
Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
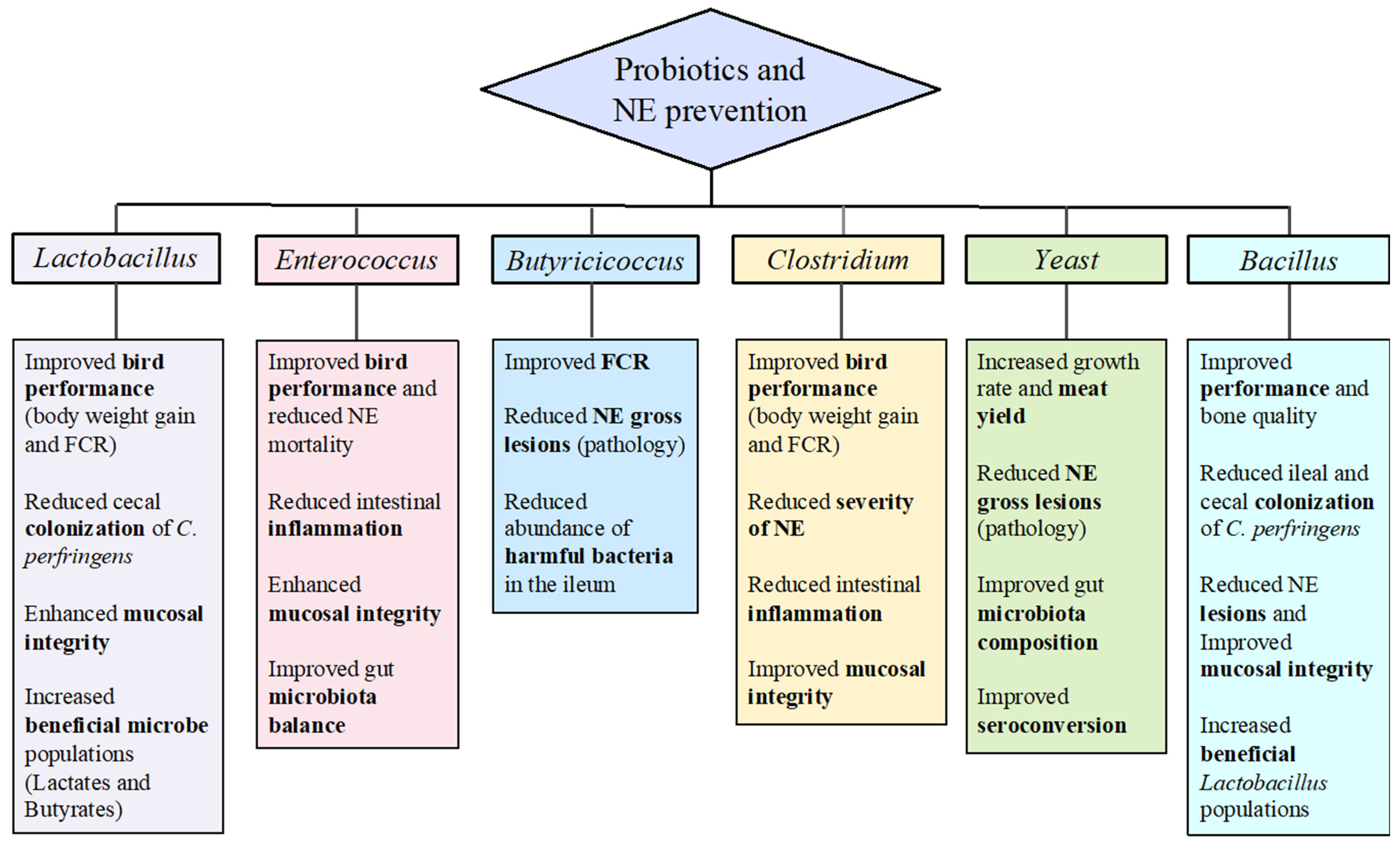
2. Lactobacillus Bacteria Used as Probiotics for Control of C. perfringens-Induced NE
3. Non-Lactobacillus Beneficial Microbes as Probiotics for NE Control
3.1. Clostridium butyricum
3.2. Enterococcus faecium
3.3. Butyricicoccus pullicaecorum
3.4. Bacillus Species
3.5. Yeasts
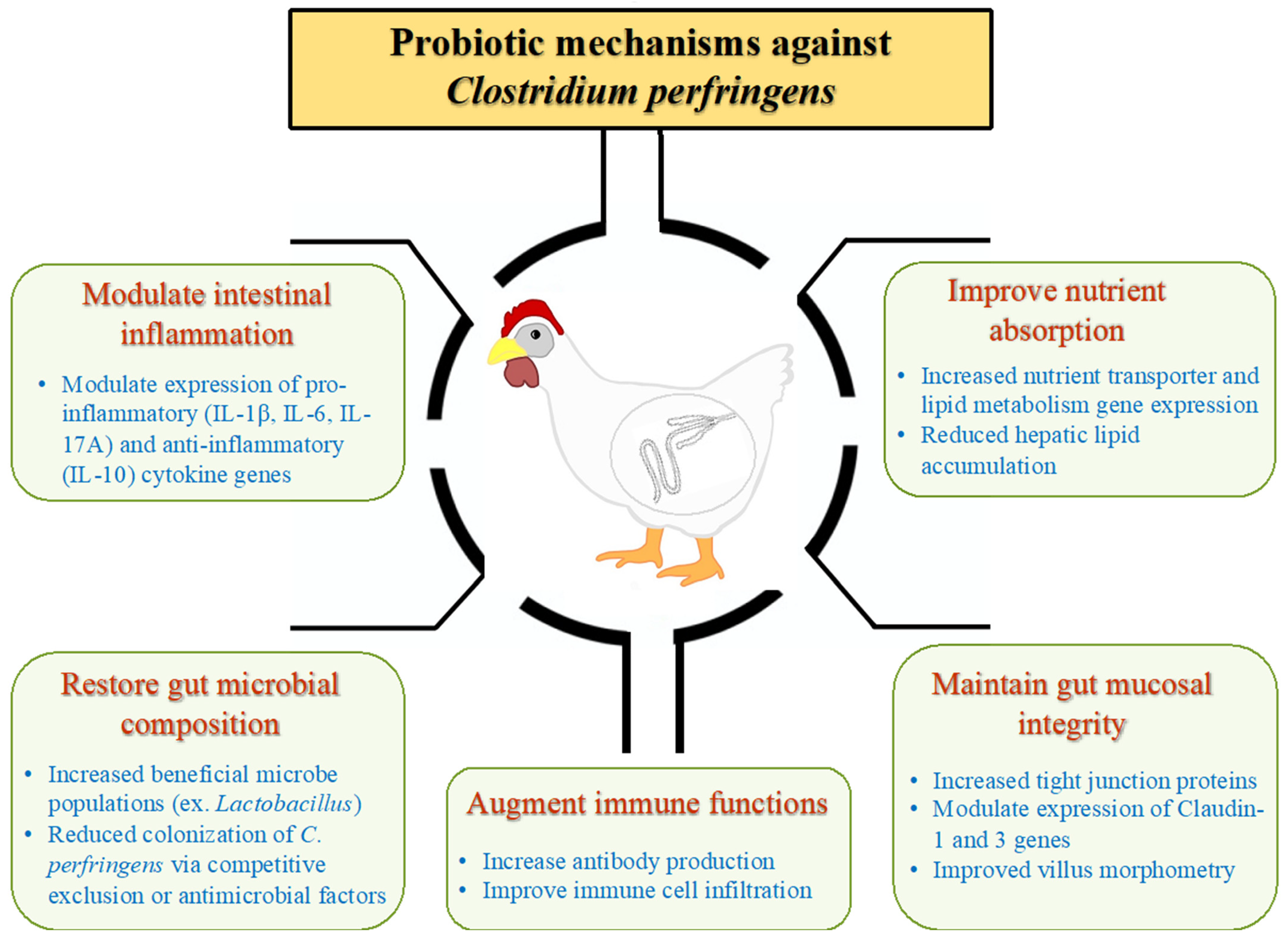
4. Probiotic Mechanisms of NE Control
4.1. Restoration of Gut Microbial Composition
4.2. Maintenance of Intestinal Epithelial Integrity, Mucus Production and Competitive Exclusion of Pathogens
4.3. Immune Modulation
4.4. Production of Antimicrobial Molecules
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Songer, J.G. Clostridial Enteric Diseases of Domestic Animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Wade, B.; Keyburn, A.L.; Seemann, T.; Rood, J.I.; Moore, R.J. Binding of Clostridium perfringens to Collagen Correlates with the Ability to Cause Necrotic Enteritis in Chickens. Vet. Microbiol. 2015, 180, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.F.; Parreira, V.R.; Gohari, M.I.; Lepp, D.; Gong, J. The Pathogenesis of Necrotic Enteritis in Chickens: What We Know and What We Need to Know: A Review. Avian Pathol. 2016, 45, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.B. Intercurrent Coccidiosis and Necrotic Enteritis of Chickens: Rational, Integrated Disease Management by Maintenance of Gut Integrity. Avian Pathol. 2005, 34, 159–180. [Google Scholar] [CrossRef]
- Gholamiandehkordi, A.R.; Timbermont, L.; Lanckriet, A.; van den Broeck, W.; Pedersen, K.; Dewulf, J.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Quantification of Gut Lesions in a Subclinical Necrotic Enteritis Model. Avian Pathol. 2007, 36, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Lovland, A.; Kaldhusdal, M. Severely Impaired Production Performance in Broiler Flocks with High Incidence of Clostridium Perfringens-Associated Hepatitis. Avian Pathol. 2001, 30, 73–81. [Google Scholar] [CrossRef]
- Skinner, J.T.; Bauer, S.; Young, V.; Pauling, G.; Wilson, J. An Economic Analysis of the Impact of Subclinical (Mild) Necrotic Enteritis in Broiler Chickens. Avian Dis. 2010, 54, 1237–1240. [Google Scholar] [CrossRef]
- Keerqin, C.; Wu, S.B.; Svihus, B.; Swick, R.; Morgan, N.; Choct, M. An Early Feeding Regime and a High-Density Amino Acid Diet on Growth Performance of Broilers under Subclinical Necrotic Enteritis Challenge. Anim. Nutr. 2017, 3, 25–32. [Google Scholar] [CrossRef]
- M’Sadeq, S.A.; Wu, S.; Swick, R.A.; Choct, M. Towards the Control of Necrotic Enteritis in Broiler Chickens with In-Feed Antibiotics Phasing-out Worldwide. Anim. Nutr. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Mack, D. Probiotics-Mixed Messages. Can. Fam. Physician 2005, 51, 1455–1457. [Google Scholar]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R.D. Alternatives to Antibiotics: Bacteriocins, Antimicrobial Peptides and Bacteriophages. Poult. Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal Microbiome of Poultry and Its Interaction with Host and Diet. Gut Microbes 2013, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Caly, D.L.; D’Inca, R.; Auclair, E.; Drider, D. Alternatives to Antibiotics to Prevent Necrotic Enteritis in Broiler Chickens: A Microbiologist’s Perspective. Front. Microbiol. 2015, 6, 1336. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on Gut Microbiota Composition in Broilers Challenged with Clostridium perfringens. PLoS ONE 2017, 12, e0188634. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on the Growth Performance and Intestinal Health of Broilers Challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, H.R.; Gong, J.; Gyles, C.L.; Hayes, M.A.; Zhou, H.; Sanei, B.; Chambers, J.R.; Sharif, S. Probiotics Stimulate Production of Natural Antibodies in Chickens. Clin. Vaccine Immunol. 2006, 13, 975–980. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Liu, D.; Zhang, B.; Li, Z.; Li, Y.; Ding, B.; Guo, Y. Two Lactobacillus Species Inhibit the Growth and α-Toxin Production of Clostridium perfringens and Induced Proinflammatory Factors in Chicken Intestinal Epithelial Cells in Vitro. Front. Microbiol. 2017, 8, 2081. [Google Scholar] [CrossRef]
