
Conjugation, Prodrug, and Co-Administration Strategies in Support of Nanotechnologies to Improve the Therapeutic Efficacy of Phytochemicals in the Central Nervous System
 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
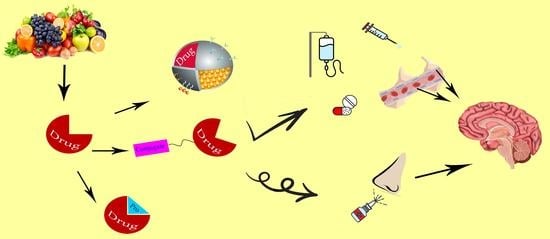
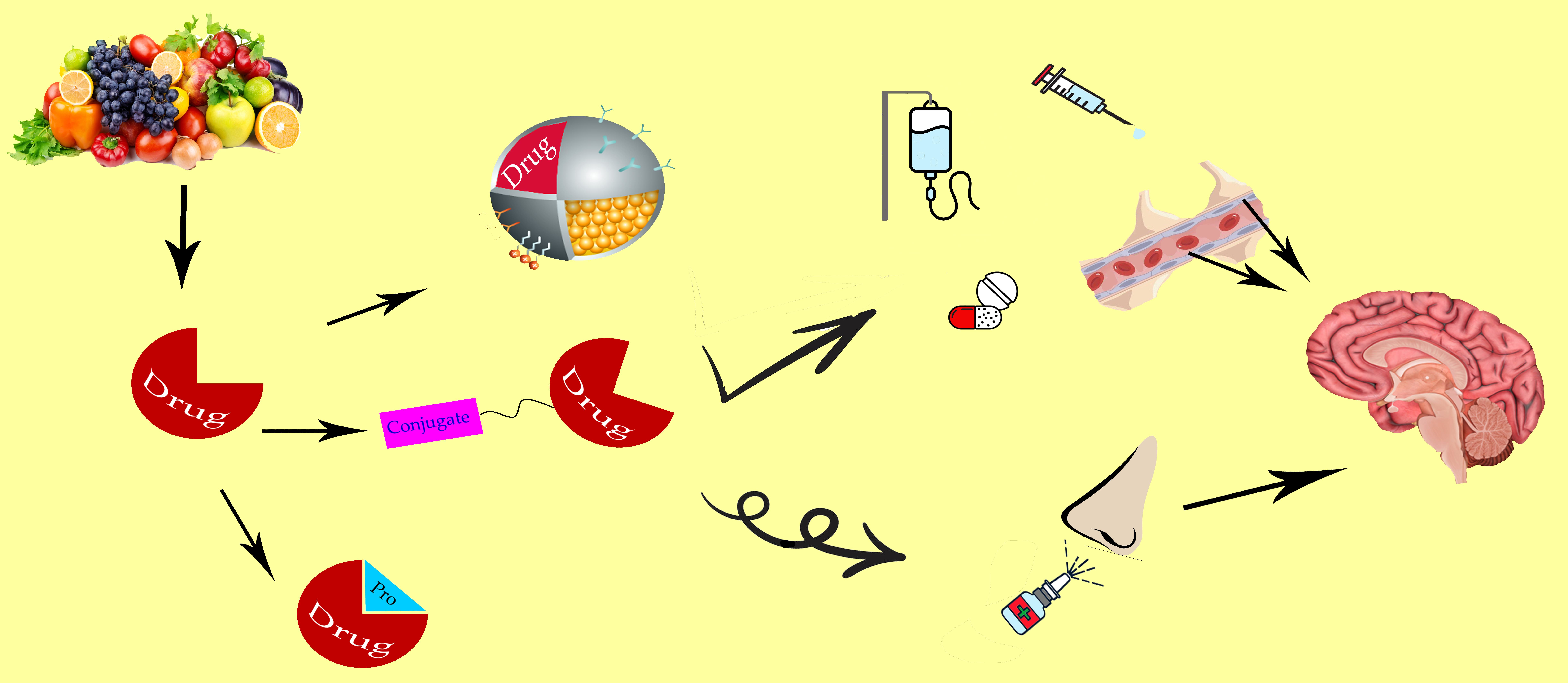
:
1. Introduction
2. Phytochemicals’ Neuroprotective or Anticancer Mechanisms of Action
2.1. Phytochemicals for the Therapy of Neurodegenerative Diseases
2.2. Phytochemicals against Cancer Relapse
3. Cellular Models to Study In Vitro the Efficacy of Phytochemicals against Brain Diseases, and Their Ability to Target the CNS
4. Innovative Formulations Designed to Enhance the Central Effects of Neurotropic Phytochemicals
4.1. Innovative Formulations Designed to Act against the Neurodegenerative Diseases
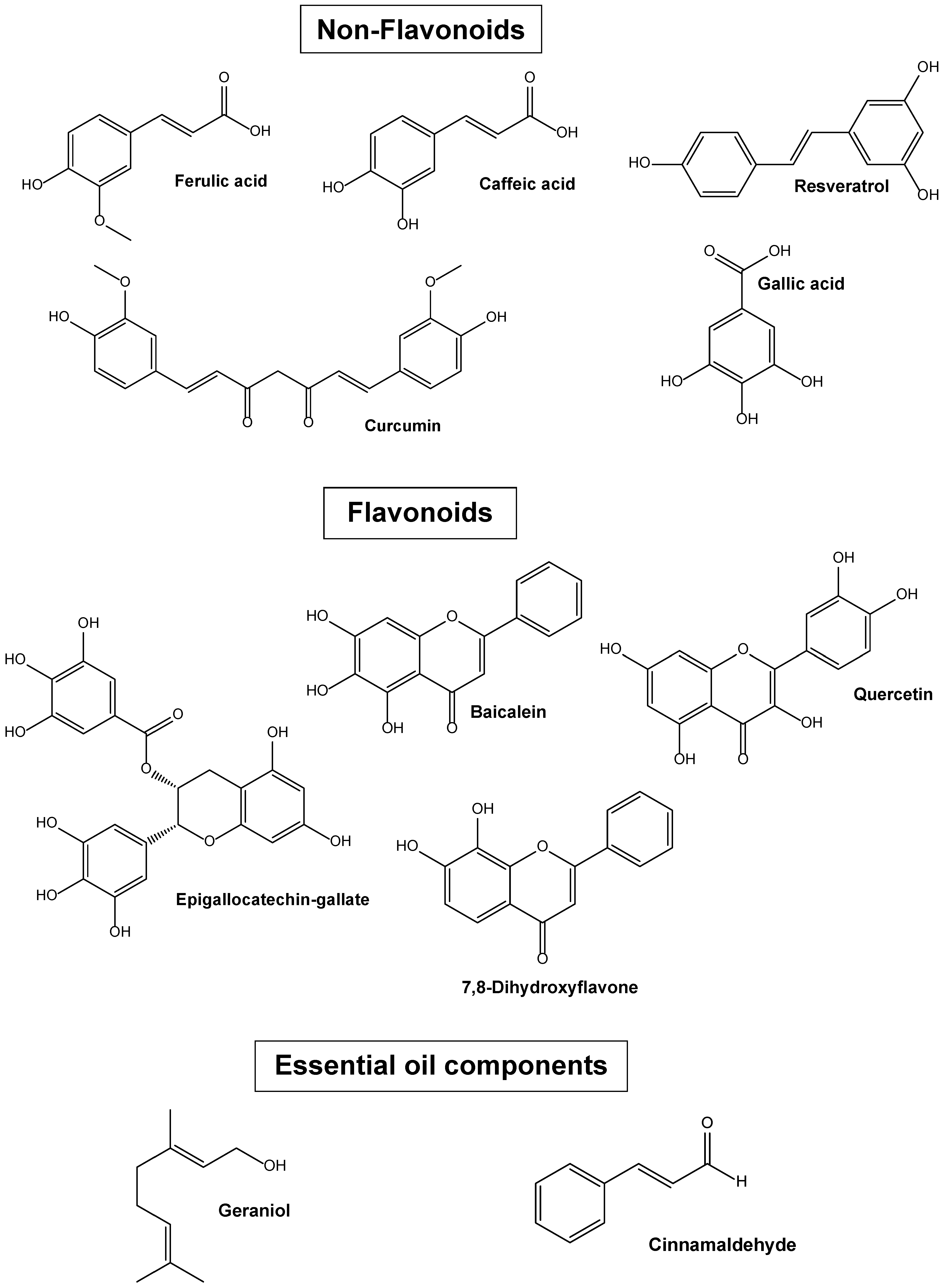
4.1.1. Ferulic Acid in the Design of New Formulations against Brain Diseases
4.1.2. Caffeic Acid in the Design of New Formulations against Brain Diseases
4.1.3. Gallic Acid in the Design of New Formulations against Brain Diseases
4.1.4. Resveratrol in the Design of New Formulations against Brain Diseases
4.1.5. Curcumin in the Design of New Formulations against Brain Diseases
4.1.6. Dihydroxyflavone in the Design of New Formulations against Brain Diseases
4.1.7. Epigallocatechin-Gallate in the Design of New Formulations against Brain Diseases
4.1.8. Baicalein Involved in the Design of New Formulations against Brain Diseases
4.1.9. Quercetin Involved in the Design of New Formulations against Brain Diseases
4.1.10. Genistein in the Design of New Formulations against Brain Diseases
4.1.11. Geraniol in the Design of New Formulations against Brain Diseases
4.2. Innovative Phytochemical Formulations Designed to Treat Glioma
4.2.1. Curcumin Involved in the Design of New Formulations for Glioma Treatment
4.2.2. Resveratrol Involved in the Design of New Formulations for Glioma Treatment
4.2.3. Cinnamaldehyde Involved in the Design of New Formulations for Glioma Treatment
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as potential anti-inflammatory molecules: A review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food components and dietary habits: Keys for a healthy gut microbiota composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef]
- Turini, M.E.; DuBois, R.N. Primary prevention: Phytoprevention and chemoprevention of colorectal cancer. Hematol. Oncol. Clin. N. Am. 2002, 16, 811–840. [Google Scholar] [CrossRef] [PubMed]
- Kerschbaum, E.; Nüssler, V. Cancer prevention with nutrition and lifestyle. Visc. Med. 2019, 35, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Van den Brink, A.C.; Brouwer-Brolsma, E.M.; Berendsen, A.A.M.; van de Rest, O. The mediterranean, dietary approaches to stop hypertension (DASH), and mediterranean-DASH intervention for neurodegenerative delay (MIND) diets are associated with less cognitive decline and a lower risk of Alzheimer’s disease-a review. Adv. Nutr. 2019, 10, 1040–1065. [Google Scholar] [CrossRef]
- Leitzmann, C. Characteristics and health benefits of phytochemicals. Forsch. Komplementmed 2016, 23, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Spisni, E.; Petrocelli, G.; Imbesi, V.; Spigarelli, R.; Azzinnari, D.; Donati Sarti, M.; Campieri, M.; Valerii, M.C. Antioxidant, anti-inflammatory, and microbial-modulating activities of essential oils: Implications in colonic pathophysiology. Int. J. Mol. Sci. 2020, 21, 4152. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Andersen-Civil, A.I.S.; Myhill, L.J.; Thamsborg, S.M.; Kot, W.; Krych, L.; Nielsen, D.S.; Blanchard, A.; Williams, A.R. The phytonutrient cinnamaldehyde limits intestinal inflammation and enteric parasite infection. J. Nutr. Biochem. 2022, 100, 108887. [Google Scholar] [CrossRef]
- Feng, W.; Jin, L.; Xie, Q.; Huang, L.; Jiang, Z.; Ji, Y.; Li, C.; Yang, L.; Wang, D. Eugenol protects the transplanted heart against ischemia/reperfusion injury in rats by inhibiting the inflammatory response and apoptosis. Exp. Ther. Med. 2018, 16, 3464–3470. [Google Scholar] [CrossRef]
- Rekha, K.R.; Selvakumar, G.P. Gene expression regulation of Bcl2, Bax and cytochrome-C by geraniol on chronic MPTP / probenecid induced C57BL/6 mice model of Parkinson’s disease. Chem. Biol. Interact. 2014, 217, 57–66. [Google Scholar] [CrossRef]
- Rekha, K.R.; Selvakumar, G.P. Geraniol ameliorates the motor behavior and neurotrophic factors inadequacy in MPTP-induced mice model of Parkinson’ s disease. J. Mol. Neurosci. 2013, 51, 851–862. [Google Scholar] [CrossRef]
- Pavan, B.; Bianchi, A.; Botti, G.; Ferraro, L.; Valerii, M.C.; Spisni, E.; Dalpiaz, A. Pharmacokinetic and permeation studies in rat brain of natural compounds led to investigate eugenol as direct activator of dopamine release in PC12 cells. Int. J. Mol. Sci. 2023, 24, 1800. [Google Scholar] [CrossRef]
- Hajinejad, M.; Ghaddaripouri, M.; Dabzadeh, M.; Forouzanfar, F.; Sahab-Negah, S. Natural cinnamaldehyde and its derivatives ameliorate neuroinflammatory pathways in neurodegenerative diseases. Biomed. Res. Int. 2020, 2020, 1034325. [Google Scholar] [CrossRef]
- Dalpiaz, A.; Paganetto, G.; Botti, G.; Pavan, B. Cancer stem cells and nanomedicine: New opportunities to combat multidrug resistance? Drug Discov. Today 2020, 25, 1651–1667. [Google Scholar] [CrossRef]
- Moselhy, J.; Srinivasan, S.; Ankem, M.K.; Damodaran, C. Natural products that target cancer stem cells. Anticancer Res. 2015, 35, 5773–5788. [Google Scholar]
- Scarpa, E.S.; Ninfali, P. Phytochemicals as innovative therapeutic tools against cancer stem cells. Int. J. Mol. Sci. 2015, 16, 15727–15742. [Google Scholar] [CrossRef]
- Xu, M.X.; Zhao, L.; Deng, C.; Yang, L.; Wang, Y.; Guo, T.; Li, L.; Lin, J.; Zhang, L. Curcumin suppresses proliferation and induces apoptosis of human hepatocellular carcinoma cells via the wnt signaling pathway. Int. J. Oncol. 2013, 43, 1951–1959. [Google Scholar] [CrossRef]
- Riccardi, G.; Giosuè, A.; Calabrese, I.; Vaccaro, O. Dietary recommendations for prevention of atherosclerosis. Cardiovasc. Res. 2022, 118, 1188–1204. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Pavan, B.; Dalpiaz, A.; Marani, L.; Beggiato, S.; Ferraro, L.; Canistro, D.; Paolini, M.; Vivarelli, F.; Valerii, M.C.; Comparone, A.; et al. Geraniol pharmacokinetics, bioavailability and its multiple effects on the liver antioxidant and xenobiotic-metabolizing enzymes. Front. Pharmacol. 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Thapliyal, S.; Singh, T.; Handu, S.; Bisht, M.; Kumari, P.; Arya, P.; Srivastava, P.; Gandham, R. A review on potential footprints of ferulic acid for treatment of neurological disorders. Neurochem. Res. 2021, 46, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.S.; Chen, L.; Hu, Y.N.; Dai, J.L.; Ma, B.; Tang, Q.F.; Tan, X.M. Self-microemulsifying drug delivery system for improved oral delivery and hypnotic efficacy of ferulic acid. Int. J. Nanomed. 2020, 15, 2059–2070. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, W.; Zhang, Y.; Wang, Q.; Qin, C.; Du, S.; Huang, L.; Ye, F.; Chen, L.; Zheng, T. Pharmacokinetics, bioavailability, and tissue distribution study of angoroside c and its metabolite ferulic acid in rat using UPLC-MS/MS. Front. Pharmacol. 2018, 9, 1186. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lou, Y.I.; Rajesh, R. Dual functional liposomes carrying antioxidants against tau hyperphosphorylation and apoptosis of neurons. J. Drug Target. 2020, 28, 949–960. [Google Scholar] [CrossRef]
- Wu, W.; Lee, S.Y.; Wu, X.; Tyler, J.Y.; Wang, H.; Ouyang, Z.; Park, K.; Xu, X.M.; Cheng, J.X. Neuroprotective ferulic acid (FA)-glycol chitosan (GC) nanoparticles for functional restoration of traumatically injured spinal cord. Biomaterials 2014, 35, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Sozio, P.; Iannitelli, A.; Cerasa, L.S.; Cacciatore, I.; Cornacchia, C.; Giorgioni, G.; Ricciutelli, M.; Nasuti, C.; Cantalamessa, F.; Di Stefano, A. New L-dopa codrugs as potential antiparkinson agents. Arch. Pharm. 2008, 341, 412–417. [Google Scholar] [CrossRef]
- Tian, C.; Asghar, S.; Xu, Y.; Chen, Z.; Zhang, M.; Huang, L.; Ye, J.; Ping, Q.; Xiao, Y. The effect of the molecular weight of hyaluronic acid on the physicochemical characterization of hyaluronic acid-curcumin conjugates and in vitro evaluation in glioma cells. Colloids Surf. B Biointerfaces 2018, 165, 45–55. [Google Scholar] [CrossRef]
- Liu, C.; Gao, Y.; Zhao, L.; Wang, R.; Xie, F.; Zhai, G.; Liu, A. The development of a redox-sensitive curcumin conjugated chitosan oligosaccharide nanocarrier for the efficient delivery of docetaxel to glioma cells. Ann. Transl. Med. 2022, 10, 297. [Google Scholar] [CrossRef]
