
Impact of the Paper by Allen and Humphreys (1979) on Anti-Tick Vaccine Research
Immunology and Vaccines Laboratory, Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro, Queretaro 76140, Mexico
Pathogens 2022, 11(11), 1253; https://doi.org/10.3390/pathogens11111253
Submission received: 10 October 2022
/
Revised: 24 October 2022
/
Accepted: 26 October 2022
/
Published: 28 October 2022
(This article belongs to the Special Issue 10th Anniversary of Pathogens—Classic Papers in Tick Research)
Abstract
:The classic paper by Allen and Humphreys “Immunisation of guinea pigs and cattle against ticks” Nature, 1979, 280: 491–493 led to a surge in the development of tick vaccines as a nonchemical method for prevention of tick infestations in susceptible hosts living in tick-endemic regions. Although observations of host resistance to ticks had been documented since the beginning of the last century, it was not until publication of this paper that the proof of concept of anti-tick vaccines was developed. The described experimental methods directly impacted further investigations on the discovery and evaluation of new anti-tick vaccines.
1. Introduction
Tick immunity or host resistance to ticks was known prior to the classic paper by Allen and Humphreys [1]. In 1918, Johnston and Bancroft documented that cattle became resistant to subsequent infestations by the Australian cattle tick Rhipicephalus (Boophilus) australis Fuller, 1899 (Acari, Ixodidae), suggesting an immune response to the substances inoculated during tick feeding [2]. In 1936, Ross observed that dogs living in endemic regions of Ixodes holocyclus Neumann, 1899 (Ixodida, Ixodidae) rarely suffered from tick paralysis and hypothesized that this may be due to immunity acquired after several tick bites [3]. Then, in 1939, Trager reported that Dermacentor andersoni Stiles, 1908 (Ixodida, Ixodidae) larvae fed on naive guinea pigs achieved an engorgement of 80%, but subsequent infestations only achieved 10% of engorgement [4]. The same year, this author published the results obtained from a series of experiments performed on D. variabilis Say, 1821 (Ixodida, Ixodidae) fed on laboratory animals [5]. Based upon these results, it was concluded that (a) one infestation of D. variabilis in guinea pigs and rabbits induced immunity preventing larval engorgement; (b) two or three infestations were needed for mice to become resistant to tick larvae; (c) repeated infestations of guinea pigs reduced the consumption of blood by ticks; and (d) immunity to ticks could be induced by subcutaneous inoculation of larval extracts or transferred by intraperitoneal inoculation of the hyperimmune serum obtained from guinea pigs after several tick infestations. This was the first time that immunization against tick infestation was demonstrated. In 1979, the publication by Allen and Humphreys [1] confirmed these results in laboratory animals, extended the concept for use in cattle, and demonstrated that immunological protection against ticks was feasible in production animals. This review aims to emphasize the contributions and the impact of this paper on anti-tick vaccine research more than four decades after its publication.
2. Discovery
Before the classic paper by Allen and Humphreys (1979) [1], two papers on the observations and findings in D. andersoni fed on laboratory animals were published [6,7]. In the first publication [6], Allen showed that resistance to D. andersoni larval infestations was acquired, as demonstrated by the obtention of fewer engorged larvae after feeding on guinea pigs infested for a second time. The role of the immune system was supported by further experiments, which showed that this resistance was prevented by using methotrexate, an immunosuppressant. In a second publication [7], Wikel and Allen documented the passive transfer of tick resistance by showing that the weight of D. andersoni larvae engorged on hosts inoculated with immune serum was slightly lower in the controls, suggesting that humoral factors contributed to host resistance.
