
Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review

 ,
,  ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
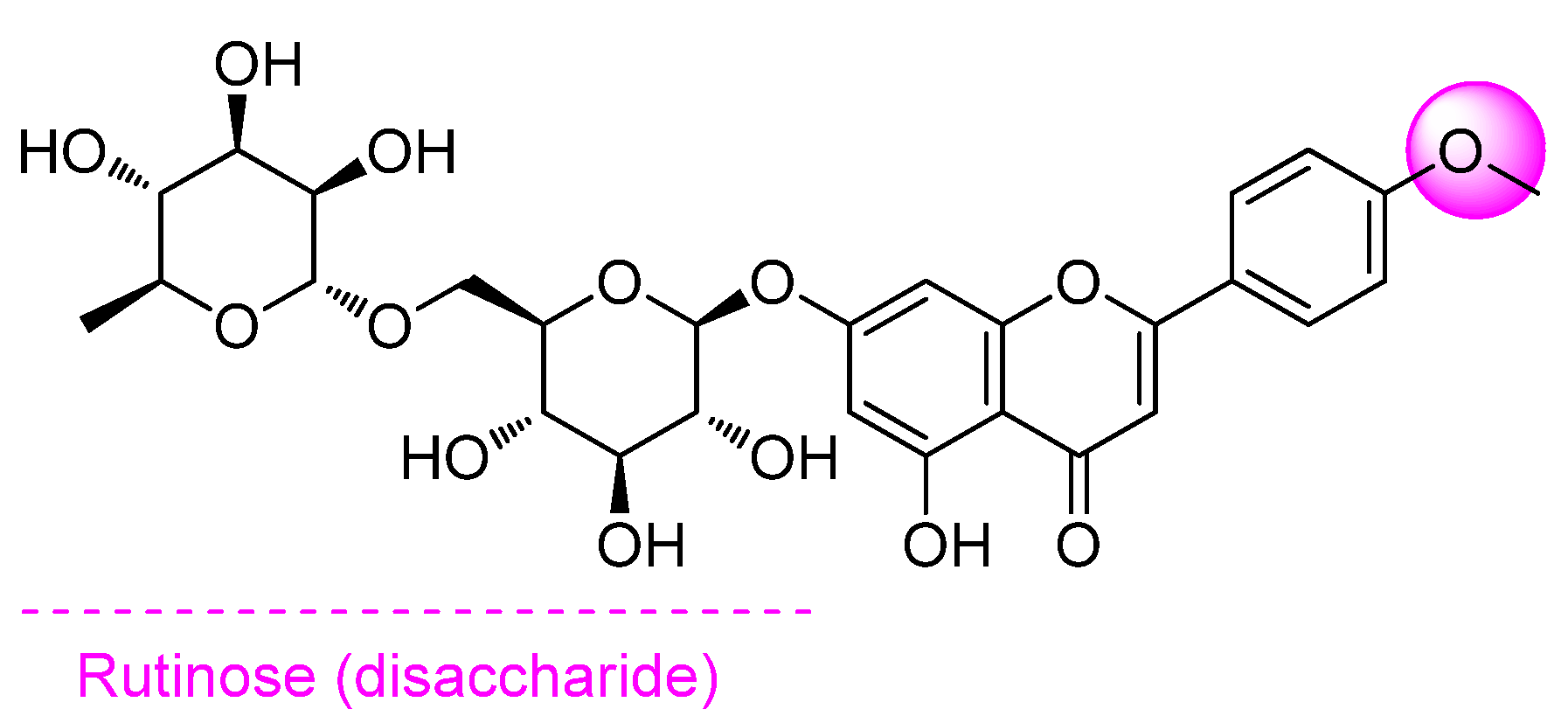{kind=link}
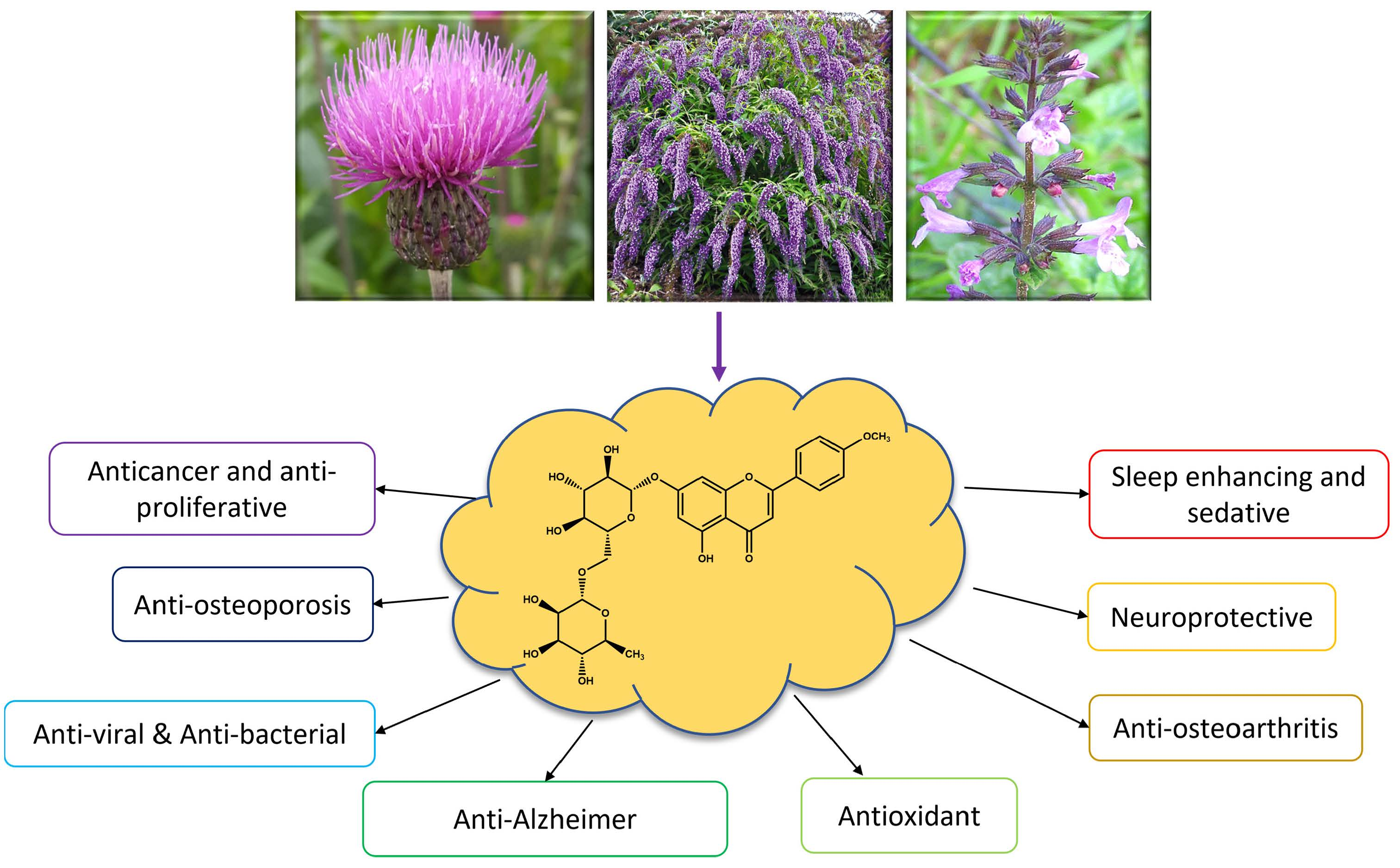
1. Introduction
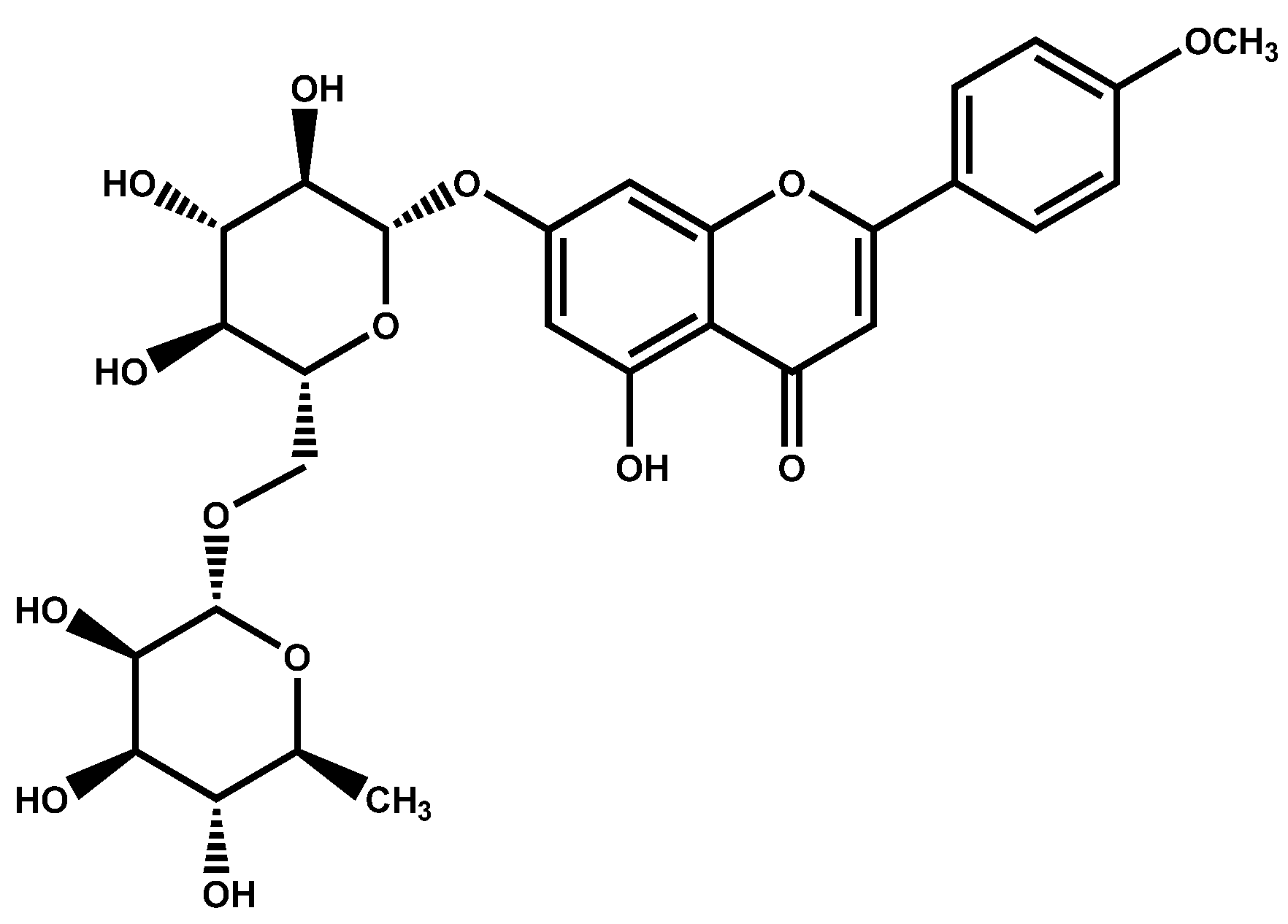
2. Phytochemistry and Chemotaxonomy of Linarin
2.1. Isolation of Linarin from Plant Species
2.1.1. Asteraceae
2.1.2. Lamiaceae
2.1.3. Scrophulariaceae
2.1.4. Miscellaneous Plants
2.2. Quantification and Qualification Analysis of Linarin in Plants
2.2.1. Asteraceae
2.2.2. Lamiaceae
2.2.3. Scrophulariaceae
2.2.4. Valerianaceae
2.2.5. Miscellaneous Plants
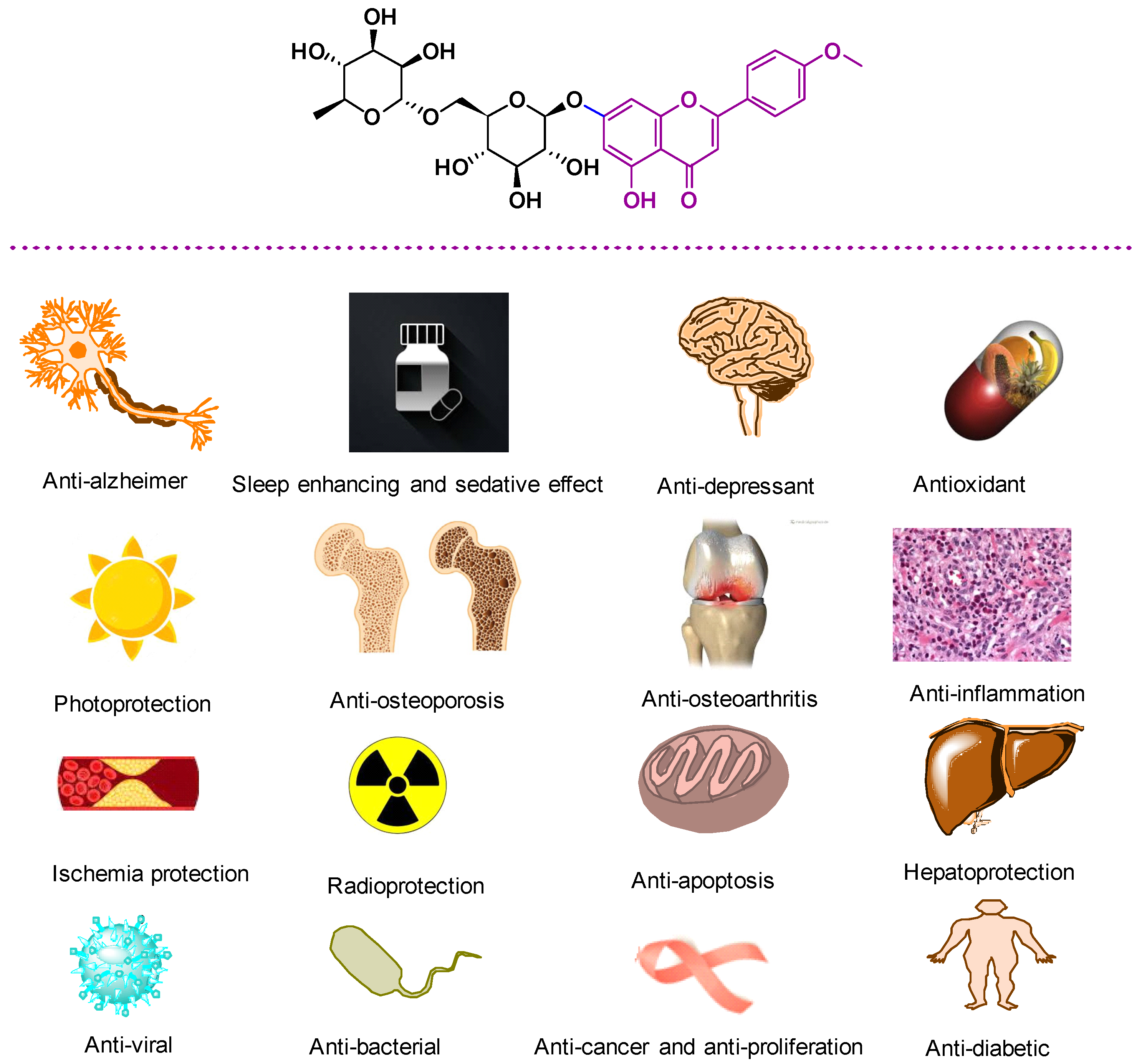
3. Biological Properties of LN
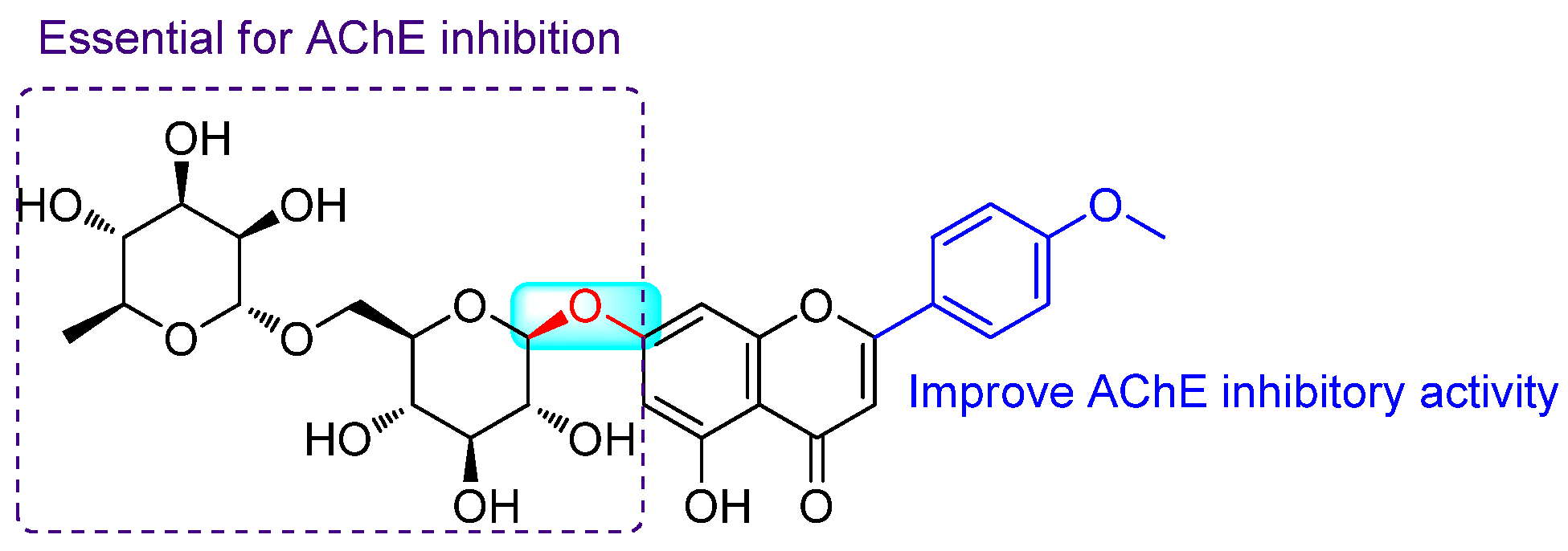
3.1. Anti-Alzheimer Properties
3.2. Antioxidant Properties
3.3. Sleep Enhancing and Sedative Effect
3.4. Anti-Osteoporosis Activity
3.5. Osteoarthritis Treatment
3.6. Ischemia Protection
3.7. Anti-Inflammation Activity
3.8. Photoprotective Properties
3.9. Radioprotection
3.10. Anti-Apoptosis Potential
3.11. Hepatoprotective Function
3.12. Non-Alcoholic Steatohepatitis Effect
3.13. Anti-Diabetic Effects
3.14. Analgesic and Anti-Pyretic Properties
3.15. Spasmolytic Properties
3.16. Treatment of Chronic Venous Hypertension
3.17. Anti-Bacterial Activity
3.18. Anti-Viral Activity
3.19. Anti-Cancer and Anti-Proliferative Activity
3.20. Negative Biological Results of LN
3.20.1. Estrogenic Activity
3.20.2. Anti-Fungal Effect
3.20.3. Anti-Depressant Properties
4. Perspectives
Anti-SARS-CoV-2 (COVID-19) Effect
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mottaghipisheh, J.; Kiss, T.; Tóth, B.; Csupor, D. The Prangos genus: A comprehensive review on traditional use, phytochemistry, and pharmacological activities. Phytochem. Rev. 2020, 19, 1449–1470. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Maghsoudlou, M.T.; Valizadeh, J.; Arjomandi, R. Antioxidant activity and chemical composition of the essential oil of Ducrosia anethifolia (DC.) Boiss. from Neyriz. J. Med. Plants By-Prod. 2014, 2, 215–218. [Google Scholar]
- Dehsheikh, A.B.; Sourestani, M.M.; Dehsheikh, P.B.; Mottaghipisheh, J.; Vitalini, S.; Iriti, M. Monoterpenes: Essential oil components with valuable features. Mini-Rev. Med. Chem. 2020, 20, 958–974. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A. High-temperature stress and metabolism of secondary metabolites in plants. In Effect of High Temperature on Crop Productivity and Metabolism of Macro Molecules; Elsevier: London, UK, 2019; pp. 391–484. [Google Scholar]
- Kwok, K.K.; Vincent, E.C.; Gibson, J.N. Antineoplastic drugs. In Pharmacology and Therapeutics for Dentistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 530–562. [Google Scholar]
- Pullaiah, T.; Raveendran, V. Camptothecin: Chemistry, biosynthesis, analogs, and chemical synthesis. In Camptothecin and Camptothecin Producing Plants; Elsevier: Amsterdam, The Netherlands, 2020; pp. 47–101. [Google Scholar]
- Dockery, L.; Daniel, M.-C. Dendronized Systems for the Delivery of Chemotherapeutics. Adv. Cancer Res. 2018, 139, 85–120. [Google Scholar] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Hao, Z.; Li, M. Isolation and structure identification of flavonoids. In Flavonoids—From Biosynthesis to Human Health; InTech: Rijeka, Croatia, 2017; pp. 17–23. [Google Scholar]
- Yamagata, K. Metabolic syndrome: Preventive effects of dietary flavonoids. Stud. Nat. Prod. Chem. 2019, 60, 1–28. [Google Scholar]
- Mottaghipisheh, J.; Stuppner, H. A comprehensive review on chemotaxonomic and phytochemical aspects of homoisoflavonoids, as rare flavonoid derivatives. Int. J. Mol. Sci. 2021, 22, 2735. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Iriti, M. Sephadex® LH-20, isolation, and purification of flavonoids from plant species: A comprehensive review. Molecules 2020, 25, 4146. [Google Scholar] [CrossRef]
- Arai, M.A.; Tanaka, M.; Tanouchi, K.; Ishikawa, N.; Ahmed, F.; Sadhu, S.K.; Ishibashi, M. Hes1-binding compounds isolated by Target Protein Oriented Natural Products Isolation (TPO-NAPI). J. Nat. Prod. 2017, 80, 538–543. [Google Scholar] [CrossRef]
- Demirtas, I.; Tufekci, A.R.; Yaglioglu, A.S.; Elmastas, M. Studies on the antioxidant and antiproliferative potentials of Cirsium arvense subsp. vestitum. J. Food Biochem. 2017, 41, e12299. [Google Scholar] [CrossRef] [Green Version]
- Kozyra, M.; Biernasiuk, A.; Malm, A.; Chowaniec, M. Chemical compositions and antibacterial activity of extracts obtained from the inflorescences of Cirsium canum (L.) all. Nat. Prod. Res. 2015, 29, 2059–2063. [Google Scholar] [CrossRef] [PubMed]
