cancer Cells. Pharmaceutics 2022, 14, 1271. [Google Scholar] [CrossRef]
- Chintakunta, R.; Buaron, N.; Kahn, N.; Moriah, A.; Lifshiz, R.; Goldbart, R.; Traitel, T.; Tyler, B.; Brem, H.; Kost, J. Synthesis, Characterization, and Self-Assembly with Plasmid DNA of a Quaternary Ammonium Derivative of Pectic Galactan and Its Fluorescent Labeling for Bioimaging Applications. Carbohydr. Polym. 2016, 150, 308–318. [Google Scholar] [CrossRef]
- Solomun, J.I.; Cinar, G.; Mapfumo, P.; Richter, F.; Moek, E.; Hausig, F.; Martin, L.; Hoeppener, S.; Nischang, I.; Traeger, A. Solely Aqueous Formulation of Hydrophobic Cationic Polymers for Efficient Gene Delivery. Int. J. Pharm. 2021, 593, 120080. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, M.; Zhang, Z.; Gong, T.; Sun, X. Induction of HIV-1 Gag Specific Immune Responses by Cationic Micelles Mediated Delivery of Gag MRNA. Drug Deliv. 2016, 23, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, M.; Fu, Y.; Li, Y.; Gong, T.; Zhang, Z.; Sun, X. Enhanced Intranasal Delivery of MRNA Vaccine by Overcoming the Nasal Epithelial Barrier via Intra- and Paracellular Pathways. J. Control Release 2016, 228, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Lee, T.-J.; Yang, G.-M.; Oh, J.; Won, J.; Han, J.; Jeong, G.-J.; Kim, J.; Kim, J.-H.; Kim, B.-S.; et al. Efficient MRNA Delivery with Graphene Oxide-Polyethylenimine for Generation of Footprint-Free Human Induced Pluripotent Stem Cells. J. Control Release 2016, 235, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Maassen, S.J.; van der Schoot, P.; Cornelissen, J.J.L.M. Experimental and Theoretical Determination of the PH inside the Confinement of a Virus-Like Particle. Small 2018, 14, 1802081. [Google Scholar] [CrossRef]
- Thaxton, C.S.; Daniel, W.L.; Giljohann, D.A.; Thomas, A.D.; Mirkin, C.A. Templated Spherical High Density Lipoprotein Nanoparticles. J. Am. Chem. Soc. 2009, 131, 1384–1385. [Google Scholar] [CrossRef] [Green Version]
- Crone, M.A.; Priestman, M.; Ciechonska, M.; Jensen, K.; Sharp, D.J.; Anand, A.; Randell, P.; Storch, M.; Freemont, P.S. A Role for Biofoundries in Rapid Development and Validation of Automated SARS-CoV-2 Clinical Diagnostics. Nat. Commun. 2020, 11, 4464. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, X.; Bian, Y.; Wang, S.; Chai, Q.; Guo, Z.; Wang, Z.; Zhu, P.; Peng, H.; Yan, X.; et al. Dual-Targeting Nanoparticle Vaccine Elicits a Therapeutic Antibody Response against Chronic Hepatitis, B. Nat. Nanotechnol. 2020, 15, 406–416. [Google Scholar] [CrossRef]
- McCarthy, H.O.; McCaffrey, J.; McCrudden, C.M.; Zholobenko, A.; Ali, A.A.; McBride, J.W.; Massey, A.S.; Pentlavalli, S.; Chen, K.-H.; Cole, G.; et al. Development and Characterization of Self-Assembling Nanoparticles Using a Bio-Inspired Amphipathic Peptide for Gene Delivery. J. Control Release 2014, 189, 141–149. [Google Scholar] [CrossRef]
- Hoerr, I.; Obst, R.; Rammensee, H.G.; Jung, G. In Vivo Application of RNA Leads to Induction of Specific Cytotoxic T Lymphocytes and Antibodies. Eur. J. Immunol. 2000, 30, 1–7. [Google Scholar] [CrossRef]
- Karabasz, A.; Bzowska, M.; Szczepanowicz, K. Biomedical Applications of Multifunctional Polymeric Nanocarriers: A Review of Current Literature. Int. J. Nanomedicine 2020, 15, 8673–8696. [Google Scholar] [CrossRef]
- Scheel, B.; Aulwurm, S.; Probst, J.; Stitz, L.; Hoerr, I.; Rammensee, H.-G.; Weller, M.; Pascolo, S. Therapeutic Anti-Tumor Immunity Triggered by Injections of Immunostimulating Single-Stranded RNA. Eur. J. Immunol. 2006, 36, 2807–2816. [Google Scholar] [CrossRef] [PubMed]
