
Preparing Collared Peccary (Pecari tajacu Linnaeus, 1758) for Reintroduction into the Wild: A Screening for Parasites and Hemopathogens of a Captive Population

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Peccaries Management
2.2. Biological Sample Collection
- -
- Flotation: A portion will be transferred to Petri dishes containing 2.5% potassium dichromate, properly labeled, where it will undergo maceration and be stored at room temperature for approximately ten days to allow for the sporulation of protozoan oocysts. After this period, 3 mL of this sample will be transferred to a 15 mL Falcon tube containing 12 mL of Sheater’s Solution to separate the feces from potassium dichromate. This material will be centrifuged at 1500 rpm for 15 min. Subsequently, a drop of the supernatant will be placed on a slide and covered with a coverslip for morphometry and identification of oocysts under an Olympus BX 40 Microscope [23].
- -
- Spontaneous sedimentation in water: Another portion of the feces will be strained and transferred to a Hoffman bowl, where it will rest for one to 24 h. Subsequently, a drop of the denser material will be mixed with 10% formalin and placed on a slide, covered with a coverslip for the identification of helminth eggs under an Olympus BX 40 Microscope [24].
2.3. Integrated Approach of Polymerase Chain Reaction (PCR) and Nucleotide Sequencing for Parasite Detection and Characterization
2.4. Analysis of Gene Transcript Sequences
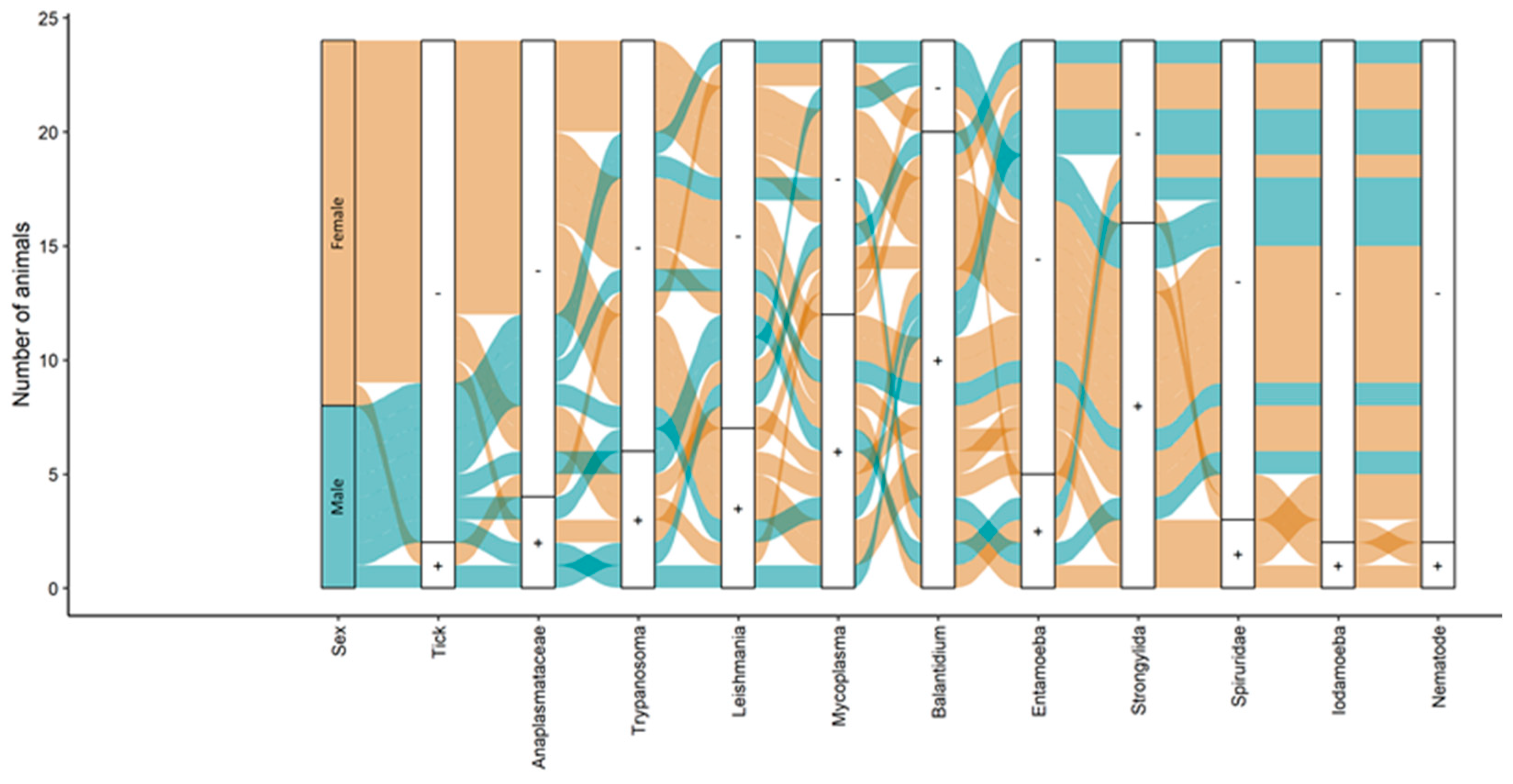
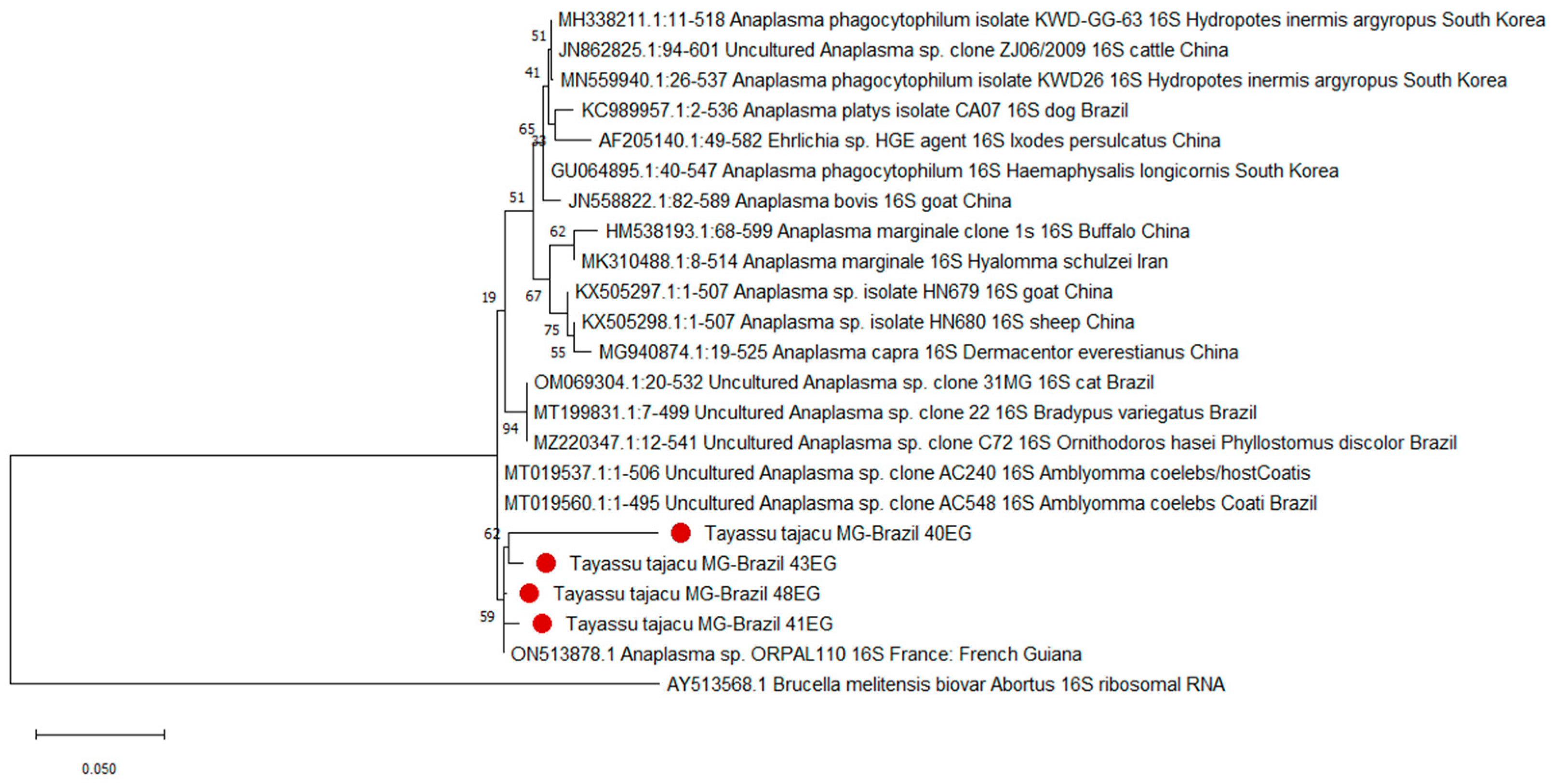
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bissonette, J.A. Ecology and Social Behavior of the Collared Peccary in Big Bend National Park, Texas; U.S. Department of the Interior, National Park Service: Washington, DC, USA, 1982. [Google Scholar]
- Sowls, L.K. Javelinas and Other Peccaries: Their Biology, Management, and Use; Texas A and M University Press: College Station, TX, USA, 1997. [Google Scholar]
- Bogoni, J.A.; Peres, C.A.; Ferraz, K.M.P.M.B. Extent, intensity and drivers of mammal defaunation: A continental-scale analysis across the Neotropics. Sci. Rep. 2020, 10, 14750. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Zamorano, J.; Benítez-López, A.; Santini, L.; Hilbers, J.P.; Huijbregts, M.A.J.; Schipper, A.M. Combined effects of land use and hunting on distributions of tropical mammals. Conserv. Biol. 2020, 34, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Keyghobadi, N. The genetic implications of habitat fragmentation for animals. Can. J. Zool. 2007, 85, 1049–1064. [Google Scholar] [CrossRef]
