
Investigating the Metabolism of Estrogens in Ligilactobacillus salivarius Strains Isolated from Human Milk and Vaginal Microbiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Bacterial Growth in the Presence of Estrogens
2.3. Extraction of Estrogens
2.4. Determination of Estrogens and Derivatives by Triple-Quadrupole Mass Spectrometry Coupled with High-Performance Liquid Chromatography (LC-QQQ-MS)
2.5. PCR Detection of Estrogen Metabolism-Related Enzymes
2.6. Statistical Analysis
3. Results
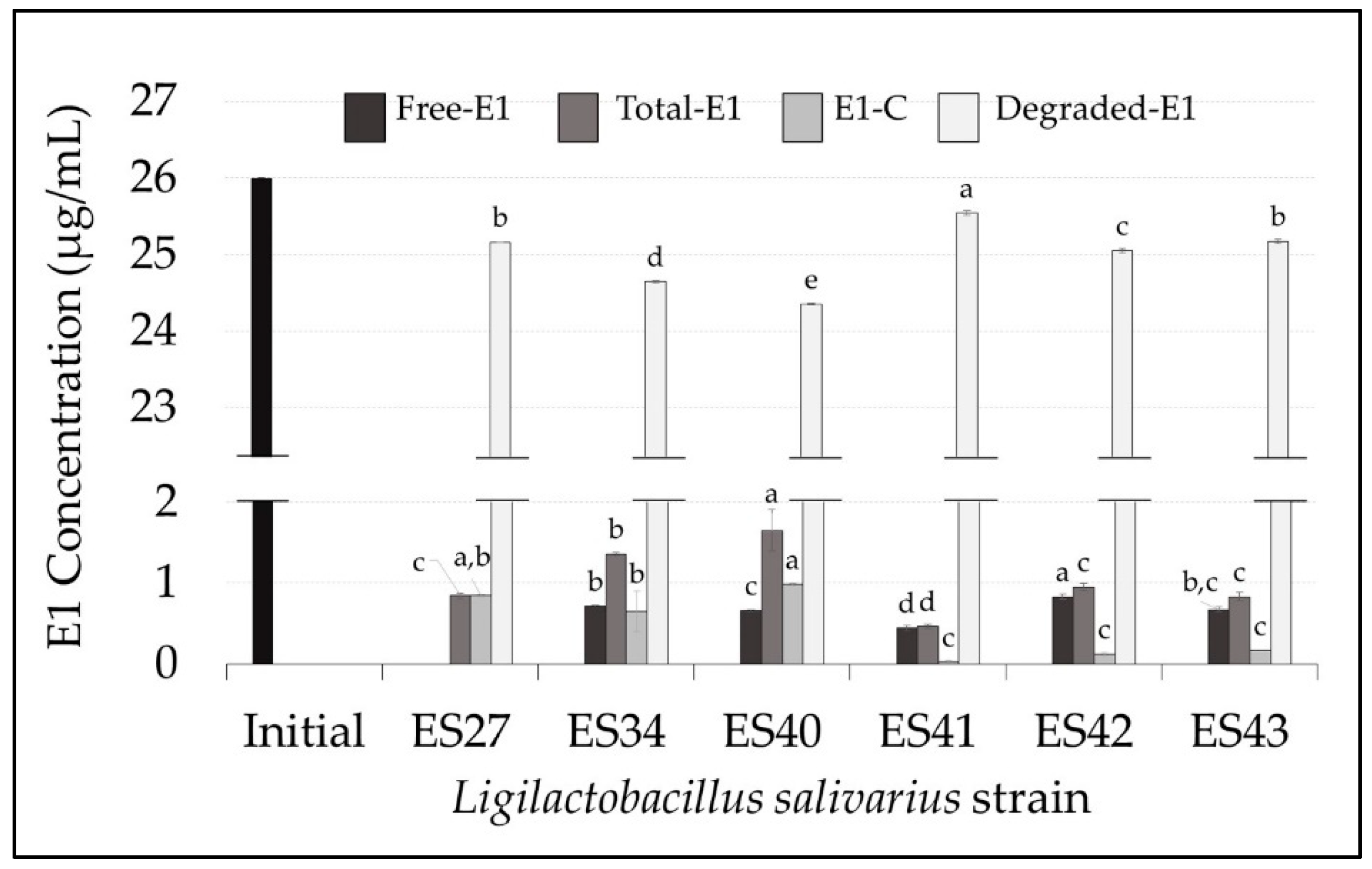
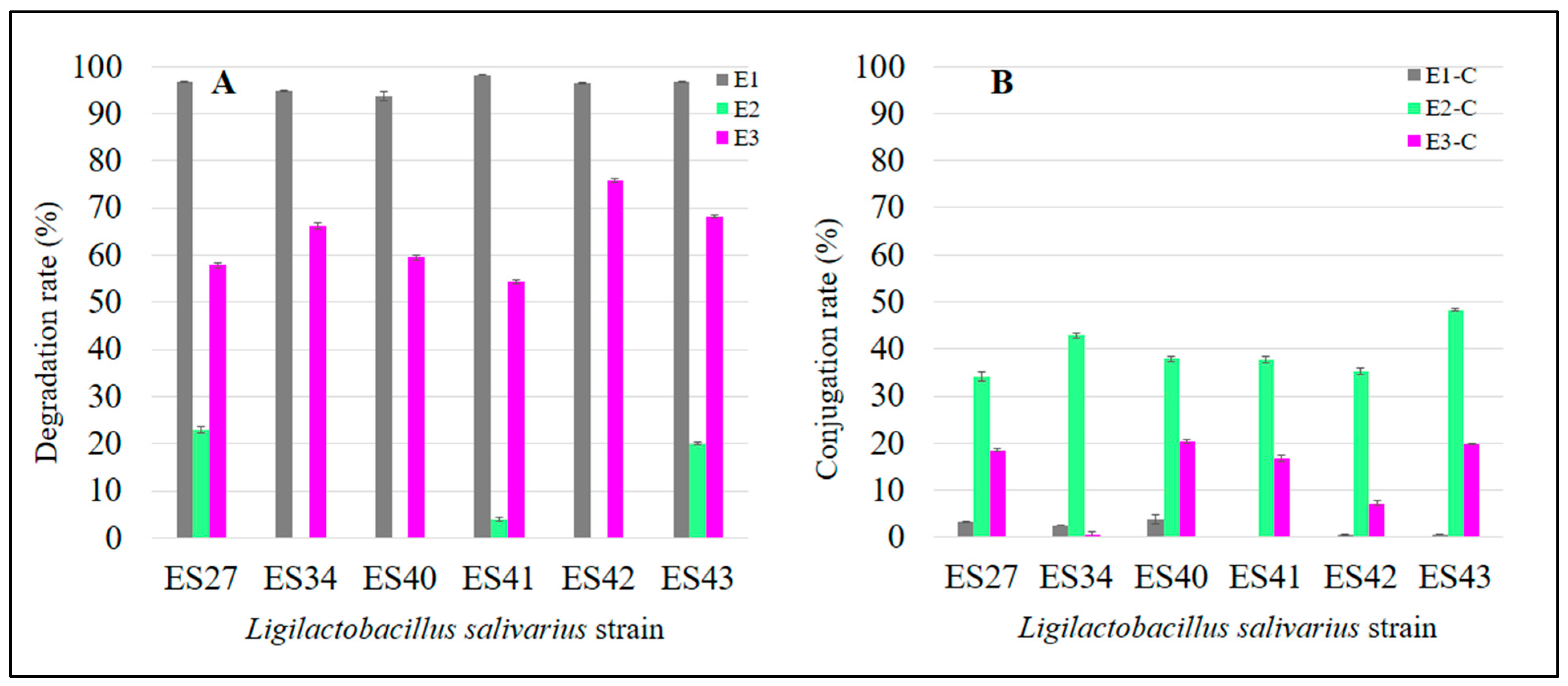
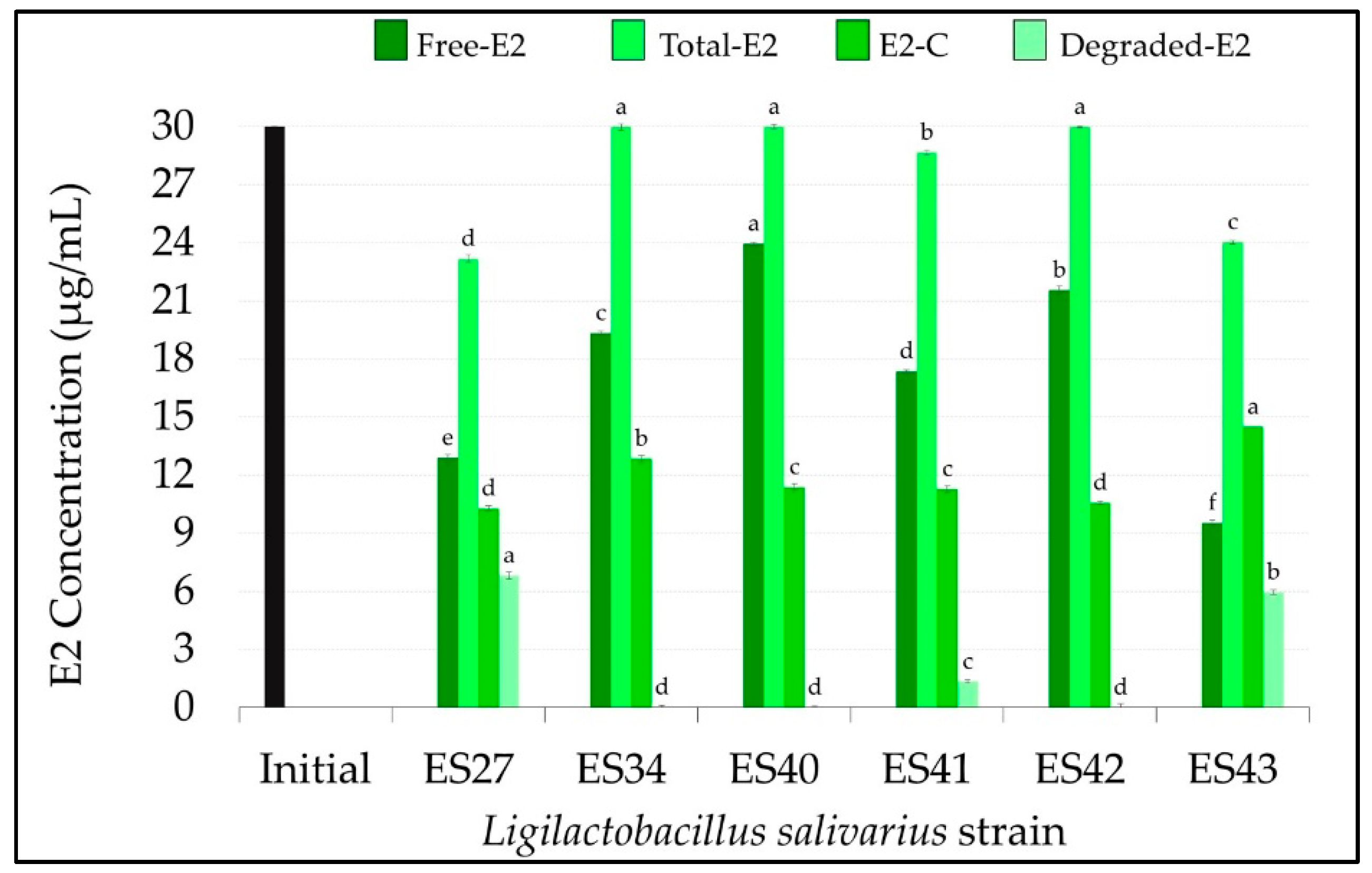
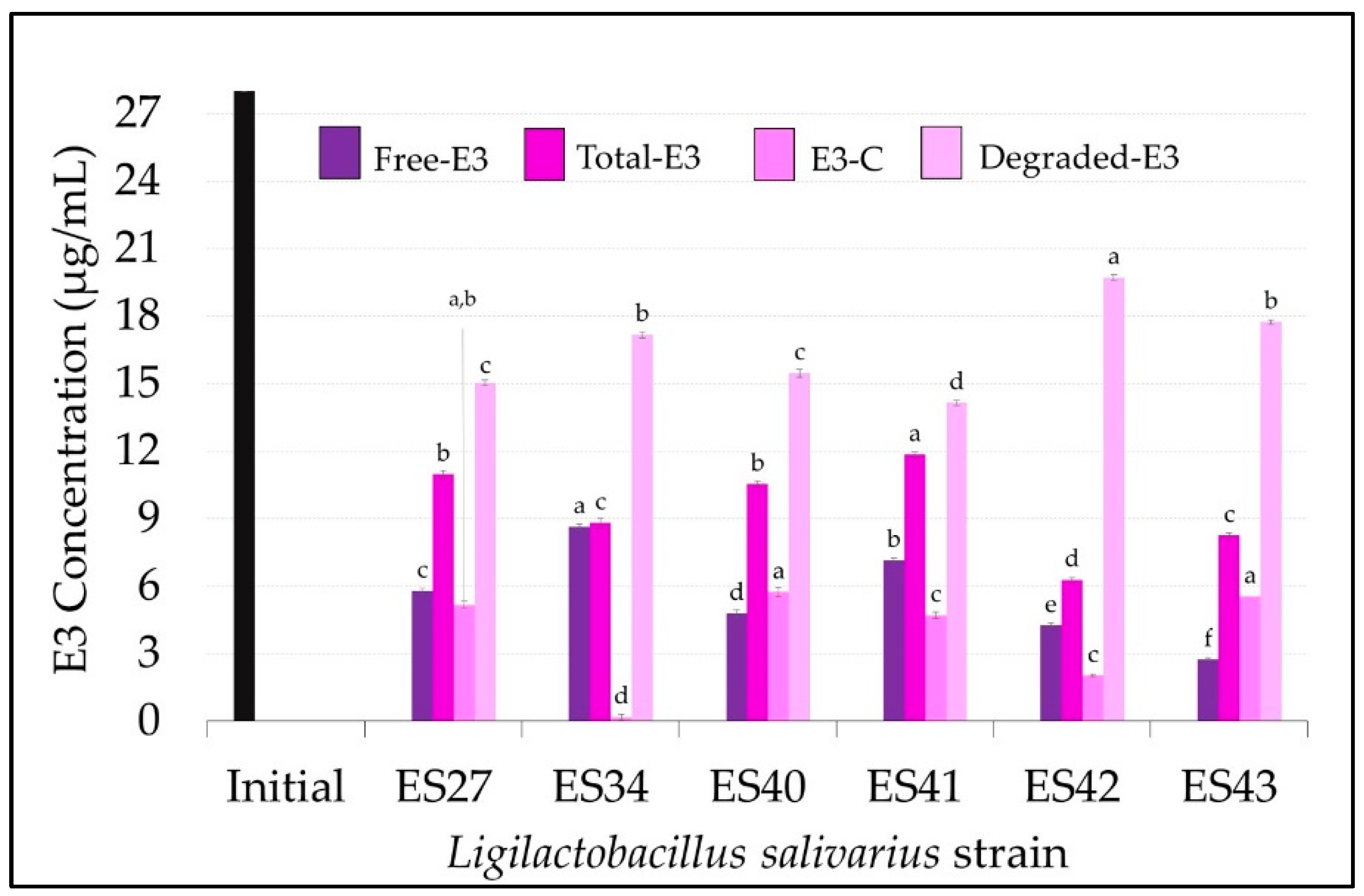
3.1. Ligilactobacillus salivarius Strains Can Degrade and Conjugate Parental Estrogens
3.2. Ligilactobacillus salivarius Strains Isolated from Human Milk and Vaginal Samples Exhibit Genetic Capacity for Estrogen Degradation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eyster, K.M. The estrogen receptors: An overview from different perspectives. Methods Mol. Biol. 2016, 1366, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, B.; Ou-Yang, L. Role of estrogen receptors in health and disease. Front. Endocrinol. 2022, 13, 839005. [Google Scholar] [CrossRef]
- Farage, M.; Maibach, H. Lifetime changes in the vulva and vagina. Arch. Gynecol. Obstet. 2006, 273, 195–202. [Google Scholar] [CrossRef]
- Muhleisen, A.L.; Herbst-Kralovetz, M.M. Menopause and the vaginal microbiome. Maturitas 2016, 91, 42–50. [Google Scholar] [CrossRef]
- Kwon, M.S.; Lee, H.K. Host and Microbiome Interplay Shapes the Vaginal Microenvironment. Front. Immunol. 2022, 13, 919728. [Google Scholar] [CrossRef] [PubMed]
- Cheskis, B.J.; Greger, J.G.; Nagpal, S.; Freedman, L.P. Signaling by estrogens. J. Cell. Physiol. 2007, 213, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Biason-Lauber, A.; Lang-Muritano, M. Estrogens: Two nuclear receptors, multiple possibilities. Mol. Cell. Endocrinol. 2022, 554, 111710. [Google Scholar] [CrossRef]
- Ingraham, H.A.; Herber, C.B.; Krause, W.C. Running the female power grid across lifespan through brain estrogen signaling. Ann. Rev. Physiol. 2022, 84, 59–85. [Google Scholar] [CrossRef]