- Ganzera, M.; Pöcher, A.; Stuppner, H. Differentiation of Cirsium japonicum and C. setosum by TLC and HPLC-MS. Phytochem. Anal. 2005, 16, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Jiang, J.-G.; Zhang, X.-M.; Zhu, W. Identification of luteolin 7-O-β-D-glucuronide from Cirsium japonicum and its anti-inflammatory mechanism. J. Funct. Foods 2018, 46, 521–528. [Google Scholar] [CrossRef]
- Zhang, Z.; Jia, P.; Zhang, X.; Zhang, Q.; Yang, H.; Shi, H.; Zhang, L. LC–MS/MS determination and pharmacokinetic study of seven flavonoids in rat plasma after oral administration of Cirsium japonicum DC. extract. J. Ethnopharmacol. 2014, 158, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Jung, Y.; Baek, J.Y.; Shin, M.S.; Lee, S.; Hahm, D.H.; Lee, S.C.; Shim, J.S.; Kim, S.N.; Kang, K.S. Cirsimaritin contributes to the estrogenic activity of Cirsium japonicum var. maackii through the activation of estrogen receptor α. Bull. Korean Chem. Soc. 2017, 38, 1486–1490. [Google Scholar] [CrossRef]
- Han, H.-S.; Shin, J.-S.; Lee, S.-B.; Park, J.C.; Lee, K.-T. Cirsimarin, a flavone glucoside from the aerial part of Cirsium japonicum var. ussuriense (Regel) Kitam. ex Ohwi, suppresses the JAK/STAT and IRF-3 signaling pathway in LPS-stimulated RAW 264.7 macrophages. Chem. Biol. Interact. 2018, 293, 38–47. [Google Scholar] [CrossRef]
- Walesiuk, A.; Nazaruk, J.; Braszko, J.J. Pro-cognitive effects of Cirsium rivulare extracts in rats. J. Ethnopharmacol. 2010, 129, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.H.; Park, E.K.; Kim, T.H. New anti-glycative flavonoids from Cirsium setidens with potent radical scavenging activities. Phytochem. Lett. 2018, 26, 115–119. [Google Scholar] [CrossRef]
- Lu, Y.; Song, W.; Liang, X.; Wei, D.; Zhou, X. Chemical fingerprint and quantitative analysis of Cirsium setosum by LC. Chromatographia 2009, 70, 125–131. [Google Scholar] [CrossRef]
- Sun, Q.; Chang, L.; Ren, Y.; Cao, L.; Sun, Y.; Du, Y.; Shi, X.; Wang, Q.; Zhang, L. Simultaneous analysis of 11 main active components in Cirsium setosum based on HPLC-ESI-MS/MS and combined with statistical methods. J. Sep. Sci. 2012, 35, 2897–2907. [Google Scholar] [CrossRef]
- Wang, B.; Lv, D.; Huang, P.; Yan, F.; Liu, C.; Liu, H. Optimization, evaluation and identification of flavonoids in Cirsium setosum (Willd.) MB by using response surface methodology. J. Food Meas. Charact. 2019, 13, 1175–1184. [Google Scholar] [CrossRef]
- Nugroho, A.; Lim, S.C.; Byeon, J.S.; Choi, J.S.; Park, H.J. Simultaneous quantification and validation of caffeoylquinic acids and flavonoids in Hemistepta lyrata and peroxynitrite-scavenging activity. J. Pharm. Biomed. Anal. 2013, 76, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.H.; Kang, S.S.; Seo, E.A.; Shin, S.W. Isolation of aldose reductase inhibitors from the flowers of Chrysanthemum boreale. Arch. Pharm. Res. 1995, 18, 65–68. [Google Scholar] [CrossRef]
- Han, A.R.; Kim, H.Y.; So, Y.; Nam, B.; Lee, I.S.; Nam, J.W.; Jo, Y.D.; Kim, S.H.; Kim, J.B.; Kang, S.Y.; et al. Quantification of antioxidant phenolic compounds in a new Chrysanthemum cultivar by high-performance liquid chromatography with diode array detection and electrospray ionization mass spectrometry. Int. J. Anal. Chem. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Dasgupta, A. Antiinflammatory herbal supplements. In Translational Inflammation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 69–91. [Google Scholar]
- Wu, X.L.; Li, C.W.; Chen, H.M.; Su, Z.Q.; Zhao, X.N.; Chen, J.N.; Lai, X.P.; Zhang, X.J.; Su, Z.R. Anti-inflammatory effect of supercritical-carbon dioxide fluid extract from flowers and buds of Chrysanthemum indicum Linnén. Evid.-Based Complement. Altern. Med. 2013, 2013, 413237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Wu, X.; Kuang, Y.; Wang, T.; Bi, K.; Li, Q. Quality assessment of Chrysanthemum indicum Flower by simultaneous quantification of six major ingredients using a single reference standard combined with HPLC fingerprint analysis. Asian J. Pharm. Sci. 2016, 11, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Seo, D.W.; Cho, Y.R.; Kim, W.; Eom, S.H. Phytochemical linarin enriched in the flower of Chrysanthemum indicum inhibits proliferation of A549 human alveolar basal epithelial cells through suppression of the Akt-dependent signaling pathway. J. Med. Food 2013, 16, 1086–1094. [Google Scholar] [CrossRef]
- Kim, S.J.; Cho, H.I.; Kim, S.J.; Park, J.H.; Kim, J.S.; Kim, Y.H.; Lee, S.K.; Kwak, J.H.; Lee, S.M. Protective effect of linarin against D-galactosamine and lipopolysaccharide-induced fulminant hepatic failure. Eur. J. Pharmacol. 2014, 738, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ji, X.; Duan, L.; Ye, P.; Yang, J.; Zhan, R.; Chen, W.; Ma, D. Gene mining and identification of a flavone synthase II involved in flavones biosynthesis by transcriptomic analysis and targeted flavonoid profiling in Chrysanthemum indicum L. Ind. Crops Prod. 2019, 134, 244–256. [Google Scholar] [CrossRef]