- Scheel, B.; Braedel, S.; Probst, J.; Carralot, J.-P.; Wagner, H.; Schild, H.; Jung, G.; Rammensee, H.-G.; Pascolo, S. Immunostimulating Capacities of Stabilized RNA Molecules. Eur. J. Immunol. 2004, 34, 537–547. [Google Scholar] [CrossRef]
- Kallen, K.-J.; Heidenreich, R.; Schnee, M.; Petsch, B.; Schlake, T.; Thess, A.; Baumhof, P.; Scheel, B.; Koch, S.D.; Fotin-Mleczek, M. A Novel, Disruptive Vaccination Technology. Hum. Vaccin Immunother. 2013, 9, 2263–2276. [Google Scholar] [CrossRef] [Green Version]
- Schnee, M.; Vogel, A.B.; Voss, D.; Petsch, B.; Baumhof, P.; Kramps, T.; Stitz, L. An MRNA Vaccine Encoding Rabies Virus Glycoprotein Induces Protection against Lethal Infection in Mice and Correlates of Protection in Adult and Newborn Pigs. PLoS Negl. Trop. Dis. 2016, 10, e0004746. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid Nanoparticles for MRNA Delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef]
- Buck, J.; Grossen, P.; Cullis, P.R.; Huwyler, J.; Witzigmann, D. Lipid-Based DNA Therapeutics: Hallmarks of Non-Viral Gene Delivery. ACS Nano 2019, 13, 3754–3782. [Google Scholar] [CrossRef] [PubMed]
- Malone, R.W.; Felgner, P.L.; Verma, I.M. Cationic Liposome-Mediated RNA Transfection. Proc. Natl. Acad. Sci. USA 1989, 86, 6077–6081. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, K.J.; Webber, M.J.; Anderson, D.G. Materials for Non-Viral Intracellular Delivery of Messenger RNA Therapeutics. J. Control Release 2016, 240, 227–234. [Google Scholar] [CrossRef]
- Hajj, K.A.; Ball, R.L.; Deluty, S.B.; Singh, S.R.; Strelkova, D.; Knapp, C.M.; Whitehead, K.A. Branched-Tail Lipid Nanoparticles Potently Deliver MRNA In Vivo Due to Enhanced Ionization at Endosomal PH. Small 2019, 15, e1805097. [Google Scholar] [CrossRef]
- Gómez-Aguado, I.; Rodríguez-Castejón, J.; Vicente-Pascual, M.; Rodríguez-Gascón, A.; Solinís, M.Á.; Del Pozo-Rodríguez, A. Nanomedicines to Deliver MRNA: State of the Art and Future Perspectives. Nanomaterials 2020, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Kim, J.; Herrera, M.; Mukherjee, A.; Kabanov, A.; Sahay, G. Brief Update on Endocytosis of Nanomedicines. Adv. Drug Deliv. Rev. 2019, 144, 90–111. [Google Scholar] [CrossRef]
- Semple, S.C.; Klimuk, S.K.; Harasym, T.O.; Dos Santos, N.; Ansell, S.M.; Wong, K.F.; Maurer, N.; Stark, H.; Cullis, P.R.; Hope, M.J.; et al. Efficient Encapsulation of Antisense Oligonucleotides in Lipid Vesicles Using Ionizable Aminolipids: Formation of Novel Small Multilamellar Vesicle Structures. Biochim. Biophys. Acta Biomembr. 2001, 1510, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Veiga, N.; Goldsmith, M.; Granot, Y.; Rosenblum, D.; Dammes, N.; Kedmi, R.; Ramishetti, S.; Peer, D. Cell Specific Delivery of Modified MRNA Expressing Therapeutic Proteins to Leukocytes. Nat. Commun. 2018, 9, 4493. [Google Scholar] [CrossRef] [Green Version]