- Hannan, F.M.; Elajnaf, T.; Vandenberg, L.N.; Kennedy, S.H.; Thakker, R.V. Hormonal regulation of mammary gland development and lactation. Nat. Rev. Endocrinol. 2023, 19, 46–61. [Google Scholar] [CrossRef]
- Knowlton, A.A.; Lee, A.R. Estrogen and the cardiovascular system. Pharmacol. Ther. 2012, 135, 54–70. [Google Scholar] [CrossRef]
- Baudry, M.; Bi, X.; Aguirre, C. Progesterone-estrogen interactions in synaptic plasticity and neuroprotection. Neuroscience 2013, 239, 280–294. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef]
- Smy, L.; Straseski, J.A. Measuring estrogens in women, men, and children: Recent advances 2012–2017. Clin. Biochem. 2018, 62, 11–23. [Google Scholar] [CrossRef]
- Patel, S.; Homaei, A.; Raju, A.B.; Meher, B.R. Estrogen: The necessary evil for human health, and ways to tame it. Biomed. Pharmacother. 2018, 102, 403–411. [Google Scholar] [CrossRef]
- Yager, J.D.; Davidson, N.E. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef]
- Meyer, M.R.; Clegg, D.J.; Prossnitz, E.R.; Barton, M. Obesity, insulin resistance and diabetes: Sex differences and role of oestrogen receptors. Acta Physiol. 2011, 203, 259–269. [Google Scholar] [CrossRef]
- Walker, S.E. Estrogen and autoimmune disease. Clin. Rev. Allergy Immunol. 2011, 40, 60–65. [Google Scholar] [CrossRef]
- Barnabei, V.M.; Cochrane, B.B.; Aragaki, A.K.; Nygaard, I.; Williams, R.S.; McGovern, P.G.; Young, R.L.; Wells, E.C.; O’Sullivan, M.J.; Chen, B.; et al. Menopausal symptoms and treatment-related effects of estrogen and progestin in the Women’s Health Initiative. Obstet. Gynecol. 2005, 105, 1063–1073. [Google Scholar] [CrossRef]
- Au, A.; Feher, A.; McPhee, L.; Jessa, A.; Oh, S.; Einstein, G. Estrogens, inflammation and cognition. Front. Neuroendocrinol. 2016, 40, 87–100. [Google Scholar] [CrossRef]
- Coyoy, A.; Guerra-Araiza, C.; Camacho-Arroyo, I. Metabolism Regulation by Estrogens and Their Receptors in the Central Nervous System Before and After Menopause. Horm. Metab. Res. 2016, 48, 489–496. [Google Scholar] [CrossRef]
- Khan, M.Z.I.; Uzair, M.; Nazli, A.; Chen, J.Z. An overview on estrogen receptors signaling and its ligands in breast cancer. Eur. J. Med. Chem. 2022, 5, 114658. [Google Scholar] [CrossRef]