- Hwang, S.; Paek, J.; Lim, S. Simultaneous ultra performance liquid chromatography determination and antioxidant activity of linarin, luteolin, chlorogenic acid and apigenin in different parts of compositae species. Molecules 2016, 21, 1609. [Google Scholar] [CrossRef] [PubMed]
- Qiaoshan, Y.; Suhong, C.; Minxia, S.; Wenjia, M.; Bo, L.; Guiyuan, L. Preparative purification of linarin extracts from Dendranthema indicum flowers and evaluation of its antihypertensive effect. Evid.-Based Complement. Altern. Med. 2014, 2014, 394276. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.J.; Lee, S.Y.; Kim, J.S.; Lee, S.; Choi, R.J.; Chung, H.S.; Kim, Y.S.; Kang, S.S. Sesquiterpenes and other constituents from Dendranthema zawadskii var. latilobum. Chem. Pharm. Bull. 2012, 60, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Agrawal, P.; Yim, D.; Agarwal, C.; Agarwal, R. Acacetin inhibits cell growth and cell cycle progression, and induces apoptosis in human prostate cancer cells: Structure-activity relationship with linarin and linarin acetate. Carcinogenesis 2005, 26, 845–854. [Google Scholar] [CrossRef]
- Li, J.; Lin, X.; Zhang, Y.; Liu, W.; Mi, X.; Zhang, J.; Su, J. Preparative purification of bioactive compounds from Flos Chrysanthemi indici and evaluation of its antiosteoporosis effect. Evid.-Based Complement. Altern. Med. 2016, 2016, 2587201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Zhang, Q.; Cui, K.; Chen, G.; Liu, X.; Wang, L. Optimization of extraction of linarin from Flos Chrysanthemi indici by response surface methodology and artificial neural network. J. Sep. Sci. 2017, 40, 2062–2070. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hao, L.-J.; Zhu, J.-J.; Wang, Z.-M.; Zhang, X.; Song, X. Comparative evaluation of Chrysanthemum Flos from different origins by HPLC-DAD-MS n and relative response factors. Food Anal. Methods 2015, 8, 40–51. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Wang, C.; Sun, W.; Zhang, Z.; Cheng, W. A gradient HPLC method for the quality control of chlorogenic acid, linarin and luteolin in Flos Chrysanthemi indici suppository. J. Pharm. Biomed. Anal. 2007, 43, 753–757. [Google Scholar] [CrossRef]
- Jung, H.A.; Park, J.J.; Islam, M.N.; Jin, S.E.; Min, B.S.; Lee, J.H.; Sohn, H.S.; Choi, J.S. Inhibitory activity of coumarins from Artemisia capillaris against advanced glycation endproduct formation. Arch. Pharm. Res. 2012, 35, 1021–1035. [Google Scholar] [CrossRef]
- Laskaris, G.G.; Gournelis, D.C.; Kokkalou, E. Phenolics of Picnomon acarna. J. Nat. Prod. 1995, 58, 1248–1250. [Google Scholar] [CrossRef]
- Kokkini, S.; Karousou, R.; Hanlidou, E. HERBS|Herbs of the Labiatae. In Encyclopedia of Food Sciences and Nutrition; Elsevier: Amsterdam, The Netherlands, 2003; pp. 3082–3090. [Google Scholar]
- Oinonen, P.P.; Jokela, J.K.; Hatakka, A.I.; Vuorela, P.M. Linarin, a selective acetylcholinesterase inhibitor from Mentha arvensis. Fitoterapia 2006, 77, 429–434. [Google Scholar] [CrossRef]
- Salin, O.; Törmäkangas, L.; Leinonen, M.; Saario, E.; Hagström, M.; Ketola, R.A.; Saikku, P.; Vuorela, H.; Vuorela, P.M. Corn mint (Mentha arvensis) extract diminishes acute Chlamydia pneumoniae infection in vitro and in vivo. J. Agric. Food Chem. 2011, 59, 12836–12842. [Google Scholar] [CrossRef]
- Erenler, R.; Telci, İ.; Elmastas, M.; Aksit, H.; Gul, F.; Tufekci, A.R.; Demirtas, İ.; Kayir, Ö. Quantification of flavonoids isolated from Mentha spicata in selected clones of Turkish mint landraces. Turk. J. Chem. 2018, 42, 1695–1705. [Google Scholar] [CrossRef]
- Dai, L.-M.; Zhao, C.-C.; Jin, H.-Z.; Tang, J.; Shen, Y.-H.; Li, H.-L.; Peng, C.-Y.; Zhang, W.-D. A new ferulic acid ester and other constituents from Dracocephalum peregrinum. Arch. Pharm. Res. 2008, 31, 1325–1329. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Peng, C.; Zhou, Q.-M.; Guo, L.; Liu, Z.-H.; Xiong, L. Alkaloids and flavonoid glycosides from the aerial parts of Leonurus japonicus and their opposite effects on uterine smooth muscle. Phytochemistry 2018, 145, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Monforte, M.T.; Lanuzza, F.; Pergolizzi, S.; Mondello, F.; Tzakou, O.; Galati, E.M. Protective effect of Calamintha officinalis Moench leaves against alcohol-induced gastric mucosa injury in rats. Macroscopic, histologic and phytochemical analysis. Phyther. Res. 2012, 26, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Marin, P.D.; Grayer, R.J.; Veitch, N.C.; Kite, G.C.; Harborne, J.B. Acacetin glycosides as taxonomic markers in Calamintha and Micromeria. Phytochemistry 2001, 58, 943–947. [Google Scholar] [CrossRef]