- Alves, R.R.N.; Rosa, I.L. Trade of animals used in Brazilian traditional medicine: Trends and implications for conservation. Hum. Ecol. 2010, 38, 691–704. [Google Scholar] [CrossRef]
- Griffiths, B.M.; Gilmore, M.P. Differential use of game species in an Amazonian Indigenous community: Navigating economics, subsistence, and social norms. J. Ethnobiol. 2022, 42, 1–12. [Google Scholar] [CrossRef]
- Alves, R.R.N.; Feijó, A.; Barboza, R.R.D.; Souto, W.M.S.; Fernandes-Ferreira, H.; Cordeiro-Estrela, P.; Langguth, A. Game mammals of the Caatinga biome. Ethnobiol. Conserv. 2016, 5, 5. [Google Scholar] [CrossRef]
- Marinho Neto, S.P.; Benigno, R.N.M.; de Albuquerque, N.I. Biota parasitária gastrointestinal de caititus “Tayassu tajacu Linnaeus 1758” (Pecari tajacu Jacq.) criados em cativeiro. In Seminário Científico da UFRA; Embrapa Amazônia Oriental: Belém, Brazil, 2009; Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/44071/1/59.pdf (accessed on 15 October 2023).
- Schettini, L.; Li, O.; Gálvez, H.; Montoya, E.; Sánchez, N. Perfil bioquímico sanguíneo hepático y renal en el sajino (Tayassu tajacu) criado en cautiverio en la amazonía peruana. Rev. Investig. Vet. Perú 2005, 16, 175–179. [Google Scholar] [CrossRef]
- Sampaio, M.; Sianto, L.; Chame, M.; Saldanha, B.; Brener, B. Intestinal Parasites in Pecari tajacu and Sus scrofa domesticus in the Caatinga from Southeastern Piauí, Brazil. J. Parasitol. 2023, 109, 274–287. [Google Scholar] [CrossRef]
- Vieira, R.F.C.; Molento, M.B.; Guimarães, A.M.S.; Santos, A.P.; Bonat, M.; Javorouski, M.L.; Popp, L.; Santos, L.C.; Moraes, W.; Cubas, Z.S. Use of a Mycoplasma suis-PCR protocol for screening a population of captive peccaries (Tayassu tajacu and Tayassu pecari). Rev. Bras. Parasitol. Veterinária 2011, 20, 75–77. [Google Scholar] [CrossRef]
- Herrera, H.M.; Abreu, U.G.P.; Keuroghlian, A.; Freitas, T.P.; Jansen, A.M. The role played by sympatric collared peccary (Tayassu tajacu), white-lipped peccary (Tayassu pecari), and feral pig (Sus scrofa) as maintenance hosts for Trypanosoma evansi and Trypanosoma cruzi in a sylvatic area of Brazil. Parasitol. Res. 2008, 103, 619–624. [Google Scholar] [CrossRef]
- Ewen, J.G.; Soorae, P.S.; Canessa, S. Reintroduction objectives, decisions and outcomes: Global perspectives from the herpetofauna. Anim. Conserv. 2014, 17, 74–81. [Google Scholar] [CrossRef]