- Carter, A.E.; Merriam, S. Menopause. Med. Clin. N. Am. 2023, 107, 199–212. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Ma, X.; Du, L.; Jia, Z.; Cui, X.; Yu, L.; Yang, J.; Xiao, L.; Zhang, B.; et al. Translocation of vaginal microbiota is involved in impairment and protection of uterine health. Nat. Comm. 2021, 12, 4191. [Google Scholar] [CrossRef]
- Minton, K. Immune checkpoint blockade breaches the mucosal firewall to induce gut microbiota translocation. Nat. Rev. Immunol. 2023, 1, 1. [Google Scholar] [CrossRef]
- Spor, A.; Koren, O.; Ley, R. Unravelling the effects of the environment and host genotype on the gut microbiome. Nat. Rev. Microbiol. 2011, 9, 279–290. [Google Scholar] [CrossRef]
- Pigrau, M.; Rodiño-Janeiro, B.K.; Casado-Bedmar, M.; Lobo, B.; Vicario, M.; Santos, J.; Alonso-Cotoner, C. The joint power of sex and stress to modulate brain-gut-microbiota axis and intestinal barrier homeostasis: Implications for irritable bowel syndrome. Neurogastroenterol. Motil. 2016, 28, 463–486. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Priya, S.; Burns, M.B.; Ward, T.; Mars, R.A.T.; Adamowicz, B.; Lock, E.F.; Kashyap, P.C.; Knights, D.; Blekhman, R. Identification of shared and disease-specific host gene-microbiome associations across human diseases using multi-omic integration. Nat. Microbiol. 2022, 7, 780–795. [Google Scholar] [CrossRef]
- Thriene, K.; Michels, K.B. Human Gut Microbiota Plasticity throughout the Life Course. Int. J. Environ. Res. Public Health 2023, 20, 1463. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen–gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Clabaut, M.; Suet, A.; Racine, P.J.; Tahrioui, A.; Verdon, J.; Barreau, M.; Maillot, O.; Le Tirant, A.; Karsybayeva, M.; Kremser, C.; et al. Effect of 17β-estradiol on a human vaginal Lactobacillus crispatus strain. Sci. Rep. 2021, 11, 7133. [Google Scholar] [CrossRef]
- Raftogianis, R.; Creveling, C.; Weinshilboum, R.; Weisz, J. Estrogen metabolism by conjugation. JNCI Monogr. 2000, 27, 113–124. [Google Scholar] [CrossRef]
