
Virus Surveys in Olive Orchards in Greece Identify Olive Virus T, a Novel Member of the Genus Tepovirus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
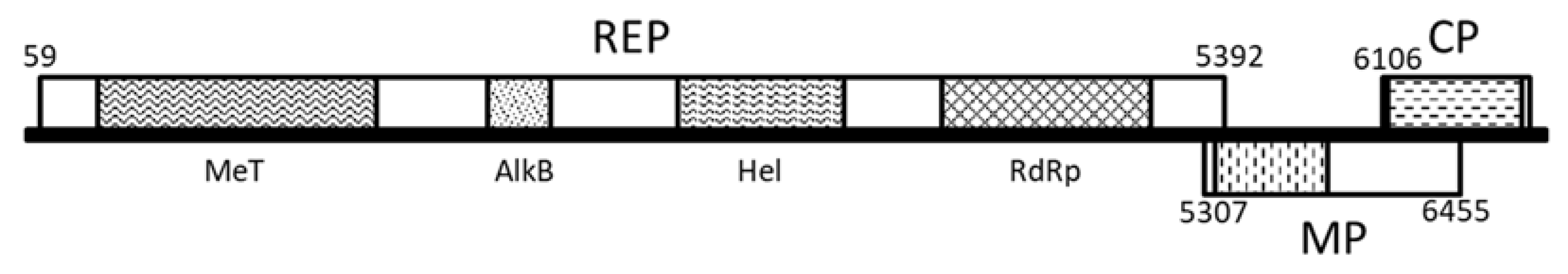
2.1. Complete Genome Sequence Determination of a Novel Tepovirus
2.2. Transmission of OlVT by Grafting
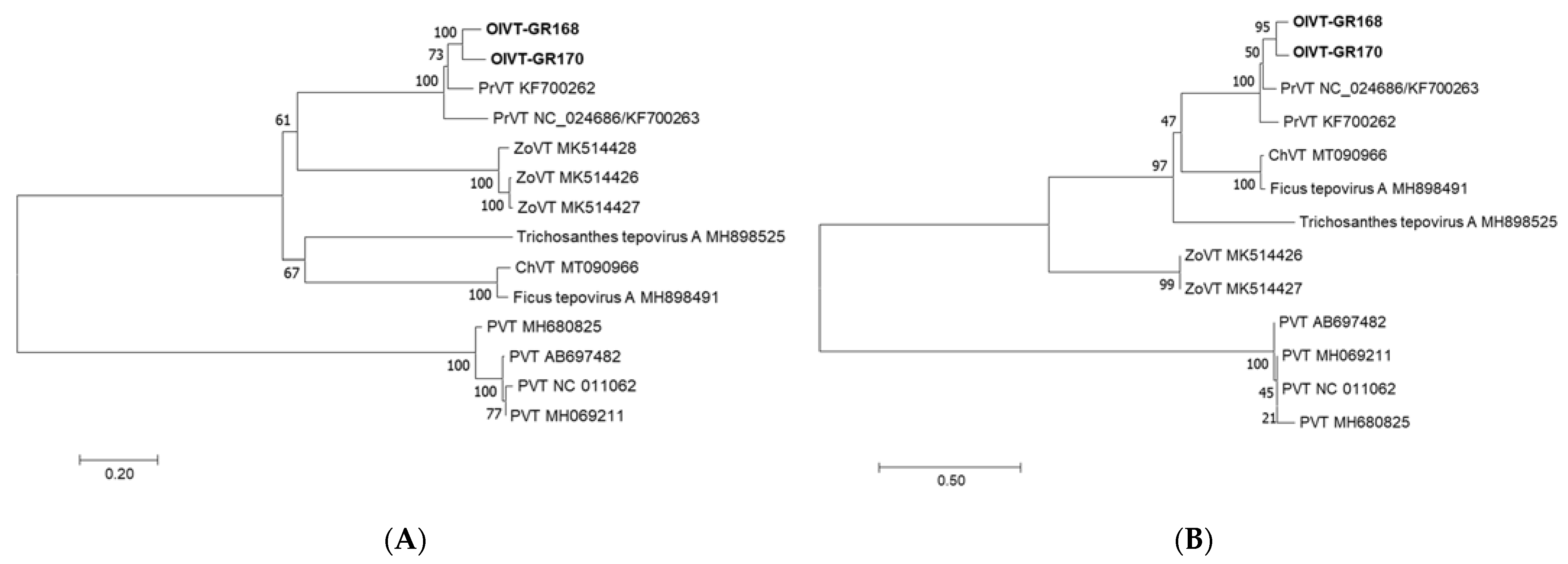
2.3. Molecular Variability of OlVT
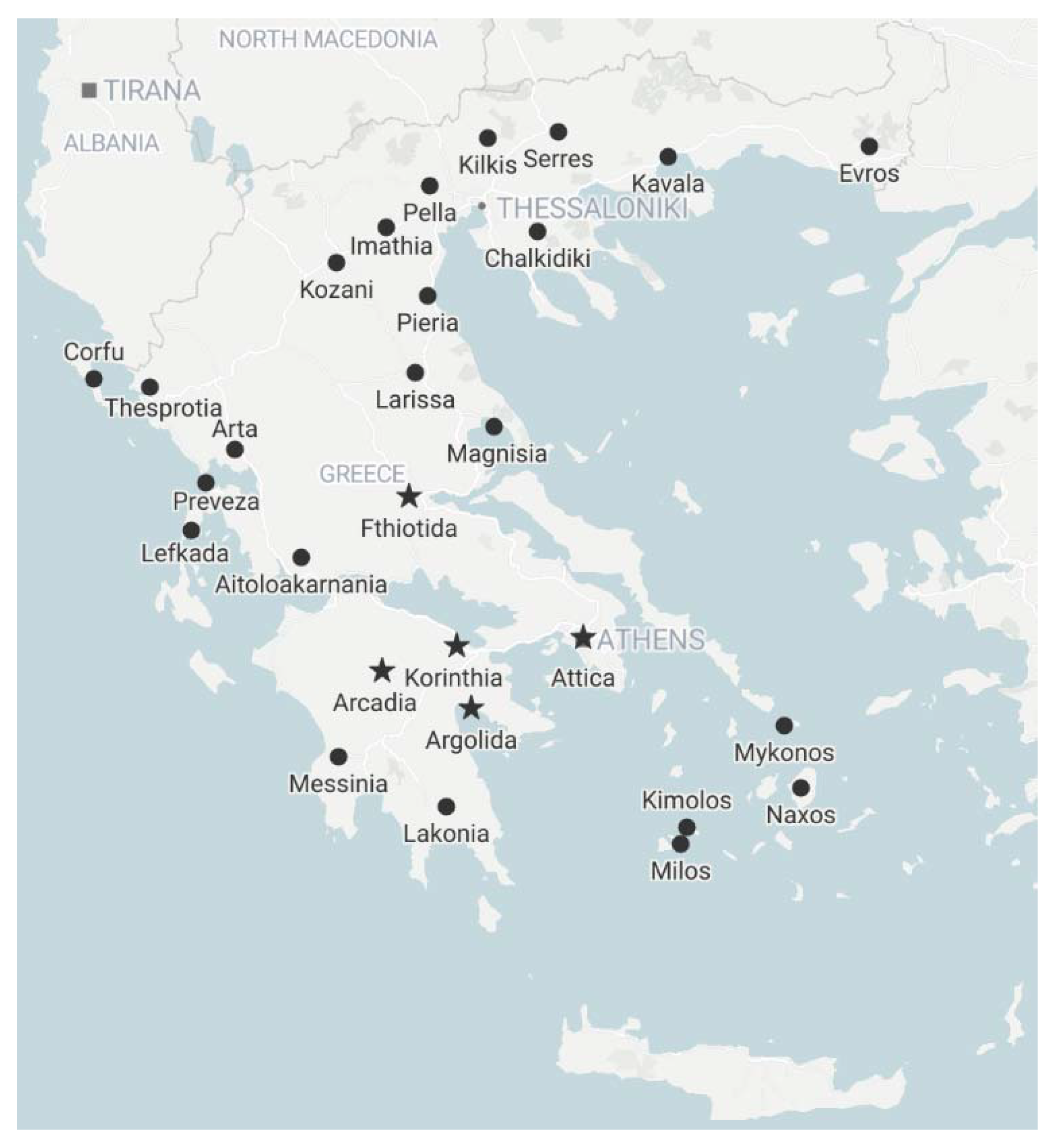
2.4. Viruses in Greek Olive Orchards
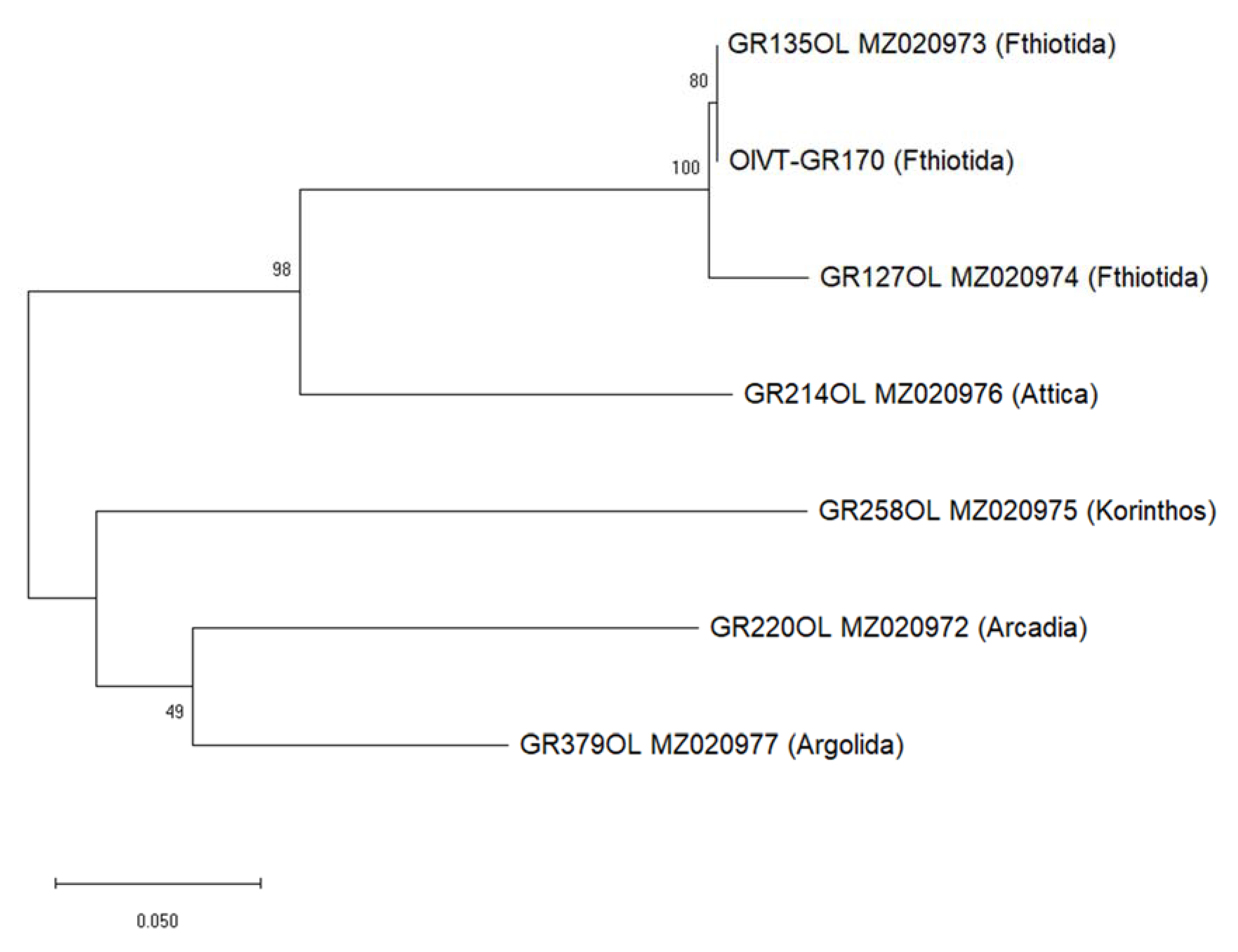
2.5. Molecular Variability of Greek Isolates of Olive Viruses
2.5.1. CMV RdRp Gene
2.5.2. OLYaV HSP70h Gene
2.5.3. CLRV 3’ UTR
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. HTS and Genome Assembly
4.3. Detection of Olive Viruses by RT–PCR
4.4. Sequence and Phylogenetic Analyses
4.5. Transmission of the Novel Tepovirus by Grafting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eurostat. 2017. Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news/-/DDN-20190301-1 (accessed on 28 March 2021).
- Martelli, G.P. A brief outline of infectious diseases of olive. Palestine Tech. Univ. Res. J. 2013, 1, 1–9. [Google Scholar]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://ictv.global/taxonomy/ (accessed on 25 March 2019).
- Kyriakopoulou, P.E. Olive fruit hump and olive fruit pox: Two new disease of possible viral origin. Phytopathol. Mediterr. 1996, 35, 231–232. [Google Scholar]
- Mathioudakis, M.M.; Saponari, M.; Hasiów-Jaroszewska, B.; Elbeaino, T.; Koubouris, G. Detection of viruses in olive cultivars in Greece, using a rapid and effective RNA extraction method, for certification of virus-tested propagation material. Phytopathol. Mediterr. 2020, 59, 203–211. [Google Scholar] [CrossRef]