- Kong, D.; Hong, W.; Yu, M.; Li, Y.; Zheng, Y.; Ying, X. Multifunctional targeting liposomes of epirubicin plus resveratrol improved therapeutic effect on brain gliomas. Int. J. Nanomed. 2022, 17, 1087–1110. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, J.; Guan, Z.; Wu, H.; Cheng, H.; Yan, G.; Tang, R. pH-triggered small molecule nano-prodrugs emulsified from tryptamine-cinnamaldehyde twin drug for targeted synergistic glioma therapy. Colloids Surf. B Biointerfaces 2021, 207, 112052. [Google Scholar] [CrossRef]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Dev. Ther. 2015, 10, 23–42. [Google Scholar] [CrossRef]
- de Oliveira Junior, E.R.; Truzzi, E.; Ferraro, L.; Fogagnolo, M.; Pavan, B.; Beggiato, S.; Rustichelli, C.; Maretti, E.; Lima, E.M.; Leo, E.; et al. Nasal administration of nanoencapsulated geraniol/ursodeoxycholic acid conjugate: Towards a new approach for the management of Parkinson’s disease. J. Control. Release 2020, 321, 540–552. [Google Scholar] [CrossRef]
- Karvandi, M.S.; Hesari, F.S.; Aref, A.R.; Mahdavi, M. The neuroprotective effects of targeting key factors of neuronal cell death in neurodegenerative diseases: The role of ER stress, oxidative stress, and neuroinflammation. Front. Cell. Neurosci. 2023, 17, 1105247. [Google Scholar] [CrossRef]
- Angeloni, C.; Malaguti, M.; Prata, C.; Freschi, M.; Barbalace, M.C.; Hrelia, S. Mechanisms underlying neurodegenerative disorders and potential neuroprotective activity of agrifood by-products. Antioxidants 2022, 12, 94. [Google Scholar] [CrossRef]
- Das, S.; Basu, S. Multi-targeting strategies for Alzheimer’s disease therapeutics: Pros and cons. Curr. Top. Med. Chem. 2017, 17, 3017–3061. [Google Scholar] [CrossRef]
- Limanaqi, F.; Biagioni, F.; Mastroiacovo, F.; Polzella, M.; Lazzeri, G.; Fornai, F. Merging the multi-target effects of phyto-chemicals in neurodegeneration: From oxidative stress to protein aggregation and inflammation. Antioxidants 2020, 9, 1022. [Google Scholar] [CrossRef]
- Yadav, D.K. Potential therapeutic strategies of phytochemicals in neurodegenerative disorders. Curr. Top. Med. Chem. 2021, 21, 2814–2838. [Google Scholar] [CrossRef]
- Pogačnik, L.; Ota, A.; Ulrih, N.P. An overview of crucial dietary substances and their modes of action for prevention of neurodegenerative diseases. Cells 2020, 9, 576. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.F. Neuroprotective role of phytochemi-cals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef]
- Moratilla-Rivera, I.; Sánchez, M.; Valdés-González, J.A.; Gómez-Serranillos, M.P. natural products as modulators of Nrf2 signaling pathway in neuroprotection. Int. J. Mol. Sci. 2023, 24, 3748. [Google Scholar] [CrossRef] [PubMed]
- Iside, C.; Scafuro, M.; Nebbioso, A.; Altucci, L. SIRT1 activation by natural phytochemicals: An overview. Front. Pharmacol. 2020, 11, 1225. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.H.; Lee, H.Y.; Min, K.J. Sirtuin signaling in cellular senescence and aging. BMB Rep. 2019, 52, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Ege, D. Action mechanisms of curcumin in Alzheimer’s disease and its brain targeted delivery. Materials 2021, 14, 3332. [Google Scholar] [CrossRef] [PubMed]
- Minocha, T.; Birla, H.; Obaid, A.A.; Rai, V.; Sushma, P.; Shivamallu, C.; Moustafa, M.; Al-Shehri, M.; Al-Emam, A.; Tikhonova, M.A.; et al. Flavonoids as promising neuroprotectants and their therapeutic potential against Alzheimer’s disease. Oxid. Med. Cell. Longev. 2022, 2022, 6038996. [Google Scholar] [CrossRef]
- Behl, T.; Rana, T.; Sehgal, A.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Meraya, A.M.; Bhatia, S.; Sachdeva, M. Phyto-chemicals targeting nitric oxide signaling in neurodegenerative diseases. Nitric Oxide 2023, 130, 1–11. [Google Scholar] [CrossRef]
- Fantacuzzi, M.; Amoroso, R.; Carradori, S.; De Filippis, B. Resveratrol-based compounds and neurodegeneration: Recent insight in multitarget therapy. Eur. J. Med. Chem. 2022, 233, 114242. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Percaccio, E.; Gullì, M.; Romano, A.; Vitalone, A.; Mazzanti, G.; Gaetani, S.; Di Sotto, A. Recent advances in the neuroprotective properties of ferulic acid in Alzheimer’s disease: A narrative review. Nutrients 2022, 14, 3709. [Google Scholar] [CrossRef]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, probiotics and neurodegenerative dis-eases: Deciphering the gut brain axis. Cell. Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef]
- Molinero, N.; Antón-Fernández, A.; Hernández, F.; Ávila, J.; Bartolomé, B.; Moreno-Arribas, M.V. Gut microbiota, an additional hallmark of human aging and neurodegeneration. Neuroscience 2023, 518, 141–161. [Google Scholar] [CrossRef]
- Saleem, A.; Qurat-Ul-Ain; Akhtar, M.F. Alternative therapy of psychosis: Potential phytochemicals and drug targets in the management of schizophrenia. Front. Pharmacol. 2022, 13, 895668. [Google Scholar] [CrossRef]
- Kuşoğlu, A.; Biray Avcı, Ç. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef]
- Niculescu, V.F. The cancer stem cell family: Atavistic origin and tumorigenic development. MOJ Tumor Res. 2018, 1, 70–73. [Google Scholar] [CrossRef]
- Dick, J.E. Looking ahead in cancer stem cell research. Nat. Biotechnol. 2009, 27, 44–46. [Google Scholar] [CrossRef]
- Peiris-Pagès, M.; Martinez-Outschoorn, U.E.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer stem cell metabolism. Breast Cancer Res. 2016, 18, 55. [Google Scholar] [CrossRef]
- Kozovska, Z.; Gabrisova, V.; Kucerova, L. Colon cancer: Cancer stem cells markers, drug resistance and treatment. Biomed. Pharmacother. 2014, 68, 911–916. [Google Scholar] [CrossRef]
- Du, L.; Wang, H.; He, L.; Zhang, J.; Ni, B.; Wang, X.; Jin, H.; Cahuzac, N.; Mehrpour, M.; Lu, Y.; et al. CD44 is of functional importance for colorectal cancer stem cells. Clin. Cancer Res. 2008, 14, 6751–6760. [Google Scholar] [CrossRef]
- Szaryńska, M.; Olejniczak, A.; Kobiela, J.; Spychalski, P.; Kmieć, Z. Therapeutic strategies against cancer stem cells in human colorectal cancer. Oncol. Lett. 2017, 14, 7653–7668. [Google Scholar] [CrossRef]
- Gupta, S.; Takebe, N.; Lorusso, P. Targeting the Hedgehog pathway in cancer. Ther. Adv. Med. Oncol. 2010, 2, 237–250. [Google Scholar] [CrossRef]
- Yuan, X.; Wu, H.; Xu, H.; Xiong, H.; Chu, Q.; Yu, S.; Wu, G.S.; Wu, K. Notch signaling: An emerging therapeutic target for cancer treatment. Cancer Lett. 2015, 369, 20–27. [Google Scholar] [CrossRef]
- Martin-Orozco, E.; Sanchez-Fernandez, A.; Ortiz-Parra, I.; Ayala-San Nicolas, M. WNT signaling in tumors: The way to evade drugs and immunity. Front. Immunol. 2019, 10, 2854. [Google Scholar] [CrossRef] [PubMed]
- Almanaa, T.N.; Geusz, M.E.; Jamasbi, R.J. Effects of curcumin on stem-like cells in human esophageal squamous carcinoma cell lines. BMC Complement. Altern. Med. 2012, 12, 195. [Google Scholar] [CrossRef]
- Kakarala, M.; Brenner, D.E.; Korkaya, H.; Cheng, C.; Tazi, K.; Ginestier, C.; Liu, S.; Dontu, G.; Wicha, M.S. Targeting breast stem cells with the cancer preventive compounds curcumin and piperine. Breast Cancer Res. Treat. 2010, 122, 777–785. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Calderón-Montaño, J.M.; Salvador, J.; Robles, A.; López-Lázaro, M. The dark side of curcumin. Int. J. Cancer. 2010, 126, 1771–1775. [Google Scholar] [CrossRef] [PubMed]
- Reddivari, L.; Charepalli, V.; Radhakrishnan, S.; Vadde, R.; Elias, R.J.; Lambert, J.D.; Vanamala, J.K. Grape compounds suppress colon cancer stem cells in vitro and in a rodent model of colon carcinogenesis. BMC Complement Altern. Med. 2016, 16, 278. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chu, S.; Yang, Y.; Zhang, Z.; Pang, Z.; Chen, N. Neuroinflammatory In vitro cell culture models and the potential applications for neurological disorders. Front. Pharmacol. 2021, 12, 671734. [Google Scholar] [CrossRef]
- He, W.J.; Lv, C.H.; Chen, Z.; Shi, M.; Zeng, C.X.; Hou, D.X.; Qin, S. The regulatory effect of phytochemicals on chronic diseases by targeting Nrf2-ARE signaling pathway. Antioxidants 2023, 12, 236. [Google Scholar] [CrossRef]
- Bednarek, R. In vitro methods for measuring the permeability of cell monolayers. Methods Protoc. 2022, 5, 17. [Google Scholar] [CrossRef]
- Toth, A.E.; Nielsen, S.S.E.; Tomaka, W.; Abbott, N.J.; Nielsen, M.S. The endo-lysosomal system of bEnd.3 and hCMEC/D3 brain endothelial cells. Fluids Barriers CNS 2019, 16, 14. [Google Scholar] [CrossRef]
- Sun, J.; Ou, W.; Han, D.; Paganini-Hill, A.; Fisher, M.J.; Sumbria, R.K. Comparative studies between the murine immortalized brain endothelial cell line (bEnd.3) and induced pluripotent stem cell-derived human brain endothelial cells for paracellular transport. PLoS ONE 2022, 17, e0268860. [Google Scholar] [CrossRef]