- Elmassry, M.M.; Kim, S.; Busby, B. Predicting drug-metagenome interactions: Variation in the microbial β-glucuronidase level in the human gut metagenomes. PLoS ONE 2021, 16, e0244876. [Google Scholar] [CrossRef]
- Neuman, H.; Debelius, J.W.; Knight, R.; Koren, O. Microbial endocrinology: The interplay between the microbiota and the endocrine system. FEMS Microbiol. Rev. 2015, 39, 509–521. [Google Scholar] [CrossRef]
- Hur, H.J.; Jeong, Y.H.; Lee, S.H.; Sung, M.J. Quercitrin ameliorates hyperlipidemia and hepatic steatosis in ovariectomized mice. Life 2020, 10, 243. [Google Scholar] [CrossRef]
- Plottel, C.S.; Blaser, M.J. Microbiome and malignancy. Cell Host Microb. 2011, 10, 324–335. [Google Scholar] [CrossRef]
- Kitawaki, J.; Kado, N.; Ishihara, H.; Koshiba, H.; Kitaoka, Y.; Honjo, H. Endometriosis: The pathophysiology as an estrogen-dependent disease. J. Steroid Biochem. Mol. Biol. 2002, 83, 149–155. [Google Scholar] [CrossRef]
- Mori, T.; Kitawaki, J. Role of estrogen and estrogen-related factors in endometriosis. In Immunology of Endometriosis; Chapter 7; Academic Press: Cambridge, MA, USA, 2022; Volume 7, pp. 105–119. ISBN 9780128206614. [Google Scholar] [CrossRef]
- Oliva, M.M.; Gambioli, R.; Forte, G.; Porcaro, G.; Aragona, C.; Unfer, V. Unopposed estrogens: Current and future perspectives. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 2975–2989. [Google Scholar]
- Hu, S.; Ding, Q.; Zhang, W.; Kang, M.; Ma, J.; Zhao, L. Gut microbial beta-glucuronidase: A vital regulator in female estrogen metabolism. Gut Microbes 2023, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Sharma, D. The Microbiome-Estrogen Connection and Breast Cancer Risk. Cells 2019, 8, 1642. [Google Scholar] [CrossRef] [PubMed]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef] [PubMed]
- Salliss, M.E.; Farland, L.V.; Mahnert, N.D.; Herbst-Kralovetz, M.M. The role of gut and genital microbiota and the estrobolome in endometriosis, infertility and chronic pelvic pain. Hum. Reprod. Updat. 2021, 28, 92–131. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Tian, G.; Liu, J.; Cao, L. The gut microbiota and endometriosis: From pathogenesis to diagnosis and treatment. Front. Cell Infect. Microbiol. 