- Alabdullah, A.; Elbeaino, T.; Minafra, A.; Digiaro, M.; Martelli, G.P. Detection and variability of olive latent virus 3 in the mediterranean region. J. Plant Pathol. 2009, 91, 521–525. [Google Scholar] [CrossRef]
- Grieco, F.; Parrella, G.; Vovlas, C. An isolate of Olive latent virus 2 infecting castor bean in Greece. J. Plant Pathol. 2002, 84, 129–131. [Google Scholar] [CrossRef]
- Gratsia, M.E.; Kyriakopoulou, P.E.; Voloudakis, A.E.; Fasseas, C.; Tzanetakis, I.E. First report of Olive mild mosaic virus and Sowbane mosaic virus in spinach in Greece. Plant Dis. 2012, 96, 1230. [Google Scholar] [CrossRef]
- Al-Rwahnih, M.; Guo, Y.; Daubert, S.; Golino, D.; Rowhani, A. Characterization of latent viral infection of olive trees in the national clonal germplasm repository in California. J. Plant Pathol. 2011, 93, 227–231. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Mustafayev, E.; Barone, M.; Alioto, D.; Candresse, T. Characterization by deep sequencing of prunus virus T, a novel Tepovirus infecting Prunus species. Phytopathology 2015, 105, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Rubino, L.; Russo, M.; de Stradis, A.; Martelli, G.P. Tepovirus, a novel genus in the family Betaflexiviridae. Arch. Virol. 2012, 157, 1629–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.J.; Candresse, T.; Hammond, J.; Kreuze, J.F.; Martelli, G.P.; Namba, S.; Pearson, M.N.; Ryu, K.H.; Saldarelli, P.; Yoshikawa, N. Family Betaflexiviridae. In Virus Taxonomy. 9th Report of the ICTV; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: Amsterdam, The Newtherland, 2012; pp. 920–941. [Google Scholar]