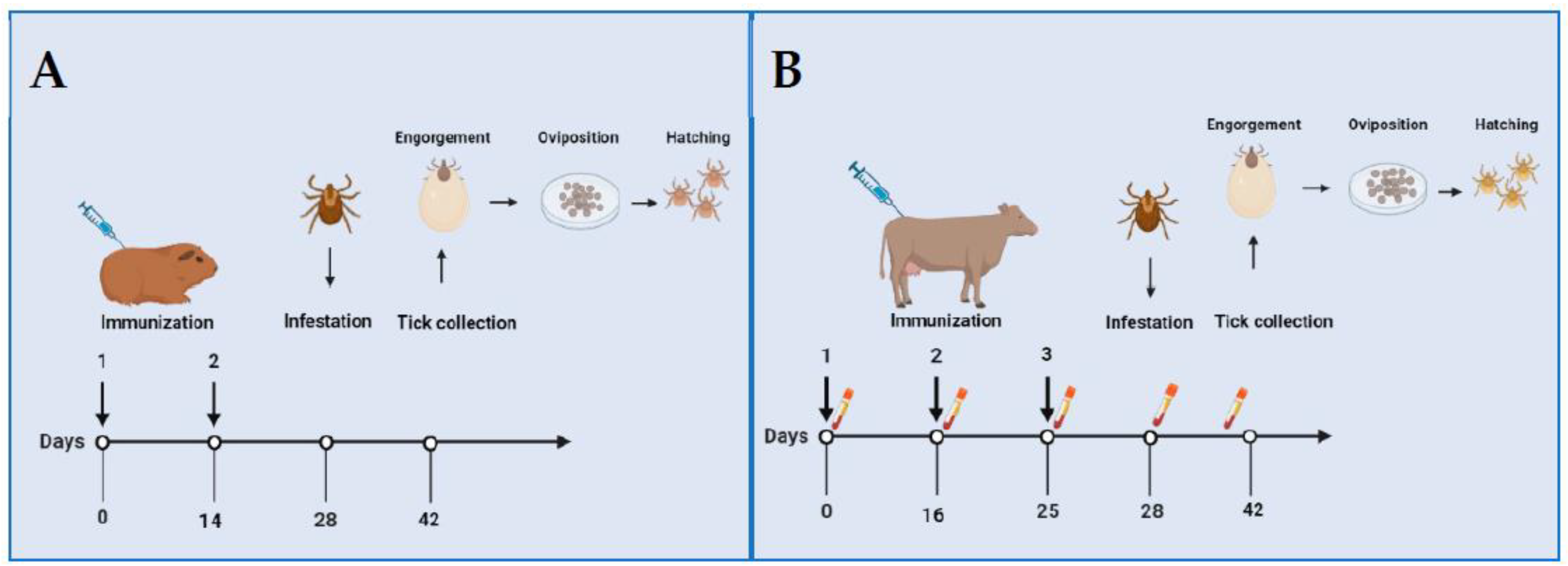
The paper by Allen and Humphreys [1] described, for the first time, experimental trials of immunization and tick challenge in two separate experiments using laboratory and large animals. The first experiment is represented in Figure 1A. Three groups of guinea pigs were intradermally immunized on days 0 and 14 with 1.2 mg/kg of D. andersoni crude extracts emulsified in Freund’s Complete Adjuvant (FCA) or FCA alone. The first group received an extract of gut and reproductive organs (antigen I), the second received an extract of all internal organs (antigen II), and the control group (C) received FCA alone. Each animal was infested with four couples of D. andersoni ticks on day 28, and the ticks were allowed to feed for two weeks, when engorgement was evaluated by the weight of each tick. Females were incubated in humidity chambers; and oviposition was evaluated by the weight of egg masses and by counting the number of larvae hatched at 35 days of incubation. These results indicated that antigen I significantly reduced mean egg weight, antigen II blocked oviposition, and both antigens blocked larval hatching. The second experiment is represented in Figure 1B. Two groups of four Hereford-cross calves were used. Calves from group 1 were intramuscularly immunized with 67 mg of antigen I emulsified in Freund’s Incomplete Adjuvant (FIA) on days 0 and 16, followed by a similar dose of antigen without adjuvant on day 25. Calves in group 2 (control) received FIA alone. Each calf was infested with 30 males and 100 females of D. andersoni on day 28. The ticks fed on calves from group 1 had a significant reduction in weight, oviposition, and hatching. Immunodiffusion studies on the sera obtained from immunized calves at 0, 16, 25, 28, and 38 days revealed single precipitating bands with the antigen and strong multiple bands on day 38, supporting the hypothesis of immune system activation.