2022, 12, 1069557. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.; Martín, V.; Jiménez, E.; Mader, I.; Rodríguez, J.M.; Fernández, L. Lactobacilli and bifidobacteria in human breast milk: Influence of antibiotherapy and other host and clinical factors. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Cárdenas, N.; Ocaña, S.; Marín, M.; Arroyo, R.; Beltrán, D.; Badiola, C.; Fernández, L.; Rodríguez, J.M. Rectal and Vaginal Eradication of Streptococcus agalactiae (GBS) in Pregnant Women by Using Lactobacillus salivarius CECT 9145, A Target-specific Probiotic Strain. Nutrients 2019, 11, 810. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Roman, J.M.; Issaq, H.J.; Keefer, L.K.; Veenstra, T.D.; Ziegler, R.G. Quantitative measurement of endogenous estrogens and estrogen metabolites in human serum by liquid Chromatography–Tandem mass spectrometry. Anal. Chem. 2007, 79, 7813–7821. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Xu, X.; Tian, K.; Zhou, D.; Meng, F.; Huo, H. Genomics analysis of the steroid estrogen-degrading bacterium Serratia nematodiphila DH-S01. Biotechnol. Biotechnol. Equip. 2020, 34, 430–440. [Google Scholar] [CrossRef]
- Qiu, Q.; Wang, P.; Kang, H.; Wang, Y.; Tian, K.; Huo, H. Genomic analysis of a new estrogen-degrading bacterial strain, Acinetobacter sp. DSSKY-A-001. Int. J. Gen. 2019, 2019, 2804134. [Google Scholar] [CrossRef]
- Baele, M.; Baele, P.; Vaneechoutte, M.; Storms, V.; Butaye, P.; Devriese, L.A.; Verschraegen, G.; Gillis, M.; Haesebrouck, F. Application of tRNA intergenic spacer PCR for identification of Enterococcus species. J. Clin. Microbiol. 2000, 38, 4201–4207. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Wu, J.; Chen, J. The role of gut microbial β-glucuronidase in estrogen reactivation and breast cancer. Front. Cell Dev. Biol. 2021, 9, 631552. [Google Scholar] [CrossRef] [PubMed]
- Szentirmai, A. Microbial physiology of sidechain degradation of sterols. J. Ind. Microbiol. 1990, 6, 101–115. [Google Scholar] [CrossRef]