- Goh, C.J.; Park, D.; Lee, J.S.; Davey, P.A.; Pernice, M.; Ralph, P.J.; Hahn, Y. Zostera virus T-a novel virus of the genus Tepovirus identified in the eelgrass, Zostera muelleri. Acta Virol. 2019, 63, 366–372. [Google Scholar] [CrossRef]
- Marais, A.; Šafářová, D.; Navrátil, M.; Faure, C.; Cornaggia, D.; Brans, Y.; Suchá, J.; Candresse, T. Complete genome sequence of cherry virus T, a novel cherry-infecting tepovirus. Arch. Virol. 2020, 165, 1711–1714. [Google Scholar] [CrossRef]
- Kutnjak, D.; Tamisier, L.; Adams, I.; Boonham, N.; Candresse, T.; Chiumenti, M.; De Jonghe, K.; Kreuze, J.F.; Lefebvre, M.; Silva, G.; et al. A primer on the analysis of high-throughput sequencing data for detection of plant viruses. Microorganisms 2021, 9, 841. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A new grapevine virus discovered by deep sequencing of virus- and viroid-derived small RNAs in Cv Pinot gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent advances on detection and characterization of fruit tree viruses using high-throughput sequencing technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, L.F.; Harrison, B.D. Host range, purification and properties of potato virus T. Ann. Appl. Biol. 1978, 89, 223–235. [Google Scholar] [CrossRef]
- Al Abdullah, A.; El Beaino, T.; Saponari, M.; Hallak, H.; Digiaro, M. Preliminary evaluation of the status of olive-infecting viruses in Syria. EPPO Bull. 2005, 35, 249–252. [Google Scholar] [CrossRef]