- Rahimi, S.; Kathariou, S.; Grimes, J.L.; Siletzky, R.M. Effect of Direct-Fed Microbials on Performance and Clostridium perfringens Colonization of Turkey Poults. Poult. Sci. 2011, 90, 2656–2662. [Google Scholar] [CrossRef]
- Emami, N.K.; Calik, A.; White, M.B.; Young, M.; Dalloul, R.A. Necrotic Enteritis in Broiler Chickens: The Role of Tight Junctions and Mucosal Immune Responses in Alleviating the Effect of the Disease. Microorganisms 2019, 7, 231. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Li, C.; Yang, X.J.; Li, Z.J.; Sun, F.F.; Wu, X.H.; Yao, J.H. Reduced Lesions in Chickens with Clostridium perfringens-Induced Necrotic Enteritis by Lactobacillus fermentum 1.20291. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef]
- Allaart, J.G.; van Asten, A.J.A.M.; Vernooij, J.C.M.; Gröne, A. Effect of Lactobacillus fermentum on Beta2 Toxin Production by Clostridium perfringens. Appl. Environ. Microbiol. 2011, 77, 4406–4411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.Y.; Chou, C.H.; Wang, C. Characterization of Toxin Genes and Quantitative Analysis of NetB in Necrotic Enteritis (NE)-Producing and Non-NE-Producing Clostridium perfringens Isolated from Chickens. Anaerobe 2018, 54, 115–120. [Google Scholar] [CrossRef]
- Jost, B.H.; Billington, S.J.; Trinh, H.T.; Bueschel, D.M.; Songer, J.G. Atypical Cpb2 Genes, Encoding Beta2-Toxin in Clostridium perfringens Isolates of Nonporcine Origin. Infect. Immun. 2005, 73, 652–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geier, M.S.; Mikkelsen, L.L.; Torok, V.A.; Allison, G.E.; Olnood, C.G.; Boulianne, M.; Hughes, R.J.; Choct, M. Comparison of Alternatives to In-Feed Antimicrobials for the Prevention of Clinical Necrotic Enteritis. J. Appl. Microbiol. 2010, 109, 1329–1338. [Google Scholar] [CrossRef]
- la Ragione, R.M.; Narbad, A.; Gasson, M.J.; Woodward, M.J. In Vivo Characterization of Lactobacillus johnsonii FI9785 for Use as a Defined Competitive Exclusion Agent against Bacterial Pathogens in Poultry. Lett. Appl. Microbiol. 2004, 38, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Lai, J.; Khalique, A.; Li, G.; Pan, K.; Jing, B.; Zeng, D. Probiotic Enhanced Intestinal Immunity in Broilers against Subclinical Necrotic Enteritis. Front. Immunol. 2017, 8, 1592. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B.; et al. Probiotic Lactobacillus johnsonii BS15 Improves Blood Parameters Related to Immunity in Broilers Experimentally Infected with Subclinical Necrotic Enteritis. Front. Microbiol. 2018, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Khalique, A.; Zeng, D.; Wang, H.; Qing, X.; Zhou, Y.; Xin, J.; Zeng, Y.; Pan, K.; Shu, G.; Jing, B.; et al. Transcriptome Analysis Revealed Ameliorative Effect of Probiotic Lactobacillus johnsonii BS15 against Subclinical Necrotic Enteritis Induced Hepatic Inflammation in Broilers. Microb. Pathog. 2019, 132, 201–207. [Google Scholar] [CrossRef]
