
Quality, Safety and Biological Studies on Campylanthus glaber Aerial Parts
 ,
,  ,
,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Botanical Quality Monographic Studies
2.1.1. Macroscopic Characterization
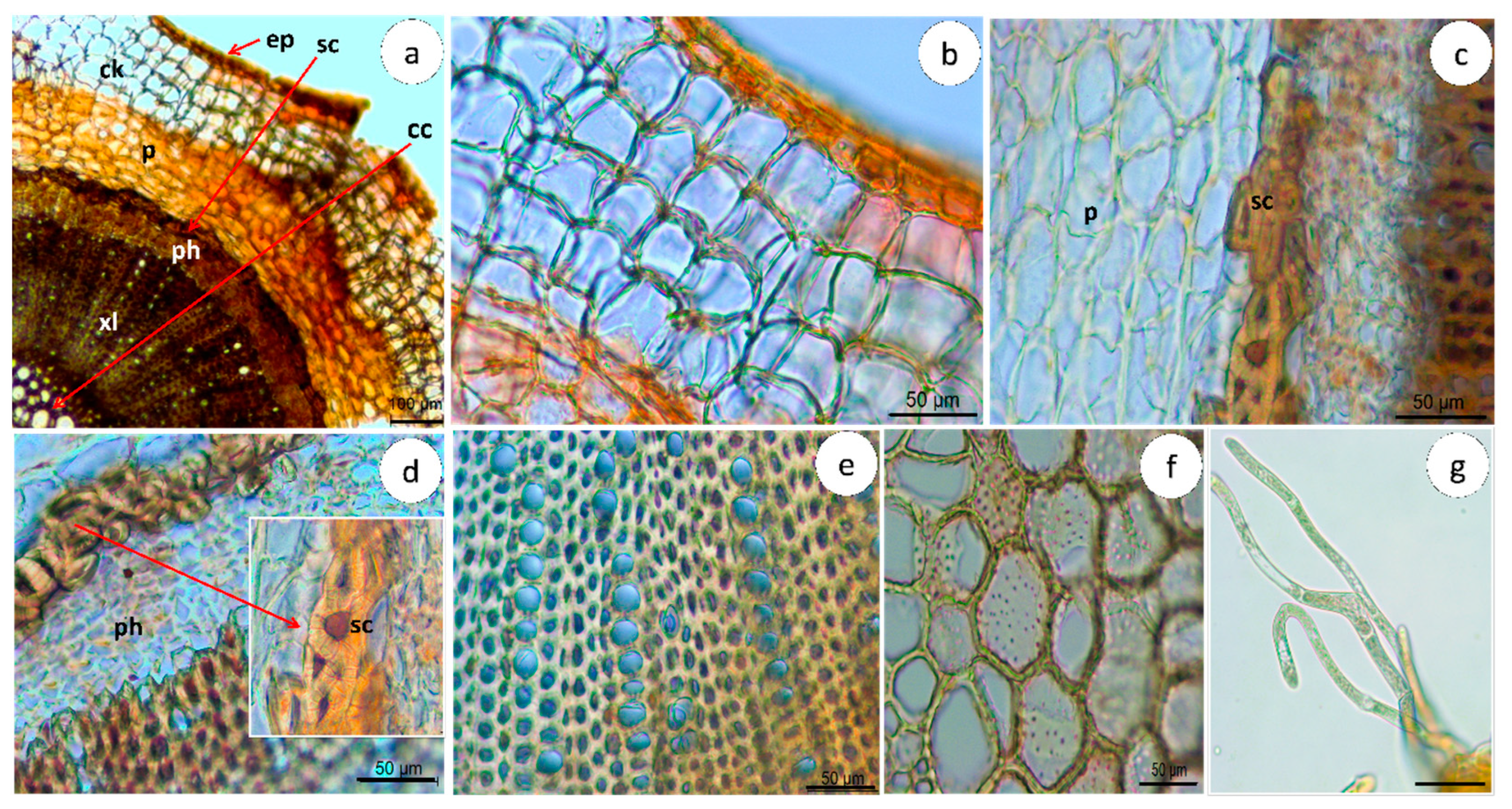
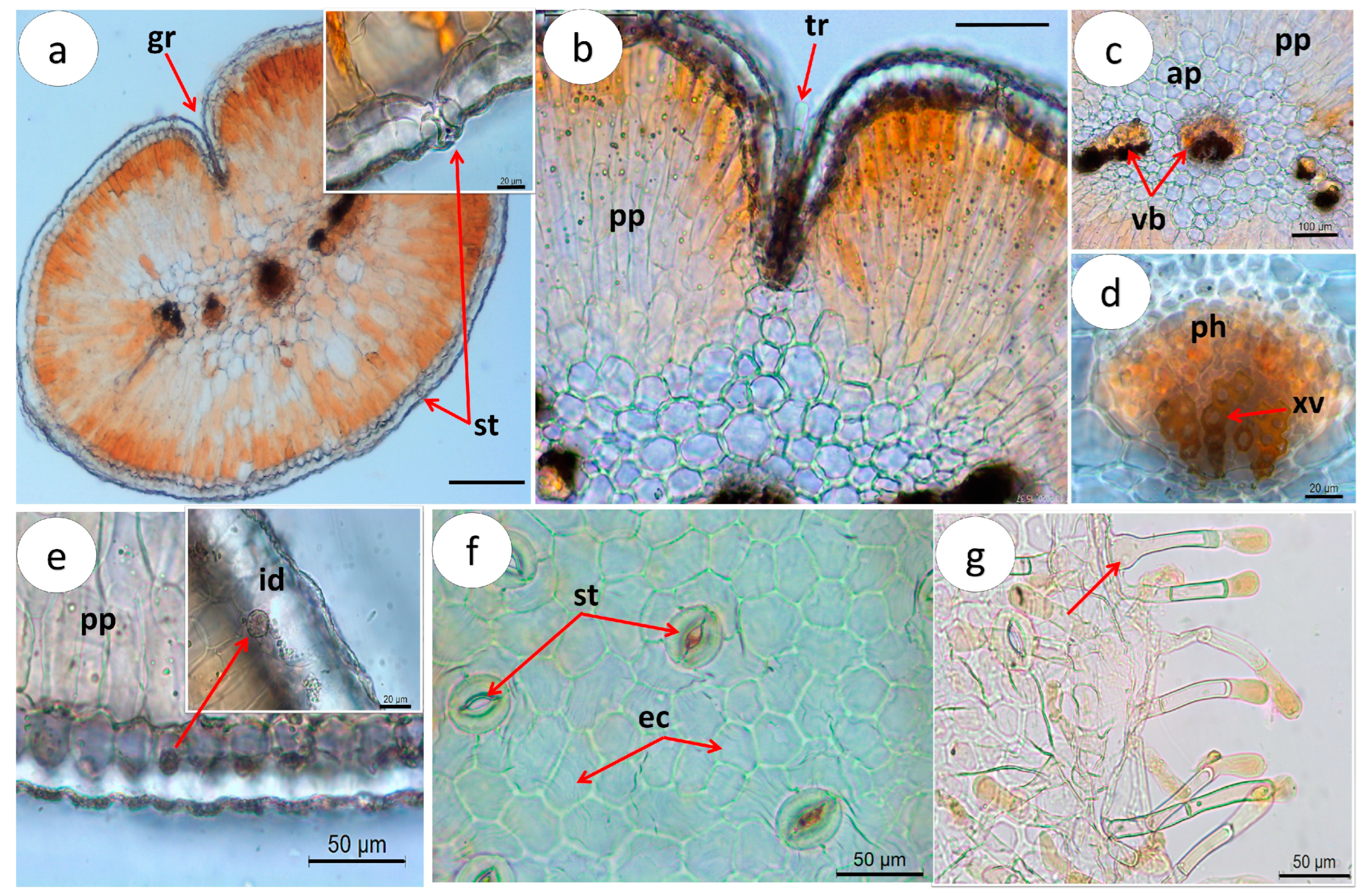
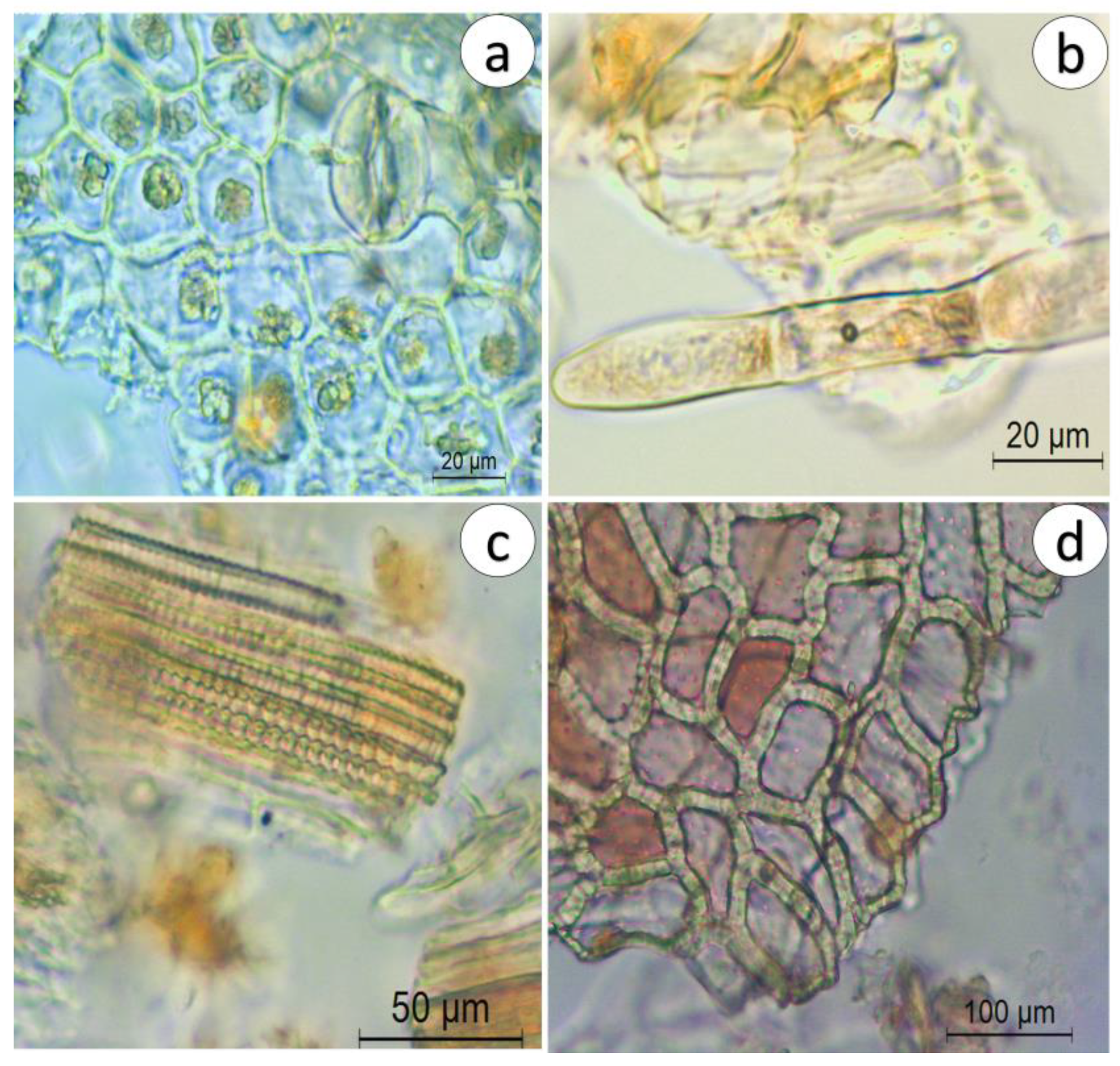
2.1.2. Microscopic Characterization
Stem
Leaf
Powdered Herbal Drug
2.2. Chemical Characterization
2.2.1. Drug–Extract Ratio
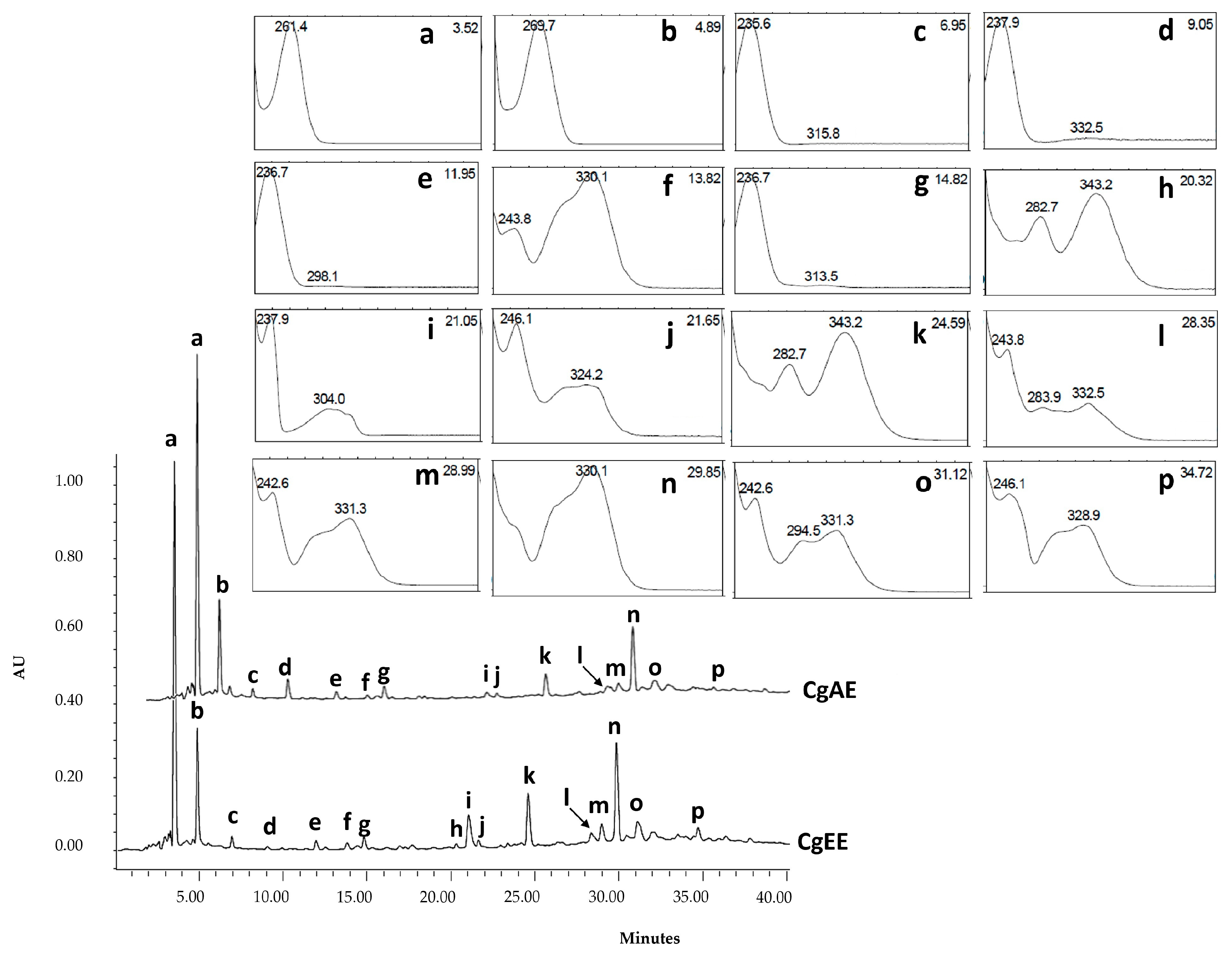
2.2.2. LC-UV/DAD Fingerprinting
2.2.3. LC-UV-ESI/MS-MS Chemical Profile
2.2.4. Quantification of Secondary Metabolites