- Bernhardt, J.R.; O’Connor, M.I.; Sunday, J.M.; Gonzalez, A. Life in fluctuating environments. Philos. Trans. R. Soc. B 2020, 375, 20190454. [Google Scholar] [CrossRef] [PubMed]
- Rezende, R.A.; Prado Filho, J.F.; Sobreira, F.G. Análise temporal da flora nativa no entorno de unidades de conservação: APA Cachoeira das Andorinhas e FLOE Uaimii, Ouro Preto, MG. Rev. Árvore 2011, 35, 435–443. [Google Scholar] [CrossRef]
- Silva, R.A.; Pimenta, E.M.L.; Rassy, F.B.; Mattoso, C.R.S.; Lima, M.P.A.; Beier, S.L. Cardiorespiratory and sedation evaluation of the combination of dexmedetomidine-butorphanol-midazolam or detomidine butorphanol-midazolam in collared peccaries (Pecari tajacu). Arq. Bras. Med. Veterinária Zootec. 2020, 72, 452–460. [Google Scholar] [CrossRef]
- Oliveira-Sequeira, T.C.G.; Amarante, A.F.T.; Ferrari, T.B.; Nunes, L.C. Prevalence of intestinal parasites in dogs from São Paulo State, Brazil. Vet. Parasitol. 2002, 103, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.S.; Pirmez, C.; Pires, M.Q.; Brazil, R.P.; Pacheco, R.S. PCR-based diagnosis for detection of Leishmania in skin and blood of rodents from an endemic area of cutaneous and visceral leishmaniasis in Brazil. Vet. Parasitol. 2005, 129, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Camargo-Neves, V.L.F. Manual de vigilância acarológica. In Manual de Vigilância Acarológica; Secretaria de Estado da Saúde: São Paulo, Brazil, 2004; 62p. [Google Scholar]
- Aragão, H.; Fonseca, F. Notas de ixodologia: VIII. Lista e chave para os representantes da fauna ixodológica brasileira: Notas de ixolodologia. Memórias Inst. Oswaldo Cruz 1961, 59, 115–129. [Google Scholar] [CrossRef]
- Barros-Battesti, D.M.; Arzua, M.; Bechara, G.H. Carrapatos de Importância Médico-Veterinária da Região Neotropical: Um Guia Ilustrado Para Identificação de Espécies; Vox/ICTTD-3/Butantan: São Paulo, Brazil, 2006. [Google Scholar]
- Duszynski, D.W.; Wilber, P.G. A guideline for the preparation of species descriptions in the Eimeriidae. J. Parasitol. 1997, 83, 333–336. [Google Scholar] [CrossRef]
- Hoffman, W.A.; Pons, J.A.; Janer, J.L. The sedimentation-concentration method in schistosomiasis mansoni. Puerto Rico J. Public Health Trop. Med. 1934, 9, 283–291. [Google Scholar]
- Barbosa, A.S.; Bastos, O.M.P.; Uchôa, C.M.A.; Pissinatti, A.; Bastos, A.C.M.P.; Souza, I.V.; Dib, L.V.; Azevedo, E.P.; Siqueira, M.P.; Cardozo, M.L. Comparison of five parasitological techniques for laboratory diagnosis of Balantidium coli cysts. Rev. Bras. Parasitol. Veterinária 2016, 25, 286–292. [Google Scholar] [CrossRef]