3. Impact on Anti-Tick Vaccine Research
Allen and Humphreys [1] hypothesized that immunization against the one-host tick Rhipicephalus (Boophilus) microplus would be more effective than in three-host ticks because engorgement of the three developmental stages occurs on the same immunized animals, thus increasing the chance of antibody uptake with blood meals resulting in host protection. This, together with previous observations by Johnston and Bancroft [2], encouraged an investigation by Australian researchers working at the Commonwealth Scientific and Industrial Research Organization (CSIRO). These studies led to the discovery of the first antigen to protect cattle against tick infestations, of which the experimental details were documented in a series of scientific papers published in 1986 [8,9,10]. Briefly, immunization of cattle with purified crude extracts from R. microplus females induced protection against infestation with R. microplus larvae, resulting in 70% and 90% reductions in the adult tick population in two experimental trials [8]. Histological studies of ticks collected from vaccinated cattle showed a gut distention in ticks with blood leakage into hemolymph. Since the tick gut cells were ruptured, it was suggested that the protective antigen was located on the plasma membranes of these cells [9]. Affected ticks showed red coloration, up to 60% of females and males presented with damaged guts, and affected females failed to engorge or lay eggs [9]. The damage to affected ticks was hypothesized to be due to an antibody response directed to a “concealed tick antigen”, so called because its location was out of reach of the host immune response [10]. This concealed antigen was named Bm86, because of the tick species and the year it was identified; is an 89 kDa membrane-bound extracellular glycoprotein, located on the surface of the R. microplus tick’s gut cells [10,11]. The recombinant protein rBm86, expressed in Escherichia coli, was used in the formulation of the first vaccine produced against ticks [12]. The production and commercialization of Bm86 under the name TickGARD started in 1994, in Australia [13]. The same year, the Bm86 antigen was isolated and expressed in the yeast Pichia pastoris by researchers at the Center for Biotechnology and Genetic Engineering, in Cuba [14]. This recombinant antigen was commercialized in Latin American countries under the name Gavac [15]. The cost and the need for a vaccine dose every three months made anti-tick vaccines unpopular with Australian farmers. Thus, TickGARD was removed from the market in the late 1990s, reintroduced in 2002, and, finally, removed again a few years later because sales represented only 4% of acaricide sales [16]. Currently, Gavac is the only tick vaccine available. More than 3 million cattle have been vaccinated with Bm86, since its introduction on the market [17].
The publication by Allen and Humphreys [1] has greatly impacted the scientific research on anti-tick vaccines. In the two decades following its publication, most of the scientific articles published on tick vaccines reported on molecular characterization, elucidation of the role of Bm86, and its efficacy on tick control [11,12,13,14]. Investigation into the immune response in laboratory and natural hosts infested with tick species other than R. microplus has also expanded, as noted by the increase in scientific publications. A search on PubMed (https://pubmed.ncbi.nlm.nih.gov, accessed on 22 September 2022), using the keywords “ticks” and “anti-tick vaccines” during the period of 1979 to 2022, yielded 235 scientific papers. From these, reviews were eliminated, and only 195 papers were considered. From these selected papers, 33% corresponded to R. microplus, followed by I. scapularis, Haemaphysalis longicornis, Amblyomma americanum, and I. ricinus with 24%, 21%, and 2%, respectively.
The discovery and use of Bm86 as an anti-tick vaccine demonstrated that immunological control of ticks was a cost-effective, environmentally friendly, and harmless strategy in comparison with chemical control [18]. Therefore, efforts from different research groups focused on the identification of new protective antigens for vaccine development against not only R. microplus but also other ticks of medical and veterinary importance such as Ixodes spp., H. longicornis, and A. americanum. The most relevant antigens discovered after the publication of the paper by Allen and Humphreys [1] are grouped in Table 1. Variations in the efficacy of Bm86 in vaccinated cattle from different geographical regions have been observed [19,20,21]. The low or null efficacy in vaccination with Bm86 in South America encouraged the isolation and evaluation of Bm86 homologues. Bm95, obtained from an Argentinian R. microplus strain, showed protection against South American tick strains, suggesting that a broad-spectrum vaccine could be obtained by the inclusion of antigens from different geographical tick strains [21]. Bm86 orthologues were also investigated, and their efficacy against R. microplus and R. annulatus was evaluated. Interestingly, more efficacious results were obtained with these orthologues (Ba86 and Bm86) against R. annulatus than against R. microplus [22]. In another study, the immunization of cattle with Haa86 reduced the infestation of Hyalomma anatolicum larvae to 67% [23].
Other identified tick antigens in R. microplus include aquaporins and metalloproteases. Aquaporins are channels that regulate transportation across cell membranes. Immunization of cattle with a recombinant aquaporin (RmAQP1) showed 75% and 68% efficacy against R. microplus in two pen trials [24]. Metalloproteases are multifunctional proteins involved in blood-meal-related functions. The vaccination of cattle with a recombinant metalloprotease (BrRm-MP4) was tested against R. microplus, with an overall protection of 60% [25].
Investigation into other potential protective antigens conducted in R. appendiculatus resulted in the identification of 64P, a putative cement protein involved in attachment and feeding [26]. Cross reactivity of 64P with R. sanguineus, I. ricinus, A. variegatum, and R. microplus was shown. Evaluation of this protein as a vaccine preparation against I. ricinus in rabbits resulted in 18–64% and 5–60% mortality of nymphs and adults, respectively [27]. In addition, 64P was shown to react against exposed antigens and protected mice from transmission of tick-borne encephalitis virus (TBEV) by I. ricinus, suggesting the feasibility of using anti-tick vaccines to protect against tick-borne pathogens [28].