- Fadel, C.; Digiaro, M.; Choueiri, E.; Beaino, T.E.; Saponari, M.; Savino, V.; Martelli, G.P. On the presence and distribution of olive viruses in Lebanon. EPPO Bull. 2005, 35, 33–36. [Google Scholar] [CrossRef]
- Faggioli, F.; Ferretti, L.; Albanese, G.; Sciarroni, R.; Pasquini, G.; Lumia, V.; Barba, M. Distribution of olive tree viruses in Italy as revealed by one-step RT-PCR. J. Plant Pathol. 2005, 87, 49–55. [Google Scholar] [CrossRef]
- Varanda, C.; Cardoso, J.M.S.; do Rosário Félix, M.; Oliveira, S.; Clara, M.I. Multiplex RT-PCR for detection and identification of three necroviruses that infect olive trees. Eur. J. Plant Pathol. 2010, 127, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Yousef, S.A.; Moawed, S.M.; El-Sayed, M.; Shalaby, A.A. Detection of olive tree viruses in Egypt by one-step RT-PCR. In Proceedings of the 21st International Conference on Virus and Other Graft Transmissible Diseases of Fruit Crops, Neustadt, Germany, 5–10 July 2009; pp. 51–52. [Google Scholar]
- Zellama, M.S.; Varanda, C.M.R.; Materatski, P.; Nabi, N.; Hafsa, A.B.; Saamali, B.M.; Chaouachi, M.; Félix, M.R. An integrated approach for understanding the high infection rates of olive viruses in Tunisia. Eur. J. Plant Pathol. 2019, 153, 1043–1054. [Google Scholar] [CrossRef]
- Valachas, C.A.; Giantsis, I.A.; Sareli, K.; Winter, S.; Zelezniakof, E.; Pentheroudaki, Z.; Chatzivassiliou, E.K. Molecular analysis of Greek isolates of cucumber mosaic virus from vegetables show a low prevalence of satellite RNAs and suggest the presence of host-associated virus strains. Arch. Virol. 2021, 166, accepted. [Google Scholar]