- McPherson, R.A.; Pincus, M.R. Henry’s Clinical Diagnosis and Management by Laboratory Methods E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Kawahara, M.; Tajima, T.; Torii, H.; Yabutani, M.; Ishii, J.; Harasawa, M.; Isogai, E.; Rikihisa, Y. Ehrlichia chaffeensis infection of sika deer, Japan. Emerg. Infect. Dis. 2009, 15, 1991. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR assay for detection of granulocytic ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J.; Ruiz-Fons, F.; Naranjo, V.; Torina, A.; Rodríguez, O.; Gortázar, C. Evidence of Anaplasma infections in European roe deer (Capreolus capreolus) from southern Spain. Res. Vet. Sci. 2008, 84, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.A.G.; Rabelo, E.M.L.; Ribeiro, M.F.B. Molecular detection of tick-borne pathogens of the family Anaplasmataceae in Brazilian brown brocket deer (Mazama gouazoubira, Fischer, 1814) and marsh deer (Blastocerus dichotomus, Illiger, 1815). Transbound. Emerg. Dis. 2012, 59, 353–360. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J.; Massung, R.F.; Wong, S.J.; Chu, F.K.; Lutz, H.; Meli, M.; Von Loewenich, F.D.; Grzeszczuk, A.; Torina, A.; Caracappa, S. Sequence analysis of the msp4 gene of Anaplasma phagocytophilum strains. J. Clin. Microbiol. 2005, 43, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Bown, K.J.; Lambin, X.; Ogden, N.H.; Petrovec, M.; Shaw, S.E.; Woldehiwet, Z.; Birtles, R. High-resolution genetic fingerprinting of European strains of Anaplasma phagocytophilum by use of multilocus variable-number tandem-repeat analysis. J. Clin. Microbiol. 2007, 45, 1771–1776. [Google Scholar] [CrossRef]
- Criado-Fornelio, A.; Martinez-Marcos, A.; Buling-Saraña, A.; Barba-Carretero, J.C. Presence of Mycoplasma haemofelis, Mycoplasma haemominutum and piroplasmids in cats from southern Europe: A molecular study. Vet. Microbiol. 2003, 93, 307–317. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; Rabelo, E.M.L.; Lacerda, A.C.R.; Borges, P.A.L.; Tomás, W.M.; Pellegrin, A.O.; Tomich, R.G.P.; Ribeiro, M.F.B. Molecular detection and identification of hemoparasites in pampas deer (Ozotoceros bezoarticus Linnaeus, 1758) from the Pantanal Brazil. Ticks Tick-Borne Dis. 2013, 4, 341–345. [Google Scholar] [CrossRef]
- Bezerra, F.S.B.; Garcia, H.A.; Alves, H.M.; Oliveira, I.R.S.; Silva, A.E.; Teixeira, M.M.G.; Batista, J.S. Trypanosoma vivax nos tecidos testicular e epididimário de ovinos experimentalmente infectados. Pesqui. Veterinária Bras. 2008, 28, 575–582. [Google Scholar] [CrossRef]