2.3. In Vitro Biological Studies
2.3.1. In Vitro Antioxidant Activity
2.3.2. In Vitro Enzymatic Inhibition of Diabetes Type 2-Related Enzymes
2.4. Safety Assessment
2.4.1. In Vitro Genotoxic/Mutagenic Potential
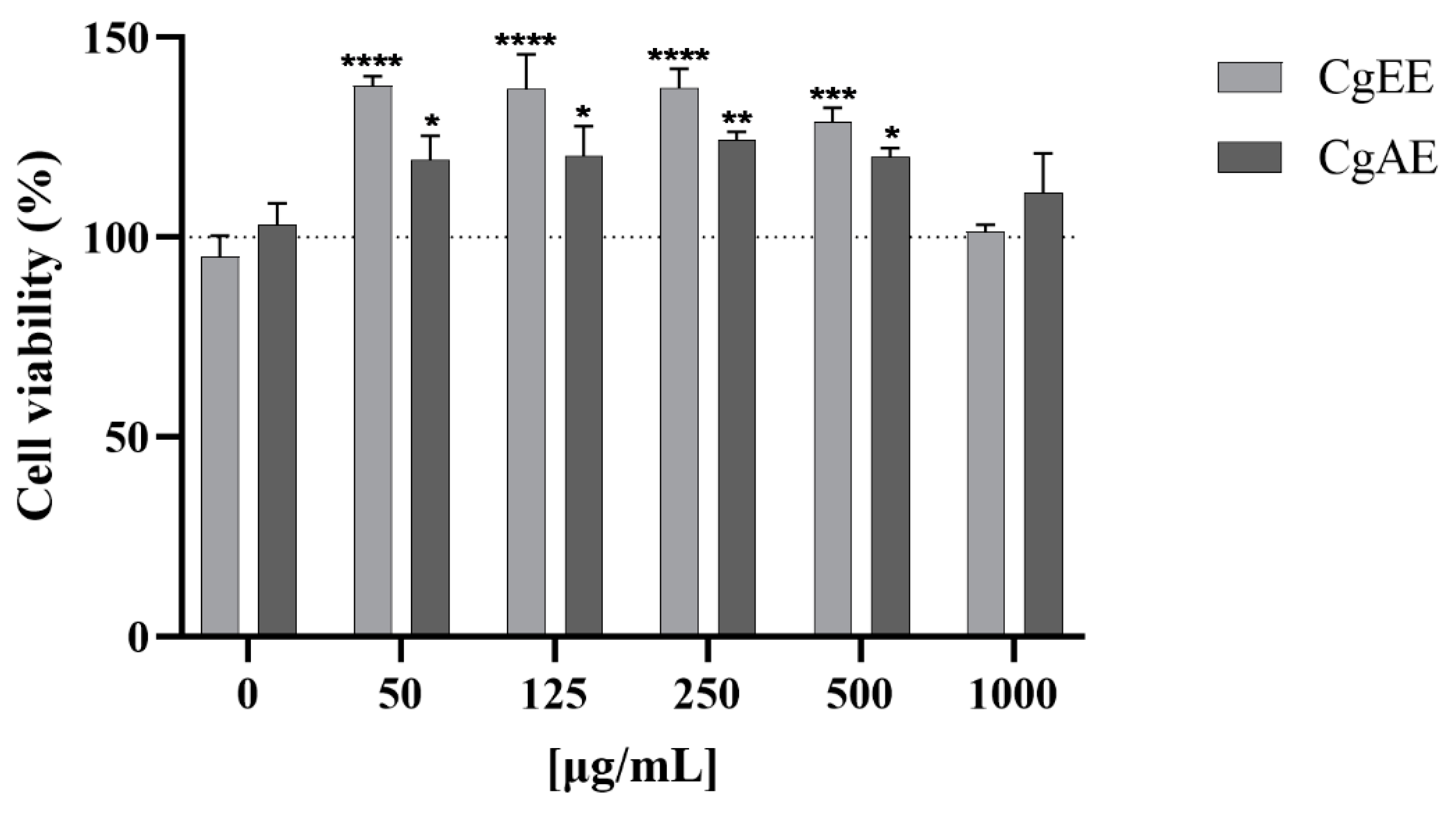
2.4.2. In Vitro Assessment of Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reference Items, and Reagents
4.2. Plant Material Collection and Processing
4.3. Preparation of Extracts
4.4. Botanical Characterization
4.4.1. Macroscopic Evaluation
4.4.2. Microscopic Evaluation
4.5. Chromatographic Conditions
4.6. Quantification of Main Classes of Secondary Metabolites
4.6.1. Total Phenolic Content
4.6.2. Flavonoid Content
4.6.3. Iridoid Content
4.7. Antioxidant Capacity
4.7.1. CUPRAC Assay
4.7.2. FRAP Assay
4.7.3. DPPH Radical Scavenging Assay
4.8. Enzymatic Inhibition
4.8.1. α-Amylase Inhibition Assay
4.8.2. α-Glucosidase Inhibition Assay
4.9. Safety Assessment
4.9.1. In Vitro Assessment of Genotoxicity/Mutagenicity
4.9.2. In Vitro Assessment of Cytotoxicity
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO Plantaginaceae Juss. |Plants of the World Online|Kew Science. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:30001136-2 (accessed on 21 August 2023).