- Ge, H.; Yang, L.; Li, B.; Feng, Y.; Wang, S.; Zheng, Y.; Feng, L.; Liu, Y.; Du, Z.; Zhang, L. A comparative study on the biodegradation of 17β-estradiol by Candida utilis CU-2 and Lactobacillus casei LC-1. Front. Energy Res. 2021, 9, 661850. [Google Scholar] [CrossRef]
- Bragin, E.Y.; Shtratnikova, V.Y.; Dovbnya, D.V.; Schelkunov, M.I.; Pekov, Y.A.; Malakho, S.G.; Egorova, O.V.; Ivashina, T.V.; Sokolov, S.L.; Ashapkin, V.V.; et al. Comparative analysis of genes encoding key steroid core oxidation enzymes in fast-growing Mycobacterium spp. strains. J. Steroid Biochem. Mol. Biol. 2013, 138, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Fujisawa, S.; Nakai, S.; Hosomi, M. Biodegradation of natural and synthetic estrogens by nitrifying activated sludge and ammonia-oxidizing bacterium Nitrosomonas europaea. Water Res. 2004, 38, 2323–2330. [Google Scholar] [CrossRef]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef]
- Xiong, W.; Yin, C.; Wang, Y.; Lin, S.; Deng, Z.; Liang, R. Characterization of an efficient estrogen-degrading bacterium Stenotrophomonas maltophilia SJTH1 in saline-, alkaline-, heavy metal-contained environments or solid soil and identification of four 17β-estradiol-oxidizing dehydrogenases. J. Hazard. Mat. 2020, 385, 121616. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, D.; Liang, R. Isolation and characterization of an estrogen-degrading Pseudomonas putida strain SJTE-1. 3 Biotech 2019, 9, 61. [Google Scholar] [CrossRef]
- Yu, C.P.; Roh, H.; Chu, K.H. 17β-Estradiol-degrading bacteria isolated from activated sludge. Environ. Sci. Technol. 2007, 41, 486–492. [Google Scholar] [CrossRef]
- Chen, Y.L.; Yu, C.P.; Lee, T.H.; Goh, K.S.; Chu, K.H.; Wang, P.H.; Ismail, W.; Shih, C.J.; Chiang, Y.R. Biochemical mechanisms and catabolic enzymes involved in bacterial estrogen degradation pathways. Cell Chem. Biol. 2017, 24, 712–724. [Google Scholar] [CrossRef]
- Bilal, M.; Barceló, D.; Iqbal, H.M. Occurrence, environmental fate, ecological issues, and redefining of endocrine disruptive estrogens in water resources. Sci. Total Environ. 2021, 800, 149635. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Raquet, S.C.G. Occurrence, fate and biodegradation of estrogens in sewage and manure. Appl. Microbiol. Biotechnol. 2010, 86, 1671–1692. [Google Scholar] [CrossRef]