{kind=link}
Table 1.
Relevant identified tick vaccine antigens evaluated in immunization and tick challenge trials, from 1979 to date.
Table 1.
Relevant identified tick vaccine antigens evaluated in immunization and tick challenge trials, from 1979 to date.
| Tick Species | Antigen | Efficacy | References |
|---|---|---|---|
| Dermacentor andersoni | Internal organ tick extracts | Reduction on tick weight, oviposition, and hatching of D. andersoni fed on guinea pigs and cattle | [1] |
| Rhipicephalus (Boophilus) microplus | Bm86 | Reduction of 70–90% of R. microplus tick population in cattle | [8,9,10,12,14] |
| Bm95 | Reduction of 58% and 89% of R. microplus Argentinian and Camcord strains, respectively | [21] | |
| Aquaporins | 75% and 68% efficacy against R. microplus | [24] | |
| Metalloprotease, rBrRm-MP4 | Overall efficacy of 60% against R. microplus | [25] | |
| R. (B.) annulatus | Ba86 orthologue | Efficacy of 83% and 71.5% against R. annulatus and R. microplus, respectively | [22] |
| Hyalomma anatolicum anatolicum | Bm86 orthologues | Reduction of 69.7% of H. anatolicum larvae | [23] |
| R. appendiculatus | Glutathione S-transferase, GST | Efficacy of 67% against R. appendiculatus. No efficacy against R. sanguineus Efficacy of 43.69% against D. marginatus fed on rabbits | [29,30,31] |
| Attachment cement protein, 64P | 18–64% mortality of nymphs and 5–60% mortality of adults fed on immunized rabbits Protects mice against TBEV infection | [26,27,28] | |
| Haemaphysalis longicornis | P0, ribosomal peptide | Overall efficacy of 90% against R. sanguineus fed on rabbits and 96% against R. microplus in cattle | [32,33,34] |
| Ixodes scapularis | Subolesin | Efficacy of 71%, 63%, and 58% against I. scapularis in mice, rabbits, and sheep, respectively Efficacy of 51% and 60% against R. microplus and R. annulatus, respectively | [35,36,37] |
| Salp15 | 50% protection of mice from Borrelia burgdorferi infection | [38,39] | |
| I. ricinus | Ferritin | Efficacy of 64% and 72% against R. microplus and R. annulatus, respectively | [40,41] |
| Microbiome Enterobacteriaceae | Significant mortality of I. ricinus nymphs fed on mice | [42,43] |
Subolesin is a tick-vaccine candidate identified in a cDNA library from I. scapularis embryos that is involved in tick feeding and fertility [35]. Other studies concluded that subolesin is involved in gene expression and several cellular pathways [44,45]. When I. scapularis larvae, nymphs, and adults were fed on subolesin-immunized mice, rabbits, and sheep, efficacies of 71%, 63%, and 58%, respectively, were obtained [36]. In subolesin-immunized cattle, efficacies of 51% and 60% against R. microplus and R. annulatus, respectively, were obtained [37]; and the efficacy of vaccination against R. microplus increased up to 67% in cattle immunized with a subolesin-derived peptide [46].
Glutathione S-transferases (GSTs) are enzymes with a function in the detoxification of xenobiotic compounds such as drugs and pesticides, and an increase in the activity of GSTs is associated with resistance to synthetic pyrethroids [29]. GSTs are required for heme processing and detoxification during tick blood meals [29]. When a recombinant GST was evaluated as a vaccine against R. appendiculatus in rabbits, a protection level of about 65% was obtained, but no protection against R. sanguineus was demonstrated [30]. In another experiment, vaccination of rabbits with a GST and challenged with D. marginatus, resulted in an overall efficacy of 43.69% [31].
Ferritin (FER2), discovered in I. ricinus, is a protein with a role that is essential in iron metabolism and was shown to affect tick feeding [40]. FER2 has been evaluated in vaccination trials against R. microplus and R. annulatus in cattle, with efficacies of 64% and 72%, respectively [40]. However, when used for vaccination against I. ricinus in cattle, significant results were not obtained [41].
Functional studies on H. longicornis resulted in the identification of Protein 0 (P0), a ribosomal protein with a principal role of the regulation of transcription, which is required for blood ingestion and tick viability [32]. A 20 aa peptide from the P0 protein was evaluated as an anti-tick vaccine against R. sanguineus in rabbits, with an efficacy of 90% [33]. Subsequent studies showed 85% efficacy against R. sanguineus and R. microplus in dogs and cattle, respectively [34].