- Tian, S.; Yu, Q.; Xin, L.; Zhou, Z.S.; Upur, H. Chemical fingerprinting by RP-RRLC-DAD and principal component analysis of Ziziphora clinopodioides from different locations. Nat. Prod. Commun. 2012, 7, 1181–1184. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Hay, A.E.; Marston, A.; Hostettmann, K. Acetylcholinesterase-inhibitory activity of Linarin from Buddleja davidii, structure-activity relationships of related flavonoids, and chemical investigation of Buddleja nitida. Pharm. Biol. 2008, 46, 596–601. [Google Scholar] [CrossRef]
- El-Domiaty, M.M.; Wink, M.; Aal, M.M.A.; Abou-Hashem, M.M.; Abd-Alla, R.H. Antihepatotoxic activity and chemical constituents of Buddleja asiatica Lour. Z. Naturforsch. C 2009, 64, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Zúñiga, M.E.; Cruz-Sosa, F.; Rodríguez-Monroy, M.; Verde-Calvo, J.R.; Vernon-Carter, E.J. Phenylpropanoid production in callus and cell suspension cultures of Buddleja cordata Kunth. Plant. Cell Tissue Organ. Cult. 2009, 97, 39–47. [Google Scholar] [CrossRef]
- Rodríguez-Zaragoza, S.; Ordaz, C.; Avila, G.; Muñoz, J.L.; Arciniegas, A.; De Vivar, A.R. In vitro evaluation of the amebicidal activity of Buddleia cordata (Loganiaceae, H.B.K.) on several strains of Acanthamoeba. J. Ethnopharmacol. 1999, 66, 327–334. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, Y.S.; Choi, E.M. Linarin isolated from Buddleja officinalis prevents hydrogen peroxide-induced dysfunction in osteoblastic MC3T3-E1 cells. Cell. Immunol. 2011, 268, 112–116. [Google Scholar] [CrossRef]
- Sun, M.; Luo, Z.; Liu, Y.; Yang, R.; Lu, L.; Yu, G.; Ma, X.; Liu, A.; Guo, Y.; Zhao, H. Identification of the major components of Buddleja officinalis extract and their metabolites in rat urine by UHPLC-LTQ-orbitrap. J. Food Sci. 2016, 81, H2587–H2596. [Google Scholar] [CrossRef]
- Tai, B.H.; Jung, B.Y.; Cuong, N.M.; Linh, P.T.; Tung, N.H.; Nhiem, N.X.; Huong, T.T.; Anh, N.T.; Kim, J.A.; Kim, S.K.; et al. Total peroxynitrite scavenging capacity of phenylethanoid and flavonoid glycosides from the flowers of Buddleja officinalis. Biol. Pharm. Bull. 2009, 32, 1952–1956. [Google Scholar] [CrossRef] [Green Version]
- Avila Acevedo, J.G.; Castañeda, C.M.C.; Benitez, F.J.C.; Durán, D.A.; Barroso, V.R.; Martínez, C.G.; Muñoz, L.J.L.; Martínez, C.A.; Romo de Vivar, A. Photoprotective activity of Buddleja scordioides. Fitoterapia 2005, 76, 301–309. [Google Scholar] [CrossRef]
- Otsuka, H. Isolation of isolinariins A and B, new flavonoid glycosides from Linaria japonica. J. Nat. Prod. 1992, 55, 1252–1255. [Google Scholar] [CrossRef]
- Vrchovská, V.; Spilková, J.; Valentão, P.; Sousa, C.; Andrade, P.B.; Seabra, R.M. Assessing the antioxidative properties and chemical composition of Linaria vulgaris infusion. Nat. Prod. Res. 2008, 22, 735–746. [Google Scholar] [CrossRef]
- Aydoğrdu, İ.; Zihnioğrlu, F.; Karayildirim, T.; Gülcemal, D.; Alankuş-Çalışkan, Ö.; Bedir, E. α-glucosidase inhibitory constituents of Linaria kurdica subsp. eriocalyx. Nat. Prod. Commun. 2010, 5, 841–844. [Google Scholar] [CrossRef] [Green Version]
- Leslie Gunatilaka, A.; Sotheeswaran, S.; Balasubramaniam, S.; Indumathie Chandrasekara, A.; Badra Sriyani, H. Linarin, a flavone glycoside from Exacum macranthum. Planta Med. 1980, 39, 66–72. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Wang, R.; Guo, F.; Yan, C. Two-dimensional liquid chromatography coupled with mass spectrometry for the analysis of Lobelia chinensis Lour. using an ESI/APCI multimode ion source. J. Sep. Sci. 2008, 31, 2388–2394. [Google Scholar] [CrossRef]
- Wang, T.; Xiao, J.; Hou, H.; Li, P.; Yuan, Z.; Xu, H.; Liu, R.; Li, Q.; Bi, K. Development of an ultra-fast liquid chromatography–tandem mass spectrometry method for simultaneous determination of seven flavonoids in rat plasma: Application to a comparative pharmacokinetic investigation of Ginkgo biloba extract and single pure ginkgo. J. Chromatogr. B 2017, 1060, 173–181. [Google Scholar] [CrossRef]
- El-Hagrassi, A.M.; Ali, M.M.; Osman, A.F.; Shaaban, M. Phytochemical investigation and biological studies of Bombax malabaricum flowers. Nat. Prod. Res. 2011, 25, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-K.; Xu, J.-K.; Zhang, L.; Du, G.-H. Flavonoids from the bran of Avena sativa. Chin. J. Nat. Med. 2012, 10, 110–114. [Google Scholar] [CrossRef]
- Ina, H.; Iida, H. Linarin monoacetate from Thalictrum aquilegifolium. Phytochemistry 1981, 20, 1176–1177. [Google Scholar] [CrossRef]