- Hajj, K.A.; Melamed, J.R.; Chaudhary, N.; Lamson, N.G.; Ball, R.L.; Yerneni, S.S.; Whitehead, K.A. A Potent Branched-Tail Lipid Nanoparticle Enables Multiplexed MRNA Delivery and Gene Editing In Vivo. Nano Lett. 2020, 20, 5167–5175. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Tonnu, N.; Tachikawa, K.; Limphong, P.; Vega, J.B.; Karmali, P.P.; Chivukula, P.; Verma, I.M. Systemic Delivery of Factor IX Messenger RNA for Protein Replacement Therapy. Proc. Natl. Acad. Sci. USA 2017, 114, E1941–E1950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, J.; Li, M.; Jin, X. Modified MRNA-LNP Vaccines Confer Protection against Experimental DENV-2 Infection in Mice. Mol. Ther. Methods Clin. Dev. 2020, 18, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Ilhan-Ayisigi, E.; Yaldiz, B.; Bor, G.; Yaghmur, A.; Yesil-Celiktas, O. Advances in Microfluidic Synthesis and Coupling with Synchrotron SAXS for Continuous Production and Real-Time Structural Characterization of Nano-Self-Assemblies. Colloids Surf. B Biointerfaces 2021, 201, 111633. [Google Scholar] [CrossRef] [PubMed]
- Tomeh, M.A.; Zhao, X. Recent Advances in Microfluidics for the Preparation of Drug and Gene Delivery Systems. Mol. Pharm. 2020, 17, 4421–4434. [Google Scholar] [CrossRef]
- Terada, T.; Kulkarni, J.A.; Huynh, A.; Chen, S.; van der Meel, R.; Tam, Y.Y.C.; Cullis, P.R. Characterization of Lipid Nanoparticles Containing Ionizable Cationic Lipids Using Design-of-Experiments Approach. Langmuir 2021, 37, 1120–1128. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell Entry Mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernán, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 MRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
- Thompson, M.G.; Burgess, J.L.; Naleway, A.L.; Tyner, H.L.; Yoon, S.K.; Meece, J.; Olsho, L.E.W.; Caban-Martinez, A.J.; Fowlkes, A.; Lutrick, K.; et al. Interim Estimates of Vaccine Effectiveness of BNT162b2 and MRNA-1273 COVID-19 Vaccines in Preventing SARS-CoV-2 Infection Among Health Care Personnel, First Responders, and Other Essential and Frontline Workers—Eight U.S. Locations, December 2020-March 2021. MMWR Morb. Mortal Wkly. Rep. 2021, 70, 495–500. [Google Scholar]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. MRNA Vaccines for Infectious Diseases: Principles, Delivery and Clinical Translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Arevalo, C.P.; Bolton, M.J.; Le Sage, V.; Ye, N.; Furey, C.; Muramatsu, H.; Alameh, M.-G.; Pardi, N.; Drapeau, E.M.; Parkhouse, K.; et al. A Multivalent Nucleoside-Modified MRNA Vaccine against All Known Influenza Virus Subtypes. Science 2022, 378, 899–904. [Google Scholar] [CrossRef]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. MRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. IJMS 2020, 21, 6582. [Google Scholar] [CrossRef]
- Sahin, U.; Oehm, P.; Derhovanessian, E.; Jabulowsky, R.A.; Vormehr, M.; Gold, M.; Maurus, D.; Schwarck-Kokarakis, D.; Kuhn, A.N.; Omokoko, T.; et al. An RNA Vaccine Drives Immunity in Checkpoint-Inhibitor-Treated Melanoma. Nature 2020, 585, 107–112. [Google Scholar] [CrossRef]
- Barbier, A.J.; Jiang, A.Y.; Zhang, P.; Wooster, R.; Anderson, D.G. The Clinical Progress of MRNA Vaccines and Immunotherapies. Nat. Biotechnol. 2022, 40, 840–854. [Google Scholar] [CrossRef]
- Lorentzen, C.L.; Haanen, J.B.; Met, Ö.; Svane, I.M. Clinical Advances and Ongoing Trials of MRNA Vaccines for Cancer Treatment. Lancet Oncol. 2022, 23, e450–e458. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Duan, J.; Yang, N.; Yang, Y.; Zhao, X. MRNA Vaccines in the Prevention and Treatment of Diseases. MedComm 2022, 3, e167. [Google Scholar] [CrossRef] [PubMed]
- Poland, G.A.; Kennedy, R.B.; Ovsyannikova, I.G.; Palacios, R.; Ho, P.L.; Kalil, J. Development of Vaccines against Zika Virus. Lancet Infect. Dis. 2018, 18, e211–e219. [Google Scholar] [CrossRef] [Green Version]