- Vallejo, G.A.; Guhl, F.; Chiari, E.; Macedo, A.M. Species specific detection of Trypanosoma cruzi and Trypanosoma rangeli in vector and mammalian hosts by polymerase chain reaction amplification of kinetoplast minicircle DNA. Acta Trop. 1999, 72, 203–212. [Google Scholar] [CrossRef]
- Schönian, G.; Nasereddin, A.; Dinse, N.; Schweynoch, C.; Schallig, H.D.F.H.; Presber, W.; Jaffe, C.L. PCR diagnosis and characterization of Leishmania in local and imported clinical samples. Diagn. Microbiol. Infect. Dis. 2003, 47, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Graça, G.C.; Volpini, A.C.; Romero, G.A.S.; Oliveira Neto, M.P.; Hueb, M.; Porrozzi, R.; Boité, M.C.; Cupolillo, E. Development and validation of PCR-based assays for diagnosis of American cutaneous leishmaniasis and identificatio nof the parasite species. Memórias Inst. Oswaldo Cruz 2012, 107, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Zahler, M.; Rinder, H.; Schein, E.; Gothe, R. Detection of a new pathogenic Babesia microti-like species in dogs. Vet. Parasitol. 2000, 89, 241–248. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.A.G.; Rabelo, É.M.L.; Ribeiro, M.F.B. Detection of Theileria and Babesia in brown brocket deer (Mazama gouazoubira) and marsh deer (Blastocerus dichotomus) in the State of Minas Gerais, Brazil. Vet. Parasitol. 2011, 177, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.A.G.; Rabelo, E.M.L.; Lima, P.C.S.; Chaves, B.N.; Ribeiro, M.F.B. Post-mortem hemoparasite detection in free-living Brazilian brown brocket deer (Mazama gouazoubira, Fischer 1814). Rev. Bras. Parasitol. Veterinária 2014, 23, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.A.G.; Valente, P.C.L.G.; Paes, P.R.O.; Vasconcelos, A.V.; Silvestre, B.T.; Ribeiro, M.F.B. The first clinical and laboratory evidence of co-infection by Anaplasma phagocytophilum and Ehrlichia canis in a Brazilian dog. Ticks Tick-Borne Dis. 2015, 6, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Ramos, V.d.N.; Piovezan, U.; Franco, A.H.A.; Osava, C.F.; Herrera, H.M.; Szabó, M.P.J. Feral pigs as hosts for Amblyomma sculptum (Acari: Ixodidae) populations in the Pantanal, Mato Grosso do Sul, Brazil. Exp. Appl. Acarol. 2014, 64, 393–406. [Google Scholar] [CrossRef]
- WHO. Leishmaniases. Available online: https://www.who.int/news-room/questions-and-answers/item/leishmaniasis (accessed on 1 November 2023).