- Çağlayan, K.; Faggioli, F.; Barba, M. Virus, phytoplasma and unknown diseases of olive trees. In Virus and Viruslike Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS: St. Paul, MN, USA, 2011; pp. 289–297. [Google Scholar]
- Afechtal, M.; Mounir, M. Preliminary evaluation of the status of olive-infecting viruses in Morocco. Mor. J. Agric. Sci. 2020, 1, 59–61. [Google Scholar]
- Jacquemond, M. Cucumber mosaic virus. Adv. Virus Res. 2012, 84, 439–504. [Google Scholar] [PubMed]
- Ruiz-García, A.B.; Candresse, T.; Canales, C.; Morán, F.; de Oliveira, C.M.; Bertolini, E.; Olmos, A. Molecular characterization of the complete coding sequence of olive leaf yellowing-associated virus. Plants 2020, 9, 1272. [Google Scholar] [CrossRef]
- Albanese, G.; Saponari, M.; Faggioli, F. Phytosanitary Certification. In Olive Germplasm. The Olive Cultivation, Table Olive and Olive Oil Industry in Italy; Muzzalupo, I., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Cutuli, M.; Lo Bianco, R.; Marra, F.P.; Caruso, T. Vegetative growth and ecophysiological aspects in young olive plants inoculated with olive leaf yellowing associated virus (OLYaV). Acta Italus Hortus. 2011, 1, 356–361. [Google Scholar]
- Campos, M.D.; Zellama, M.S.; Varanda, C.; Materatski, P.; Peixe, A.; Chaouachi, M.; Félix, M. do R. Establishment of a sensitive qPCR methodology for detection of the olive-infecting viruses in Portuguese and Tunisian orchards. Front. Plant Sci. 2019, 10, 694. [Google Scholar] [CrossRef]
- Büttner, C.; von Bargen, S.; Bandte, M.; Myrta, A. Cherry leaf roll virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St Paul, MN, USA, 2011; pp. 119–125. [Google Scholar]