- Rønsted, N.; Jensen, S.R. Iridoid Glucosides and Caffeoyl Phenylethanoid Glycosides from Campylanthus salsaloides and Campylanthus glaber. Biochem. Syst. Ecol. 2002, 30, 1091–1095. [Google Scholar] [CrossRef]
- Thiv, M.; Thulin, M.; Hjertson, M.; Kropf, M.; Linder, H.P. Evidence for a Vicariant Origin of Macaronesian-Eritreo/Arabian Disjunctions in Campylanthus Roth (Plantaginaceae). Mol. Phylogenet. Evol. 2010, 54, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Hjertson, M. Revision of the Disjunct Genus Campylanthus (Scrophulariaceae). Edinb. J. Bot. 2003, 60, 131–174. [Google Scholar] [CrossRef]
- Brochmann, C.; Rustan, Ø.H.; Lobin, W.; Kilian, N. The Endemic Vascular Plants of the Cape Verde Islands, W Africa. Sommerfeltia 1997, 24, 1–363. [Google Scholar] [CrossRef]
- Gomes, A.; Vasconcelos, T.; Almeida, M.H. Plantas Na Medicina Tradicional de Cabo Verde. In Proceedings of the Workshop Plantas Medicinais e Fitoterapêuticas nos Trópicos, Instituto de Investigação Científica Tropical/Centro Científico e Cultural de Macau, Lisboa, Portugal, 29–31 October 2008; pp. 1–13. [Google Scholar]
- Gomes, S. Plantas Endémicas Medicinais de Cabo Verde. Available online: http://portaldoconhecimento.gov.cv/handle/10961/3838 (accessed on 11 January 2020).
- Roque, S.; Duarte, I.; Graziani, P.; Nascimento, M.; Ramos, N. Usos Medicinais e Tradicionais Da Flora Endémica, Indígena e Exótica Do Monte Gordo; Projecto Áreas Protegidas CVI/02/G31/A/1G/99/2006; Ministério do Ambiente e Agricultura: São Nicolau, Cabo Verde, 2008. [Google Scholar]
- WHO. WHO Global Report on Traditional and Complementary Medicine 2019; WHO: Geneva, Switzerland, 2019.
- Essoh, A.; Liberal, Â.; Fernandes, Â.; Dias, M.I.; Pereira, C.; Mandim, F.; Moldão-Martins, M.; Cravo, P.; Duarte, M.P.; Moura, M.; et al. Evaluation of the Polyphenolic Composition and Bioactivities of Three Native Cabo Verde Medicinal Plants. Pharmaceuticals 2022, 15, 1162. [Google Scholar] [CrossRef] [PubMed]
- Romeiras, M.M.; Essoh, A.P.; Catarino, S.; Silva, J.; Lima, K.; Varela, E.; Moura, M.; Gomes, I.; Duarte, M.P.C.; Duarte, M.P.C. Diversity and Biological Activities of Medicinal Plants of Santiago Island (Cabo Verde). Heliyon 2023, 9, e14651. [Google Scholar] [CrossRef] [PubMed]
- De Moura Sperotto, N.D.; Steffens, L.; Veríssimo, R.M.; Henn, J.G.; Péres, V.F.; Vianna, P.; Chies, J.A.B.; Roehe, A.; Saffi, J.; Moura, D.J. Wound Healing and Anti-Inflammatory Activities Induced by a Plantago australis Hydroethanolic Extract Standardized in Verbascoside. J. Ethnopharmacol. 2018, 225, 178–188. [Google Scholar] [CrossRef]
- Sripahco, T.; Khruengsai, S.; Charoensup, R.; Tovaranonte, J.; Pripdeevech, P. Chemical Composition, Antioxidant, and Antimicrobial Activity of Elsholtzia beddomei C. B. Clarke Ex Hook. f. Essential Oil. Sci. Rep. 2022, 12, 2225. [Google Scholar] [CrossRef]
- Fritzen, M.; Dutra, R.; Crivelli, S.; Farmacognosia, I.; Cabral, S.; Farias, P.; Paes, R.; Linhares, G. (Eds.) 1a Edição; SESES: Rio de Janeiro, Brazil, 2016; ISBN 2013206534. [Google Scholar]
- Rabizadeh, F.; Mirian, M.S.; Doosti, R.; Kiani-Anbouhi, R.; Eftekhari, E. Phytochemical Classification of Medicinal Plants Used in the Treatment of Kidney Disease Based on Traditional Persian Medicine. Evid. Based Complement. Alternat. Med. 2022, 2022, 8022599. [Google Scholar] [CrossRef]
- Jain, C.; Khatana, S.; Vijayvergia, R. Bioactivity of Secondary Metabolites of Various Plants: A Review. Int. J. Pharm. Sci. Res. 2019, 10, 494–504. [Google Scholar] [CrossRef]
- Catarino, S.; Duarte, M.C.; Romeiras, M.M. Campylanthus glaber . IUCN Red List Threat. Species 2017, 2017, e.T107423655A107468087. [Google Scholar] [CrossRef]
- Romeiras, M.M.; Catarino, S.; Gomes, I.; Fernandes, C.; Costa, J.C.; Caujapé-Castells, J.; Duarte, M.C. IUCN Red List Assessment of the Cape Verde Endemic Flora: Towards a Global Strategy for Plant Conservation in Macaronesia. Bot. J. Linn. Soc. 2016, 180, 413–425. [Google Scholar] [CrossRef]
- Moyo, M.; Aremu, A.O.; Van Staden, J. Medicinal Plants: An Invaluable, Dwindling Resource in Sub-Saharan Africa. J. Ethnopharmacol. 2015, 174, 595–606. [Google Scholar] [CrossRef]
- Tahir, N.I.; Shaari, K.; Abas, F.; Ahmad Parveez, G.K.; Hashim, A.T.; Ramli, U.S. Identification of Oil Palm (Elaeis guineensis) Spear Leaf Metabolites Using Mass Spectrometry and Neutral Loss Analysis. J. Oil Palm Res. 2013, 25, 72–83. [Google Scholar]
- Fernández-Fernández, R.; López-Martínez, J.C.; Romero-González, R.; Martínez-Vidal, J.L.; Alarcón Flores, M.I.; Garrido Frenich, A. Simple LC-MS Determination of Citric and Malic Acids in Fruits and Vegetables. Chromatographia 2010, 72, 55–62. [Google Scholar] [CrossRef]