- Desjeux, P. Leishmaniasis: Current situation and new perspectives. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 305–318. [Google Scholar] [CrossRef]
- Desquesnes, M.; Holzmuller, P.; Lai, D.; Dargantes, A.; Lun, Z.R.; Jittaplapong, S. Trypanosoma evansi and surra: A review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. BioMed Res. Int. 2013, 2013, 194176. [Google Scholar] [CrossRef]
- Gutierrez, C.; Desquesnes, M.; Touratier, L.; Büscher, P. Trypanosoma evansi: Recent outbreaks in Europe. Vet. Parasitol. 2010, 174, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Aregawi, W.G.; Agga, G.E.; Abdi, R.D.; Büscher, P. Systematic review and meta-analysis on the global distribution, host range, and prevalence of Trypanosoma evansi. Parasites Vectors 2019, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Ventura, R.M.; Takata, C.S.A.; Silva, R.A.M.S.; Nunes, V.L.; Takeda, G.F.; Teixeira, M.M.G. Molecular and morphological studies of Brazilian Trypanosoma evansi stocks: The total absence of kDNA in trypanosomes from both laboratory stocks and naturally infected domestic and wild mammals. J. Parasitol. 2000, 86, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Herrera, H.M.; Norek, A.; Freitas, T.P.T.; Rademaker, V.; Fernandes, O.; Jansen, A.M. Domestic and wild mammals infection by Trypanosoma evansi in a pristine area of the Brazilian Pantanal region. Parasitol. Res. 2005, 96, 121–126. [Google Scholar] [CrossRef]
- Perrone, T.; Sánchez, E.; Hidalgo, L.; Mijares, A.; Balzano-Nogueira, L.; Gonzatti, M.I.; Aso, P. Infectivity and virulence of Trypanosoma evansi and Trypanosoma equiperdum Venezuelan strains from three different host species. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef]
- Duron, O.; Koual, R.; Musset, L.; Buysse, M.; Lambert, Y.; Jaulhac, B.; Blanchet, D.; Alsibai, K.D.; Lazrek, Y.; Epelboin, L. Novel Chronic Anaplasmosis in Splenectomized Patient, Amazon Rainforest. Emerg. Infect. Dis. 2022, 28, 1673. [Google Scholar] [CrossRef]
- Millán, J.; Travaini, A.; Cevidanes, A.; Sacristán, I.; Rodríguez, A. Assessing the natural circulation of canine vector-borne pathogens in foxes, ticks and fleas in protected areas of Argentine Patagonia with negligible dog participation. Int. J. Parasitol. Parasites Wildl. 2019, 8, 63–70. [Google Scholar] [CrossRef]
- Willi, B.; Novacco, M.; Meli, M.L.; Wolf-Jäckel, G.A.; Boretti, F.S.; Wengi, N.; Lutz, H.; Hofmann-Lehmann, R. Haemotropic mycoplasmas of cats and dogs: Transmission, diagnosis, prevalence and importance in Europe. Schweiz. Arch. Tierheilkd. 2010, 152, 237. [Google Scholar] [CrossRef]
- Biondo, A.W.; Santos, A.P.; Guimarães, A.M.S.; Vieira, R.F.C.; Vidotto, O.; Macieira, D.B.; Almosny, N.R.P.; Molento, M.B.; Timenetsky, J.; Morais, H.A. A review of the occurrence of hemoplasmas (hemotrophic mycoplasmas) in Brazil. Rev. Bras. Parasitol. Veterinária 2009, 18, 1–7. [Google Scholar] [CrossRef]
- Dias, G.B.; do Amaral, R.B.; Gatto, I.R.H.; Lapera, I.M.; de Oliveira, L.G.; Hoppe, E.G.L.; Machado, R.Z.; André, M.R. Molecular detection of Mycoplasma suis in captive white-lipped peccaries (Tayassu pecari) and wild boars (Sus scrofa) in Brazil. Comp. Immunol. Microbiol. Infect. Dis. 2019, 63, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Benvenuto, A.; Sacristán, C.; Ewbank, A.C.; Sacristán, I.; Zamana-Ramblas, R.; Gravena, W.; Mello, M.D.; da Silva, V.M.F.; Marmontel, M.; Carvalho, V.L.; et al. Hemotropic Mycoplasma spp. in Aquatic Mammals, Amazon Basin, Brazil. Emerg. Infect. Dis. 2022, 28, 2556. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.P.; Roy, S.S.; Cowan, D.P.; Massei, G.; Smith, G.C.; Ji, W.; Rossi, S.; Woodroffe, R.; Wilson, G.J.; Delahay, R.J. Options for the control of disease 2: Targeting hosts. In Management of Disease in Wild Mammals; Springer: Berlin/Heidelberg, Germany, 2009; pp. 121–146. [Google Scholar]