- Dowd, K.A.; DeMaso, C.R.; Pelc, R.S.; Speer, S.D.; Smith, A.R.Y.; Goo, L.; Platt, D.J.; Mascola, J.R.; Graham, B.S.; Mulligan, M.J.; et al. Broadly Neutralizing Activity of Zika Virus-Immune Sera Identifies a Single Viral Serotype. Cell Rep. 2016, 16, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- VanBlargan, L.A.; Himansu, S.; Foreman, B.M.; Ebel, G.D.; Pierson, T.C.; Diamond, M.S. An MRNA Vaccine Protects Mice against Multiple Tick-Transmitted Flavivirus Infections. Cell Rep. 2018, 25, 3382–3392.e3. [Google Scholar] [CrossRef] [Green Version]
- Richner, J.M.; Himansu, S.; Dowd, K.A.; Butler, S.; Salazar, V.; Fox, J.M.; Julander, J.G.; Tang, W.W.; Shresta, S.; Pierson, T.C.; et al. Modified MRNA Vaccines Protect against Zika Virus Infection. Cell 2017, 168, 1114. [Google Scholar] [CrossRef] [Green Version]
- Luisi, K.; Morabito, K.M.; Burgomaster, K.E.; Sharma, M.; Kong, W.-P.; Foreman, B.M.; Patel, S.; Fisher, B.; Aleshnick, M.A.; Laliberte, J.; et al. Development of a Potent Zika Virus Vaccine Using Self-Amplifying Messenger RNA. Sci. Adv. 2020, 6, eaba5068. [Google Scholar] [CrossRef]
- Wollner, C.J.; Richner, J.M. MRNA Vaccines against Flaviviruses. Vaccines 2021, 9, 148. [Google Scholar] [CrossRef]
- Schlake, T.; Thess, A.; Thran, M.; Jordan, I. MRNA as Novel Technology for Passive Immunotherapy. Cell Mol. Life Sci. 2019, 76, 301–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, N.A.C.; Kester, K.E.; Casimiro, D.; Gurunathan, S.; DeRosa, F. The Promise of MRNA Vaccines: A Biotech and Industrial Perspective. NPJ Vaccines 2020, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Espeseth, A.S.; Cejas, P.J.; Citron, M.P.; Wang, D.; DiStefano, D.J.; Callahan, C.; Donnell, G.O.; Galli, J.D.; Swoyer, R.; Touch, S.; et al. Modified MRNA/Lipid Nanoparticle-Based Vaccines Expressing Respiratory Syncytial Virus F Protein Variants Are Immunogenic and Protective in Rodent Models of RSV Infection. NPJ Vaccines 2020, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Hajiaghapour Asr, M.; Dayani, F.; Saedi Segherloo, F.; Kamedi, A.; Neill, A.O.; MacLoughlin, R.; Doroudian, M. Lipid Nanoparticles as Promising Carriers for MRNA Vaccines for Viral Lung Infections. Pharmaceutics 2023, 15, 1127. [Google Scholar] [CrossRef] [PubMed]
- Kremsner, P.; Mann, P.; Bosch, J.; Fendel, R.; Gabor, J.J.; Kreidenweiss, A.; Kroidl, A.; Leroux-Roels, I.; Leroux-Roels, G.; Schindler, C.; et al. Phase 1 Assessment of the Safety and Immunogenicity of an MRNA- Lipid Nanoparticle Vaccine Candidate Against SARS-CoV-2 in Human Volunteers. MedRxiv 2020. [Google Scholar] [CrossRef]
- de Alwis, R.; Gan, E.S.; Chen, S.; Leong, Y.S.; Tan, H.C.; Zhang, S.L.; Yau, C.; Low, J.G.H.; Kalimuddin, S.; Matsuda, D.; et al. A Single Dose of Self-Transcribing and Replicating RNA-Based SARS-CoV-2 Vaccine Produces Protective Adaptive Immunity in Mice. Mol. Ther. 2021, 29, 1970–1983. [Google Scholar] [CrossRef] [PubMed]
- SEC Filing | BioNTech. Available online: https://investors.biontech.de/node/8126/html (accessed on 5 June 2023).
- BioNTech. Next Generation Immunotherapy. Available online: https://investors.biontech.de/static-files/057be080-d0bd-4b58-8ad5-6acb6f7e421b (accessed on 5 June 2023).
- Financials—SEC Filings—SEC Filings Details. Available online: https://investors.modernatx.com/financials/sec-filings/sec-filings-details/default.aspx?FilingId=13959984 (accessed on 5 June 2023).