- Botti, G.; Bianchi, A.; Pavan, B.; Tedeschi, P.; Albanese, V.; Ferraro, L.; Spizzo, F.; Del Bianco, L.; Dalpiaz, A. Effects of mi-croencapsulated ferulic acid or its prodrug methyl ferulate on neuroinflammation induced by muramyl dipeptide. Int. J. Environ. Res. Public Health 2022, 19, 10609. [Google Scholar] [CrossRef]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef]
- Formolo, C.A.; Williams, R.; Gordish-Dressman, H.; MacDonald, T.J.; Lee, N.H.; Hathout, Y. Secretome signature of invasive glioblastoma multiforme. J. Proteome Res. 2011, 10, 3149–3159. [Google Scholar] [CrossRef]
- Ghosh, S.; Basak, P.; Dutta, S.; Chowdhury, S.; Sil, P.C. New insights into the ameliorative effects of ferulic acid in pathophysiological conditions. Food Chem. Toxicol. 2017, 103, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, C.; Zhang, Y.; Mi, S.; Wang, N. Pharmacokinetics of ferulic acid and potential interactions with Honghua and clopidogrel in rats. J. Ethnopharmacol. 2011, 137, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, P.; Arbabi, E.; Atyabi, F.; Dinarvand, R. Ferulic acid-loaded nanostructured lipid carriers: A promising nanoformulation against the ischemic neural injuries. Life Sci. 2018, 193, 64–76. [Google Scholar] [CrossRef]
- Saini, S.; Sharma, T.; Jain, A.; Kaur, H.; Katare, O.P.; Singh, B. Systematically designed chitosan-coated solid lipid nanoparticles of ferulic acid for effective management of Alzheimer’s disease: A preclinical evidence. Colloids Surf. B Biointerfaces 2021, 205, 111838. [Google Scholar] [CrossRef]
- Qin, J.; Chen, D.; Hu, H.; Cui, Q.; Qiao, M.; Chen, B. Surface modification of RGD-liposomes for selective drug delivery to monocytes/neutrophils in brain. Chem. Pharm. Bull. 2007, 55, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Chen, D.; Hu, H.; Qiao, M.; Zhao, X.; Chen, B. Body distribution of RGD-mediated liposome in brain-targeting drug delivery. Yakugaku Zasshi 2007, 127, 1497–1501. [Google Scholar] [CrossRef]
- Puris, E.; Gynther, M.; Huttunen, J.; Auriola, S.; Huttunen, K.M. L-type amino acid transporter 1 utilizing prodrugs of ferulic acid revealed structural features supporting the design of prodrugs for brain delivery. Eur. J. Pharm. Sci. 2019, 129, 99–109. [Google Scholar] [CrossRef]
- Montaser, A.; Huttunen, J.; Ibrahim, S.A.; Huttunen, K.M. Astrocyte-targeted transporter-utilizing derivatives of ferulic acid can have multifunctional effects ameliorating inflammation and oxidative stress in the brain. Oxid. Med. Cell. Longev. 2019, 2019, 3528148. [Google Scholar] [CrossRef]
- Rosini, M.; Simoni, E.; Caporaso, R.; Basagni, F.; Catanzaro, M.; Abu, I.F.; Fagiani, F.; Fusco, F.; Masuzzo, S.; Albani, D.; et al. Merging memantine and ferulic acid to probe connections between NMDA receptors, oxidative stress and amyloid-β peptide in Alzheimer’s disease. Eur. J. Med. Chem. 2019, 180, 111–120. [Google Scholar] [CrossRef]
- Pavlíková, N. Caffeic acid and diseases—Mechanisms of action. Int. J. Mol. Sci. 2023, 24, 588. [Google Scholar] [CrossRef]
- Grabska-Kobylecka, I.; Kaczmarek-Bak, J.; Figlus, M.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Glabinski, A.; Nowak, D. The presence of caffeic acid in cerebrospinal fluid: Evidence that dietary polyphenols can cross the blood-brain barrier in humans. Nutrients 2020, 12, 1531. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic implications of caffeic acid in cancer and neurological diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Choi, K.C.; Kang, M.S.; Oh, J.S.; Jeong, Y.; Lee, H.C. Self-organized nanoparticles of caffeic acid conjugated polysaccharide and its anticancer activity. J. Nanosci. Nanotechnol. 2015, 15, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Mehtap Kutlu, H.; Genc, L.; Guney, G. The impact of caffeic acid loaded solid lipid nanoparticles on cancer treatment. Current Nanoscience 2013, 9, 698–703. [Google Scholar] [CrossRef]
- Šebestík, J.; Marques, S.M.; Falé, P.L.; Santos, S.; Arduíno, D.M.; Cardoso, S.M.; Oliveira, C.R.; Serralheiro, M.L.M.; Santos, M.A. Bifunctional phenolic-choline conjugates as anti-oxidants and acetylcholinesterase inhibitors. J. Enzyme Inhib. Med. Chem. 2011, 26, 485–497. [Google Scholar] [CrossRef]
- Akomolafe, S.F. The effects of caffeine, caffeic acid, and their combination on acetylcholin-esterase, adenosine deaminase and arginase activities linked with brain function. J. Food Biochem. 2017, 41, e12401. [Google Scholar] [CrossRef]
- Mishra, R.K.; Ahmad, A.; Kanika; Kumar, A.; Vyawahare, A.; Sakla, R.; Nadeem, A.; Siddiqui, N.; Raza, S.S.; Khan, R. Caffeic acid-conjugated budesonide-loaded nanomicelle attenuates inflammation in experimental colitis. Mol. Pharm. 2023, 20, 172–182. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Pal, S.M.; Avneet, G.; Siddhraj, S.S. Gallic acid: Pharmacogical promising lead molecule: A review. Int. J. Pharmacogn. Phytochem. Res. 2018, 10, 132–138. [Google Scholar] [CrossRef]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and diseases: A mechanistic review. Iran. J. Basic Med. Sci. 2019, 22, 225–237. [Google Scholar] [CrossRef]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A role of gallic acid in oxidative damage diseases: A comprehensive review. Nat. Prod. Commun. 2019, 14, 1–9. [Google Scholar] [CrossRef]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and po-tential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Choubey, S.; Varughese, L.; Kumar, V.; Beniwal, V. Medicinal importance of gallic acid and its ester derivatives: A patent review. Pharm. Pat. Anal. 2015, 4, 305–315. [Google Scholar] [CrossRef]
- Chen, L.; Huanga, G.L.; Lü, M.H.; Zhang, J.X.; Xu, J.; Bai, S.P. Amide derivatives of Gallic acid: Design, synthesis and evaluation of inhibitory activities against in vitro α-synuclein aggregation. Bioorg. Med. Chem. 2020, 28, 1155961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chang, R.; Ren, Y.; He, Y.; Guo, S.; Guan, F.; Yao, M. Injectable and reactive oxygen species-scavenging gelatin hydrogel promotes neural repair in experimental trau-matic brain injury. Int. J. Biol. Macromol. 2022, 219, 844–863. [Google Scholar] [CrossRef] [PubMed]
- Madhusudanan, P.; Raju, G.; Shankarappa, S. Hydrogel systems and their role in neural tissue engineering. J. R. Soc. Interface 2020, 17, 20190505. [Google Scholar] [CrossRef]
- Yao, M.; Gao, F.; Xu, R.; Zhang, J.; Chen, Y.; Guan, F. A dual-enzymatically crosslinked in-jectable gelatin hydrogel loaded with BMSC improves neurological function recovery of traumatic brain injury in rats. Biomater. Sci. 2019, 7, 4088–4098. [Google Scholar] [CrossRef] [PubMed]
- Bostancieri, N.; Elbe, H.; Esrefoglu, M.; Vardi, N. Cardioprotective potential of melatonin, quercetin and resveratrol in an experimental model of diabetes. Biotech. Histochem. 2022, 97, 152–157. [Google Scholar] [CrossRef]
- Bonferoni, M.C.; Rossi, S.; Sandri, G.; Ferrari, F. Nanoparticle formulations to enhance tu-mor targeting of poorly soluble polyphenols with potential anticancer properties. Semin. Cancer Biol. 2017, 46, 205–214. [Google Scholar] [CrossRef]
- Fonseca-Santos, B.; Chorilli, M. The uses of resveratrol for neurological diseases treatment and insights for nanotechnology based-drug delivery systems. Int. J. Pharm. 2020, 589, 119832. [Google Scholar] [CrossRef]
- Andrade, S.; Ramalho, M.J.; Pereira, M.D.C.; Loureiro, J.A. Resveratrol Brain Delivery for Neurological Disorders Prevention and Treatment. Front. Pharmacol. 2018, 9, 1261. [Google Scholar] [CrossRef]
- Komorowska, J.; Wątroba, M.; Szukiewicz, D. Review of beneficial effects of resveratrol in neurodegenerative diseases such as Alzheimer’s disease. Adv. Med. Sci. 2020, 65, 415–423. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, C.X.; Kakar, M.U.; Khan, M.S.; Wu, P.F.; Amir, R.M.; Dai, D.F.; Naveed, M.; Li, Q.Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological re-view and call for further research. Biomed. Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, T.; Li, W.; Gao, N.; Zhang, T. Resveratrol attenuates oxidative damage through activating mitophagy in an in vitro model of Alzheimer’s disease. Toxicol. Lett. 2018, 282, 100–108. [Google Scholar] [CrossRef]
- Bastianetto, S.; Zheng, W.H.; Quirion, R. Neuroprotective abilities of resveratrol and other red wine constituents against nitric oxide-related toxicity in cultured hippocampal neu-rons. Br. J. Pharmacol. 2000, 131, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, R.; Anada, M.; Miyaguchi, A.; Nomi, Y.; Matsumoto, H. Evaluation of Blood-Brain Barrier Permeability of Polyphenols, Anthocyanins, and Their Metabolites. J. Agric. Food Chem. 2021, 69, 11676–11686. [Google Scholar] [CrossRef] [PubMed]