- Ma, B.-X.; Meng, X.-S.; Tong, J.; Ge, L.-L.; Zhou, G.; Wang, Y.-W. Protective effects of Coptis chinensis inflorescence extract and linarin against carbon tetrachloride-induced damage in HepG2 cells through the MAPK/Keap1-Nrf2 pathway. Food Funct. 2018, 9, 2353–2361. [Google Scholar] [CrossRef]
- Rios, M.Y.; Córdova-Albores, L.C.; Ramírez-Cisneros, M.Á.; King-DÍaz, B.; Lotina-Hennsen, B.; Rivera, I.L.; Miranda-Sánchez, D. Phytotoxic potential of Zanthoxylum affine and its major compound linarin as a possible natural herbicide. ACS Omega 2018, 3, 14779–14787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, G.R.; da Fonseca, T.S.; Martínez-Fructuoso, L.; Simas, R.C.; Silva, F.T.; Salimena, F.R.G.; Alviano, D.S.; Alviano, C.S.; Leitão, G.G.; Pereda-Miranda, R.; et al. Antifungal phenylpropanoid glycosides from Lippia rubella. J. Nat. Prod. 2019, 82, 566–572. [Google Scholar] [CrossRef]
- Liu, S.; Luo, X.; Li, D.; Zhang, J.; Qiu, D.; Liu, W.; She, L.; Yang, Z. Tumor inhibition and improved immunity in mice treated with flavone from Cirsium japonicum DC. Int. Immunopharmacol. 2006, 6, 1387–1393. [Google Scholar] [CrossRef]
- Byun, J.-H.; Choi, C.-W.; Jang, M.-J.; Lim, S.H.; Han, H.J.; Choung, S.-Y. Anti-osteoarthritic mechanisms of Chrysanthemum zawadskii var. latilobum in MIA-induced osteoarthritic rats and interleukin-1β-induced SW1353 human chondrocytes. Medicina (B. Aires). 2020, 56, 685. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, S.; Xuan, Z.; Ge, D.; Chen, X.; Zhang, J.; Wang, Q.; Wu, Y.; Liu, B. The phenolic fraction of Mentha haplocalyx and its constituent linarin ameliorate inflammatory response through inactivation of NF-kB and MAPKs in lipopolysaccharide-induced RAW264.7 cells. Molecules 2017, 22, 811. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-M.; An, D.-Q.; Guo, L.-L.; Yang, N.-H.; Zhang, H. Identification and screening of active components from Ziziphora clinopodioides Lam. in regulating autophagy. Nat. Prod. Res. 2019, 33, 2549–2553. [Google Scholar] [CrossRef]
- Navarrete, A.; Avula, B.; Choi, Y.-W.; Khan, I.A. Chemical fingerprinting of valeriana species: Simultaneous determination of valerenic acids, flavonoids, and phenylpropanoids using liquid chromatography with ultraviolet detection. J. AOAC Int. 2006, 89, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Ahmad, R.; Khare, S.K. Alzheimer’s disease and its treatment by different approaches: A review. Eur. J. Med. Chem. 2021, 216, 113320. [Google Scholar] [CrossRef]
- Feng, X.; Wang, X.; Liu, Y.; Di, X. Linarin inhibits the acetylcholinesterase activity in-vitro and ex-vivo. Iran. J. Pharm. Res. 2015, 14, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhang, J.; Wang, Y.; Cui, K.; Cao, Y.; Wang, L.; Wu, Y. Linarin improves the dyskinesia recovery in Alzheimer’s disease zebrafish by inhibiting the acetylcholinesterase activity. Life Sci. 2019, 222, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.; Giraldez-Alvarez, L.D.; Ávila-Rodriguez, M.; Capani, F.; Galembeck, E.; Neto, A.G.; Barreto, G.E.; Andrade, B. SUR1 receptor interaction with hesperidin and linarin predicts possible mechanisms of action of Valeriana officinalis in Parkinson. Front. Aging Neurosci. 2016, 8, 1–12. [Google Scholar] [CrossRef]
- Jang, M.; Kim, K.-H.; Kim, G.-H. Antioxidant capacity of thistle (Cirsium japonicum) in various drying methods and their protection effect on neuronal PC12 cells and Caenorhabditis elegans. Antioxidants 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Hu, W.; Li, H.; Liu, J.; Zhang, P.; Gu, Y.; Yu, Y.; Wang, W.; Wei, Y. Purification of linarin and hesperidin from Mentha haplocalyx by aqueous two-phase flotation coupled with preparative HPLC and evaluation of the neuroprotective effect of linarin. J. Sep. Sci. 2021, 44, 2496–2503. [Google Scholar] [CrossRef]
- Fernández, S.P.; Wasowski, C.; Loscalzo, L.M.; Granger, R.E.; Johnston, G.A.R.; Paladini, A.C.; Marder, M. Central nervous system depressant action of flavonoid glycosides. Eur. J. Pharmacol. 2006, 539, 168–176. [Google Scholar] [CrossRef]
- Nugroho, A.; Lim, S.C.; Choi, J.; Park, H.J. Identification and quantification of the sedative and anticonvulsant flavone glycoside from Chrysanthemum boreale. Arch. Pharm. Res. 2013, 36, 51–60. [Google Scholar] [CrossRef]