- Khattak, A.; Carlino, M.; Meniawy, T.; Ansstas, G.; Medina, T.; Taylor, M.H.; Kim, K.B.; McKean, M.; Long, G.V.; Sullivan, R.J.; et al. A Personalized Cancer Vaccine, MRNA-4157 (V940), Combined With Pembrolizumab Versus Pembrolizumab Alone in Patients With Resected High-Risk Melanoma: Efficacy and Safety Results From the Randomized, Open-Label Phase 2 MRNA-4157-P201/KEYNOTE-942 Trial. CancerRes. 2023, 83, CT001. [Google Scholar] [CrossRef]
- Papachristofilou, A.; Hipp, M.M.; Klinkhardt, U.; Früh, M.; Sebastian, M.; Weiss, C.; Pless, M.; Cathomas, R.; Hilbe, W.; Pall, G.; et al. Phase Ib Evaluation of a Self-Adjuvanted Protamine Formulated MRNA-Based Active Cancer Immunotherapy, BI1361849 (CV9202), Combined with Local Radiation Treatment in Patients with Stage IV Non-Small Cell Lung Cancer. J. Immunother. Cancer 2019, 7, 38. [Google Scholar] [CrossRef]
- Chen, J.; Xu, Z.; Hu, C.; Zhang, S.; Zi, M.; Yuan, L.; Cheng, X. Targeting CLDN18.2 in Cancers of the Gastrointestinal Tract: New Drugs and New Indications. Front. Oncol. 2023, 13, 1132319. [Google Scholar] [CrossRef]
- Muik, A.; Garralda, E.; Altintas, I.; Gieseke, F.; Geva, R.; Ben-Ami, E.; Maurice-Dror, C.; Calvo, E.; LoRusso, P.M.; Alonso, G.; et al. Preclinical Characterization and Phase I Trial Results of a Bispecific Antibody Targeting PD-L1 and 4-1BB (GEN1046) in Patients with Advanced Refractory Solid Tumors. Cancer Discov. 2022, 12, 1248–1265. [Google Scholar] [CrossRef]
- Muik, A.; Adams 3rd, H.C.; Gieseke, F.; Altintas, I.; Schoedel, K.B.; Blum, J.M.; Sänger, B.; Burm, S.M.; Stanganello, E.; Verzijl, D.; et al. DuoBody-CD40x4-1BB Induces Dendritic-Cell Maturation and Enhances T-Cell Activation through Conditional CD40 and 4-1BB Agonist Activity. J. Immunother. Cancer 2022, 10, e004322. [Google Scholar] [CrossRef]
- Mackensen, A.; Haanen, J.B.A.G.; Koenecke, C.; Alsdorf, W.; Wagner-Drouet, E.; Heudobler, D.; Borchmann, P.; Bokemeyer, C.; Klobuch, S.; Smit, E.; et al. LBA38 BNT211-01: A Phase I Trial to Evaluate Safety and Efficacy of CLDN6 CAR T Cells and CLDN6-Encoding MRNA Vaccine-Mediated in Vivo Expansion in Patients with CLDN6-Positive Advanced Solid Tumours. Ann. Oncol. 2022, 33, S1404–S1405. [Google Scholar] [CrossRef]
- Moderna Reports Second Quarter Fiscal Year 2021 Financial Results and Provides Business Updates. Available online: https://investors.modernatx.com/news/news-details/2021/Moderna-Reports-Second-Quarter-Fiscal-Year-2021-Financial-Results-and-Provides-Business-Updates/default.aspx (accessed on 5 June 2023).
- Moderna. Annual R&D Day R&D Day. Available online: https://investors.modernatx.com/events-and-presentations/events/event-details/2020/RD-Day/default.aspx (accessed on 5 June 2023).
- Moderna Announces Advances Across Its Industry-Leading mRNA Pipeline and Provides Business Update. Available online: https://investors.modernatx.com/news/news-details/2022/Moderna-Announces-Advances-Across-Its-Industry-Leading-mRNA-Pipeline-and-Provides-Business-Update/default.aspx (accessed on 5 June 2023).
- Cohen, J. What Went Wrong with CureVac’s MRNA Vaccine? Science 2021, 372, 1381. [Google Scholar] [CrossRef]
- Low, J.G.; de Alwis, R.; Chen, S.; Kalimuddin, S.; Leong, Y.S.; Mah, T.K.L.; Yuen, N.; Tan, H.C.; Zhang, S.L.; Sim, J.X.Y.; et al. A Phase I/II Randomized, Double-Blinded, Placebo-Controlled Trial of a Self-Amplifying Covid-19 MRNA Vaccine. NPJ Vaccines 2022, 7, 161. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II Study of COVID-19 RNA Vaccine BNT162b1 in Adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.; Koch, M.; Wu, K.; Chu, L.; Ma, L.; Hill, A.; Nunna, N.; Huang, W.; Oestreicher, J.; Colpitts, T.; et al. Safety and Immunogenicity of SARS-CoV-2 Variant MRNA Vaccine Boosters in Healthy Adults: An Interim Analysis. Nat. Med. 2021, 27, 2025–2031. [Google Scholar] [CrossRef]