- Katila, N.; Duwa, R.; Bhurtel, S.; Khanal, S.; Maharjan, S.; Jeong, J.H.; Lee, S.; Choi, D.Y.; Yook, S. Enhancement of blood-brain barrier penetration and the neuroprotective effect of resveratrol. J. Control. Release 2022, 346, 1–19. [Google Scholar] [CrossRef]
- Shen, Y.; Cao, B.; Snyder, N.R.; Woeppel, K.M.; Eles, J.R.; Cui, X.T. ROS responsive resvera-trol delivery from LDLR peptide conjugated PLA-coated mesoporous silica nanoparticles across the blood-brain barrier. J. Nanobiotechnology 2018, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Intagliata, S.; Modica, M.N.; Santagati, L.M.; Montenegro, L. Strategies to improve resveratrol systemic and topical bioavailability: An update. Antioxidants 2019, 8, 244. [Google Scholar] [CrossRef]
- Chao, J.; Li, H.; Cheng, K.W.; Yu, M.S.; Chang, R.C.; Wang, M. Protective effects of pinostil-bene, a resveratrol methylated derivative, against 6- hydroxydopamine-induced neuro-toxicity in SH-SY5Y cells. J. Nutr. Biochem. 2010, 21, 482e489. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Rimando, A.; Pallas, M.; Camins, A.; Porquet, D.; Reeves, J.; Shukitt-Hale, B.; Smith, M.A.; Joseph, J.A.; Casadesus, G. Low-dose pterostilbene, but not resveratrol, is a potent neuromodulator in aging and Alzheimer’s disease. Neurobiol. Aging 2012, 33, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Thangthaeng, N.; Miller, M.G.; Shukitt-Hale, B. Effects of pterostilbene and resveratrol on brain and behaviour. Neurochem. Int. 2015, 89, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lu, F.; Zhang, X.; Liu, S.; Mu, P. SIRT1 is involved in the neuroprotection of pterostilbene against amyloid β 25–35-induced cognitive deficits in mice. Front. Pharmacol. 2022, 13, 877098. [Google Scholar] [CrossRef]
- Potdar, S.; Parmar, M.S.; Ray, S.D.; Cavanaugh, J.E. Protective effects of the resveratrol analog piceid in dopaminergic SH-SY5Y cells. Arch. Toxicol. 2018, 92, 669–677. [Google Scholar] [CrossRef]
- Peñalver, P.; Belmonte-Reche, E.; Adán, N.; Caro, M.; Mateos-Martín, M.L.; Delgado, M.; González-Rey, E.; Morales, J.C. Alkylated resveratrol prodrugs and metabolites as poten-tial therapeutics for neurodegenerative diseases. Eur. J. Med. Chem. 2018, 146, 123–138. [Google Scholar] [CrossRef]
- Belmonte-Reche, E.; Peñalver, P.; Caro-Moreno, M.; Mateos-Martín, M.L.; Adán, N.; Delga-do, M.; González-Rey, E.; Morales, J.C. Silyl resveratrol derivatives as potential therapeutic agents for neurodegenerative and neurological diseases. Eur. J. Med. Chem. 2021, 223, 113655. [Google Scholar] [CrossRef]
- Grau, L.; Soucek, R.; Pujol, M.D. Resveratrol derivatives: Synthesis and their biological activities. Eur. J. Med. Chem. 2023, 246, 114962. [Google Scholar] [CrossRef]
- Wightman, E.L.; Reay, J.L.; Haskell, C.F.; Williamson, G.; Dew, T.P.; Kennedy, D.O. Effects of resveratrol alone or in combination with piperine on cerebral blood flow parameters and cognitive performance in human subjects: A randomised, double-blind, place-bo-controlled, cross-over investigation. Br. J. Nutr. 2014, 112, 203–313. [Google Scholar] [CrossRef]
- Haq, I.-U.; Imran, M.; Nadeem, M.; Tufail, T.; Gondal, T.A.; Mubarak, M.S. Piperine: A review of its biological effects. Phytother. Res. 2021, 35, 680–700. [Google Scholar] [CrossRef]
- Johnson, J.J.; Nihal, M.; Siddiqui, I.A.; Scarlett, C.O.; Bailey, H.H.; Mukhtar, H.; Ahmad, N. Enhancing the bioavailability of resveratrol by combining it with piperine. Mol. Nutr. Food Res. 2011, 55, 1169–1176. [Google Scholar] [CrossRef]
- Belkacemi, A.; Doggui, S.; Dao, L.; Ramassamy, C. Challenges associated with curcumin therapy in Alzheimer disease. Expert. Rev. Mol. Med. 2011, 13, e34. [Google Scholar] [CrossRef]
- Mathew, A.; Fukuda, T.; Nagaoka, Y.; Hasumura, T.; Morimoto, H.; Yoshida, Y.; Maekawa, T.; Venugopal, K.; Kumar, D.S. Cur-cumin loaded-PLGA nanoparticles conjugated with Tet-1 peptide for potential use in Alzheimer’s disease. PLoS ONE 2012, 7, e32616. [Google Scholar] [CrossRef]
- Lazar, A.N.; Mourtas, S.; Youssef, I.; Parizot, C.; Dauphin, A.; Delatour, B.; Antimisiaris, S.G.; Duyckaerts, C. Curcu-min-conjugated nanoliposomes with high affinity for Aβ deposits: Possible applications to Alzheimer disease. Nanomedicine 2013, 9, 712–721. [Google Scholar] [CrossRef]
- Wei, X.L.; Han, Y.R.; Quan, L.H.; Liu, C.Y.; Liao, Y.H. Oily nanosuspension for long-acting intramuscular delivery of curcumin didecanoate prodrug: Preparation, characterization and in vivo evaluation. Eur. J. Pharm. Sci. 2013, 49, 286–293. [Google Scholar] [CrossRef]
- Mourtas, S.; Lazar, A.N.; Markoutsa, E.; Duyckaerts, C.; Antimisiaris, S.G. Multifunctional nanoliposomes with curcumin-lipid derivative and brain targeting functionality with potential applications for Alzheimer disease. Eur. J. Med. Chem. 2014, 80, 175–183. [Google Scholar] [CrossRef]
- Frielingsdorf, H.; Simpson, D.R.; Thal, L.J.; Pizzo, D.P. Nerve growth factor promotes survival of new neurons in the adult hip-pocampus. Neurobiol. Dis. 2007, 26, 47–55. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lin, C.C. Rescuing apoptotic neurons in Alzheimer’s disease using wheat germ agglutinin-conjugated and cardi-olipin-conjugated liposomes with encapsulated nerve growth factor and curcumin. Int. J. Nanomed. 2015, 10, 2653–2672. [Google Scholar] [CrossRef]
- Paka, G.D.; Ramassamy, C. Optimization of curcumin-loaded PEG-PLGA nanoparticles by GSH functionalization: Investigation of the internalization pathway in neuronal cells. Mol. Pharm. 2017, 14, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Lu, S.; Liu, X.G.; Zhu, J.; Wang, Y.J.; Liu, R.T. PLGA nanoparticles modified with a BBB-penetrating peptide co-delivering Aβ generation inhibitor and curcumin attenuate memory deficits and neuropathology in Alzheimer’s disease mice. Oncotarget 2017, 8, 81001–81013. [Google Scholar] [CrossRef]
- Fan, S.; Zheng, Y.; Liu, X.; Fang, W.; Chen, X.; Liao, W.; Jing, X.; Lei, M.; Tao, E.; Ma, Q.; et al. Curcumin-loaded PLGA-PEG nanoparticles conjugated with B6 peptide for potential use in Alzheimer’s disease. Drug Deliv. 2018, 25, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, X.; Kang, T.; Jiang, M.; Miao, D.; Gu, G.; Hu, Q.; Song, Q.; Yao, L.; Tu, Y.; et al. B6 peptide-modified PEG-PLA nanoparticles for enhanced brain delivery of neuroprotective peptide. Bioconjugate Chem. 2013, 24, 997–1007. [Google Scholar] [CrossRef]
- Pepe, G.; Calce, E.; Verdoliva, V.; Saviano, M.; Maglione, V.; Di Pardo, A.; De Luca, S. Curcumin-loaded nanoparticles based on amphiphilic hyaluronan-conjugate explored as targeting delivery system for neurodegenerative disorders. Int. J. Mol. Sci. 2020, 21, 8846. [Google Scholar] [CrossRef] [PubMed]
- Barzegarzadeh, B.; Hatami, H.; Dehghan, G.; Khajehnasiri, N.; Khoobi, M.; Sadeghian, R. Conjugated linoleic acid-curcumin attenuates cognitive deficits and oxidative stress parameters in the ethidium bromide–induced model of demyelination. Neurotox. Res. 2021, 39, 815–825. [Google Scholar] [CrossRef]
- Phongpradist, R.; Thongchai, W.; Thongkorn, K.; Lekawanvijit, S.; Chittasupho, C. Surface modification of curcumin microemul-sions by coupling of KLVFF peptide: A prototype for targeted bifunctional microemulsions. Polymers 2022, 14, 443. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Z.; Zhang, Z.; Liu, X.; Kang, S.S.; Zhang, Y.; Ye, K. The prodrug of 7,8-dihydroxyflavone development and therapeutic efficacy for treating Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, 578–583. [Google Scholar] [CrossRef]
- Zuccato, C.; Cattaneo, E. Brain-derived neurotrophic factor in neurodegenerative diseases. Nat. Rev. Neurol. 2009, 5, 311–322. [Google Scholar] [CrossRef]
- Ando, S.; Kobayashi, S.; Waki, H.; Kon, K.; Fukui, F.; Tadenuma, T.; Iwamoto, M.; Takeda, Y.; Izumiyama, N.; Watanabe, K.; et al. Animal model of dementia induced by entorhinal synaptic damage and partial restoration of cognitive deficits by BDNF and carnitine. J. Neurosci. Res. 2002, 70, 519–527. [Google Scholar] [CrossRef]
- Jang, S.W.; Liu, X.; Yepes, M.; Shepherd, K.R.; Miller, G.W.; Liu, Y.; Wilson, W.; Xiao, G.; Blanchi, B.; Sun, Y.E.; et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc. Natl. Acad. Sci. USA 2010, 107, 2687–2692. [Google Scholar] [CrossRef]
- Liu, X.; Qi, Q.; Xiao, G.; Li, J.; Luo, H.R.; Ye, K. O-methylated metabolite of 7,8-dihydroxyflavone activates TrkB receptor and displays antidepressant activity. Pharmacology 2013, 91, 185–200. [Google Scholar] [CrossRef]
- English, A.W.; Carrasco, D.; Hoffman, D.; Isaacson, R.; Kang, S.S.; Khan, S.; Liu, X.; Ye, K. Oral Treatments with the TrkB ligand prodrug, R13, promote enhanced axon regeneration following peripheral nerve injury. Front Cell. Neurosci. 2022, 16, 857664. [Google Scholar] [CrossRef]