- Qing, X.; Zeng, D.; Wang, H.; Ni, X.; Liu, L.; Lai, J.; Khalique, A.; Pan, K.; Jing, B. Preventing Subclinical Necrotic Enteritis through Lactobacillus johnsonii BS15 by Ameliorating Lipid Metabolism and Intestinal Microflora in Broiler Chickens. AMB Express 2017, 7, 139. [Google Scholar] [CrossRef]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. BioMed Res. Int. 2018, 2018, 9361614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Chen, Y.; Yu, L.; Wang, J.; Huang, M.; Zhu, N. Effects of Lactobacillus plantarum on Intestinal Integrity and Immune Responses of Egg-Laying Chickens Infected with Clostridium perfringens under the Free-Range or the Specific Pathogen Free Environment. BMC Vet. Res. 2020, 16, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuprys-Caruk, M.; Michalczuk, M.; Chablowska, B.; Stefanska, I.; Kotyrba, D.; Parzeniecka-Jaworska, M. Efficacy and Safety Assessment of Microbiological Feed Additive for Chicken Broilers in Tolerance Studies. J. Vet. Res. 2018, 62, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizerwetter-Świda, M.; Binek, M. Assessment of Potentially Probiotic Properties of Lactobacillus Strains Isolated from Chickens. Pol. J. Vet. Sci. 2016, 19, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Dec, M.; Puchalski, A.; Nowaczek, A.; Wernicki, A. Antimicrobial Activity of Lactobacillus Strains of Chicken Origin against Bacterial Pathogens. Int. Microbiol. 2016, 19, 57–67. [Google Scholar] [CrossRef]
- Monteiro, C.R.A.V.; do Carmo, M.S.; Melo, B.O.; Alves, M.S.; dos Santos, C.I.; Monteiro, S.G.; Bomfim, M.R.Q.; Fernandes, E.S.; Monteiro-Neto, V. In Vitro Antimicrobial Activity and Probiotic Potential of Bifidobacterium and Lactobacillus against Species of Clostridium. Nutrients 2019, 11, 448. [Google Scholar] [CrossRef] [Green Version]
- Schoster, A.; Kokotovic, B.; Permin, A.; Pedersen, P.D.; Bello, F.D.; Guardabassi, L. In Vitro Inhibition of Clostridium difficile and Clostridium perfringens by Commercial Probiotic Strains. Anaerobe 2013, 20, 36–41. [Google Scholar] [CrossRef]
- Kizerwetter-Świda, M.; Binek, M. Selection of Potentially Probiotic Lactobacillus Strains towards Their Inhibitory Activity against Poultry Enteropathogenic Bacteria. Pol. J. Microbiol. 2005, 54, 287–294. [Google Scholar]
- Kizerwetter-Swida, M.; Binek, M. Protective Effect of Potentially Probiotic Lactobacillus Strain on Infection with Pathogenic Bacteria in Chickens. Pol. J. Vet. Sci. 2009, 12, 15–20. [Google Scholar]
- Brisbin, J.T.; Gong, J.; Orouji, S.; Esufali, J.; Mallick, A.I.; Parvizi, P.; Shewen, P.E.; Sharif, S. Oral Treatment of Chickens with Lactobacilli Influences Elicitation of Immune Responses. Clin. Vaccine Immunol. 2011, 18, 1447–1455. [Google Scholar] [CrossRef]
- Koenen, M.E.; Kramer, J.; van der Hulst, R.; Heres, L.; Jeurissen, S.H.M.; Boersma, W.J.A. Immunomodulation by Probiotic Lactobacilli in Layer- And Meat-Type Chickens. Br. Poult. Sci. 2004, 45, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Karimi, S.H.; Bavananthasivam, J.; Kulkarni, R.R.; Sharif, S. Effects of in Ovo Inoculation of Multi-Strain Lactobacilli on Cytokine Gene Expression and Antibody-Mediated Immune Responses in Chickens. Front. Vet. Sci. 2020, 7, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layton, S.L.; Hernandez-Velasco, X.; Chaitanya, S.; Xavier, J.; Menconi, A.; Latorre, J.D.; Kallapura, G.; Kuttappan, V.A.; Wolfenden, R.E.; Filho, R.L.A.; et al. The Effect of a Lactobacillus-Based Probiotic for the Control of Necrotic Enteritis in Broilers. Food Nutr. Sci. 2013, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- de la Mora, Z.V.; Nuño, K.; Vázquez-Paulino, O.; Avalos, H.; Castro-Rosas, J.; Gómez-Aldapa, C.; Angulo, C.; Ascencio, F.; Villarruel-López, A. Effect of a Synbiotic Mix on Intestinal Structural Changes, and Salmonella Typhimurium and Clostridium perfringens Colonization in Broiler Chickens. Animals 2019, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Shanmugasundaram, R.; Markazi, A.; Mortada, M.; Ng, T.T.; Applegate, T.J.; Bielke, L.R.; Syed, B.; Pender, C.M.; Curry, S.; Murugesan, G.R.; et al. Research Note: Effect of Synbiotic Supplementation on Caecal Clostridium perfringens Load in Broiler Chickens with Different Necrotic Enteritis Challenge Models. Poult. Sci. 2020, 99, 2452–2458. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, J.; Waneck, C.; Byrd, J.; Genovese, K.; Duke, S.; Nisbet, D. Efficacy of Multistrain Direct-Fed Microbial and Phytogenetic Products in Reducing Necrotic Enteritis in Commercial Broilers. Poult. Sci. 2009, 88, 2075–2080. [Google Scholar] [CrossRef]