- Gandhi, L.; Aufiero Ramirez, K.; Schwarzenberger, P.; Ricciardi, T.; Macri, M.J.; Ryan, A.; Venhaus, R.R. Phase 1/2 Study of MRNA Vaccine Therapy + Durvalumab (Durva) ± Tremelimumab (Treme) in Patients with Metastatic Non-Small Cell Lung Cancer (NSCLC). JCO 2018, 36, TPS9107. [Google Scholar] [CrossRef]
- Shimabukuro, T.T.; Cole, M.; Su, J.R. Reports of Anaphylaxis After Receipt of MRNA COVID-19 Vaccines in the US-14 December 2020–18 January 2021. JAMA 2021, 325, 1101–1102. [Google Scholar] [CrossRef] [PubMed]
- McNeil, M.M.; Weintraub, E.S.; Duffy, J.; Sukumaran, L.; Jacobsen, S.J.; Klein, N.P.; Hambidge, S.J.; Lee, G.M.; Jackson, L.A.; Irving, S.A.; et al. Risk of Anaphylaxis after Vaccination in Children and Adults. J. Allergy Clin. Immunol. 2016, 137, 868–878. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, M.D.; Carrasco, M.J.; Alishetty, S.; Paige, M.; Alameh, M.G.; Weissman, D. Nanomaterial Delivery Systems for MRNA Vaccines. Vaccines 2021, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-Intron Circular RNAs Regulate Transcription in the Nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Koch, L. Translated Circular RNAs. Nat. Rev. Genet. 2017, 18, 272–273. [Google Scholar] [CrossRef]
- Enuka, Y.; Lauriola, M.; Feldman, M.E.; Sas-Chen, A.; Ulitsky, I.; Yarden, Y. Circular RNAs Are Long-Lived and Display Only Minimal Early Alterations in Response to a Growth Factor. Nucleic Acids Res. 2016, 44, 1370–1383. [Google Scholar] [CrossRef] [Green Version]
- Wesselhoeft, R.A.; Kowalski, P.S.; Anderson, D.G. Engineering Circular RNA for Potent and Stable Translation in Eukaryotic Cells. Nat. Commun. 2018, 9, 2629. [Google Scholar] [CrossRef] [Green Version]
- Wesselhoeft, R.A.; Kowalski, P.S.; Parker-Hale, F.C.; Huang, Y.; Bisaria, N.; Anderson, D.G. RNA Circularization Diminishes Immunogenicity and Can Extend Translation Duration In Vivo. Mol. Cell 2019, 74, 508–520.e4. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, S.K.; Belk, J.A.; Amaya, L.; Li, Z.; Cardenas, A.; Abe, B.T.; Chen, C.-K.; Wender, P.A.; Chang, H.Y. Engineering Circular RNA for Enhanced Protein Production. Nat. Biotechnol. 2023, 41, 262–272. [Google Scholar] [CrossRef]
- Segel, M.; Lash, B.; Song, J.; Ladha, A.; Liu, C.C.; Jin, X.; Mekhedov, S.L.; Macrae, R.K.; Koonin, E.V.; Zhang, F. Mammalian Retrovirus-like Protein PEG10 Packages Its Own MRNA and Can Be Pseudotyped for MRNA Delivery. Science 2021, 373, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Fusogenix Drug Delivery System. Entos Pharmaceuticals. Available online: https://www.entospharma.com/fusogenix (accessed on 8 June 2023).
- Sun, Y.; Lau, S.Y.; Lim, Z.W.; Chang, S.C.; Ghadessy, F.; Partridge, A.; Miserez, A. Phase-Separating Peptides for Direct Cytosolic Delivery and Redox-Activated Release of Macromolecular Therapeutics. Nat. Chem. 2022, 14, 274–283. [Google Scholar] [CrossRef]
- Miao, L.; Li, L.; Huang, Y.; Delcassian, D.; Chahal, J.; Han, J.; Shi, Y.; Sadtler, K.; Gao, W.; Lin, J.; et al. Delivery of MRNA Vaccines with Heterocyclic Lipids Increases Anti-Tumor Efficacy by STING-Mediated Immune Cell Activation. Nat. Biotechnol. 2019, 37, 1174–1185. [Google Scholar] [CrossRef]
- Lee, S.M.; Cheng, Q.; Yu, X.; Liu, S.; Johnson, L.T.; Siegwart, D.J. A Systematic Study of Unsaturation in Lipid Nanoparticles Leads to Improved MRNA Transfection In Vivo. Angew. Chem. Int. Ed. Engl. 2021, 60, 5848–5853. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Gong, N.; Shepherd, S.J.; Xiong, X.; Liao, X.; Han, X.; Zhao, G.; Song, C.; Huang, X.; Zhang, H.; et al. Rational Design of Bisphosphonate Lipid-like Materials for MRNA Delivery to the Bone Microenvironment. J. Am. Chem. Soc. 2022, 144, 9926–9937. [Google Scholar] [CrossRef]