- Pervin, M.; Unno, K.; Takagaki, A.; Isemura, M.; Nakamura, Y. Function of green tea catechins in the brain: Epigallocatechin gallate and its metabolites. Int. J. Mol. Sci. 2019, 20, 3630. [Google Scholar] [CrossRef]
- Levites, Y.; Weinreb, O.; Maor, G.; Youdim, M.B.; Mandel, S. Green tea polyphenol (-)-epigallocatechin-3-gallate prevents N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced dopaminergic neurodegeneration. J. Neurochem. 2001, 78, 1073–1082. [Google Scholar] [CrossRef]
- Ferri, P.; Angelino, D.; Gennari, L.; Benedetti, S.; Ambrogini, P.; Del Grande, P.; Ninfali, P. Enhancement of flavonoid ability to cross the blood-brain barrier of rats by co-administration with α-tocopherol. Food Funct. 2015, 6, 394–400. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Wang, I.H.; Rajesh, R. Use of leptin-conjugated phosphatidic acid liposomes with resveratrol and epigallocatechin gallate to protect dopaminergic neurons against apoptosis for Parkinson’s disease therapy. Acta Biomater. 2021, 119, 360–374. [Google Scholar] [CrossRef]
- Banks, W.A. Leptin transport across the blood-brain barrier: Implications for the cause and treatment of obesity. Curr. Pharm. Des. 2001, 7, 125–133. [Google Scholar] [CrossRef]
- Lu, J.; Park, C.S.; Lee, S.K.; Shin, D.W.; Kang, J.H. Leptin inhibits 1-methyl-4-phenylpyridinium-induced cell death in SH-SY5Y cells. Neurosci. Lett. 2006, 407, 240–243. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, J.; Hölscher, C. Therapeutic potential of baicalein in Alzheimer’s disease and Parkinson’s disease. CNS Drugs 2017, 31, 639–652. [Google Scholar] [CrossRef]
- Tarragó, T.; Kichik, N.; Claasen, B.; Prades, R.; Teixidó, M.; Giralt, E. Baicalin, a Prodrug able to reach the CNS, is a prolyl oligopeptidase inhibitor. Bioorg. Med. Chem. 2008, 16, 7516–7524. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-Z.; Wang, H.-T.; Huang, H.-J.; Lo, Y.-L.; Lin, A.M.-Y. Neuroprotective effects of baicalein on acrolein-induced neurotoxicity in the nigrostriatal dopaminergic system of rat brain. Mol. Neurobiol. 2018, 55, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Peng, L.; Gong, P.; Zheng, X.; Sun, T.; Zhang, X.; Huo, J. Baicalein mediates mi-tochondrial autophagy via mir-30b and the NIX/BNIP3 signaling pathway in Parkinson’s disease. Biochem. Res. Int. 2021, 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Li, Y.; Zhang, Y.; Chen, J.; Gao, J.; Zhang, T.; Shang, X.; Zhang, X. Baicalein amelio-rates Aβ-induced memory deficits and neuronal atrophy via inhibition of PDE2 and PDE4. Front. Pharmacol. 2021, 12, 794458. [Google Scholar] [CrossRef]
- Chen, B.; Luo, M.; Liang, J.; Zhang, C.; Gao, C.; Wang, J.; Wang, J.; Li, Y.; Xu, D.; Liu, L.; et al. Surface modification of PGP for a neutrophil–nanoparticle co-vehicle to enhance the anti-depressant effect of baicalein. Acta Pharm. Sin. B 2018, 8, 64–73. [Google Scholar] [CrossRef]
- Jia, X.; Jia, M.; Yang, Y.; Wang, D.; Zhou, F.; Zhang, W.; Huang, X.; Guo, W.; Cai, D.; Chen, H.; et al. Synthesis of novel baicalein amino acid derivatives and biological evaluation as neuroprotective agents. Molecules 2019, 24, 3647. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The pharmacological activity, biochemical properties, and pharmacokinetics of the major natural polyphenolic flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef]
- Dajas, F.; Abin-Carriquiry, J.A.; Arredondo, F.; Blasina, F.; Echeverry, C.; Martínez, M.; Rivera, F.; Vaamonde, L. Quercetin in brain diseases: Potential and limits. Neurochem. Int. 2015, 89, 140–148. [Google Scholar] [CrossRef]
- Porcu, E.P.; Cossu, M.; Rassu, G.; Giunchedi, P.; Cerri, G.; Pourová, J.; Najmanová, I.; Migkos, T.; Pilařová, V.; No-váková, L.; et al. Aqueous injection of quercetin: An approach for confirmation of its direct in vivo cardiovascular effects. Int. J. Pharm. 2018, 541, 224–233. [Google Scholar] [CrossRef]
- Manta, K.; Papakyriakopoulou, P.; Chountoulesi, M.; Diamantis, D.A.; Spaneas, D.; Vakali, V.; Naziris, N.; Chat-ziathanasiadou, M.V.; Andreadelis, I.; Moschovou, K.; et al. Preparation and bio-physical characterization of quercetin inclusion complexes with β-cyclodextrin derivatives to be formulated as possible nose-to-brain quercetin delivery systems. Mol. Pharm. 2020, 17, 4241–4255. [Google Scholar] [CrossRef]
- Dhawan, S.; Kapil, R.; Singh, B. Formulation development and systematic optimization of solid lipid nanoparticles of quercetin for improved brain delivery. J. Pharm. Pharmacol. 2011, 63, 342–351. [Google Scholar] [CrossRef]
- Pinheiro, R.G.R.; Granja, A.; Loureiro, J.A.; Pereira, M.C.; Pinheiro, M.; Neves, A.R.; Reis, S. RVG29-functionalized li-pid nanoparticles for quercetin brain delivery and Alzheimer’s disease. Pharm. Res. 2020, 37, 139. [Google Scholar] [CrossRef]
- Guo, L.; Huang, Z.; Huang, L.; Liang, J.; Wang, P.; Zhao, L.; Shi, Y. Surface-modified engineered exosomes attenuat-ed cerebral ischemia/reperfusion injury by targeting the delivery of quercetin towards impaired neurons. J. Nanobiotech. 2021, 19, 141. [Google Scholar] [CrossRef]
- Cen, J.; Zhang, R.; Zhao, T.; Zhang, X.; Zhang, C.; Cui, J.; Zhao, K.; Duan, S.; Guo, Y. A water-soluble quercetin conjugate with triple targeting exerts neuron-protective effect on cerebral ischemia by mitophagy activation. Adv. Healthc. Mater. 2022, 11, e2200817. [Google Scholar] [CrossRef]
- Ebrahimpour, S.; Esmaeili, A.; Beheshti, S. Effect of quercetin-conjugated superparamagnetic iron oxide nanoparti-cles on diabetes-induced learning and memory impairment in rats. Int. J. Nanomed. 2018, 13, 6311–6324. [Google Scholar] [CrossRef]
- Ebrahimpour, S.; Esmaeili, A.; Dehghanian, F.; Beheshti, S. Effects of quercetin-conjugated with superparamagnetic iron oxide nanoparticles on learning and memory improvement through targeting microRNAs/NF-ΚB Pathway. Sci. Rep. 2020, 10, 15070. [Google Scholar] [CrossRef]
- Enteshari Najafabadi, R.; Kazemipour, N.; Esmaeili, A.; Beheshti, S.; Nazifi, S. Using superparamagnetic iron oxide nanoparticles to enhance bioavailability of quercetin in the intact rat brain. BMC Pharmacol. Toxicol. 2018, 19, 59. [Google Scholar] [CrossRef]
- Yarjanli, Z.; Ghaedi, K.; Esmaeili, A.; Zarrabi, A.; Rahgozar, S. The antitoxic effects of quercetin and quercetin-conjugated iron oxide nanoparticles (QNPs) against H2O2-induced toxicity in PC12 cells. Int. J. Nanomed. 2019, 14, 6813–6830. [Google Scholar] [CrossRef]
- Hashemian, M.; Ghasemi-Kasman, M.; Ghasemi, S.; Akbari, A.; Moalem-Banhangi, M.; Zare, L.; Ahmadian, S.R. Fabrication and evaluation of novel quercetin-conjugated Fe3O4–β-Cyclodextrin nanoparticles for potential use in epilepsy disorder. Int. J. Nanomed. 2019, 14, 6481–6495. [Google Scholar] [CrossRef]
- Bardestani, A.; Ebrahimpour, S.; Esmaeili, A.; Esmaeili, A. Quercetin attenuates neurotoxicity induced by iron oxide nanoparticles. J. Nanobiotech. 2021, 19, 327. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.X.; Jalil, J.; Lam, K.W.; Husain, K.; Premakumar, C.M. Genistein: A review on its anti-inflammatory properties. Front. Pharmacol. 2022, 13, 820969. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Y.; Gu, J.; Liang, P.; Shen, M.; Xi, J.; Qin, J. Anti-invasive effect and pharmacological mechanism of genistein against colorectal cancer. BioFactors 2020, 46, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, P.; Puga-Olguín, A.; Rodríguez-Landa, J.F.; Zepeda, R.C. Genistein as potential therapeutic candidate formenopausal symptoms and other related diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef]
- Sansai, K.; Na Takuathung, M.; Khatsri, R.; Teekachunhatean, S.; Hanprasertpong, N.; Koonrungsesomboon, N. Effects of isoflavone interventions on bone mineral density in postmenopausal women: A systematic review and meta-analysis of randomized controlled trials. Osteoporos. Int. 2020, 31, 1853–1864. [Google Scholar] [CrossRef]
- Sarkaki, A.; Amani, R.; Badavi, M.; Moghaddam, A.Z.; Aligholi, H.; Safahani, M.; Haghighizadeh, M.H. Pre-treatment effect of different doses of soy isoflavones on spatial learning and memory in an ovariectomized animal model of Alzheimer’s disease. Pak. J. Biol. Sci. 2008, 11, 1114–1119. [Google Scholar] [CrossRef]
- Duro-Castano, A.; Nebot, V.J.; Niño-Pariente, A.; Armiñán, A.; Arroyo-Crespo, J.J.; Paul, A.; Feiner-Gracia, N.; Albertazzi, L.; Vicent, M.J. Capturing “extraordinary” soft-assembled charge-like polypeptides as a strategy for nanocarrier design. Adv. Mater. 2017, 29, 1702888. [Google Scholar] [CrossRef]