- Prevedello, J.A.; Vieira, M.V. Does the type of matrix matter? A quantitative review of the evidence. Biodivers. Conserv. 2010, 19, 1205–1223. [Google Scholar] [CrossRef]
- Bowman, D.D. Ascaris and Toxocara as foodborne and waterborne pathogens. Res. Vet. Sci. 2021, 135, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Ijaz, M.; Ayyub, R.M.; Ghaffar, A.; Ghauri, H.N.; Aziz, M.U.; Ali, S.; Altaf, M.; Awais, M.; Naveed, M. Balantidium coli in domestic animals: An emerging protozoan pathogen of zoonotic significance. Acta Trop. 2020, 203, 105298. [Google Scholar] [CrossRef] [PubMed]
- Sodeman, W.A., Jr. Intestinal protozoa: Amebas. In Medical Microbiology, 4th ed.; National Library of Medicine: Bethesda, MD, USA, 1996. [Google Scholar]
- Melo, Y.J.O.; Ferraz, H.T.; Saturnino, K.C.; Silva, T.D.P.; Braga, I.A.; Amaral, A.V.C.; Meirelles-Bartoli, R.B.; Ramos, D.G.S. Gastrointestinal parasites in captive and free-living wild birds in Goiania Zoo. Braz. J. Biol. 2021, 82, e240386. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.G.; Batista, A.I.V.; Silva, W.S.I.; Oliveira Neto, M.B.; Schettino, S.C.; Oliveira, M.R.; Ramos, R.A.N.; Alves, L.C.; Bezerra-Santos, M.; Lima, V.F.S. Gastrointestinal parasites in captive wild animals from two Brazilian Zoological Gardens. Res. Soc. Dev. 2022, 11, e28411426637. [Google Scholar] [CrossRef]
- Mbaya, A.W.; Udendeye, U.J. Gastrointestinal parasites of captive and free-roaming primates at the Afi Mountain Primate Conservation Area in Calabar, Nigeria and their zoonotic implications. Pak. J. Biol. Sci. PJBS 2011, 14, 709–714. [Google Scholar] [CrossRef]
- Gray, M.J.; Spatz, J.A.; Carter, E.D.; Yarber, C.M.; Wilkes, R.P.; Miller, D.L. Poor biosecurity could lead to disease outbreaks in animal populations. PLoS ONE 2018, 13, e0193243. [Google Scholar] [CrossRef]
- Dobson, A.; Cattadori, I.; Holt, R.D.; Ostfeld, R.S.; Keesing, F.; Krichbaum, K.; Rohr, J.R.; Perkins, S.E.; Hudson, P.J. Sacred cows and sympathetic squirrels: The importance of biological diversity to human health. PLoS Med. 2006, 3, e231. [Google Scholar] [CrossRef]
- Mangini, P.R.S. Medicina da Conservação: Aspectos Gerais; Roca: São Paulo, Brazil, 2006; pp. 1258–1268. [Google Scholar]
- Colombo, S.A.P.; Grencis, R.K. Immunity to Soil-Transmitted Helminths: Evidence From the Field and Laboratory Models. Front. Immunol. 2020, 11, 1286. [Google Scholar] [CrossRef] [PubMed]
- Ovaska, U.; Bläuer, A.; Kroløkke, C.; Kjetså, M.; Kantanen, J.; Honkatukia, M. The conservation of native domestic animal breeds in Nordic countries: From genetic resources to cultural heritage and good governance. Animals 2021, 11, 2730. [Google Scholar] [CrossRef] [PubMed]
- Paulson, N.; Laudati, A.; Doolittle, A.; Welch-Devine, M.; Pena, P. Indigenous peoples Participation in global conservation: Looking beyond headdresses and face paint. Environ. Values 2012, 21, 255–276. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Hemopathogen | Sequence (5′→ 3′) | Primer | Target | Products (pb) | References |
|---|---|---|---|---|---|
| Babesia/Theileria 1st reaction | CGGGATCCAACCTGGTTGATCCTGC CCGAATTCCTTGTTACGACTTCTC | RIB-19 RIB-20 | 18S rRNA | 1700 | [39] |