- Li, J.; Hao, L.; Wu, J.; Zhang, J.; Su, J. Linarin promotes osteogenic differentiation by activatingthe BMP-2/RUNX2 pathway via protein kinase A signaling. Int. J. Mol. Med. 2016, 37, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-I.; Kim, Y.H.; Kang, B.G.; Kang, M.K.; Lee, E.J.; Kim, D.Y.; Oh, H.; Oh, S.Y.; Na, W.; Lim, S.S.; et al. Linarin and its aglycone acacetin abrogate actin ring formation and focal contact to bone matrix of bone-resorbing osteoclasts through inhibition of αvβ3 integrin and core-linked CD44. Phytomedicine 2020, 79, 153351. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Chen, Y.; Sun, S.; Xu, X.; Zhan, J.; Yan, Z.; Shang, P.; Pan, X.; Liu, H. Inhibiting TLR4 signaling by linarin for preventing inflammatory response in osteoarthritis. Aging (Albany NY) 2021, 13, 5369–5382. [Google Scholar] [CrossRef]
- Yu, Q.; Li, X.; Cao, X. Linarin could protect myocardial tissue from the injury of Ischemia-reperfusion through activating Nrf-2. Biomed. Pharmacother. 2017, 90, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chengyu, Y.; Long, Z.; Bin, Z.; Hong, L.; Xuefei, S.; Congjuan, L.; Caixia, C.; Yan, X. Linarin protects the kidney against ischemia/reperfusion injury via the inhibition of bioactive ETS2/IL-12. Biol. Pharm. Bull. 2021, 44, 25–31. [Google Scholar] [CrossRef]
- Tian, D.; Yang, Y.; Yu, M.; Han, Z.Z.; Wei, M.; Zhang, H.W.; Jia, H.M.; Zou, Z.M. Anti-inflammatory chemical constituents of Flos Chrysanthemi indici determined by UPLC-MS/MS integrated with network pharmacology. Food Funct. 2020, 11, 6340–6351. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Chen, Y.; Chen, T.; Huang, D.; Li, S.; Cui, S. A systematic study of mechanism of Sargentodoxa cuneata and Patrinia scabiosifolia against pelvic inflammatory disease with dampness-heat stasis syndrome via network pharmacology approach. Front. Pharmacol. 2020, 11, 1856. [Google Scholar] [CrossRef]
- Acevedo, J.G.A.; Espinosa González, A.M.; y Campos, D.M.D.M.; Benitez Flores, J.d.C.; Delgado, T.H.; Maya, S.F.; Contreras, J.C.; López, J.L.M.; García Bores, A.M. Photoprotection of Buddleja cordata extract against UVB-induced skin damage in SKH-1 hairless mice. BMC Complement. Altern. Med. 2014, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.-H.; Han, A.-R.; Chung, H.-J.; Ha, I.-H.; Um, H.-D. Linarin inhibits radiation-induced cancer invasion by downregulating MMP-9 expression via the suppression of NF-κB activation in human non-small-cell lung cancer A549. Nat. Prod. Res. 2019, 33, 3582–3586. [Google Scholar] [CrossRef]
- Peng, C.; Stewart, A.G.; Woodman, O.L.; Ritchie, R.H.; Qin, C.X. Non-alcoholic steatohepatitis: A review of its mechanism, models and medical treatments. Front. Pharmacol. 2020, 11, 1864. [Google Scholar] [CrossRef]
- Zhuang, Z.J.; Shan, C.W.; Li, B.; Pang, M.X.; Wang, H.; Luo, Y.; Liu, Y.L.; Song, Y.; Wang, N.N.; Chen, S.H.; et al. Linarin enriched extract attenuates liver injury and inflammation induced by high-fat high-cholesterol diet in rats. Evid.-Based Complement. Altern. Med. 2017, 2017, 4701570. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Kim, H.K.; Lee, H.S. Hypoglycemic effect of standardized Chrysanthemum zawadskii ethanol extract in high-fat diet/streptozotocin-induced diabetic mice and rats. Food Sci. Biotechnol. 2018, 27, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Kim, S.-E.; Lee, H.S.; Hong, S.-Y.; Kim, S.-E.; Kim, Y.J.; Lee, J.H.; Park, S.J.; Kim, J.H.; Park, Y.-J.; et al. Comparison of linarin content and biological activity in ethanol extraction of Chrysanthemum zawadskii. J. Korean Soc. Food Sci. Nutr. 2016, 45, 1414–1421. [Google Scholar] [CrossRef]
- Chenafa, H.; Mesli, F.; Daoud, I.; Achiri, R.; Ghalem, S.; Neghra, A. In silico design of enzyme α-amylase and α-glucosidase inhibitors using molecular docking, molecular dynamic, conceptual DFT investigation and pharmacophore modelling. J. Biomol. Struct. Dyn. 2021, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vázquez, M.; Ramírez Apan, T.O.; Aguilar, M.H.; Bye, R. Analgesic and antipyretic activities of an aqueous extract and of the flavone linarin of Buddleia cordata. Planta Med. 1996, 62, 137–140. [Google Scholar] [CrossRef]
- Sándor, Z.; Mottaghipisheh, J.; Veres, K.; Hohmann, J.; Bencsik, T.; Horváth, A.; Kelemen, D.; Papp, R.; Barthó, L.; Csupor, D. Evidence supports tradition: The in vitro effects of roman chamomile on smooth muscles. Front. Pharmacol. 2018, 9, 323. [Google Scholar] [CrossRef]
- Ghayur, M.N.; Khan, H.; Gilani, A.H. Antispasmodic, bronchodilator and vasodilator activities of (+)-catechin, a naturally occurring flavonoid. Arch. Pharm. Res. 2007, 30, 970–975. [Google Scholar] [CrossRef]
- Mendel, M.; Chłopecka, M.; Dziekan, N.; Karlik, W. Antispasmodic effect of selected Citrus flavonoids on rat isolated jejunum specimens. Eur. J. Pharmacol. 2016, 791, 640–646. [Google Scholar] [CrossRef] [PubMed]