- Shojadoost, B.; Alizadeh, M.; Boodhoo, N.; Astill, J.; Karimi, S.H.; Doost, J.S.; Taha-Abdelaziz, K.; Kulkarni, R.; Sharif, S. Effects of Treatment with Lactobacilli on Necrotic Enteritis in Broiler Chickens. Probiotics Antimicrob. Proteins 2022. [Google Scholar] [CrossRef]
- Woo, T.D.H.; Oka, K.; Takahashi, M.; Hojo, F.; Osaki, A.; Hanawa, T.; Kurata, S.; Yonezawa, H.; Kamiya, S. Inhibition of the Cytotoxic Effect of Clostridium difficile in Vitro by Clostridium butyricum MIYAIRI 588 Strain. J. Med. Microbiol. 2011, 60, 1617–1625. [Google Scholar] [CrossRef]
- Huang, T.; Peng, X.Y.; Gao, B.; Wei, Q.L.; Xiang, R.; Yuan, M.G.; Xu, Z.H. The Effect of Clostridium butyricum on Gut Microbiota, Immune Response and Intestinal Barrier Function During the Development of Necrotic Enteritis in Chickens. Front. Microbiol. 2019, 10, 2309. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; McCartney, E.; Knox, A.; Francesch, M.; Oka, K.; Wada, K.; Ideno, M.; Uno, K.; Kozłowski, K.; Jankowski, J.; et al. Effects of the Butyric Acid-Producing Strain Clostridium butyricum MIYAIRI 588 on Broiler and Piglet Zootechnical Performance and Prevention of Necrotic Enteritis. Anim. Sci. J. 2018, 89, 895–905. [Google Scholar] [CrossRef]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with Probiotic Enterococcus faecium NCIMB 11181 Ameliorates Necrotic Enteritis-Induced Intestinal Barrier Injury in Broiler Chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef] [PubMed]
- Huyghebaert, G.; Ducatelle, R.; Immerseel, F. van An Update on Alternatives to Antimicrobial Growth Promoters for Broilers. Vet. J. 2010, 187, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eeckhaut, V.; Wang, J.; van Parys, A.; Haesebrouck, F.; Joossens, M.; Falony, G.; Raes, J.; Ducatelle, R.; van Immerseel, F. The Probiotic Butyricicoccus pullicaecorum Reduces Feed Conversion and Protects from Potentially Harmful Intestinal Microorganisms and Necrotic Enteritis in Broilers. Front. Microbiol. 2016, 7, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, R.A.; Doranalli, K.; Rinttilä, T.; Vienola, K.; Jurgens, G.; Apajalahti, J. The Impact of Bacillus subtilis DSM 32315 on the Pathology, Performance, and Intestinal Microbiome of Broiler Chickens in a Necrotic Enteritis Challenge. Poult. Sci. 2019, 98, 3450–3463. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Zhang, N.; Han, J.-C.; Chang, C.-W.; Hsiao, F.S.-H.; Yu, Y.-H. Optimization of Surfactin Production from Bacillus Subtilis in Fermentation and Its Effects on Clostridium Perfringens- Induced Necrotic Enteritis and Growth Performance in Broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1232–1244. [Google Scholar] [CrossRef]
- Al-Baadani, H.H.; Abudabos, A.M.; Al-Mufarrej, S.I.; Al-Baadani, A.A.; Alhidary, I.A. Dietary Supplementation of Bacillus subtilis, Saccharomyces cerevisiae and Their Symbiotic Effect on Serum Biochemical Parameters in Broilers Challenged with Clostridium Perfringens. J. Appl. Anim. Res. 2018, 46, 1064–1072. [Google Scholar] [CrossRef] [Green Version]
- Sokale, A.O.; Menconi, A.; Mathis, G.F.; Lumpkins, B.; Sims, M.D.; Whelan, R.A.; Doranalli, K. Effect of Bacillus subtilis DSM 32315 on the Intestinal Structural Integrity and Growth Performance of Broiler Chickens under Necrotic Enteritis Challenge. Poult. Sci. 2019, 98, 5392–5400. [Google Scholar] [CrossRef]