- Duro-Castano, A.; Borrás, C.; Herranz-Pérez, V.; Blanco-Gandía, M.C.; Conejos-Sánchez, I.; Armiñán, A.; Mas-Bargues, C.; Inglés, M.; Miñarro, J.; Rodríguez-Arias, M.; et al. Targeting Alzheimer’s disease with multimodal polypeptide-based nanoconjugates. Sci. Adv. 2021, 7, eabf9180. [Google Scholar] [CrossRef]
- Mortiboys, H.; Furmston, R.; Bronstad, G.; Aasly, J.; Elliott, C.; Bandmann, O. UDCA exerts beneficial effect on mitochondrial dysfunction in LRRK2(G2019S) carriers and in vivo. Neurology 2015, 85, 8468–8552. [Google Scholar] [CrossRef]
- Bonferoni, M.C.; Ferraro, L.; Pavan, B.; Beggiato, S.; Cavalieri, E.; Giunchedi, P.; Dalpiaz, A. Uptake in the central nervous system of geraniol oil encapsulated in chitosan oleate following nasal and oral administration. Pharmaceutics 2019, 11, 106. [Google Scholar] [CrossRef]
- Truzzi, E.; Rustichelli, C.; de Oliveira Junior, E.R.; Ferraro, L.; Maretti, E.; Graziani, D.; Botti, G.; Beggiato, S.; Iannuccelli, V.; Lima, E.M.; et al. Nasal biocompatible powder of Geraniol oil complexed with cyclodextrins for neurodegenerative diseases: Physicochemical characterization and in vivo evidences of nose to brain delivery. J. Control. Release 2021, 335, 91–202. [Google Scholar] [CrossRef]
- Price, R.L.; Chiocca, E.A. Evolution of malignant glioma treatment: From chemotherapy to vaccines to viruses. Neurosurgery 2014, 61 (Suppl. 1), 74–83. [Google Scholar] [CrossRef]
- Gao, H. Progress and perspectives on targeting nanoparticles for brain drug delivery. Acta Pharm. Sin. B 2016, 6, 268–286. [Google Scholar] [CrossRef]
- Qin, L.; Wang, C.Z.; Fan, H.J.; Zhang, C.J.; Zhang, H.W.; Lv, M.H.; Cui, S.D. A dual-targeting liposome conjugated with transferrin and arginine-glycine-aspartic acid peptide for glioma-targeting therapy. Oncol. Lett. 2014, 8, 2000–2006. [Google Scholar] [CrossRef]
- Pardridge, W.M. Brain drug development and brain drug targeting. Pharm. Res. 2007, 24, 1729–1732. [Google Scholar] [CrossRef]
- Zangui, M.; Atkin, S.L.; Majeed, M.; Sahebkar, A. Current evidence and future perspectives for curcumin and its analogues as promising adjuncts to oxaliplatin: State-of-the-art. Pharmacol. Res. 2019, 141, 343–356. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, P.; Law, S.; Tian, H.; Leung, W.; Xu, C. Preventive effect of curcumin against chemotherapy-induced side-effects. Front. Pharmacol. 2018, 9, 1374. [Google Scholar] [CrossRef]
- Tønnesen, H.H. Solubility, chemical and photochemical stability of curcumin in surfactant solutions. Studies of curcumin and curcuminoids, XXVIII. Pharmazie 2002, 57, 820–824. [Google Scholar]
- Sun, Z.J.; Sun, B.; Tao, R.B.; Xie, X.; Lu, X.L.; Dong, D.L. A poly(glycerol-sebacate-curcumin) polymer with potential use for brain gliomas. J. Biomed. Mater Res. A 2013, 101, 253–260. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer. 2010, 10, 9–22. [Google Scholar] [CrossRef]
- Mas-Moruno, C.; Rechenmacher, F.; Kessler, H. Cilengitide: The first anti-angiogenic small molecule drug candidate design, synthesis and clinical evaluation. Anticancer Agents Med. Chem. 2010, 10, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, X.; Hua, H.; Wang, A.; Liu, W.; Li, Y.; Fu, F.; Shi, Y.; Sun, K. Cyclic hexapeptide-conjugated nanoparticles enhance curcumin delivery to glioma tumor cells and tissue. Int. J. Nanomed. 2017, 12, 5717–5732. [Google Scholar] [CrossRef] [PubMed]
- Pavan, B.; Paganetto, G.; Rossi, D.; Dalpiaz, A. Multidrug resistance in cancer or inefficacy of neuroactive agents: Innovative strategies to inhibit or circumvent the active efflux transporters selectively. Drug Discov. Today 2014, 19, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Basso, J.; Mendes, M.; Silva, J.; Sereno, J.; Cova, T.; Oliveira, R.; Fortuna, A.; Castelo-Branco, M.; Falcão, A.; Sousa, J.; et al. Peptide-lipid nanoconstructs act site-specifically towards glioblastoma growth impairment. Eur. J. Pharm. Biopharm. 2020, 155, 177–189. [Google Scholar] [CrossRef]
- Sharma, M.; Sahu, K.; Singh, S.P.; Jain, B. Wound healing activity of curcumin conjugated to hyaluronic acid: In vitro and in vivo evaluation. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1009–1017. [Google Scholar] [CrossRef]
- Balendiran, G.K.; Dabur, R.; Fraser, D. The role of glutathione in cancer. Cell Biochem. Funct. 2004, 22, 343–352. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Tian, C.; Asghar, S.; Xu, Y.; Chen, Z.; Zhang, J.; Ping, Q.; Xiao, Y. Tween 80-modified hyaluronic acid-ss-curcumin micelles for targeting glioma: Synthesis, characterization and their in vitro evaluation. Int. J. Biol. Macromol. 2018, 120, 2579–2588. [Google Scholar] [CrossRef]
- Gastaldi, L.; Battaglia, L.; Peira, E.; Chirio, D.; Muntoni, E.; Solazzi, I.; Gallarate, M.; Dosio, F. Solid lipid nanoparticles as vehicles of drugs to the brain: Current state of the art. Eur. J. Pharm. Biopharm. 2014, 87, 433–444. [Google Scholar] [CrossRef]
- Sun, W.; Xie, C.; Wang, H.; Hu, Y. Specific role of polysorbate 80 coating on the targeting of nanoparticles to the brain. Biomaterials 2004, 25, 3065–3071. [Google Scholar] [CrossRef]
- Peter, K.; Kar, S.K.; Gothalwal, R.; Gandhi, P. Curcumin in Combination with other adjunct therapies for brain tumor treatment: Existing knowledge and blueprint for future research. Int. J. Mol. Cell. Med. 2021, 10, 163–181. [Google Scholar] [CrossRef]
- Saul, J.M.; Annapragada, A.; Natarajan, J.V.; Bellamkonda, R.V. Controlled targeting of liposomal doxorubicin via the folate receptor in vitro. J. Control. Release 2003, 92, 49–67. [Google Scholar] [CrossRef]
- Miura, Y.; Takenaka, T.; Toh, K.; Wu, S.; Nishihara, H.; Kano, M.R.; Ino, Y.; Nomoto, T.; Matsumoto, Y.; Koyama, H.; et al. Cyclic RGD-linked polymeric micelles for targeted delivery of platinum anticancer drugs to glioblastoma through the blood-brain tumor barrier. ACS Nano 2013, 7, 8583–8592. [Google Scholar] [CrossRef]
- Zhao, B.X.; Zhao, Y.; Huang, Y.; Luo, L.M.; Song, P.; Wang, X.; Chen, S.; Yu, K.F.; Zhang, X.; Zhang, Q. The efficiency of tumor-specific pH-responsive peptide-modified polymeric micelles containing paclitaxel. Biomaterials 2012, 33, 2508–2520. [Google Scholar] [CrossRef]
- Zhao, Y.; Ren, W.; Zhong, T.; Zhang, S.; Huang, D.; Guo, Y.; Yao, X.; Wang, C.; Zhang, W.Q.; Zhang, X.; et al. Tumor-specific pH-responsive peptide-modified pH-sensitive liposomes containing doxorubicin for enhancing glioma targeting and anti-tumor activity. J. Control. Release 2016, 222, 56–66. [Google Scholar] [CrossRef]
- Gabay, M.; Weizman, A.; Zeineh, N.; Kahana, M.; Obeid, F.; Allon, N.; Gavish, M. Liposomal carrier conjugated to APP-derived peptide for brain cancer treatment. Cell. Mol. Neurobiol. 2021, 41, 1019–1029. [Google Scholar] [CrossRef]
- Zhu, L.; Li, R.; Jiao, S.; Wei, J.; Yan, Y.; Wang, Z.A.; Li, J.; Du, Y. Blood-brain barrier permeable chitosan oligosaccharides interfere with β-amyloid aggregation and alleviate β-amyloid protein mediated neurotoxicity and neuroinflammation in a dose- and degree of polymerization-dependent manner. Mar. Drugs 2020, 18, 488. [Google Scholar] [CrossRef]
- Jiang, H.; Shang, X.; Wu, H.; Huang, G.; Wang, Y.; Al-Holou, S.; Gautam, S.C.; Chopp, M. Combination treatment with resveratrol and sulforaphane induces apoptosis in human U251 glioma cells. Neurochem. Res. 2010, 35, 152–161. [Google Scholar] [CrossRef]
- Castino, R.; Pucer, A.; Veneroni, R.; Morani, F.; Peracchio, C.; Lah, T.T.; Isidoro, C. Resveratrol reduces the invasive growth and promotes the acquisition of a long-lasting differentiated phenotype in human glioblastoma cells. J. Agric. Food Chem. 2011, 59, 4264–4272. [Google Scholar] [CrossRef]
- Yousef, M.; Vlachogiannis, I.A.; Tsiani, E. Effects of resveratrol against lung cancer: In vitro and in vivo studies. Nutrients 2017, 9, 1231. [Google Scholar] [CrossRef]
- Sanna, V.; Roggio, A.M.; Siliani, S.; Piccinini, M.; Marceddu, S.; Mariani, A.; Sechi, M. Development of novel cationic chitosan-and anionic alginate-coated poly(D,L-lactide-co-glycolide) nanoparticles for controlled release and light protection of resveratrol. Int. J. Nanomed. 2012, 7, 5501–5516. [Google Scholar] [CrossRef]
- Behin, A.; Hoang-Xuan, K.; Carpentier, A.F.; Delattre, J.Y. Primary brain tumours in adults. Lancet 2003, 361, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Q.; Qu, Y.H.; Ke, W.L.; Zhu, J.H.; Pei, Y.Y.; Jiang, C. Efficient gene delivery targeted to the brain using a transferrin-conjugated polyethyleneglycol-modified polyamidoamine dendrimer. FASEB J. 2007, 21, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, A.; Larocca, L.M.; Deaglio, S.; Finisguerra, V.; Boe, A.; Raggi, C.; Ricci-Vitani, L.; Pierconti, F.; Malavasi, F.; De Maria, R.; et al. Transferrin receptor 2 is frequently and highly expressed in glioblastomas. Transl. Oncol. 2010, 3, 123–134. [Google Scholar] [CrossRef]