- Wieacker, P. Genetic Aspects of premature ovarian failure. J. Reprod. Med. Endocrinol. 2009, 6, 17–18. [Google Scholar]
- Liang, J.; Shang, Y. Estrogen and cancer. Ann. Rev. Physiol. 2013, 75, 225–240. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yang, S.; Fang, Z.; Gurates, B.; Tamura, M.; Sebastian, S. Estrogen production and metabolism in endometriosis. Ann. N. Y. Acad. Sci. 2002, 955, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Van Der Veer, C.; Hertzberger, R.Y.; Bruisten, S.M.; Tytgat, H.L.; Swanenburg, J.; Angelino-Bart, A.; Schuren, F.; Molenaar, D.; Reid, G.; de Vries, H.; et al. Comparative genomics of human Lactobacillus crispatus isolates reveals genes for glycosylation and glycogen degradation: Implications for in vivo dominance of the vaginal microbiota. Microbiome 2019, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Sun, H.; Huang, S.; Feng, X.; Jiang, T.; Li, C. Structure-guided engineering of the substrate specificity of a fungal β-glucuronidase toward triterpenoid saponins. J. Biol. Chem. 2018, 293, 433–443. [Google Scholar] [CrossRef]
- Coulon, S.; Chemardin, P.; Gueguen, Y.; Arnaud, A.; Galzy, P. Purification and characterization of an intracellular β-glucosidase from Lactobacillus casei ATCC 393. Appl. Biochem. Biotechnol. 1998, 74, 105–114. [Google Scholar] [CrossRef]
- Dabek, M.; McCrae, S.I.; Stevens, V.J.; Duncan, S.H.; Louis, P. Distribution of β-glucosidase and β-glucuronidase activity and of β-glucuronidase gene gus in human colonic bacteria. FEMS Microbiol. Ecol. 2008, 66, 487–495. [Google Scholar] [CrossRef]
- Muccee, F.; Ghazanfar, S.; Ajmal, W.; Al-Zahrani, M. In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients. Genes 2022, 13, 1545. [Google Scholar] [CrossRef] [PubMed]
- Mroczynska, M.; Libudzisz, Z. Beta-glucuronidase and beta-glucosidase activity of Lactobacillus and Enterococcus isolated from human feces. Pol. J. Microbiol. 2010, 59, 265–269. [Google Scholar] [CrossRef]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.; Muñoz, R. Bioactivation of phytoestrogens: Intestinal bacteria and health. Crit. Rev. Food Sci. Nut. 2016, 56, 1826–1843. [Google Scholar] [CrossRef]