- Fu, Z.; Xue, R.; Li, Z.; Chen, M.; Sun, Z.; Hu, Y.; Huang, C. Fragmentation Patterns Study of Iridoid Glycosides in Fructus Gardeniae by HPLC-Q/TOF-MS/MS. Biomed. Chromatogr. 2014, 28, 1795–1807. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Liu, Z.; Song, F.; Xing, J.; Zheng, Z.; Liu, S. A Strategy for Identification and Structural Characterization of Compounds from Plantago Asiatica L. by Liquid Chromatography-Mass Spectrometry Combined with Ion Mobility Spectrometry. Molecules 2022, 27, 4302. [Google Scholar] [CrossRef]
- Kucharska, A.Z.; Fecka, I. Identification of Iridoids in Edible Honeysuckle Berries (Lonicera caerulea L. Var. kamtschatica Sevast.) by UPLC-ESI-QTOF-MS/MS. Molecules 2016, 21, 1157. [Google Scholar] [CrossRef]
- Yang, L.; He, J. Lagopsis supina Extract and Its Fractions Exert Prophylactic Effects against Blood Stasis in Rats via Anti-Coagulation, Anti-Platelet Activation and Anti-Fibrinolysis and Chemical Characterization by UHPLC-QTOF-MS/MS. Biomed. Pharmacother. 2020, 132, 110899. [Google Scholar] [CrossRef]
- Liao, X.; Hong, Y.; Chen, Z. Identification and Quantification of the Bioactive Components in Osmanthus fragrans Roots by HPLC-MS/MS. J. Pharm. Anal. 2021, 11, 299–307. [Google Scholar] [CrossRef]
- Dimaki, V.D.; Zeliou, K.; Nakka, F.; Stavreli, M.; Bakratsas, I.; Papaioannou, L.; Iatrou, G.; Lamari, F.N. Characterization of Sideritis clandestina Subsp. peloponnesiaca Polar Glycosides and Phytochemical Comparison to Other Mountain Tea Populations. Molecules 2022, 27, 7613. [Google Scholar] [CrossRef] [PubMed]
- Kelber, O.; Wegener, T.; Steinhoff, B.; Staiger, C.; Wiesner, J.; Knöss, W.; Kraft, K. Assessment of Genotoxicity of Herbal Medicinal Products: Application of the “Bracketing and Matrixing” Concept Using the Example of Valerianae radix (Valerian Root). Phytomedicine 2014, 21, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Dantas, F.G.; de Castilho, P.F.; de Almeida-Apolonio, A.A.; de Araújo, R.P.; de Oliveira, K.M.P. Mutagenic Potential of Medicinal Plants Evaluated by the Ames Salmonella/Microsome Assay: A Systematic Review. Mutat. Res. Mutat. Res. 2020, 786, 108338. [Google Scholar] [CrossRef] [PubMed]
- Maron, D.M.; Ames, B.N. Revised Methods for the Salmonella Mutagenicity Test. Mutat. Res. Mutagen. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef] [PubMed]
- Malmir, M.; Lima, K.; Póvoas Camões, S.; Manageiro, V.; Duarte, M.P.; Paiva Miranda, J.; Serrano, R.; da Silva, I.M.; Silva Lima, B.; Caniça, M.; et al. Identification of Marker Compounds and In Vitro Toxicity Evaluation of Two Portuguese Asphodelus Leaf Extracts. Molecules 2023, 28, 2372. [Google Scholar] [CrossRef]
- Malú, Q.; Lima, K.; Malmir, M.; Pinto, R.; da Silva, I.M.; Catarino, L.; Duarte, M.P.; Serrano, R.; Rocha, J.; Lima, B.S.; et al. Contribution to the Preclinical Safety Assessment of Lannea velutina and Sorindeia juglandifolia Leaves. Plants 2023, 12, 130. [Google Scholar] [CrossRef]
- Kilian, A.N.; Hein, P.; Bahah, S.O.; Kilian, N.; Hein, P.; Bahah, S.O.; Kilian, A.N.; Hein, P.; Bahah, S.O.; Kilian, N.; et al. A New Species of Campylanthus (Scrophulariaceae) from Ras Fartak, Al-Mahra, and Notes on Other Species of the Genus in Yemen. Willdenowia 2002, 32, 271–279. [Google Scholar] [CrossRef]
- Kummerow, J. Comparative Anatomy of Sclerophylls of Mediterranean Climatic Areas; Springer: Berlin/Heidelberg, Germany, 1973; pp. 157–167. [Google Scholar]
- Simioni, P.F.; Eisenlohr, P.V.; Pessoa, M.J.G.; da Silva, I.V. Elucidating Adaptive Strategies from Leaf Anatomy: Do Amazonian Savannas Present Xeromorphic Characteristics? Flora 2017, 226, 38–46. [Google Scholar] [CrossRef]
- Abd Elhalim, M.E.; Abo-Alatta, O.K.; Habib, S.A.; Abd Elbar, O.H. The Anatomical Features of the Desert Halophytes Zygophyllum album L.F. and Nitraria retusa (Forssk.) Asch. Ann. Agric. Sci. 2016, 61, 97–104. [Google Scholar] [CrossRef]
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Leaves. In Plant Anatomy; Springer: Cham, Switzerland, 2018; pp. 395–441. [Google Scholar] [CrossRef]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of Extraction Solvent on Total Phenol Content, Total Flavonoid Content, and Antioxidant Activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef]
- Kooiman, P. The Occurrence of Iridoid Glycosides in the Scrophulariaceae. Acta Bot. Neerl. 1975, 24, 459–468. [Google Scholar] [CrossRef]
- Albach, D.C.; Meudt, H.M.; Oxelman, B. Piecing Together the “New” Plantaginaceae. Am. J. Bot. 2005, 92, 297–315. [Google Scholar] [CrossRef]
- Gonda, S.; Nguyen, N.M.; Batta, G.; Gyémánt, G.; Máthé, C.; Vasas, G. Determination of Phenylethanoid Glycosides and Iridoid Glycosides from Therapeutically Used Plantago Species by CE-MEKC. Electrophoresis 2013, 34, 2577–2584. [Google Scholar] [CrossRef]