- Zhang, D.; Atochina-Vasserman, E.N.; Maurya, D.S.; Liu, M.; Xiao, Q.; Lu, J.; Lauri, G.; Ona, N.; Reagan, E.K.; Ni, H.; et al. Targeted Delivery of MRNA with One-Component Ionizable Amphiphilic Janus Dendrimers. J. Am. Chem. Soc. 2021, 143, 17975–17982. [Google Scholar] [CrossRef]
- Park, J.H.; Mohapatra, A.; Zhou, J.; Holay, M.; Krishnan, N.; Gao, W.; Fang, R.H.; Zhang, L. Virus-Mimicking Cell Membrane-Coated Nanoparticles for Cytosolic Delivery of MRNA. Angew. Chem. Int. Ed. Engl. 2022, 61, e202113671. [Google Scholar] [CrossRef]
- Zhang, S.; Dong, Y.; Wang, Y.; Sun, W.; Wei, M.; Yuan, L.; Yang, G. Selective Encapsulation of Therapeutic MRNA in Engineered Extracellular Vesicles by DNA Aptamer. Nano Lett. 2021, 21, 8563–8570. [Google Scholar] [CrossRef]
- Kedmi, R.; Veiga, N.; Ramishetti, S.; Goldsmith, M.; Rosenblum, D.; Dammes, N.; Hazan-Halevy, I.; Nahary, L.; Leviatan-Ben-Arye, S.; Harlev, M.; et al. A Modular Platform for Targeted RNAi Therapeutics. Nat. Nanotechnol. 2018, 13, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Gutkin, A.; Kedmi, R.; Ramishetti, S.; Veiga, N.; Jacobi, A.M.; Schubert, M.S.; Friedmann-Morvinski, D.; Cohen, Z.R.; Behlke, M.A.; et al. CRISPR-Cas9 Genome Editing Using Targeted Lipid Nanoparticles for Cancer Therapy. Sci. Adv. 2020, 6, eabc9450. [Google Scholar] [CrossRef] [PubMed]
- Su, F.-Y.; Zhao, Q.H.; Dahotre, S.N.; Gamboa, L.; Bawage, S.S.; Silva Trenkle, A.D.; Zamat, A.; Phuengkham, H.; Ahmed, R.; Santangelo, P.J.; et al. In Vivo MRNA Delivery to Virus-Specific T Cells by Light-Induced Ligand Exchange of MHC Class I Antigen-Presenting Nanoparticles. Sci. Adv. 2022, 8, eabm7950. [Google Scholar] [CrossRef] [PubMed]
- Tombácz, I.; Laczkó, D.; Shahnawaz, H.; Muramatsu, H.; Natesan, A.; Yadegari, A.; Papp, T.E.; Alameh, M.-G.; Shuvaev, V.; Mui, B.L.; et al. Highly Efficient CD4+ T Cell Targeting and Genetic Recombination Using Engineered CD4+ Cell-Homing MRNA-LNPs. Mol. Ther. 2021, 29, 3293–3304. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, L.; Wang, X.; Jin, H. RNA-Based Therapeutics: An Overview and Prospectus. Cell Death Dis. 2022, 13, 644. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug Delivery Systems for RNA Therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef]
- Roth, N.; Schön, J.; Hoffmann, D.; Thran, M.; Thess, A.; Mueller, S.O.; Petsch, B.; Rauch, S. CV2CoV, an Enhanced MRNA-Based SARS-CoV-2 Vaccine Candidate, Supports Higher Protein Expression and Improved Immunogenicity in Rats. BioRxiv 2021. [Google Scholar] [CrossRef]
- Uddin, M.N.; Roni, M.A. Challenges of Storage and Stability of MRNA-Based COVID-19 Vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.; Xu, Q. Current Developments and Challenges of MRNA Vaccines. Annu. Rev. Biomed. Eng. 2022, 24, 85–109. [Google Scholar] [CrossRef]
- Knezevic, I.; Liu, M.A.; Peden, K.; Zhou, T.; Kang, H.-N. Development of MRNA Vaccines: Scientific and Regulatory Issues. Vaccines 2021, 9, 81. [Google Scholar] [CrossRef]
- Liu, T.; Liang, Y.; Huang, L. Development and Delivery Systems of MRNA Vaccines. Front Bioeng. Biotechnol. 2021, 9, 718753. [Google Scholar] [PubMed]
- Kannan, S.; Shaik Syed Ali, P.; Sheeza, A. Evolving Biothreat of Variant SARS-CoV-2—Molecular Properties, Virulence and Epidemiology. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4405–4412. [Google Scholar] [PubMed]
- Coronavirus Disease (COVID-19): Variants of SARS-COV-2. Available online: https://www.who.int/news-room/questions-and-answers/item/coronavirus-disease-(covid-19)-variants-of-sars-cov-2 (accessed on 8 June 2023).