- Musa, B.B.; Duan, Y.; Khawar, H.; Sun, Q.; Ren, Z.; Elsiddig Mohamed, M.A.; Abbasi, I.H.R.; Yang, X. Bacillus Subtilis B21 and Bacillus licheniformis B26 Improve Intestinal Health and Performance of Broiler Chickens with Clostridium perfringens-Induced Necrotic Enteritis. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1039–1049. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Wang, H.; Qing, X.; Sun, N.; Xin, J.; Luo, M.; Khalique, A.; Pan, K.; Shu, G.; et al. Dietary Probiotic Bacillus licheniformis H2 Enhanced Growth Performance, Morphology of Small Intestine and Liver, and Antioxidant Capacity of Broiler Chickens Against Clostridium perfringens–Induced Subclinical Necrotic Enteritis. Probiotics Antimicrob. Proteins 2020, 12, 883–895. [Google Scholar] [CrossRef]
- Hernandez-Patlan, D.; Solis-Cruz, B.; Pontin, K.P.; Hernandez-Velasco, X.; Merino-Guzman, R.; Adhikari, B.; López-Arellano, R.; Kwon, Y.M.; Hargis, B.M.; Arreguin-Nava, M.A.; et al. Impact of a Bacillus Direct-Fed Microbial on Growth Performance, Intestinal Barrier Integrity, Necrotic Enteritis Lesions, and Ileal Microbiota in Broiler Chickens Using a Laboratory Challenge Model. Front. Vet. Sci. 2019, 6, 108. [Google Scholar] [CrossRef]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 Improves Intestinal Health of Broiler Chickens Challenged with Clostridium perfringens-Induced Necrotic Enteritis. Poult. Sci. 2013, 92, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Das, P.P.; Saini, P.C.; Roy, B.; Chatterjee, P.N. Use of Bacillus subtilis PB6 as a Potential Antibiotic Growth Promoter Replacement in Improving Performance of Broiler Birds. Poult. Sci. 2017, 96, 2614–2622. [Google Scholar] [CrossRef] [PubMed]
- Teo, A.Y.L.; Tan, H.M. Inhibition of Clostridium Perfringens by a Novel Strain of Bacillus subtilis Isolated from the Gastrointestinal Tracts of Healthy Chickens. Appl. Environ. Microbiol. 2005, 71, 4185–4190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.S.; Kim, I.H. Effect of Bacillus Subtilis C-3102 Spores as a Probiotic Feed Supplement on Growth Performance, Noxious Gas Emission, and Intestinal Microflora in Broilers. Poult. Sci. 2014, 93, 3097–3103. [Google Scholar] [CrossRef] [PubMed]
- Rhayat, L.; Jacquier, V.; Brinch, K.S.; Nielsen, P.; Nelson, A.; Geraert, P.A.; Devillard, E. Bacillus subtilis Strain Specificity Affects Performance Improvement in Broilers. Poult. Sci. 2017, 96, 2274–2280. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, C.; Vieira, B.S.; de Paula Dorigam, J.C.; Menconi, A.; Sokale, A.; Doranalli, K.; Applegate, T.J. Bacillus subtilis Dsm 32315 Supplementation Attenuates the Effects of Clostridium perfringens Challenge on the Growth Performance and Intestinal Microbiota of Broiler Chickens. Microorganisms 2019, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Latorre, J.D.; Hernandez-Velasco, X.; Kuttappan, V.A.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Bielke, L.R.; Prado-Rebolledo, O.F.; Morales, E.; Hargis, B.M.; et al. Selection of Bacillus Spp. for Cellulase and Xylanase Production as Direct-Fed Microbials to Reduce Digesta Viscosity and Clostridium perfringens Proliferation Using an in Vitro Digestive Model in Different Poultry Diets. Front. Vet. Sci. 2015, 2, 25. [Google Scholar] [CrossRef]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the Use of Bacillus Species for Industrial Production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Zhou, M.; Zeng, D.; Ni, X.; Tu, T.; Yin, Z.; Pan, K.; Jing, B. Effects of Bacillus licheniformis on the Growth Performance and Expression of Lipid Metabolism-Related Genes in Broiler Chickens Challenged with Clostridium perfringens-Induced Necrotic Enteritis. Lipids Health Dis. 2016, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xu, S.; Zeng, D.; Ni, X.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; et al. Disruption in the Cecal Microbiota of Chickens Challenged with Clostridium perfringens and Other Factors Was Alleviated by Bacillus licheniformis Supplementation. PLoS ONE 2017, 12, e0182426. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B.; et al. Bacillus licheniformis Normalize the Ileum Microbiota of Chickens Infected with Necrotic Enteritis. Sci. Rep. 2018, 8, 1744. [Google Scholar] [CrossRef] [PubMed]