Salp15 is a 15 kDa protein from tick saliva that was identified in I. scapularis, which was associated with immunosuppression by inhibiting cellular and complement activity in the tick-bite area [47]. Subsequent experiments with I. scapularis engorged on Borrelia burgdorferi-infected mice demonstrated that Salp15 binds OspC, an outer-surface protein produced by B. burgdorferi, which facilitates transmission during tick feeding [38]. Immunization of mice with Salp15 resulted in 50% protection against B. burgdorferi infection, suggesting that the vaccination of hosts with the target molecules required for pathogen transmission may be used for disease prevention [48]. Recently, an mRNA vaccine, encoding for 19 salivary proteins from I. scapularis (including Salp15), was tested in guinea pigs, and a reduction in tick feeding and B. burgdorferi infection were confirmed [39].
Tick microbiome manipulation through host vaccination is a novel approach that has been recently investigated using an I. ricinus–mouse model [42]. Keystone bacteria family (Enterobacteriaceae) in the microbiome of I. ricinus were targeted through vaccination and the evaluation of nymph-feeding performance on vaccinated mice. High mortality during nymph feeding suggested that anti-tick microbiota vaccines may be used to evaluate the function of a specific taxon in tick microbiome, thus facilitating the identification of new targets for tick interventions and blocking the transmission of tick-borne pathogens [43].
In conclusion, since the publication of Allen and Humphreys in 1979 [1], anti-tick vaccine research has greatly advanced. Two commercial vaccines against the cattle tick R. microplus have been independently produced, and many new molecular targets from different tick species have been identified. Although anti-tick vaccine research has expanded to different tick species, R. microplus remains the most studied, as effective anti-tick vaccines against this tick would provide the most significant contribution to a reduction in the chemical products used to treat cattle raised in tick-endemic regions. Vaccines for the reduction in tick populations and blocking the transmission of pathogens causing human diseases are also needed. Developing anti-tick vaccines is challenging, but considering the experience with Bm86 vaccines, the availability of genomics and transcriptomics resources, and the newly reported tools for manipulation of the tick microbiome, effective anti-tick vaccines for targeting both ticks and tick-vectored pathogens are expected.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thanks Alejandro Cabezas-Cruz for the invitation to write this contribution to the Special Issue “10th Anniversary of Pathogens–Classic Papers in Tick Research”. Jeff Blair is acknowledged for reviewing this manuscript and providing valuable suggestions.
Conflicts of Interest
The author declares no conflict of interest.
References
- Allen, J.R.; Humphreys, S.J. Immunisation of guinea pigs and cattle against ticks. Nature 1979, 280, 491–493. [Google Scholar] [CrossRef]
- Johnston, T.H.; Bancroft, M.J. A tick resistant condition in cattle. Proc. R Soc. Qld. 1918, 30, 219–317. [Google Scholar]
- Ross, I.C. An experimental study of tick paralysis in Australia. Parasitology 1926, 18, 410–429. [Google Scholar] [CrossRef]
- Trager, W. Acquired immunity to ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Trager, W. Further observations on acquired immunity to the tick Dermacentor variabilis Say. J. Parasitol. 1939, 25, 137–139. [Google Scholar] [CrossRef]
- Allen, J.R. Tick resistance: Basophils in skin reactions of resistant guinea pigs. Int. J. Parasitol. 1973, 3, 195–200. [Google Scholar] [CrossRef]
- Wikel, S.K.; Allen, J.R. Acquired resistance to ticks. I. Passive transfer of resistance. Immunology 1976, 30, 311. [Google Scholar] [PubMed]
- Johnston, L.A.Y.; Kemp, D.H.; Pearson, R.D. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Effects of induced immunity on tick populations. Int. J. Parasitol. 1986, 16, 27–34. [Google Scholar] [CrossRef]