- Gonçalves, S.; Romano, A. The Medicinal Potential of Plants from the Genus Plantago (Plantaginaceae). Ind. Crops Prod. 2016, 83, 213–226. [Google Scholar] [CrossRef]
- Apak, R.; Capanoglu, E.; Shahidi, F. Measurment of Antioxidant Activity and Capacity: Recent Trends and Applications; Apak, R., Capanoglu, E., Shahidi, F., Eds.; Wiley: Oxford, UK, 2018; ISBN 9781119135357. [Google Scholar]
- Atoui, A.; Mansouri, A.; Boskou, G.; Kefalas, P. Tea and Herbal Infusions: Their Antioxidant Activity and Phenolic Profile. Food Chem. 2005, 89, 27–36. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Zhang, D.; Nie, S.; Xie, M.; Hu, J. Antioxidant and Antibacterial Capabilities of Phenolic Compounds and Organic Acids from Camellia oleifera Cake. Food Sci. Biotechnol. 2020, 29, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Deb, P.K.; Khound, P.; Bhattacharjee, S.; Choudhury, P.; Sarma, H.; Devi, R.; Sarkar, B. Variation in Chemical Constituents, In-Vitro Bioactivity and Toxicity Profile among Different Parts of Clerodendrum glandulosum Lindl. (C. colebrookianum Walp.). S. Afr. J. Bot. 2021, 140, 50–61. [Google Scholar] [CrossRef]
- Quazi, A.; Patwekar, M.; Patwekar, F.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Islam, F. In Vitro Alpha-Amylase Enzyme Assay of Hydroalcoholic Polyherbal Extract: Proof of Concept for the Development of Polyherbal Teabag Formulation for the Treatment of Diabetes. Evid.-Based Complement. Altern. Med. 2022, 2022, 1577957. [Google Scholar] [CrossRef]
- Somtimuang, C.; Olatunji, O.J.; Ovatlarnporn, C. Evaluation of In Vitro α-Amylase and α-Glucosidase Inhibitory Potentials of 14 Medicinal Plants Constituted in Thai Folk Antidiabetic Formularies. Chem. Biodivers. 2018, 15, e1800025. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase Inhibitors Isolated from Medicinal Plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef]
- Galli, A.; Marciani, P.; Marku, A.; Ghislanzoni, S.; Bertuzzi, F.; Rossi, R.; Di Giancamillo, A.; Castagna, M.; Perego, C. Verbascoside Protects Pancreatic β-Cells against ER-Stress. Biomedicines 2020, 8, 582. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Xue, H.Y.; Jin, L.J.; Li, S.Y.; Xu, Y.P. Antioxidant and Pancreas-Protective Effect of Aucubin on Rats with Streptozotocin-Induced Diabetes. Eur. J. Pharmacol. 2008, 582, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhu, Z.; Zhang, J.; Ren, C.; Zhang, Q. Aucubin Alleviates Diabetic Nephropathy by Inhibiting NF-ΚB Activation and Inducing SIRT1/SIRT3-FOXO3a Signaling Pathway in High-Fat Diet/Streptozotocin-Induced Diabetic Mice. J. Funct. Foods 2020, 64, 103702. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, L.; Zheng, Z.; Pi, Z.; Liu, Z.; Song, F. Online Microdialysis-Ultra Performance Liquid Chromatography–Mass Spectrometry Method for Comparative Pharmacokinetic Investigation on Iridoids from Gardenia jasminoides Ellis in Rats with Different Progressions of Type 2 Diabetic Complications. J. Pharm. Biomed. Anal. 2017, 140, 146–154. [Google Scholar] [CrossRef]
- Thengyai, S.; Thiantongin, P.; Sontimuang, C.; Ovatlarnporn, C.; Puttarak, P. α-Glucosidase and α-Amylase Inhibitory Activities of Medicinal Plants in Thai Antidiabetic Recipes and Bioactive Compounds from Vitex glabrata R. Br. Stem Bark. J. Herb. Med. 2020, 19, 100302. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, Q.V.; Nguyen, A.D.; Wang, S.L. Screening and Evaluation of α-Glucosidase Inhibitors from Indigenous Medicinal Plants in Dak Lak Province, Vietnam. Res. Chem. Intermed. 2017, 43, 3599–3612. [Google Scholar] [CrossRef]
- Verschaeve, L. Genotoxicity and Antigenotoxicity Studies of Traditional Medicinal Plants: How Informative and Accurate Are the Results? Nat. Prod. Commun. 2015, 10, 1489–1493. [Google Scholar] [CrossRef]
- Aydin, A.; Aktay, G.; Yesilada, E. A Guidance Manual for the Toxicity Assessment of Traditional Herbal Medicines. Nat. Prod. Commun. 2016, 11, 1763–1773. [Google Scholar] [CrossRef]
- Bardoloi, A.; Soren, A.D. Genotoxicity Induced by Medicinal Plants. Bull. Natl. Res. Cent. 2022, 46, 119. [Google Scholar] [CrossRef]
- Zhou, J.; Ouedraogo, M.; Qu, F.; Duez, P. Potential Genotoxicity of Traditional Chinese Medicinal Plants and Phytochemicals: An Overview. Phyther. Res. 2013, 27, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, G.; Adam, M.L.; Silva, C.D.; Silva Soley, B.; De Mello-Sampayo, C.; Cabrini, D.A.; Correr, C.J.; Otuki, M.F. Quantitative Genotoxicity Assays for Analysis of Medicinal Plants: A Systematic Review. J. Ethnopharmacol. 2016, 178, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Mortelmans, K.; Zeiger, E. The Ames Salmonella/Microsome Mutagenicity Assay. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2000, 455, 29–60. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, M.; Cohen, I.; Campbell, M. Reduction of MTT by Aqueous Herbal Extracts in the Absence of Cells. J. Ethnopharmacol. 2004, 93, 381–384. [Google Scholar] [CrossRef]