- das Graças, C.; de Souza, M.; Cyrino, F.Z.; de Carvalho, J.J.; Blanc-Guillemaud, V.; Bouskela, E. Protective effects of Micronized Purified Flavonoid Fraction (MPFF) on a novel experimental model of chronic venous hypertension. Eur. J. Vasc. Endovasc. Surg. 2018, 55, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zeng, L.; Yang, J.; Wang, Y.; Yao, F.; Wu, Y.; Wang, D.; Hu, Y.; Liu, J. Anti-DHAV-1 reproduction and immuno-regulatory effects of a flavonoid prescription on duck virus hepatitis. Pharm. Biol. 2017, 55, 1545–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zeng, L.; Lu, Y.; Yang, Y.; Xu, M.; Wang, Y.; Liu, J. Treatment effect of a flavonoid prescription on duck virus hepatitis by its hepatoprotective and antioxidative ability. Pharm. Biol. 2017, 55, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Zarenezhad, E.; Farjam, M.; Iraji, A. Synthesis and biological activity of pyrimidines-containing hybrids: Focusing on pharmacological application. J. Mol. Struct. 2021, 1230, 129833. [Google Scholar] [CrossRef]
- Yonekawa, M.; Shimizu, M.; Kaneko, A.; Matsumura, J.; Takahashi, H. Suppression of R5-type of HIV-1 in CD4 + NKT cells by Vδ1 + T cells activated by flavonoid glycosides, hesperidin and linarin. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Jiang, W.; Jia, H.; Zheng, L.; Xing, J.; Liu, A.; Du, G. Discovery of multitarget-directed ligands against influenza a virus from compound yizhihao through a predictive system for compound-protein interactions. Front. Cell. Infect. Microbiol. 2020, 10, 16. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Fedotcheva, N.I.; Shimanovsky, N.L. Progestins as anticancer drugs and chemosensitizers, new targets and applications. Pharmaceutics 2021, 13, 1616. [Google Scholar] [CrossRef]
- Talib, W.H.; Alsayed, A.R.; Barakat, M.; Abu-Taha, M.I.; Mahmod, A.I. Targeting drug chemo-resistance in cancer using natural products. Biomedicines 2021, 9, 1353. [Google Scholar] [CrossRef]
- Ailioaie, L.M.; Ailioaie, C.; Litscher, G. Latest innovations and nanotechnologies with curcumin as a nature-inspired photosensitizer applied in the photodynamic therapy of cancer. Pharmaceutics 2021, 13, 1562. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Zhen, Z.G.; Ren, S.H.; Ji, H.M.; Ma, J.H.; Ding, X.M.; Feng, F.Q.; Chen, S.L.; Zou, P.; Ren, J.R.; Jia, L. Linarin suppresses glioma through inhibition of NF-κB/p65 and up-regulating p53 expression in vitro and in vivo. Biomed. Pharmacother. 2017, 95, 363–374. [Google Scholar] [CrossRef]
- Alqahtani, M.S.; Al-Yousef, H.M.; Alqahtani, A.S.; Tabish Rehman, M.; AlAjmi, M.F.; Almarfidi, O.; Amina, M.; Alshememry, A.; Syed, R. Preparation, characterization, and in vitro-in silico biological activities of Jatropha pelargoniifolia extract loaded chitosan nanoparticles. Int. J. Pharm. 2021, 606, 120867. [Google Scholar] [CrossRef]
- Sharif Siam, M.K.; Sarker, A.; Sayeem, M.M.S. In silico drug design and molecular docking studies targeting Akt1 (RAC-alpha serine/threonine-protein kinase) and Akt2 (RAC-beta serine/threonine-protein kinase) proteins and investigation of CYP (cytochrome P450) inhibitors against MAOB (monoamine oxidase B) for OSCC (oral squamous cell carcinoma) treatment. J. Biomol. Struct. Dyn. 2021, 39, 6467–6479. [Google Scholar] [CrossRef]
- De La Peña, J.B.I.; Kim, C.A.; Lee, H.L.; Yoon, S.Y.; Kim, H.J.; Hong, E.Y.; Kim, G.H.; Ryu, J.H.; Lee, Y.S.; Kim, K.M.; et al. Luteolin mediates the antidepressant-like effects of Cirsium japonicum in mice, possibly through modulation of the GABAA receptor. Arch. Pharm. Res. 2014, 37, 263–269. [Google Scholar] [CrossRef]
- Bouazzaoui, A.; Abdellatif, A.A.H.; Al-Allaf, F.A.; Bogari, N.M.; Al-Dehlawi, S.; Qari, S.H. Strategies for vaccination: Conventional vaccine approaches versus new-generation strategies in combination with adjuvants. Pharmaceutics 2021, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.B.; Pal, S.; Javed, N.; Shekhar, R. COVID-19 vaccination in developing nations: Challenges and opportunities for innovation. Infect. Dis. Rep. 2021, 13, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Jirjees, F.; Saad, A.K.; Al Hano, Z.; Hatahet, T.; Al Obaidi, H.; Dallal Bashi, Y.H. COVID-19 treatment guidelines: Do they really reflect best medical practices to manage the pandemic? Infect. Dis. Rep. 2021, 13, 259–284. [Google Scholar] [CrossRef]
- Shah, S.A.A.; ul Hassan, S.S.; Bungau, S.; Si, Y.; Xu, H.; Rahman, M.H.; Behl, T.; Gitea, D.; Pavel, F.-M.; Corb Aron, R.A.; et al. Chemically diverse and biologically active secondary metabolites from marine Phylum chlorophyta. Mar. Drugs 2020, 18, 493. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Balmeh, N.; Mohammadi, N.; Sadeghian-Rizi, T. The novel drug discovery to combat COVID-19 by repressing important virus proteins involved in pathogenesis using medicinal herbal compounds. Avicenna J. Med. Biotechnol. 2021, 13, 108. [Google Scholar] [CrossRef]