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 Protein Interaction Map Reveals Targets for Drug Repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Plante, J.A.; Mitchell, B.M.; Plante, K.S.; Debbink, K.; Weaver, S.C.; Menachery, V.D. The Variant Gambit: COVID-19’s next Move. Cell Host Microbe 2021, 29, 508–515. [Google Scholar] [CrossRef]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an Inactivated Vaccine Candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.-S.; Webby, R.J. An MRNA Vaccine for Influenza. Nat. Biotechnol. 2012, 30, 1202–1204. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The Dawn of MRNA Vaccines: The COVID-19 Case. J. Control Release 2021, 333, 511–520. [Google Scholar] [CrossRef]
- Rajapakse, N.; Dixit, D. Human and Novel Coronavirus Infections in Children: A Review. Paediatr. Int. Child Health 2021, 41, 36–55. [Google Scholar] [CrossRef]
- Staff, A.O.L. Coronavirus Pandemic Now Driven by Younger Adults: WHO. Available online: https://www.aol.com/article/news/2020/08/18/coronavirus-pandemic-now-driven-by-younger-adults-who/24593184/ (accessed on 8 June 2023).
- Collier, D.A.; Ferreira, I.A.T.M.; Kotagiri, P.; Datir, R.P.; Lim, E.Y.; Touizer, E.; Meng, B.; Abdullahi, A.; CITIID-NIHR BioResource COVID-19 Collaboration; Elmer, A.; et al. Age-Related Immune Response Heterogeneity to SARS-CoV-2 Vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef]
- Sherman, S.M.; Smith, L.E.; Sim, J.; Amlôt, R.; Cutts, M.; Dasch, H.; Rubin, G.J.; Sevdalis, N. COVID-19 Vaccination Intention in the UK: Results from the COVID-19 Vaccination Acceptability Study (CoVAccS), a Nationally Representative Cross-Sectional Survey. Hum. Vaccin Immunother. 2021, 17, 1612–1621. [Google Scholar] [CrossRef]
- Kreps, S.; Prasad, S.; Brownstein, J.S.; Hswen, Y.; Garibaldi, B.T.; Zhang, B.; Kriner, D.L. Factors Associated With US Adults’ Likelihood of Accepting COVID-19 Vaccination. JAMA Netw. Open 2020, 3, e2025594. [Google Scholar] [CrossRef] [PubMed]
- Soiza, R.L.; Scicluna, C.; Thomson, E.C. Efficacy and Safety of COVID-19 Vaccines in Older People. Age Ageing 2021, 50, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Zent, O.; Arras-Reiter, C.; Broeker, M.; Hennig, R. Immediate Allergic Reactions after Vaccinations--a Post-Marketing Surveillance Review. Eur. J. Pediatr. 2002, 161, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Thone, M.N.; Kwon, Y.J. COVID-19 Vaccines: The Status and Perspectives in Delivery Points of View. Adv. Drug Deliv. Rev. 2021, 170, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Barda, N.; Dagan, N.; Ben-Shlomo, Y.; Kepten, E.; Waxman, J.; Ohana, R.; Hernán, M.A.; Lipsitch, M.; Kohane, I.; Netzer, D.; et al. Safety of the BNT162b2 MRNA Covid-19 Vaccine in a Nationwide Setting. N Engl. J. Med. 2021, 385, 1078–1090. [Google Scholar] [CrossRef]
- Hampton, L.M.; Aggarwal, R.; Evans, S.J.W.; Law, B. General Determination of Causation between Covid-19 Vaccines and Possible Adverse Events. Vaccine 2021, 39, 1478–1480. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Fayez, N.; Nassar, M.S.; Alshehri, A.A.; Alnefaie, M.K.; Almughem, F.A.; Alshehri, B.Y.; Alawad, A.O.; Tawfik, E.A. Recent Advancement in mRNA Vaccine Development and Applications. Pharmaceutics 2023, 15, 1972. https://doi.org/10.3390/pharmaceutics15071972
Al Fayez N, Nassar MS, Alshehri AA, Alnefaie MK, Almughem FA, Alshehri BY, Alawad AO, Tawfik EA. Recent Advancement in mRNA Vaccine Development and Applications. Pharmaceutics. 2023; 15(7):1972. https://doi.org/10.3390/pharmaceutics15071972
Chicago/Turabian StyleAl Fayez, Nojoud, Majed S. Nassar, Abdullah A. Alshehri, Meshal K. Alnefaie, Fahad A. Almughem, Bayan Y. Alshehri, Abdullah O. Alawad, and Essam A. Tawfik. 2023. "Recent Advancement in mRNA Vaccine Development and Applications" Pharmaceutics 15, no. 7: 1972. https://doi.org/10.3390/pharmaceutics15071972