- European Directorate for the Quality of Medicines & Healthcare. European Pharmacopoeia, 10th ed.; Newsroom: Strasbourg, France, 2019. [Google Scholar]
- Loebler, M.; Sánchez, C.; Maurício, E.M.; Diogo, E.; Santos, M.; Vasilenko, P.; Cruz, A.S.; Mendes, B.; Gonçalves, M.; Duarte, M.P. Potential Application of Propolis Extracts to Control the Growth of Stemphylium vesicarium in ‘Rocha’ pear. Appl. Sci. 2020, 10, 1990. [Google Scholar] [CrossRef]
- Barros, L.; Carvalho, A.M.; Morais, J.S.; Ferreira, I.C.F.R. Strawberry-Tree, Blackthorn and Rose Fruits: Detailed Characterisation in Nutrients and Phytochemicals with Antioxidant Properties. Food Chem. 2010, 120, 247–254. [Google Scholar] [CrossRef]
- Nizioł-Lukaszewska, Z.; Wasilewski, T.; Bujak, T.; Osika, P. Iridoids from Cornus mas L. and Their Potential as Innovative Ingredients in Cosmetics. Polish J. Chem. Technol. 2017, 19, 122–127. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: CUPRAC Method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Lima, K.; Silva, O.; Figueira, M.E.; Pires, C.; Cruz, D.; Gomes, S.; Maurício, E.M.; Duarte, M.P. Influence of the In Vitro Gastrointestinal Digestion on the Antioxidant Activity of Artemisia gorgonum Webb and Hyptis pectinata (L.) Poit. Infusions from Cape Verde. Food Res. Int. 2019, 115, 150–159. [Google Scholar] [CrossRef]
- Miceli, N.; Trovato, A.; Dugo, P.; Cacciola, F.; Donato, P.; Marino, A.; Bellinghieri, V.; La Barbera, T.M.; Güvenç, A.; Taviano, M.F. Comparative Analysis of Flavonoid Profile, Antioxidant and Antimicrobial Activity of the Berries of Juniperus communis L. Var. communis and Juniperus communis L. Var. Saxatilis Pall. from Turkey. J. Agric. Food Chem. 2009, 57, 6570–6577. [Google Scholar] [CrossRef]
- Ferron, L.; Colombo, R.; Mannucci, B.; Papetti, A. A New Italian Purple Corn Variety (Moradyn) Byproduct Extract: Antiglycative and Hypoglycemic In Vitro Activities and Preliminary Bioaccessibility Studies. Molecules 2020, 25, 1958. [Google Scholar] [CrossRef] [PubMed]
- Rouzbehan, S.; Moein, S.; Homaei, A.; Moein, M.R. Kinetics of α-Glucosidase Inhibition by Different Fractions of Three Species of Labiatae Extracts: A New Diabetes Treatment Model. Pharm. Biol. 2017, 55, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 471: Bacterial Reverse Mutation Test, OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, France, 1997. Available online: http://www.oecd-ilibrary.org/environment/test-no-471-bacterial-reverse-mutation-test_9789264071247-en (accessed on 14 September 2021).
- ICH International Conference on Harmonization (ICH). Consensus Guideline S2(R1) Guidance on Genotoxicity Testing and Data Interpretation for Pharmaceuticals Intended for Human Use. Step 4 Version of November 2011; ICH Guideline; ICH: Geneva, Switzerland, 2011.
- Santos, J.M.; Camões, S.P.; Filipe, E.; Cipriano, M.; Barcia, R.N.; Filipe, M.; Teixeira, M.; Simões, S.; Gaspar, M.; Mosqueira, D.; et al. Three-Dimensional Spheroid Cell Culture of Umbilical Cord Tissue-Derived Mesenchymal Stromal Cells Leads to Enhanced Paracrine Induction of Wound Healing. Stem Cell Res. Ther. 2015, 6, 90. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anatomical Parameters | Min–Max | Mean | ±SD |
|---|---|---|---|
| Stem | |||
| Cork (no. of cell layers) | 3–7 | 4 | 1 |
| Sclereid cell wall thickness (µm) | 3.3–10.2 | 6.8 | 2.1 |
| Xylem vessel area (µm2) | 82.1–324.1 | 208 | 74 |
| Central cylinder cell area (µm2) | 770.9–4340.5 | 2304 | 1024 |
| Leaf | |||
| Cuticule thickness (µm) | 13.7–25.6 | 19.2 | 3.3 |
| Pallissade parenchyma thickness (µm) | 154–323 | 254.7 | 34.2 |
| Epidermal cell length (µm) | 22.1–55.4 | 36.4 | 7.9 |
| Epidermal cell width (µm) | 13.6–38.5 | 24.8 | 5.8 |
| Stomata length (µm) | 31.3–46.6 | 38.4 | 3.8 |
| Stomata width (µm) | 25.9–38.6 | 32.4 | 3.2 |
| Stomata index | - | 4.3 | 1.2 |
| Idioblast area (µm2) | 73.1–96.8 | 88.7 | 6.5 |
| Thrichome stalk length (µm) | 49.4–120.7 | 77.1 | 16.6 |
| Peak | tr (min) | UVλmax (nm) | [M-H]− | MS-MS Fragment Ions m/z (Relative Abundance) | Proposed Compound | Class |
|---|---|---|---|---|---|---|
| a | 3.52 | 261 | 133 | 133 (88); 115 (100); 71(40); 73 (18) | Malic acid | Organic acid |
| b | 4.89 | 269 | 191 | 191(45); 111(100); 87(38); 85 (27) | Citric acid | Organic acid |
| c | 6.95 | 234 | 373 | 123 (100); 193(75); 149 (48) | Gardoside | Iridoid |