- Knap, I.; Lund, B.; Kehlet, A.B.; Hofacre, C.; Mathis, G. Bacillus licheniformis Prevents Necrotic Enteritis in Broiler Chickens. Avian Dis. 2010, 54, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Piao, X.; Ru, Y.; Zhang, H.; Peron, A.; Zhang, H. Effect of Bacillus amyloliquefaciens-based Direct-fed Microbial on Performance, Nutrient Utilization, Intestinal Morphology and Cecal Microflora in Broiler Chickens. Asian-Australas. J. Anim. Sci. 2015, 28, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerzsele, A.; Szeker, K.; Csizinszky, R.; Gere, E.; Jakab, C.; Mallo, J.J.; Galfi, P. Efficacy of Protected Sodium Butyrate, a Protected Blend of Essential Oils, Their Combination, and Bacillus amyloliquefaciens Spore Suspension against Artificially Induced Necrotic Enteritis in Broilers. Poult. Sci. 2012, 91, 837–843. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.J.K.; Sakomura, N.K.; de Paula Dorigam, J.C.; Doranalli, K.; Soares, L.; Viana, G.D.S. Bacillus amyloliquefaciens CECT 5940 Alone or in Combination with Antibiotic Growth Promoters Improves Performance in Broilers under Enteric Pathogen Challenge. Poult. Sci. 2019, 98, 4391–4400. [Google Scholar] [CrossRef]
- Tsukahara, T.; Inoue, R.; Nakayama, K.; Inatomi, T. Inclusion of Bacillus amyloliquefaciens Strain TOA5001 in the Diet of Broilers Suppresses the Symptoms of Coccidiosis by Modulating Intestinal Microbiota. Anim. Sci. J. 2018, 89, 679–687. [Google Scholar] [CrossRef]
- Mu, Y.; Cong, Y. Bacillus coagulans and Its Applications in Medicine. Benef. Microbes 2019, 10, 679–688. [Google Scholar] [CrossRef]
- Kawarizadeh, A.; Tabatabaei, M.; Hosseinzadeh, S.; Farzaneh, M.; Pourmontaseri, M. The Effects of Probiotic Bacillus coagulans on the Cytotoxicity and Expression of Alpha Toxin Gene of Clostridium Perfringens Type A. Anaerobe 2019, 59, 61–67. [Google Scholar] [CrossRef]
- Calik, A.; Omara, I.I.; White, M.B.; Evans, N.P.; Karnezos, T.P.; Dalloul, R.A. Dietary Non-Drug Feed Additive as an Alternative for Antibiotic Growth Promoters for Broilers during a Necrotic Enteritis Challenge. Microorganisms 2019, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus coagulans in the Food Industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Patlan, D.; Solis-Cruz, B.; Latorre, J.D.; Merino-Guzman, R.; Morales Rodríguez, M.; Ausland, C.; Hernandez-Velasco, X.; Ortiz Holguin, O.; Delgado, R.; Hargis, B.M.; et al. Whole-Genome Sequence and Interaction Analysis in the Production of Six Enzymes From the Three Bacillus Strains Present in a Commercial Direct-Fed Microbial (NorumTM) Using a Bliss Independence Test. Front. Vet. Sci. 2022, 9, 784387. [Google Scholar] [CrossRef] [PubMed]
- Krueger, L.A.; Spangler, D.A.; Vandermyde, D.R.; Sims, M.D.; Ayangbile, G.A. Avi-Lution® Supplemented at 1.0 or 2.0 g/Kg in Feed Improves the Growth Performance of Broiler Chickens during Challenge with Bacitracin-Resistant Clostridium Perfringens. Poult. Sci. 2017, 96, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Granstad, S.; Kristoffersen, A.B.; Benestad, S.L.; Sjurseth, S.K.; David, B.; Sørensen, L.; Fjermedal, A.; Edvardsen, D.H.; Sanson, G.; Løvland, A.; et al. Effect of Feed Additives as Alternatives to In-Feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens. Animals 2020, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-M.; Cheng, W.T.K.; Chang, Y.-C.; Chang, T.-J.; Chen, H.-L. Growth Enhancement of Fowls by Dietary Administration of Recombinant Yeast Cultures Containing Enriched Growth Hormone. Life Sci. 2000, 67, 2103–2115. [Google Scholar] [CrossRef]