- Egert, S.; Rimbach, G. Which sources of flavonoids: Complex diets or dietary supplements? Adv. Nutr. 2011, 2, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Slámová, K.; Kapešová, J.; Valentová, K. “Sweet flavonoids”: Glycosidase-catalyzed modifications. Int. J. Mol. Sci. 2018, 19, 2126. [Google Scholar] [CrossRef] [Green Version]
- Fernández, S.; Wasowski, C.; Paladini, A.C.; Marder, M. Sedative and sleep-enhancing properties of linarin, a flavonoid-isolated from Valeriana officinalis. Pharmacol. Biochem. Behav. 2004, 77, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Chari, V.M.; Jordan, M.; Wagner, H.; Thies, P.W. A 13C-NMR study of the structure of an acyllinarin from Valeriana wallichii. Phytochemistry 1977, 16, 1110–1112. [Google Scholar] [CrossRef]
- Martínez-Vázquez, M.; Ramírez Apan, T.O.; Lastra, A.L.; Bye, R. A comparative study of the analgesic and anti-inflammatory activities of pectolinarin isolated from Cirsium subcoriaceum and linarin isolated from Buddleia cordata. Planta Med. 1998, 64, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Sung, K.; Yim, D.; Lee, S.; Lee, C.; Ha, N.; Kim, K. The effect of linarin on LPS-induced cytokine production and nitric oxide inhibition in murine macrophages cell line RAW264.7. Arch. Pharm. Res. 2002, 25, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Kang, H.; Sun, H.; Lee, Y.; Park, J.; Chun, S. Isolation and identification of flavonoids from Gujeolcho (Chrysanthemum zawadskii var. latilobum) as inhibitor of histamine release. Food Sci. Biotechnol. 2012, 21, 613–617. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, X.; Lan, Y.; Han, C.; Zhang, Y.; Chen, G. Linarin sensitizes tumor necrosis factor-related apoptosis (TRAIL)-induced ligand-triggered apoptosis in human glioma cells and in xenograft nude mice. Biomed. Pharmacother. 2017, 95, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.-Y.; Zhang, Y.-P.; Gao, J.-L.; Yu, J.-J.; Lei, J.; Zhang, Z.-R.; Li, B.; Zhan, R.-J.; Chen, S.-H. Combined antihypertensive effect of luteolin and buddleoside enriched extracts in spontaneously hypertensive rats. J. Ethnopharmacol. 2013, 150, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, J.H.; Seo, M.J.; Eom, S.H.; Kim, W. Linarin down-regulates phagocytosis, pro-inflammatory cytokine production, and activation marker expression in RAW264.7 macrophages. Food Sci. Biotechnol. 2016, 25, 1437–1442. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, H.S.; Kim, D.K.; Lee, J.H.; Yoo, Y.H.; Kim, J.Y.; Park, S.K.; Nam, S.T.; Kim, H.W.; Park, Y.H.; et al. The extract of Chrysanthemum zawadskii var. latilobum ameliorates collagen-induced arthritis in mice. Evid.-Based Complement. Altern. Med. 2016, 2016, 3915013. [Google Scholar] [CrossRef]
- Han, X.; Wu, Y.I.-C.; Meng, M.; Sun, Q.-S.; Gao, S.-M.; Sun, H. Linarin prevents LPS-induced acute lung injury by suppressing oxidative stress and inflammation via inhibition of TXNIP/NLRP3 and NF-κB pathways. Int. J. Mol. Med. 2018, 42, 1460–1472. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Yang, J.; Wei, X.; Xu, Q.; Qin, M. Separation of acteoside and linarin from Buddlejae Flos by high-speed countercurrent chromatography and their anti-inflammatory activities. J. Sep. Sci. 2020, 48, 1–8. [Google Scholar] [CrossRef]
- Benitez, F.J.C.; Acevedo, J.G.A.; Castan, C.M.C.; Dura, D.A.; Barroso, V.R.; Martı, C.G.; Vivar, A.R. De Photoprotective activity of Buddleja scordioides. Fitoterapia 2005, 76, 301–309. [Google Scholar] [CrossRef]
- Chow, N.; Fretz, M.; Hamburger, M.; Butterweck, V. Telemetry as a tool to measure sedative effects of a valerian root extract and its single constituents in mice. Planta Med. 2011, 77, 795–803. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mottaghipisheh, J.; Taghrir, H.; Boveiri Dehsheikh, A.; Zomorodian, K.; Irajie, C.; Mahmoodi Sourestani, M.; Iraji, A. Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review. Pharmaceuticals 2021, 14, 1104. https://doi.org/10.3390/ph14111104
Mottaghipisheh J, Taghrir H, Boveiri Dehsheikh A, Zomorodian K, Irajie C, Mahmoodi Sourestani M, Iraji A. Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review. Pharmaceuticals. 2021; 14(11):1104. https://doi.org/10.3390/ph14111104
Chicago/Turabian StyleMottaghipisheh, Javad, Hadi Taghrir, Anahita Boveiri Dehsheikh, Kamiar Zomorodian, Cambyz Irajie, Mohammad Mahmoodi Sourestani, and Aida Iraji. 2021. "Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review" Pharmaceuticals 14, no. 11: 1104. https://doi.org/10.3390/ph14111104