| 2nd reaction | ACCTCACCAGGTCCAGACAG GTACAAAGGGCAGGGACGTA | BAB-rumF BAB-rumR | 18S rRNA | 430 | [40] |
| A. marginale/A. ovis 1st reaction | GGGAGCTCCTATGAATTACAGAGAATTGTTTAC CCGGATCCTTAGCTGAACAGGAATCTTGC | MSP45 MSP43 | msp4 | 872 | [29] |
| 2nd reaction | CGCCAGCAAACTTTTCCAAA ATATGGGGACACAGGCAAAT | AnapF AnapR | msp4 | 294 | [30] |
| A. phagocytophilum 1st reaction | ATGAATTACAGAGAATTGCTTGTAGG TTAATTGAAAGCAAATCTTGCTCCTATG | Msp4AP1F Msp4AP1R | msp4 | - | [31] |
| 2nd reaction | CTATTGGYGGNGCYAGAGT GTTCATCGAAAATTCCGTGGTA | Msp4AP2F Msp4AP2R | msp4 | 450 | [32] |
| Monocytic Anaplasmataceae 1st reaction | ACGGACAATTGCTTATAGCCTT ACAACTTTTATGGATTAGCTAAAT | NS16SCH1F NS16SCH1R | 16S rRNA | 1195 | [27] |
| 2nd reaction | GGGCACGTAGGTGGACTAG CCTGTTAGGAGGGATACGAC | NS16SCH2F NS16SCH2R | 16S rRNA | 443 | [27] |
| Granulocytic/platelet Anaplasmataceae 1st reaction | CACATGCAAGTCGAACGGATTATTC TTCCGTTAAGAAGGATCTAATCTCC | GE3a GE10r | 16S rRNA | 932 | [28] |
| 2nd reaction | AACGGATTATTCTTTATAGCTTGCT GGCAGTATTAAAAGCAGCTCCAGG | GE9f GE2 | 16S rRNA | 546 | [28] |
| Trypanosoma evansi 1st reaction | GCACAGTATGCAACCAAAAA GTGGTCAACAGGGAGAAAAT | Te1F Te1R | ITS | 280 | [34] |
| 2nd reaction | CATGTATGTGTTTCTATATG | Te2F | ITS | 219 | [34] |
| Trypanosoma vivax | GCCATCGCCAAGTACCTCGCGA TTAGAATTCCCAGGAGTTCTTGATGATCCAGTA | Tvi2 DTO156 | Catepsin L gene | 177 | [35] |
| Trypanosoma cruzi | AAATAATGTACGGGKGAGATGCATGA GGTTCGATTGGGGTTGGTGTAATATA- | S35 S36 | kDNA | 333 | [36] |
| Leishmania spp. | GGACGAGATCGAGCGCATGGT TCCTTCGACGCCTCCTGGTTG | hsp70F hsp70R | Hsp70 | 234 pb | [37] |
| Leishmania spp. | CTGGATCATTTTCCGATG TGATACCACTTATCGCACTT | LITSR L5.8S | ITS 1 | 300–350 | [38] |
| Hemotropic Mycoplasma spp. | ATACGGCCCATATTCCTACG TGCTCCACCACTTGTTCA | HBT-F 16S Fw HBT-R 16S Rv | 16S rRNA | 618 | [33] |
| Species | Targets | |
|---|---|---|
| ITS1 | HSP70 | |
| Leishmania infantum | ~190, 70 and 60 | 90, 80 and 70 |
| Leishmania amazonensis | ~190 and 140 | 230 |
| Leishmania braziliensis | ~150 and 140 | 140 |
| Leishmania guyanensis | ~150 and 130 | 180 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silveira, J.A.G.d.; Moreira, S.M.; Nascimento, A.F.d.; Oliveira, M.M.d.; Santos, H.A.d.; Estevam, L.G.T.d.M.; Pereira, C.R.; Oliveira, A.G.G.; D’Elia, M.L.; Araujo, A.d.C.; et al. Preparing Collared Peccary (Pecari tajacu Linnaeus, 1758) for Reintroduction into the Wild: A Screening for Parasites and Hemopathogens of a Captive Population. Pathogens 2024, 13, 47. https://doi.org/10.3390/pathogens13010047
Silveira JAGd, Moreira SM, Nascimento AFd, Oliveira MMd, Santos HAd, Estevam LGTdM, Pereira CR, Oliveira AGG, D’Elia ML, Araujo AdC, et al. Preparing Collared Peccary (Pecari tajacu Linnaeus, 1758) for Reintroduction into the Wild: A Screening for Parasites and Hemopathogens of a Captive Population. Pathogens. 2024; 13(1):47. https://doi.org/10.3390/pathogens13010047
Chicago/Turabian StyleSilveira, Júlia Angélica Gonçalves da, Simone Magela Moreira, Ariane Flávia do Nascimento, Marco Miguel de Oliveira, Hudson Andrade dos Santos, Letícia Gracielle Tôrres de Miranda Estevam, Carine Rodrigues Pereira, Anna Gabriela Guimarães Oliveira, Mirella Lauria D’Elia, Andreina de Carvalho Araujo, and et al. 2024. "Preparing Collared Peccary (Pecari tajacu Linnaeus, 1758) for Reintroduction into the Wild: A Screening for Parasites and Hemopathogens of a Captive Population" Pathogens 13, no. 1: 47. https://doi.org/10.3390/pathogens13010047