- Agbede, R.I.S.; Kemp, D.H. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Histopathology of ticks feeding on vaccinated cattle. Int. J. Parasitol. 1986, 16, 35–41. [Google Scholar] [CrossRef]
- Kemp, D.H.; Agbede, R.I.; Johnston, L.A.; Gough, J.M. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Feeding and survival of the parasite on vaccinated cattle. Int. J. Parasitol. 1986, 16, 115–120. [Google Scholar] [CrossRef]
- Willadsen, P.; Kemp, D.H. Vaccination with ‘concealed’ antigens for tick control. Parasitol. Today 1988, 4, 196–198. [Google Scholar] [CrossRef]
- Rand, K.N.; Moore, T.; Sriskantha, A.; Spring, K.; Tellam, R.; Willadsen, P.; Cobon, G.S. Cloning and expression of a protective antigen from the cattle tick Boophilus Microplus. Proc. Natl. Acad. Sci. USA 1989, 86, 9657–9661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willadsen, P.; Bird, P.; Cobon, G.S.; Hungerford, J. Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 1995, 110, S43–S50. [Google Scholar] [CrossRef]
- Rodríguez, M.; Rubiera, R.; Penichet, M.; Montesinos, R.; Cremata, J.; Falcón, V.; Sánchez, G.; Bringas, R.; Cordovés, C.; Mario Valdés, M.; et al. High level expression of the B. microplus Bm86 antigen in the yeast Pichia pastoris forming highly immunogenic particles for cattle. J. Biotechnol. 1994, 33, 135–146. [Google Scholar] [CrossRef]
- Vanegas, L.F.; Parra, S.A.; Vanegas, C.G.; de la Fuente, J. Commercialization of the recombinant vaccine Gavac against Boophilus microplus in Colombia. In Recombinant Vaccines for the Control of Cattle Tick; De la Fuente, J., Ed.; Elfos Scientiae: La Habana, Cuba, 1995; pp. 195–199. [Google Scholar]
- Playford, M. Review of Research Needs for Cattle Tick Control—Phases I and II; Meat & Livestock: Sydney, Australia, 2005. [Google Scholar]
- Rodríguez-Mallón, A.; Javier González, L.; Encinosa Guzmán, P.E.; Bechara, G.H.; Sanches, G.S.; Pousa, S.; Estrada, M.P. Functional and mass spectrometric evaluation of an anti-tick antigen based on the P0 peptide conjugated to Bm86 protein. Pathogens 2020, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Rodríguez, M.; Redondo, M.; Montero, C.; García-García, J.C.; Méndez, L.; Serrano, E.; Valdés, M.; Enriquez, A.; Canales, M.; et al. Field studies and cost-effectiveness analysis of vaccination with Gavac against the cattle tick Boophilus microplus. Vaccine 1998, 16, 366–373. [Google Scholar] [CrossRef]
- Willadsen, P.; Eisemann, C.H.; Tellam, R.L. Concealed antigens: Expanding the range of immunological targets. Parasitol. Today 1993, 9, 132–135. [Google Scholar] [CrossRef]
- Riding, G.A.; Jarmey, J.; McKenna, R.V.; Pearson, R.; Cobon, G.S.; Willadsen, P. A protective “concealed” antigen from Boophilus microplus. Purification, localization, and possible function. J. Immunol. 1994, 153, 5158–5166. [Google Scholar]
- García-García, J.C.; Montero, C.; Redondo, M.; Vargas, M.; Canales, M.; Boue, O.; Rodríguez, M.; Joglar, M.; Machado, H.; González, I.L.; et al. Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine 2000, 18, 2275–2287. [Google Scholar] [CrossRef]
- Canales, M.; Almazán, C.; Naranjo, V.; Jongejan, F.; de la Fuente, J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol. 2009, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; de La Fuente, J.; Kumar, R.; Ghosh, S. Comparative efficacy of rHaa86 and rBm86 against Hyalomma anatolicum anatolicum and Rhipicephalus (Boophilus) microplus. Parasit. Immunol. 2012, 34, 297–301. [Google Scholar]
- Guerrero, F.D.; Andreotti, R.; Bendele, K.G.; Cunha, R.C.; Miller, R.J.; Yeater, K.; Pérez de León, A.A. Rhipicephalus (Boophilus) microplus aquaporin as an effective vaccine antigen to protect against cattle tick infestations. Parasit. Vectors 2014, 7, 475–487. [Google Scholar] [PubMed] [Green Version]
- Ali, A.; Parizi, L.F.; Guizzo, M.G.; Tirloni, L.; Seixas, A.; Vaz Ida, S., Jr.; Termignoni, C. Immunoprotective potential of a Rhipicephalus (Boophilus) microplus metalloprotease. Vet. Parasitol. 2015, 207, 107–114. [Google Scholar]