- de los Santos, G.J.R.; Storch, O.B.; Fernandes, C.G.; Gil-Turnes, C. Evaluation in Broilers of the Probiotic Properties of Pichia pastoris and a Recombinant P. pastoris Containing the Clostridium perfringens Alpha Toxin Gene. Vet. Microbiol. 2012, 156, 448–451. [Google Scholar] [CrossRef]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Ahiwe, E.U.; Chang’a, E.P.; Abdallh, M.E.; Al-Qahtani, M.; Kheravii, S.K.; Wu, S.; Graham, H.; Iji, P.A. Dietary Hydrolysed Yeast Cell Wall Extract Is Comparable to Antibiotics in the Control of Subclinical Necrotic Enteritis in Broiler Chickens. Br. Poult. Sci. 2019, 60, 757–765. [Google Scholar] [CrossRef]
- Thompson, D.R.; Parreira, V.R.; Kulkarni, R.R.; Prescott, J.F. Live Attenuated Vaccine-Based Control of Necrotic Enteritis of Broiler Chickens. Vet. Microbiol. 2006, 113, 25–34. [Google Scholar] [CrossRef]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Immunization of Broiler Chickens against Clostridium perfringens-Induced Necrotic Enteritis. Clin. Vaccine Immunol. 2007, 14, 1070–1077. [Google Scholar] [CrossRef] [Green Version]
- Daneshmand, A.; Kermanshahi, H.; Mohammed, J.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M.; Alizadeh, M.; Razmyar, J.; Kulkarni, R.R. Intestinal Changes and Immune Responses during Clostridium perfringens-Induced Necrotic Enteritis in Broiler Chickens. Poult. Sci. 2022, 101, 101652. [Google Scholar] [CrossRef]
- Mcreynolds, J.L.; Byrd, J.A.; Anderson, R.C.; Moore, R.W.; Edrington, T.S.; Genovese, K.J.; Poole, T.L.; Kubena, L.F.; Nisbet, D.J. Evaluation of Immunosuppressants and Dietary Mechanisms in an Experimental Disease Model for Necrotic Enteritis. Poult. Sci. 2004, 83, 1948–1952. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The Successful Experimental Induction of Necrotic Enteritis in Chickens by Clostridium Perfringens: A Critical Review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacey, J.A.; Stanley, D.; Keyburn, A.L.; Ford, M.; Chen, H.; Johanesen, P.; Lyras, D.; Moore, R.J. Clostridium perfringens-Mediated Necrotic Enteritis Is Not Influenced by the Pre-Existing Microbiota but Is Promoted by Large Changes in the Post-Challenge Microbiota. Vet. Microbiol. 2018, 227, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Heak, C.; Sukon, P.; Sornplang, P. Effect of Direct-Fed Microbials on Culturable Gut Microbiotas in Broiler Chickens: A Meta-Analysis of Controlled Trials. Asian-Australas. J. Anim. Sci. 2018, 31, 1781–1794. [Google Scholar] [CrossRef]
- Feng, Y.; Gong, J.; Yu, H.; Jin, Y.; Zhu, J.; Han, Y. Identification of Changes in the Composition of Ileal Bacterial Microbiota of Broiler Chickens Infected with Clostridium perfringens. Vet. Microbiol. 2010, 140, 116–121. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Kokkotou, E.G.; Mustafa, N.; Bhaskar, K.R.; Sougioultzis, S.; O’Brien, M.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii Inhibits ERK1/2 Mitogen-Activated Protein Kinase Activation Both in Vitro and in Vivo and Protects against Clostridium difficile Toxin A-Induced Enteritis. J. Biol. Chem. 2006, 281, 24449–24454. [Google Scholar] [CrossRef] [Green Version]
- Ramlucken, U.; Ramchuran, S.O.; Moonsamy, G.; Lalloo, R.; Thantsha, M.S.; Jansen van Rensburg, C. A Novel Bacillus Based Multi-Strain Probiotic Improves Growth Performance and Intestinal Properties of Clostridium perfringens Challenged Broilers. Poult. Sci. 2020, 99, 331–341. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergström, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schütte, A.; van der Post, S.; Svensson, F.; Rodríguez-Piñeiro, A.M.; Nyström, E.E.L.; et al. The Mucus and Mucins of the Goblet Cells and Enterocytes Provide the First Defense Line of the Gastrointestinal Tract and Interact with the Immune System. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of Commensal and Pathogenic Microorganisms with the Intestinal Mucosal Barrier. Nat. Rev. Microbiol. 2018, 16, 457–470. [Google Scholar] [CrossRef]