| d | 9.05 | 238 | 373 | 123 (100); 149 (80) | Geniposidic acid * | Iridoid |
| e | 11.95 | 235 | 375 | 169 (100); 89 (65); 151 (59); 213(50) | 8-Epiloganic acid | Iridoid |
| f | 13.82 | 244; 330 | 487 | 179 (100); 135(13); 161 (12); 341 (5) | Cistanoside derivative | Phenylethanoid glycoside |
| g | 14.82 | 236 | 451 | 243(100); 101(50) | Iridoid derivative | Iridoid |
| h | 20.32 | 283; 343 | 667 | 351 (100); 113 (12) | Unknown | Flavonoid |
| i | 21.05 | 238; 304 | 453 | 89 (100); 159 (90) | Unknown | Unknown |
| j | 21.65 | 246; 283sh; 324 | 639 | 639 (100); 621(33); 161(26); 179 (23); 451 (22) | β-Hydroxyverbascoside isomer | Phenylethanoid glycoside |
| k | 24.59 | 283; 343 | 667 | 351 (100); 315 (28); 113 (25) | Unknown | Flavonoid |
| l | 28.35 | 244; 284sh; 332 | 637 | 637 (100); 251 (70); 62(63); 623 (25) | Unknown | Phenylethanoid glycoside |
| m | 28.99 | 331 | 755 | 755 (100); 161 (22); 593 (12) | Lavandulifolioside | Phenylethanoid glycoside |
| n | 29.85 | 330 | 623 | 623 (100); 161 (28); 461 (12); 135 (3) | Verbascoside | Phenylethanoid glycoside |
| o | 31.12 | 331 | 623 | 623 (100); 161 (28); 461 (12); 135 (3) | cis-Verbascoside | Phenylethanoid glycoside |
| p | 34.72 | 328 | 623 | 623 (100); 161 (28); 461 (12); 135 (3) | Isoverbascoside | Phenylethanoid glycoside |
| Plant Extracts | DER | TPC (mg GAE/g) | TFC (mg CE/g) | TIC (mg AuE/g) |
|---|---|---|---|---|
| CgAE | 4.39:1 | 131.3 b ± 3.9 | 71.0 b ± 0.6 | 4.9 a ± 0.6 |
| CgEE | 2.89:1 | 148.1 a ± 9.5 | 75.8 a ± 0.2 | 6.1 a ± 1.3 |
| Samples | CUPRAC (µg AA/g) | FRAP (µg AA/g) | DPPH• IC50 (µg/mL) |
|---|---|---|---|
| CgAE | 197.9 b ± 2.6 | 109.8 a ± 3.2 | 130.9 a ± 1.4 |
| CgEE | 203.8 a ± 1.8 | 104.0 a ± 3.9 | 134.3 a ± 3.1 |
| Ascorbic acid | - | - | 17.3 ± 0.3 |
| Samples | α-Amylase IC50 (mg/mL) | α-Glucosidase IC50 (µg/mL) |
|---|---|---|
| CgAE | 7.21 c ± 0.23 | nd |
| CgEE | 5.77 b ± 0.10 | 827.9 b ± 11.2 |
| Acarbose | 0.011 a ± 0.001 | 350.3 a ± 15.4 |
| CgAE (µg/Plate) | Revertant Colonies per Plate (Mean ± SD) without Metabolic Activation | ||||
|---|---|---|---|---|---|
| TA98 | TA100 | TA102 | TA1535 | TA1537 | |
| 0 | 26 ± 4 | 148 ± 6 | 361 ± 9 | 20 ± 4 | 28 ±5 |
| 250 | 26 ± 4 | 167 ± 20 | 358 ± 24 | 14 ± 1 | 31 ± 9 |
| 625 | 25 ± 6 | 166 ± 22 | 388 ± 17 | 16 ± 5 | 26 ± 6 |
| 1250 | 26 ± 2 | 154 ± 15 | 365 ± 20 | 22 ± 5 | 27 ± 6 |
| 2500 | 25 ± 2 | 162 ± 13 | 377 ± 19 | 16 ± 2 | 21 ± 1 |
| 3750 | 28 ± 2 | 170 ± 61 | 384 ± 21 | 20 ± 6 | 30 ± 2 |
| 5000 | 26 ± 2 | 178 ± 5 | 406 ± 35 | 14 ± 3 | 27 ± 8 |
| CgEE (µg/plate) | Revertant colonies per plate (mean ± SD) without metabolic activation | ||||
| TA98 | TA100 | TA102 | TA1535 | TA1537 | |
| 0 | 26 ± 5 | 196 ± 5 | 344 ± 5 | 13 ± 3 | 25 ± 3 |
| 250 | 25 ± 5 | 179 ± 22 | 364 ± 5 | 14 ± 3 | 25 ± 3 |
| 625 | 25 ± 7 | 192 ± 22 | 361 ± 7 | 14 ± 1 | 28 ± 3 |
| 1250 | 26 ± 6 | 199 ± 6 | 347 ± 6 | 12 ± 1 | 26 ± 3 |
| 2500 | 29 ± 8 | 204 ± 8 | 391 ± 8 | 14 ± 3 | 21 ± 5 |
| 3750 | 28 ± 5 | 213 ± 22 | 388 ± 5 | 20 ± 7 | 25 ± 8 |
| 5000 | 30 ± 3 | 221 ± 6 | 388 ± 3 | 11 ± 1 | 24 ± 4 |
| PC | (1) | (2) | (3) | (2) | (4) |
| 488 ± 30 * | 1048 ± 43 * | 881 ± 26 * | 827 ± 13 * | 1354 ± 5 * | |
| CgEE (µg/Plate) | Revertant Colonies per Plate (Mean ± SD) with Metabolic Activation | ||||
|---|---|---|---|---|---|
| TA98 | TA100 | TA102 | TA1535 | TA1537 | |
| 0 | 47 ± 4 | 157 ± 6 | 172 ± 2 | 11 ± 2 | 12 ± 1 |
| 625 | 61 ± 7 | 146 ± 7 | 180 ± 35 | 12 ± 2 | 16 ± 3 |
| 1250 | 65 ± 4 | 167 ± 18 | 200 ± 11 | 16 ± 1 | 14 ± 3 |
| 2500 | 50 ± 6 | 175 ± 16 | 205 ± 3 | 16 ± 1 | 15 ± 2 |
| 5000 | 37 ± 3 | 162 ± 3 | 180 ± 35 | 12 ± 1 | 15 ± 4 |
| PC | (1) | (2) | (1) | (1) | (1) |
| 832 ± 35 * | 947 ± 148 * | 732 ± 12 * | 266 ± 1 * | 306 ± 50 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, K.; Malmir, M.; Camões, S.P.; Hasan, K.; Gomes, S.; Moreira da Silva, I.; Figueira, M.E.; Miranda, J.P.; Serrano, R.; Duarte, M.P.; et al. Quality, Safety and Biological Studies on Campylanthus glaber Aerial Parts. Pharmaceuticals 2023, 16, 1373. https://doi.org/10.3390/ph16101373
Lima K, Malmir M, Camões SP, Hasan K, Gomes S, Moreira da Silva I, Figueira ME, Miranda JP, Serrano R, Duarte MP, et al. Quality, Safety and Biological Studies on Campylanthus glaber Aerial Parts. Pharmaceuticals. 2023; 16(10):1373. https://doi.org/10.3390/ph16101373
Chicago/Turabian StyleLima, Katelene, Maryam Malmir, Sérgio P. Camões, Kamrul Hasan, Samuel Gomes, Isabel Moreira da Silva, Maria Eduardo Figueira, Joana P. Miranda, Rita Serrano, Maria Paula Duarte, and et al. 2023. "Quality, Safety and Biological Studies on Campylanthus glaber Aerial Parts" Pharmaceuticals 16, no. 10: 1373. https://doi.org/10.3390/ph16101373