- Trimnell, A.R.; Hails, R.S.; Nuttall, P.A. Dual action ectoparasite vaccine targeting ‘exposed’and ‘concealed’antigens. Vaccine 2020, 29, 3560–3568. [Google Scholar]
- Trimnell, A.R.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P.A. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 2005, 23, 329–4341. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Trimnell, A.R.; Licková, M.; Kazimírová, M.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P.A. An antivector vaccine protects against a lethal vector-borne pathogen. PLoS Pathog. 2006, 2, e27. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Vaz, J.I.; Imamura, S.; Ohashi, K.; Onuma, M. Cloning, expression and partial characterization of a Haemaphysalis longicornis and a Rhipicephalus appendiculatus glutathione S-transferase. Insect Mol. Biol. 2004, 13, 329–335. [Google Scholar] [CrossRef]
- Sabadin, G.A.; Parizi, L.F.; Kiio, I.; Xavier, M.A.; da Silva Matos, R.; Camargo-Mathias, M.I.; Githaka, N.W.; Nene, V.; da Silva Vaz, I.J. Effect of recombinant glutathione S-transferase as vaccine antigen against Rhipicephalus appendiculatus and Rhipicephalus sanguineus infestation. Vaccine 2017, 35, 6649–6656. [Google Scholar] [CrossRef]
- Huercha, S.R.; Li, M.; Fan, X.; Hu, Z.; Wu, L.; Li, Y.; Zhang, W.; Zhang, Y.; Ma, Y.; Bayin, C. Characterization of glutathione S-transferase of Dermacentor marginatus and effect of the recombinant antigen as a potential anti-tick vaccine. Vet. Parasitol. 2020, 279, 109043. [Google Scholar] [CrossRef]
- Gong, H.; Liao, M.; Zhou, J.; Hatta, T.; Huang, P.; Zhang, G.; Kanuka, H.; Nishikawa, Y.; Xuan, X.; Fujisaki, K. Gene silencing of ribosomal protein P0 is lethal to the tick Haemaphysalis longicornis. Vet. Parasitol. 2008, 151, 268–278. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A.; Fernández, E.; Encinosa, P.E.; Bello, Y.; Méndez-Pérez, L.; Cepero, L.; Pérez, D.; González, M.; Garay, H.; Reyes, O. A novel tick antigen shows high vaccine efficacy against the dog tick, Rhipicephalus sanguineus. Vaccine 2012, 30, 1782–1789. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A.; Encinosa, P.E.; Méndez-Pérez, L.; Bello, Y.; Rodríguez Fernández, R.; Garay, H.; Cabrales, A.; Méndez, L.; Borroto, C.; Estrada, M.P. High efficacy of a 20 amino acid peptide of the acidic ribosomal protein P0 against the cattle tick, Rhipicephalus microplus. Ticks Tick Borne Dis. 2015, 6, 530–537. [Google Scholar] [CrossRef]
- Almazán, C.; Kocan, K.; Bergman, D.; Garcia-Garcia, J.; Blouin, E.; de la Fuente, J. Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 2003, 21, 1492–1501. [Google Scholar] [CrossRef]
- Almazán, C.; Kocan, K.M.; Blouin, E.F.; de la Fuente, J. Vaccination with recombinant tick antigens for the control of Ixodes scapularis adult infestations. Vaccine 2005, 23, 5294–5298. [Google Scholar] [CrossRef]
- Almazán, C.; Lagunes, R.; Villar, M.; Canales, M.; Rosario-Cruz, R.; Jongejan, F.; de la Fuente, J. Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitol. Res. 2010, 106, 471–479. [Google Scholar] [CrossRef]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Fikrig, E. The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Sajid, A.; Matias, J.; Arora, G.; Kurokawa, C.; DePonte, K.; Tang, X.; Fikrig, E. mRNA vaccination induces tick resistance and prevents transmission of the Lyme disease agent. Sci. Transl. Med. 2021, 13, eabj9827. [Google Scholar] [CrossRef]
- Hajdusek, O.; Almazán, C.; Loosova, G.; Villar, M.; Canales, M.; Grubhoffer, L.; Kopacek, P.; de la Fuente, J. Characterization of ferritin 2 for the control of tick infestations. Vaccine 2010, 28, 2993–2998. [Google Scholar] [CrossRef]
- Knorr, S.; Anguita, J.; Cortazar, J.T.; Hajdusek, O.; Kopáček, P.; Trentelman, J.J.; Nijhof, A.M. Preliminary evaluation of tick protein extracts and recombinant ferritin 2 as anti-tick vaccines targeting Ixodes ricinus in cattle. Front. Physiol. 2018, 9, 1696. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Hernández, L.; Obregón, D.; Maye, J.; Borneres, J.; Versille, N.; de la Fuente, J.; Estrada-Peña, A.; Hodžic, A.; Šimo, L.; Cabezas-Cruz, A. Anti-tick microbiota vaccine impacts Ixodes ricinus performance during feeding. Vaccines 2020, 8, 702. [Google Scholar] [CrossRef]