- Mattar, A.F.; Teitelbaum, D.H.; Drongowski, R.A.; Yongyi, F.; Harmon, C.M.; Coran, A.G. Probiotics Up-Regulate MUC-2 Mucin Gene Expression in a Caco-2 Cell-Culture Model. Pediatric Surg. Int. 2002, 18, 586–590. [Google Scholar]
- Boekhorst, J.; Helmer, Q.; Kleerebezem, M.; Siezen, R.J. Comparative Analysis of Proteins with a Mucus-Binding Domain Found Exclusively in Lactic Acid Bacteria. Microbiology 2006, 152, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion Mechanisms Mediated by Probiotics and Prebiotics and Their Potential Impact on Human Health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between Commensal Bacteria and the Gut-Associated Immune System of the Chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisbin, J.T.; Davidge, L.; Roshdieh, A.; Sharif, S. Characterization of the Effects of Three Lactobacillus Species on the Function of Chicken Macrophages. Res. Vet. Sci. 2015, 100, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Kulkarni, R.R.; Brisbin, J.T.; Quinteiro-Filho, W.; Alkie, T.N.; Sharif, S. Interactions between Lactobacilli and Chicken Macrophages Induce Antiviral Responses against Avian Influenza Virus. Res. Vet. Sci. 2019, 125, 441–450. [Google Scholar] [CrossRef]
- Cox, C.M.; Dalloul, R.A. Immunomodulatory Role of Probiotics in Poultry and Potential in Ovo Application. Benef. Microbes 2015, 6, 45–52. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Astill, J.; Kulkarni, R.R.; Read, L.R.; Najarian, A.; Farber, J.M.; Sharif, S. In Vitro Assessment of Immunomodulatory and Anti-Campylobacter Activities of Probiotic Lactobacilli. Sci. Rep. 2019, 9, 17903. [Google Scholar] [CrossRef]
- Plantinga, T.S.; van Maren, W.W.C.; van Bergenhenegouwen, J.; Hameetman, M.; Nierkens, S.; Jacobs, C.; de Jong, D.J.; Joosten, L.A.B.; Van’t Land, B.; Garssen, J.; et al. Differential Toll-like Receptor Recognition and Induction of Cytokine Profile by Bifidobacterium breve and Lactobacillus Strains of Probiotics. Clin. Vaccine Immunol. 2011, 18, 621–628. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and Immune Health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Qi, L.; Wu, T.; Wang, J. Clostridium butyricum Activates TLR2-Mediated MyD88-Independent Signaling Pathway in HT-29 Cells. Mol. Cell Biochem. 2012, 361, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Diergeneeskunde, F. Virulence mechanisms of Clostridium perfringens in Broiler Necrotic Enteritis. Ph.D. Thesis, Faculty of Veterinary Medicine, Department of Pathology, Bacteriology, and Poultry Diseases, Ghent University, Ghent, Belgium, 2009. [Google Scholar]
- Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Charrier-Hisamuddin, L.; Sitaraman, S.V.; Merlin, D. Butyrate Transcriptionally Enhances Peptide Transporter PepT1 Expression and Activity. PLoS ONE 2008, 3, e2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, A.; Cotter, P.D.; Paul Ross, R.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins-a Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Lagha, B.A.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial Potential of Bacteriocins in Poultry and Swine Production. Vet. Res. 2017, 48, 22. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.N.; Kogut, M.H.; Genovese, K.; He, H.; Kazemi, S.; Arsenault, R.J. Administration of a Postbiotic Causes Immunomodulatory Responses in Broiler Gut and Reduces Disease Pathogenesis Following Challenge. Microorganisms 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens 2022, 11, 692. https://doi.org/10.3390/pathogens11060692
Kulkarni RR, Gaghan C, Gorrell K, Sharif S, Taha-Abdelaziz K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens. 2022; 11(6):692. https://doi.org/10.3390/pathogens11060692
Chicago/Turabian StyleKulkarni, Raveendra R., Carissa Gaghan, Kaitlin Gorrell, Shayan Sharif, and Khaled Taha-Abdelaziz. 2022. "Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens" Pathogens 11, no. 6: 692. https://doi.org/10.3390/pathogens11060692