- Kaponi, M.S.; Kyriakopoulou, P.E. Olive viruses in Greece. Phytopathol. Mediterr. 2008, 47, 152–153. [Google Scholar]
- Sclavounos, A.P.; Kyriakopoulou, P.E.; Holeva, M.C.; Kostas, P.; Voloudakis, A.E. Detection of Cherry leaf roll (CLRV) from walnut (Juglans regia L.) in Greece. Phytopathol. Mediterr. 2008, 47, 159. [Google Scholar]
- Saponari, M.; Savino, V.; Martelli, G.P. Seed transmission in olive of two olive infecting viruses. J. Plant Pathol. 2002, 84, 167–168. [Google Scholar]
- Rebenstorf, K.; Candresse, T.; Dulucq, M.J.; Büttner, C.; Obermeier, C. Host species-dependent population structure of a pollen-borne plant virus, cherry leaf roll virus. J. Virol. 2006, 80, 2453–2462. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xylogianni, E.; Margaria, P.; Knierim, D.; Sareli, K.; Winter, S.; Chatzivassiliou, E.K. Virus Surveys in Olive Orchards in Greece Identify Olive Virus T, a Novel Member of the Genus Tepovirus. Pathogens 2021, 10, 574. https://doi.org/10.3390/pathogens10050574
Xylogianni E, Margaria P, Knierim D, Sareli K, Winter S, Chatzivassiliou EK. Virus Surveys in Olive Orchards in Greece Identify Olive Virus T, a Novel Member of the Genus Tepovirus. Pathogens. 2021; 10(5):574. https://doi.org/10.3390/pathogens10050574
Chicago/Turabian StyleXylogianni, Evanthia, Paolo Margaria, Dennis Knierim, Kyriaki Sareli, Stephan Winter, and Elisavet K. Chatzivassiliou. 2021. "Virus Surveys in Olive Orchards in Greece Identify Olive Virus T, a Novel Member of the Genus Tepovirus" Pathogens 10, no. 5: 574. https://doi.org/10.3390/pathogens10050574