- Mateos-Hernández, L.; Obregon, D.; Wu-Chuang, A.; Maye, J.; Bornères, J.; Versillé, N.; de la Fuente, J.; Díaz-Sánchez, S.; Bermúdez-Humarán, L.G.; Torres-Maravilla, E.; et al. Anti-microbiota vaccines modulate the tick microbiome in a taxon-specific manner. Front. Immunol. 2021, 12, 704621. [Google Scholar] [CrossRef] [PubMed]
- Galindo, R.C.; Doncel-Perez, E.; Zivkovic, Z.; Naranjo, V.; Gortazar, C.; Mangold, A.J.; de la Fuente, J. Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev. Comp. Immunol. 2009, 33, 612–617. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Maritz-Olivier, C.; Naranjo, V.; Ayoubi, P.; Nijhof, A.M.; Almazán, C.; Canales, M.; Pérez de la Lastra, J.M.; Galindo, R.C.; Blouin, E.F.; et al. Evidence of the role of tick subolesin in gene expression. BMC Genom. 2008, 9, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza-Martínez, N.; Alonso-Díaz, M.A.; Merino, O.; Fernández-Salas, A.; Lagunes-Quintanilla, R. Protective efficacy of the peptide Subolesin antigen against the cattle tick Rhipicephalus microplus under natural infestation. Vet. Parasitol. 2021, 299, 109577. [Google Scholar] [CrossRef]
- Anguita, J.; Ramamoorthi, N.; Hovius, J.W.; Das, S.; Thomas, V.; Persinski, R.; Conze, D.; Askenase, P.W.; Rincón, M.; Kantor, F.S.; et al. Salp15, an Ixodes scapularis salivary protein, inhibits CD4(+) T cell activation. Immunity 2002, 16, 849–859. [Google Scholar] [CrossRef]
- Dai, J.; Wang, P.; Adusumilli, S.; Booth, C.J.; Narasimhan, S.; Anguita, J.; Fikrig, E. Antibodies against a tick protein, Salp15, protect mice from the Lyme disease agent. Cell Host Microbe 2009, 6, 482–492. [Google Scholar] [CrossRef]
Figure 1.
Schematical representation of experiments of immunization against Dermacentor andersoni performed by Allen and Humphreys (1979). (A) Three groups of guinea pigs were immunized at days 0 and 14 with D. andersoni crude extracts from gut and reproductive organs and all internal organs. (B) Two groups of 4 Hereford-cross calves were immunized at days 0, 16, and 25 with crude extracts from gut and reproductive organs. Serum samples were obtained at days 0, 16, 25, 28, and 38 for immunodiffusion. Guinea pigs were infested with 4 couples and cattle with 30 males and 100 females of D. andersoni adult ticks at day 28. Engorged ticks were collected after two weeks of feeding. Efficacy of immunization was evaluated based on weight of engorged ticks, oviposition, and hatching. Created with Biorender.com.
Figure 1.
Schematical representation of experiments of immunization against Dermacentor andersoni performed by Allen and Humphreys (1979). (A) Three groups of guinea pigs were immunized at days 0 and 14 with D. andersoni crude extracts from gut and reproductive organs and all internal organs. (B) Two groups of 4 Hereford-cross calves were immunized at days 0, 16, and 25 with crude extracts from gut and reproductive organs. Serum samples were obtained at days 0, 16, 25, 28, and 38 for immunodiffusion. Guinea pigs were infested with 4 couples and cattle with 30 males and 100 females of D. andersoni adult ticks at day 28. Engorged ticks were collected after two weeks of feeding. Efficacy of immunization was evaluated based on weight of engorged ticks, oviposition, and hatching. Created with Biorender.com.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Almazán, C. Impact of the Paper by Allen and Humphreys (1979) on Anti-Tick Vaccine Research. Pathogens 2022, 11, 1253. https://doi.org/10.3390/pathogens11111253
AMA Style
Almazán C. Impact of the Paper by Allen and Humphreys (1979) on Anti-Tick Vaccine Research. Pathogens. 2022; 11(11):1253. https://doi.org/10.3390/pathogens11111253
Chicago/Turabian StyleAlmazán, Consuelo. 2022. "Impact of the Paper by Allen and Humphreys (1979) on Anti-Tick Vaccine Research" Pathogens 11, no. 11: 1253. https://doi.org/10.3390/pathogens11111253
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.