- Guo, W.; Li, A.; Jia, Z.; Yuan, Y.; Dai, H.; Li, H. Transferrin modified PEG-PLA-resveratrol conjugates: In vitro and in vivo studies for glioma. Eur. J. Pharmacol. 2013, 718, 41–47. [Google Scholar] [CrossRef]
- Hong, S.H.; Ismail, I.A.; Kang, S.M.; Han, D.; Kwon, B.M. Cinnamaldehydes in cancer chemotherapy. Phytother. Res. 2016, 30, 754–767. [Google Scholar] [CrossRef]
- Zhao, H.; Xie, Y.; Yang, Q.; Cao, Y.; Tu, H.; Cao, W.; Wang, S. Pharmacokinetic study of cinnamaldehyde in rats by GC-MS after oral and intravenous administration. J. Pharm. Biomed. Anal. 2014, 89, 150–157. [Google Scholar] [CrossRef]
- Merzak, A.; Koochekpour, S.; Fillion, M.P.; Fillion, G.; Pilkington, G.J. Expression of serotonin receptors in human fetal astrocytes and glioma cell lines: A possible role in glioma cell proliferation and migration. Brain Res. Mol. Brain Res. 1996, 41, 1–7. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Design | Formulation | In Vitro Cellular or Tissue Models | In Vivo Evaluation | Ref. |
|---|---|---|---|---|
| Ferulic Acid | ||||
| Use of inflammatory cells as carriers of ferulic acid-loaded liposomes decorated with peptide substrates of integrin receptors expressed by cells taken up by the brain in response to neural inflammation | Ferulic acid-loaded liposomes decorated with RGD peptide using conjugation to cholesterol via a succinic spacer | Pro-monocytic U937 cell line for binding and antioxidant studies | Intravenous administration to rat models of brain inflammation (intra-striatal micro-injections of human recombinant IL-1β) | [77,78] |
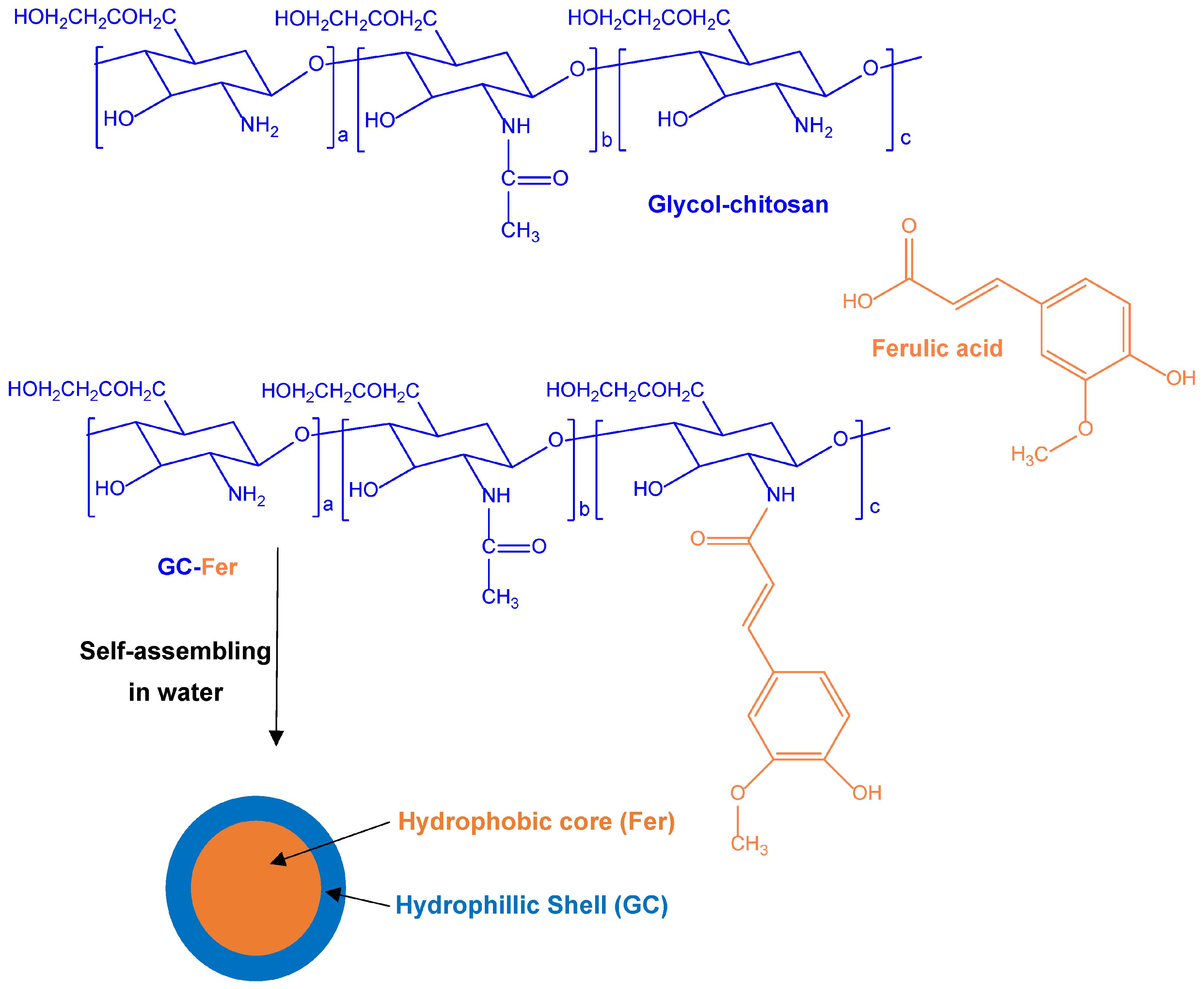
| Ferulic acid and glycol chitosan conjugates self-assembled as nanoparticles for a co-delivery strategy of injured spinal cord restoration | Self-assembled nanoparticles obtained using amidic conjugates of ferulic acid with glycol–chitosan | Rat primary neurons for protection studies against glutamate-induced excitotoxicity | Model rats of spinal cord contusion | [26] |
| Synthesis of prodrugs recognised by L-type amino acid transporters (LAT1) | Prodrug of ferulic acid obtained using amidic or ester conjugation with amino acids recognised by LAT1 | Retinal pigment epithelia ARPE-19 cells; mice primary astrocytes from cortex and hippocampi | Intraperitoneal administration to mice | [80,81] |
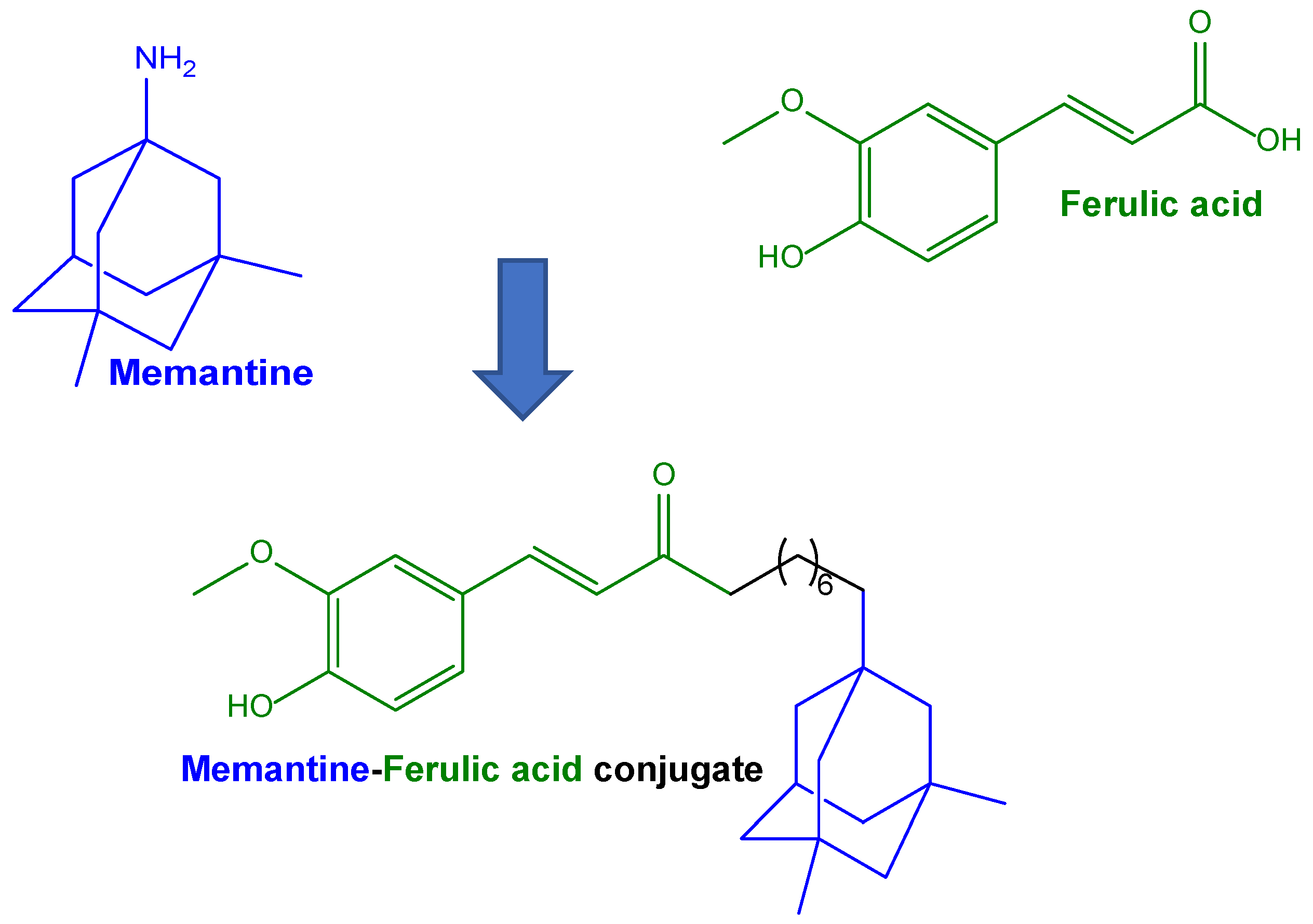
| Conjugation of ferulic acid with memantine, which is able to limit the overactivation of N-methyl-D-aspartate receptors | Conjugates of ferulic acid with memantine via a hexamethylene spacer | SH-SY5Y cells as neuronal model | Conjugate proposed for potential use against AD | [82] |
| Methyl ester derivative of ferulic acid as a prodrug able to increase loading in solid lipid microparticles | Methyl-ferulate-loaded solid lipid microparticles (SLMs) based on tristearin or stearic acid | PC12 cells chosen as a model for neural differentiation | SLMs proposed for potential use as a nasal formulation for methyl-ferulate brain-targeting | [71] |
| Caffeic acid | ||||
| Ionic complex between caffeic acid and chitosan carboxymethyl dextran-b-poly(ethylene glycol) for anticancer drug delivery | Nanoparticles of caffeic acid conjugated with chitosan and carboxymethyl dextran-b-poly(ethylene glycol) loading doxorubicin | CT26 cells (murine colorectal carcinoma cell line) | [86] | |
| Caffeic acid conjugated to stearic acid by means of ethylenediamine as a linker to enhance the aqueous dispersibility of micelles | Nanosized-micelles encapsulating budesonide for rectal administration | hTERT-BJ cells (fibroblast cells) | Colitic mouse model | [90] |
| Gallic acid | ||||
| Conjugation of gallic acid with gelatine to improve antioxidant properties | Conjugates of gallic acid linked to oxidised dextran to obtain an injectable hydrogel | HT22 cells (neuronal cell line) | Traumatic brain injury mouse model | [98] |
| Resveratrol | ||||
| PLGA nanoparticles able to induce the brain’s uptake of resveratrol in cases of PD, due to the presence on the surface of lactoferrin | Resveratrol-loaded PLGA nanoparticles decorated with lactoferrin for intravenous injection | SH-SY5Y cells (neuroblastoma cells) | MPTP-induced PD mice model. | [110] |
| PLA nanoparticles loaded with resveratrol conjugated with ligands for low-density lipoprotein receptors to cross the BBB via receptor-mediated transcytosis | PLA-coated mesoporous silica nanoparticles | HAPI cells (brain-derived microglial cell line) | [111] | |
| Curcumin | ||||
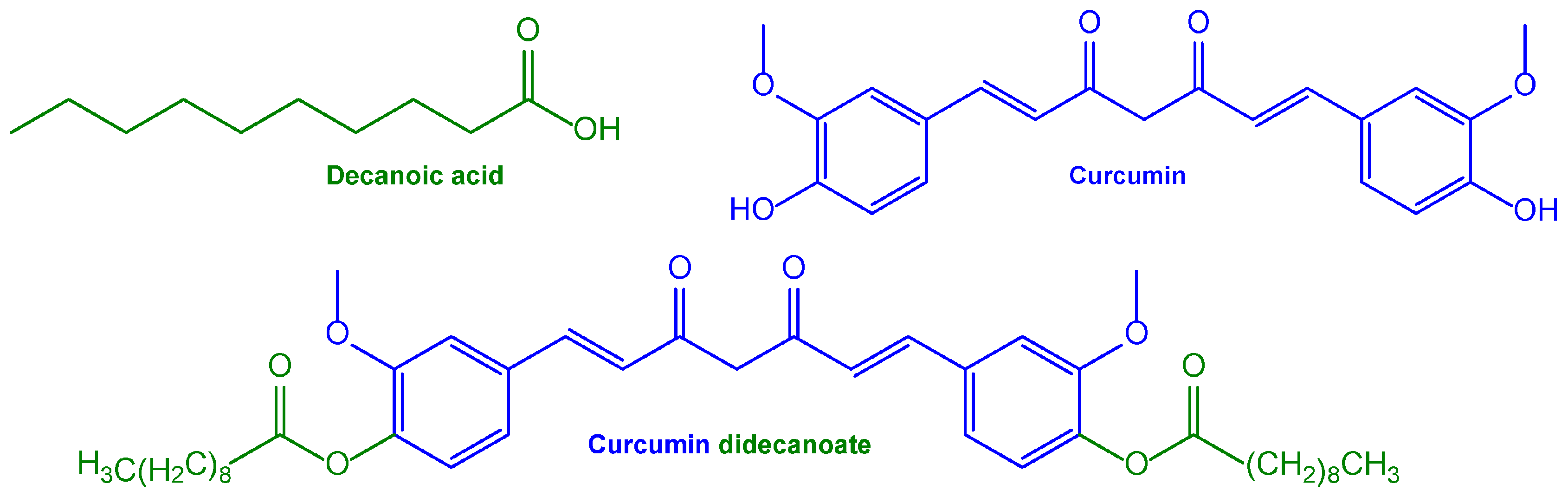
| Nanosuspension based on a curcumin prodrug for intramuscular administration with consequent systemic curcumin’s prolonged release | Nanosuspension based on a curcumin prodrug obtained using ester conjugation with decanoic acid | Intramuscular administration to male Wistar rats | [127] | |
| PLGA nanoparticles able to induce curcumin uptake in neurons via the presence of the targeting moiety-Tet-1 peptide on their surface | Curcumin-loaded nanoparticles decorated on their surface with the Tet-1 peptide (HLNILSTLWKYR) | LAG cell lines (mouse fibroblast-like connective tissue) for toxicity studies; GI-1 glioma cells for uptake studies | Nanocarriers are proposed as a potential formulation for the treatment of AD | [125] |
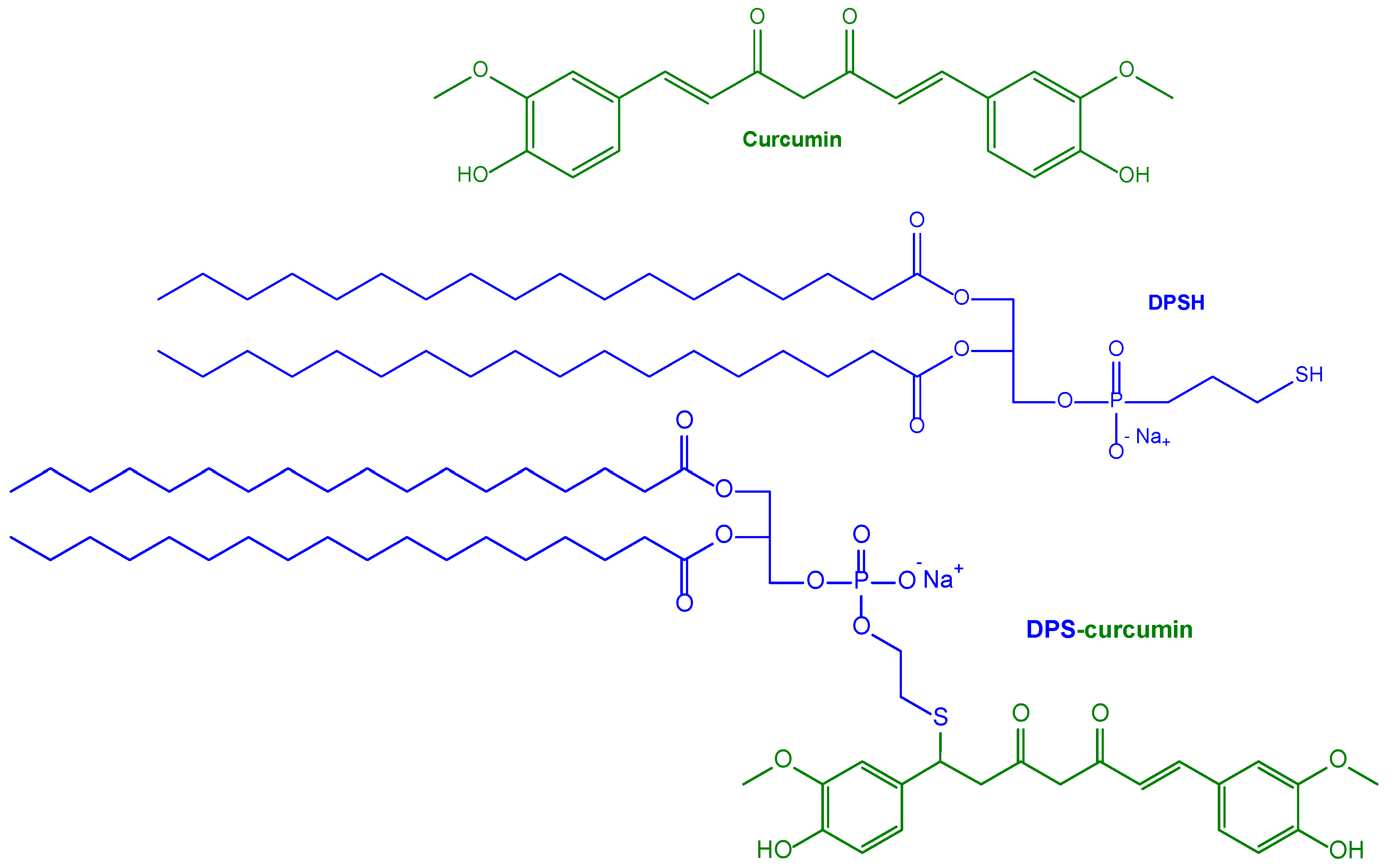
| Nanoliposomes with high amyloid affinity using surface decoration with curcumin for the AD treatment | SUV obtained in the presence 1,2-dipalmitoyl-sn-glycero-3-phosphothioethanol conjugated to curcumin | HEK cells, control SH-SY5Y cells and hAPP SH-SY5Y cells stably overexpressing the human APP gene causing familial AD | Transgenic mouse AD model (APPxPS1 mice) for injection into the brain of the nanocarrier | [126] |
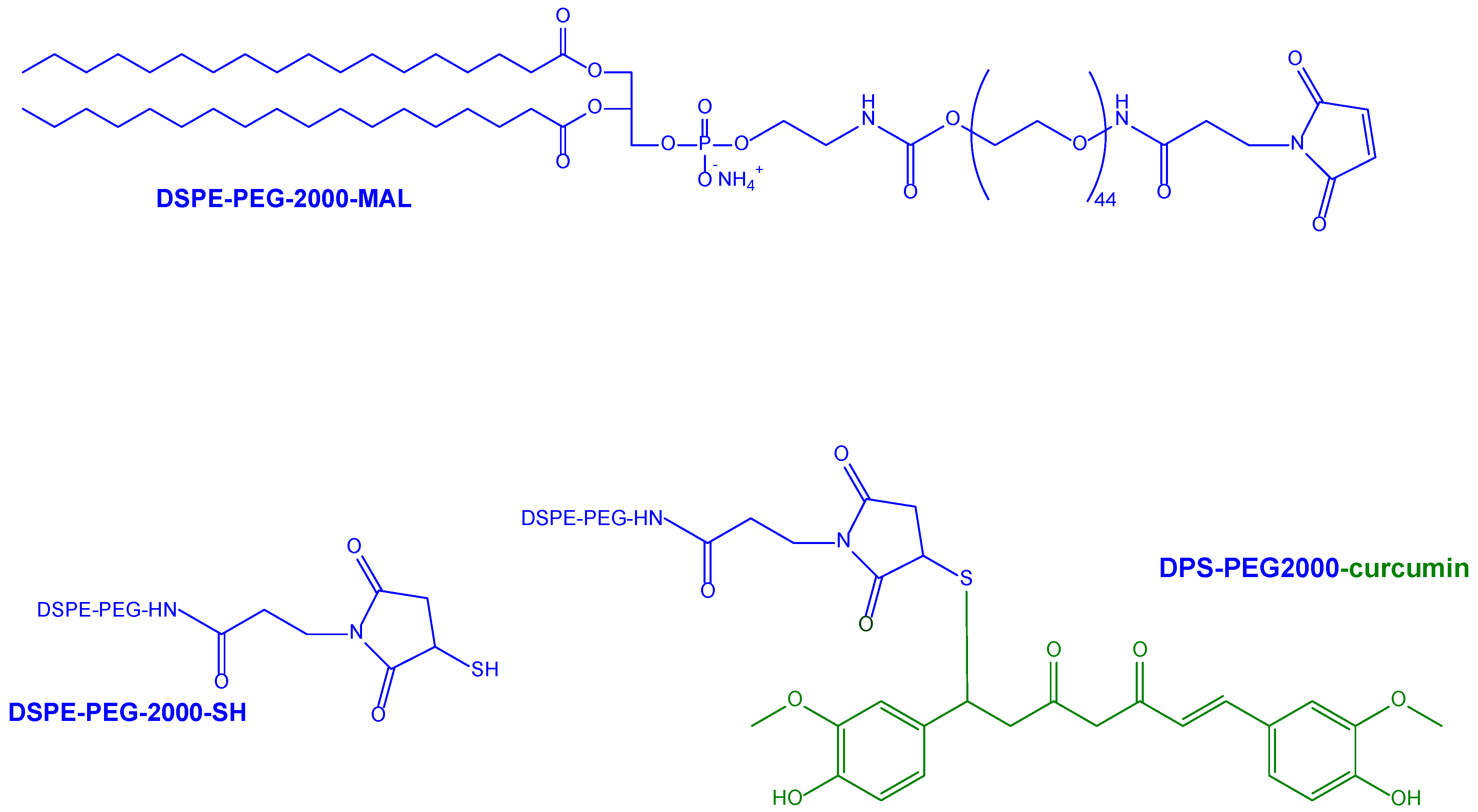
| Previous nanoliposomes further implemented with anti-transferrin antibody decoration for BBB targeting | SUV obtained in the presence of phospholipids conjugated with PEG2000 as anchoring points for curcumin or anti-transferrin antibodies | Human brain capillary endothelial cells (hCMEC/D3) as an in vitro model for BBB permeation studies | Proposed as potential formulations for the treatment of AD | [128] |
| Multifunctional liposomes for co-delivery of curcumin and NGF to the brain | Curcumin-loaded and NGF liposomes obtained in the presence of cardiolipin in their bilayer structure, and decorated with WGA anchored to a PEG spacer | Human brain microvascular endothelial cells (HBMECs) and astrocytes (HAs) for BBB permeation studies; SK-N-MC cells for the Aβ toxicity model | Proposed as potential pharmacotherapy vehicles for AD therapy | [130] |
| PLGA nanoparticles provided with GSH functionalization to increase the neuronal internalization of curcumin. | Curcumin-loaded PLGA nanoparticles decorated with PEG spacers inserted between polymer and GSH, which was conjugated in the presence of a maleimide moiety | SK-N-SH cells, a human neuroblastoma cell line, as a neuronal internalization model | [131] | |
| PLGA nanoparticles functionalised with the brain-targeting peptide CRT for the co-delivery of curcumin and the Aβ generation inhibitor S1 to the brain | Curcumin and S1-loaded PLGA nanoparticles functionalised with the CRT peptide by conjugation to a PEG spacer | Neuroblastoma SH-SY5Y cells for cytotoxicity studies; brain microvascular bEnd.3 cells as a model for BBB permeation studies | Intravenous administration to male BALB/c nude mice; intraperitoneal injection to AD model (APP/PS1dE9) mice | [132] |
| PLGA nanoparticles functionalised with B6 peptide for curcumin drug delivery to the CNS | Curcumin-loaded PLGA nanoparticles functionalised with the B6 peptide by conjugation to a PEG spacer | Immortalised mouse hippocampal HT-22 cell line for intracellular studies | Intraperitoneal administration to APP/PS1 transgenic mice for spatial learning and memory capability studies | [133] |
| Evaluation the uptake of nanoparticles in an in vitro model of Huntington’s disease and cell susceptibility to apoptosis | Nanoparticles based on an amphiphilic conjugate of hyaluronic acid and palmitic acid loaded with curcumin | Striatal-derived immortalised cell line expressing Huntington’s mutation (STHdh111/111) | [135] | |
| Effects of curcumin conjugate on spatial memory and oxidative stress in a rat model of multiple sclerosis | Conjugate of curcumin with linoleic acid | Intracerebroventricular injection into male Wistar rats after ethidium bromide treatment as an animal model of multiple sclerosis | [136] | |
| Curcumin encapsulated in bifunctional microemulsions as a targeting agent for Aβ and an inhibitor of fibrils aggregation | Microemulsions based on a modified Pluronic conjugated to the peptide KLVFF for specific binding to Aβ fibrils | Ex vivo permeation study on porcine nasal mucosa; proposed as a potential nasal formulation | [137] | |
| 7,8-dihydroxyflavone | ||||
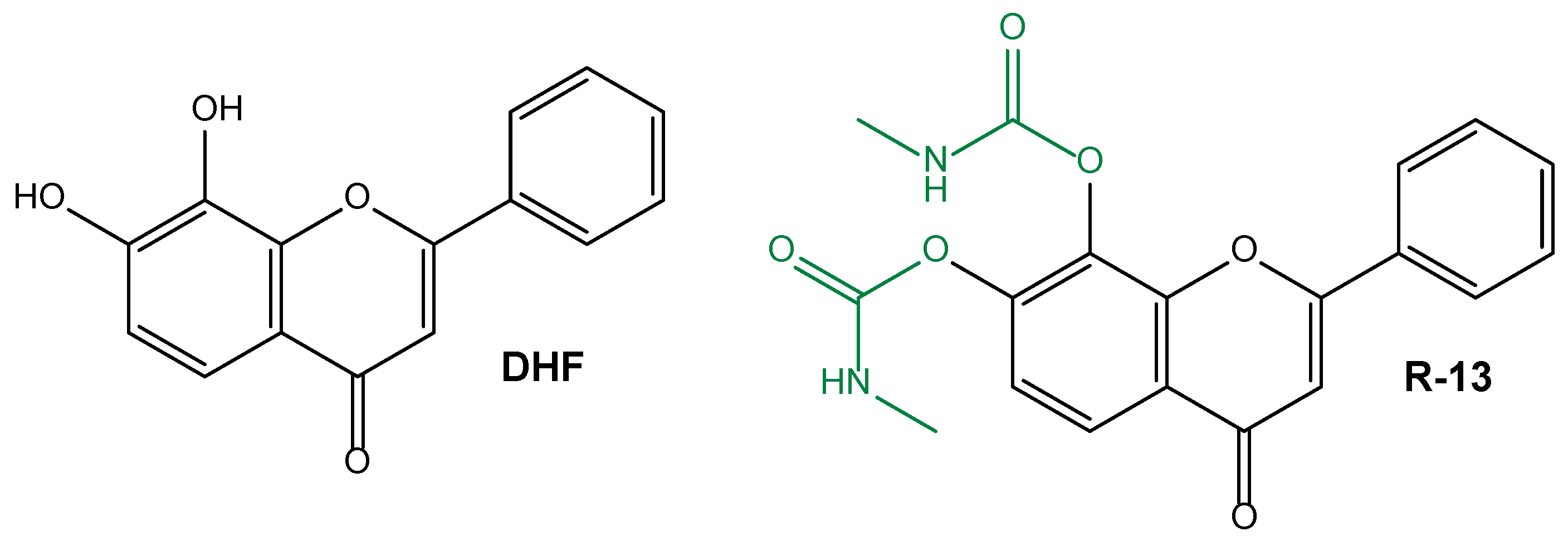
| Synthesis of a prodrug able to increase the poor oral bioavailability and half-life of dihydroxyflavone for AD treatment | R-13 prodrug | Caco-2 cell line as intestinal model for permeability studies | 5XFAD mice as an in vivo model of AD | [138] |
| Use of R13 prodrug for the oral treatment of peripheral nerve injury | R-13 prodrug | C57B6 wild-type mice | [143] | |
| Epigallocatechin gallate | ||||
| Formulation of liposomes with the ability to cross the BBB for the co-delivery of EGCG and resveratrol against PD | Resveratrol and EGCG-loaded liposomes decorated on their surface with leptin | Human astrocytes, brain vascular pericytes and brain microvascular endothelial cells for an in vitro model of BBB; SH-SY5Y cells insulted with MPP as an in vitro neurodegenerative model | [147] | |
| Quercetin | ||||
| Evaluation the ability of α-tocopherol to promote the transport of quercetin across the BBB | Co-administration of quercetin and α-tocopherol | Oral administration to male Sprague Dawley rats | [146] | |
| Water-soluble quercetin conjugate to increase the BBB permeability and to potentially treat cerebral ischemia | Conjugate of quercetin with hyaluronic acid targeted with a penetrating polypeptide, SS31 | Human neural cell lines (PC12 and SH-SY5Y cells) and primary cortical neurons | Intraperitoneal injection or through tail vein to male Sprague Dawley rats after middle cerebral artery occlusion as a model of permanent cerebral ischemia | [165] |
| Quercetin conjugate for improving diabetes-related memory impairment | Conjugate of quercetin with dextran-coated superparamagnetic iron oxide nanoparticles | Oral administration by gavage to male Wistar rats treated with streptozotocin to induce Type 1 diabetes | [166,167] | |
| Quercetin conjugate to enhance the its bioavailability in the brain for the potential treatment of neurodegenerative disorders | Conjugate of quercetin with dextran-coated superparamagnetic iron oxide nanoparticles | Oral administration by gavage to male Wistar rats | [168] | |
| In vitro evaluation of antitoxic properties of free quercetin and quercetin conjugate | Conjugate of quercetin with dextran-coated superparamagnetic iron oxide nanoparticles | Human neural cell lines (PC12 cells) | [169] | |
| In vivo evaluation of anticonvulsant activity of quercetin conjugate | Conjugate of quercetin with superparamagnetic iron oxide nanoparticles coated with β-cyclodextrin and pluronic F68 | Intraperitoneal injection to male NMRI mice treated with pentylenetetrazole as a kindling model | [170] | |
| Genistein | ||||
| Genistein-carrying polypeptide conjugate with propargylamine and Angiopep-2 to enhance the ability to cross the BBB | Genistein-loaded PGA- nanocarrier modified with neuroprotective propargylamine residues together with Angiopep-2 | Neuroblastoma SH-SY5Y cell line | Mouse model of AD mice | [178] |
| Geraniol | ||||
| Nasal formulation as solid lipid nanoparticles loaded with a prodrug obtained via the conjugation of geraniol with ursodeoxycholic acid | Solid lipid nanoparticles based on compritol loaded with an ester conjugate of geraniol and ursodeoxycholic acid | Rat liver and brain homogenates | Intravenous and nasal administration to rats | [33] |
| Geraniol encapsulated in chitosan oleate as oral and nasal nanoformulations | Geraniol nanoemulsion obtained in the presence of chitosan oleate as surfactant | Oral and nasal administration to rats | [180] | |
| Design | Formulation | In Vitro Cellular Models | In Vivo Evaluation | Ref. |
|---|---|---|---|---|
| Curcumin | ||||
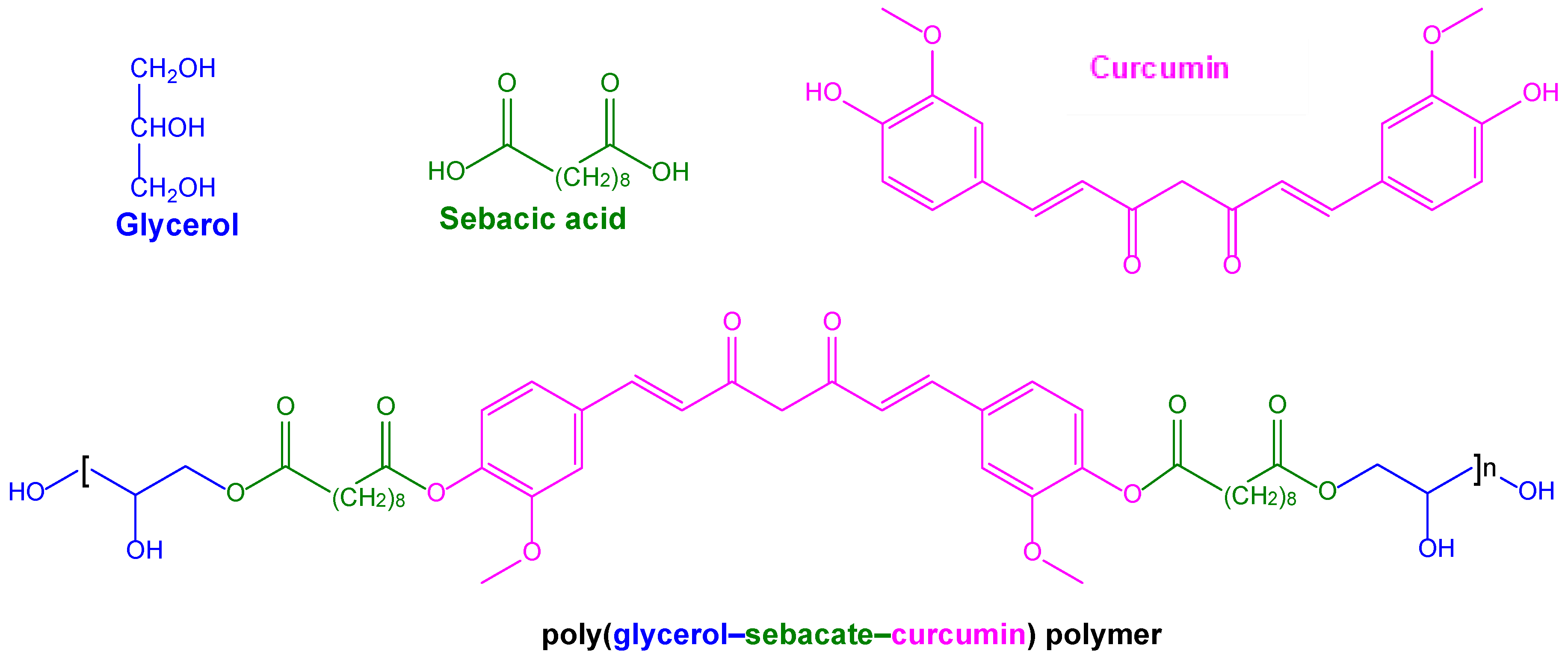
| Conjugation of curcumin into biocompatible and biodegradable polymers for local treatment of tumours | Polymers based on poly(glycerol–sebacate–curcumin) unities | Glioblastoma U-87 and neuroblastoma T-98 cell lines | Proposed for potential use in the local treatment of glioma | [189] |
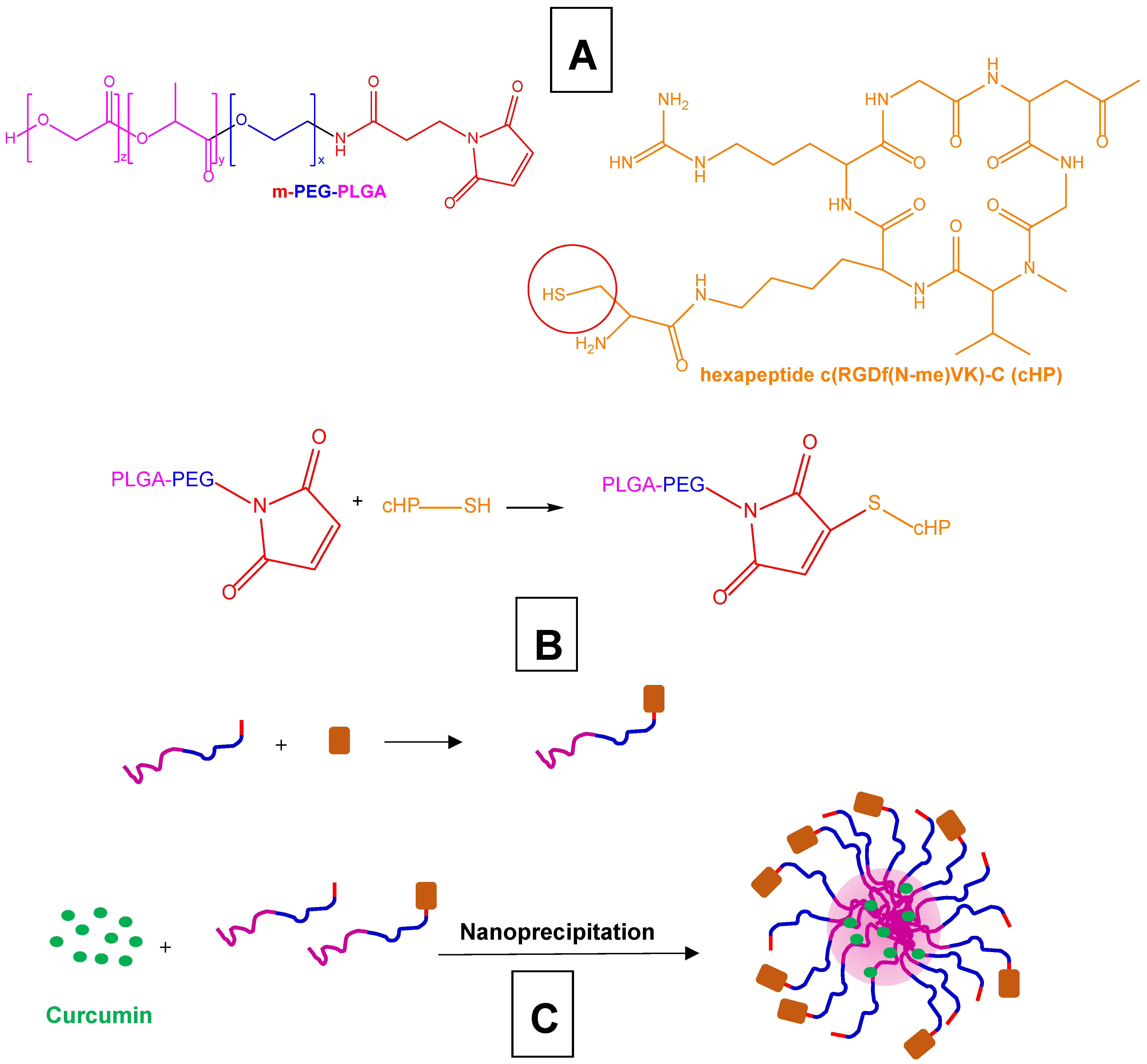
| Polymeric nanoparticles decorated with the esapeptide c(RGDf(N-me)VK)-C (cHP) for the targeting of integrins overexpressed by glioma | Curcumin-loaded PEG-PLGA nanoparticles decorated with cHP | C6 rat glioma cell line, C6 cell spheres | Intravenous administration to rats undergoing C6 cell transplantation | [192] |
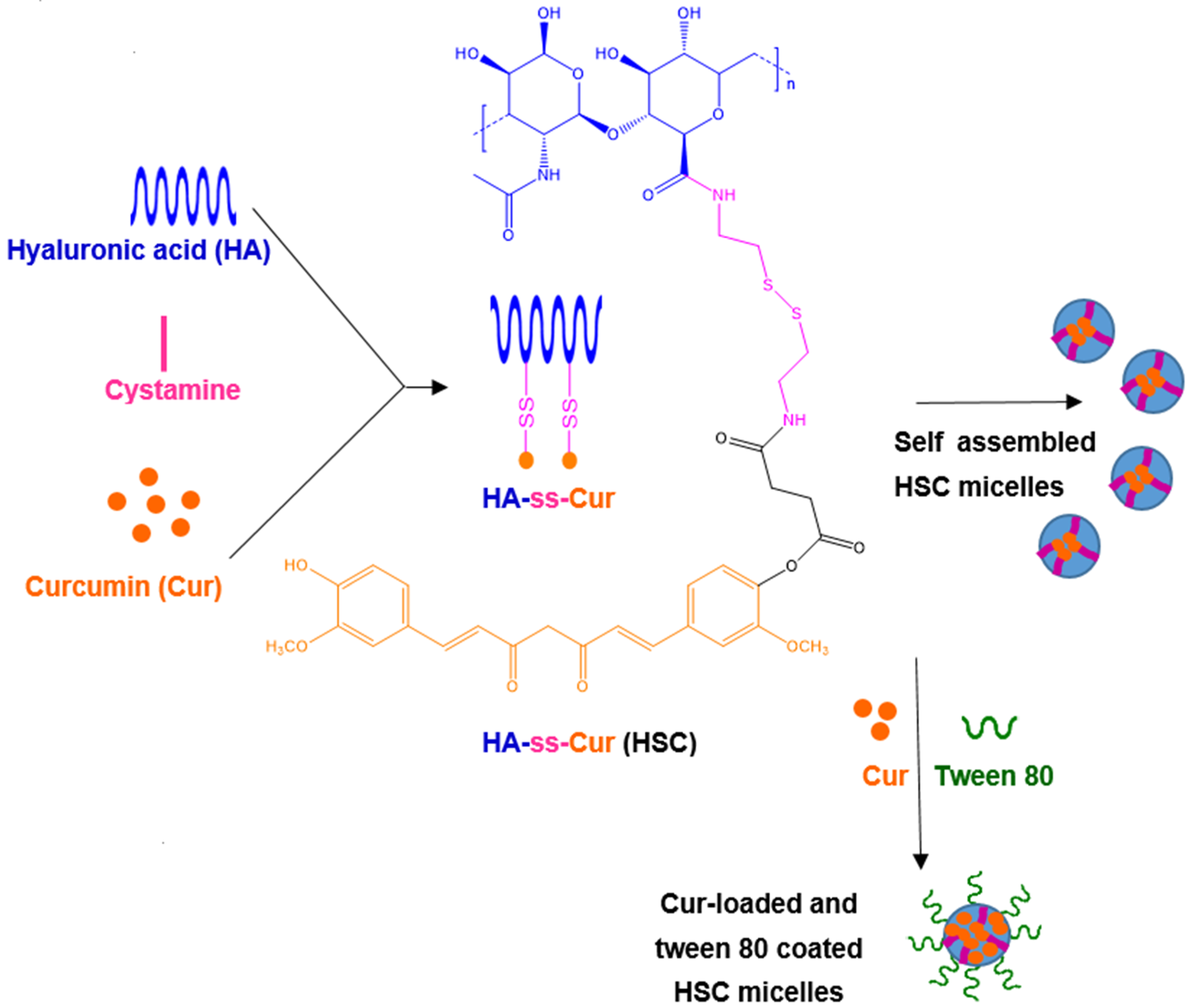
| Self-assembled nanoparticles obtained via conjugates of curcumin and hyaluronic acid via a GSH-sensitive disulphide bond. Hyaluronic acid can target glioma cells via CD44 markers. Tween 80 coating induces the BBB permeation. | Curcumin-loaded and Tween 80-coated nanoparticles obtained by self-assembling curcumin–hyaluronic acid conjugates. | Glioblastoma G422 cell line | Intravenous administration to glioma-bearing mice | [28,198] |
| Co-delivery of curcumin- and atorvastatin-loaded ultra-small nanostructured lipid carriers decorated with folic acid and peptides able to induce targeting and internalization in glioma cells | Curcumin- and atorvastatin-loaded ultra-small nanostructured lipid carriers bio-conjugated via electrostatic binding with hyaluronic acid conjugated with folic acid or specific peptides cRGDfK and H7K(R2)2 | Glioblastoma U-87 cells | Intraperitoneal administration to mice that underwent intracranial injection of U87 glioblastoma | [194] |
| Co-delivery of temozolomide-, curcumin-, and doxorubicin-loaded liposomes decorated with a peptide able to induce the BBB crossing | Temozolomide-, curcumin- and doxorubicin-loaded liposomes decorated with the pentapeptide RERMS | In vitro model of BBB, based on porcine brain-derived capillary endothelial cells | Injection to mice underwenting intracranial injection of human U87 glioblastoma to mice. | [206] |
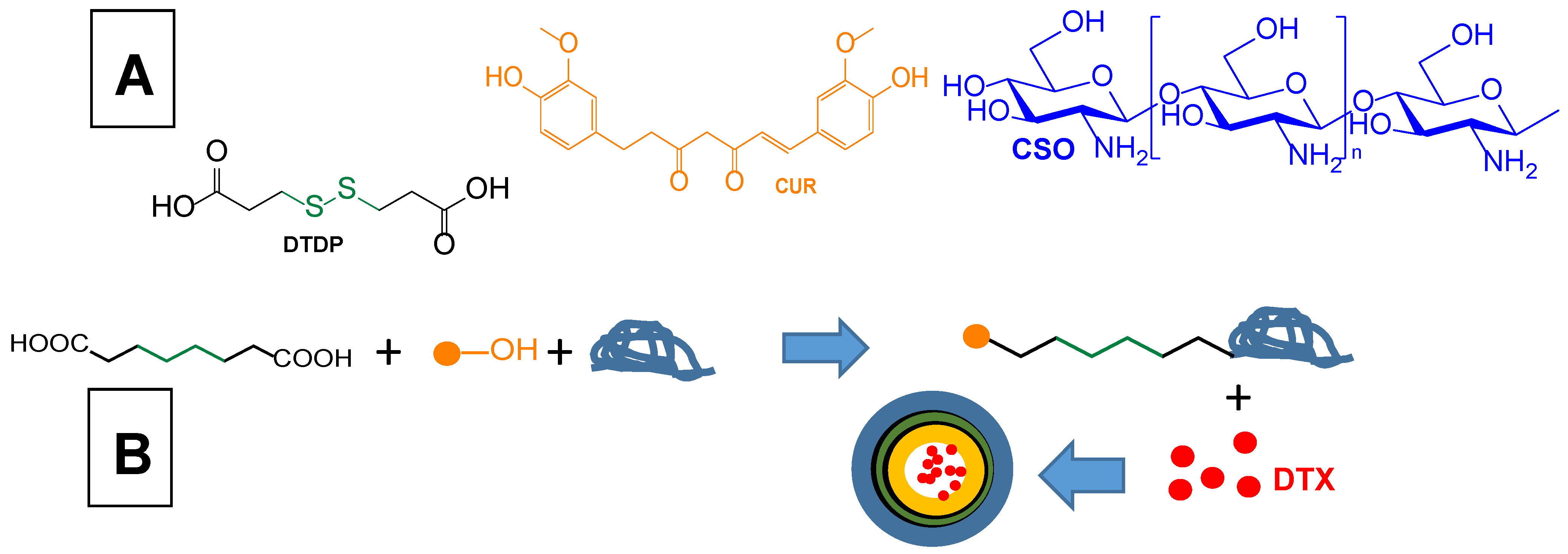
| Curcumin coupling via a disulphide bond with chitosan oligosaccharide (CSO) in order to obtain self- assembled nanomicelles loaded with docetaxel for co-delivery to glioma cells | Loaded docetaxel self-assembled nanomicelles based on conjugates of curcumin to CSO by a disulphide bond | Glioma C6 cell line | Intravenous administration to C6 tumour-bearing mouse model | [29] |
| Resveratrol | ||||
| Polymeric nanoparticles decorated with transferrin to increase BBB penetration | Resveratrol-loaded PEG-PLGA nanoparticles decorated with transferrin | Glioma C6 and glioblastoma U-87 cell lines | Intraperitoneal administration to glioma-bearing rats | [215] |
| Co-delivery of resveratrol and epirubicin-loaded polyfunctional liposomes able to cross the BBB and target glioma cells | Resveratrol- and epirubicin-loaded liposomes decorated with p-aminophenyl-α-D-manno-pyranoside (MAN) and germ agglutinin (WGA) | Mouse brain microvascular endothelial (bEnd.3) cells; glioma C6 cell lines; C6 glioma spheroids | Intravenous administration to glioma bearing rats | [30] |
| Cinnamaldehyde | ||||
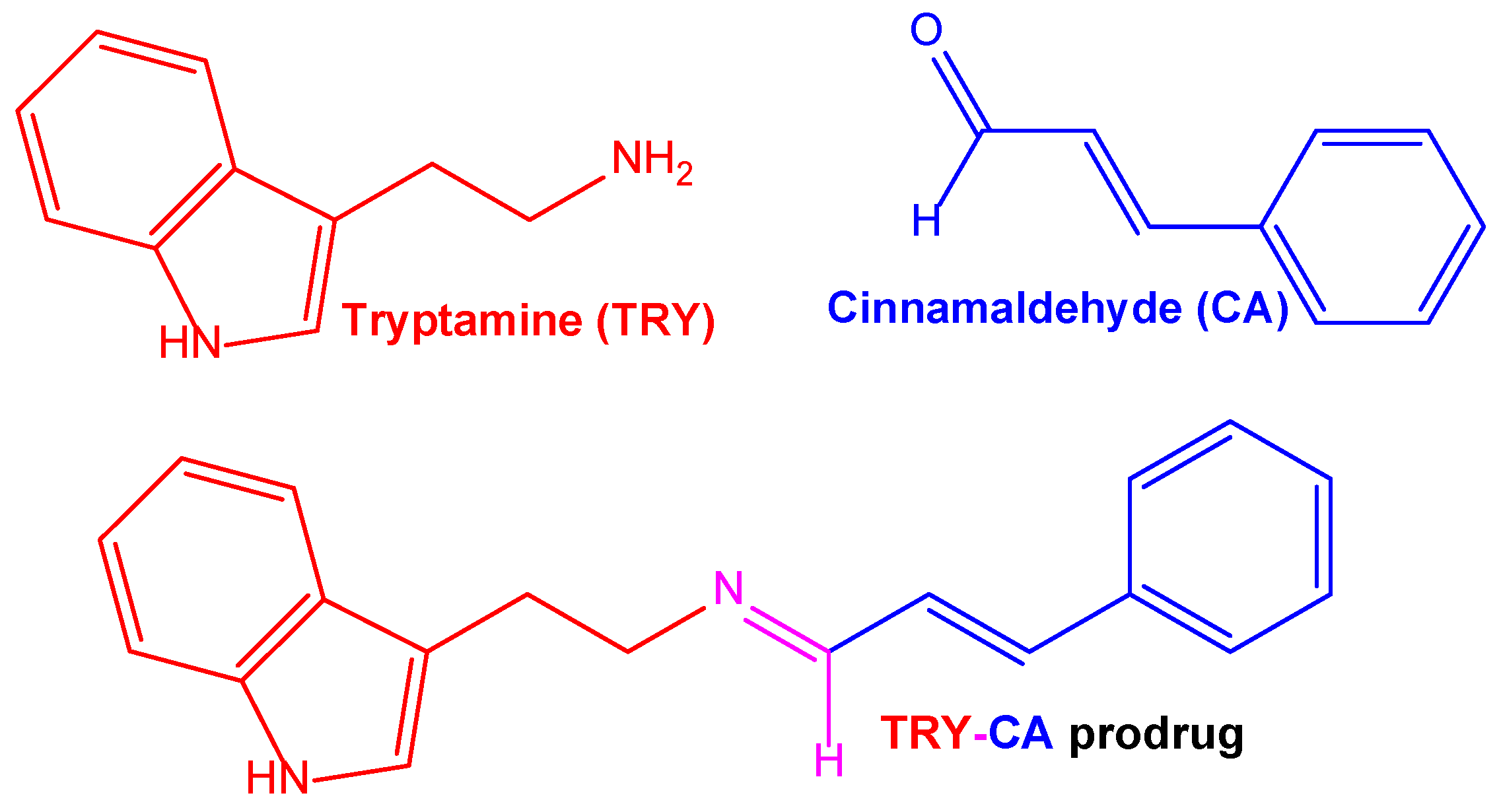
| Co-delivery of cinnamldehyde and tryptamine conjugated to each other via aldimine condensation and self-assembled as nanospheres | Self-assembled nanospheres of a prodrug of cinnamldehyde and tryptamine, obtained using their aldimine condensation | SH-SY5Y neuroblastoma cells; SH-SY5Y multicellular spheroids | Intravenous administration to mice | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rassu, G.; Sorrenti, M.; Catenacci, L.; Pavan, B.; Ferraro, L.; Gavini, E.; Bonferoni, M.C.; Giunchedi, P.; Dalpiaz, A. Conjugation, Prodrug, and Co-Administration Strategies in Support of Nanotechnologies to Improve the Therapeutic Efficacy of Phytochemicals in the Central Nervous System. Pharmaceutics 2023, 15, 1578. https://doi.org/10.3390/pharmaceutics15061578
Rassu G, Sorrenti M, Catenacci L, Pavan B, Ferraro L, Gavini E, Bonferoni MC, Giunchedi P, Dalpiaz A. Conjugation, Prodrug, and Co-Administration Strategies in Support of Nanotechnologies to Improve the Therapeutic Efficacy of Phytochemicals in the Central Nervous System. Pharmaceutics. 2023; 15(6):1578. https://doi.org/10.3390/pharmaceutics15061578
Chicago/Turabian StyleRassu, Giovanna, Milena Sorrenti, Laura Catenacci, Barbara Pavan, Luca Ferraro, Elisabetta Gavini, Maria Cristina Bonferoni, Paolo Giunchedi, and Alessandro Dalpiaz. 2023. "Conjugation, Prodrug, and Co-Administration Strategies in Support of Nanotechnologies to Improve the Therapeutic Efficacy of Phytochemicals in the Central Nervous System" Pharmaceutics 15, no. 6: 1578. https://doi.org/10.3390/pharmaceutics15061578