- Beck, A.P.; Li, H.; Ervin, S.M.; Redinbo, M.R.; Mani, S. Inhibition of Microbial Beta-Glucuronidase Does Not Prevent Breast Carcinogenesis in the Polyoma Middle T Mouse. bioRxiv 2019, 746602. [Google Scholar] [CrossRef]
- Nakamura, J.; Kubota, Y.; Miyaoka, M.; Saitoh, T.; Mizuno, F.; Benno, Y. Comparison of four microbial enzymes in Clostridia and Bacteroides isolated from human feces. Microbiol. Immunol. 2002, 46, 487–490. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Strain Code | Origin |
|---|---|---|
| Ligilactobacillus salivarius | ES27 | Vagina |
| L. salivarius | ES34 | Vagina |
| L. salivarius | ES40 | Human milk |
| L. salivarius | ES41 | Human milk |
| L. salivarius | ES42 | Human milk |
| L. salivarius | ES43 | Human milk |
| Estrogen | RT (min) | Q1 | CE (V) | Q3 | LOD (mg/L) | LOQ (mg/L) |
|---|---|---|---|---|---|---|
| Estrone | 7.30 | 271.2 | −13 | 253.05 | 0.03 | 0.10 |
| −25 | 133.10 | |||||
| 17β-Estradiol | 6.70 | 273.0 | −27 | 107.10 | 0.86 | 2.88 |
| −18 | 135.00 | |||||
| Estriol | 3.10 | 289.1 | −25 | 107.00 | 0.03 | 0.09 |
| −14 | 253.05 | |||||
| 2-Hydroxyestrone-3-methyl ether | 8.00 | 301.0 | −18 | 186.95 | 0.01 | 0.02 |
| −19 | 240.20 | |||||
| 2-Methoxyestradiol | 7.30 | 303.2 | −24 | 137.00 | 0.01 | 0.04 |
| −19 | 135.20 | |||||
| 4-Methoxy-17β-estradiol | 6.98 | 303.2 | −25 | 137.05 | 0.06 | 0.18 |
| −20 | 135.20 | |||||
| 4-Methoxyestrone | 7.60 | 301.0 | −14 | 283.20 | 0.02 | 0.07 |
| −23 | 163.00 | |||||
| 16ἀ-Hydroxyestrone | 4.40 | 287.0 | −15 | 251.15 | 0.01 | 0.04 |
| −20 | 199.10 | |||||
| 2-Hydroxyestrone | 6.00 | 287.0 | −13 | 269.25 | 0.02 | 0.06 |
| −14 | 175.05 |
| Enzyme (EC Number) | Gene (ID) | Primer Sequence (5′-3′) | Annealing Temperature (°C) | Amplicon Size (pb) | References |
|---|---|---|---|---|---|
| α-glucosidase (3.2.1.20) | ygjK (BQ1177_RS04000) | F: GGACGATGTGGAGGAGCTAA | 55.0 | 205 | |
| R: CCCAAATGCGGAACCATGAT | |||||
| β-glucosidase (3.2.1.86) | bglH (BQ1177_RS09625) | F: AATGGCCTTGGTGCAAAAGA | 55.0 | 184 | |
| R: AATTTCGCCGGAACTAGCAC | |||||
| α-glucuronidase (3.2.1.139) | H0A38_RS06600 | F: TACGCTCGCTTTCTTGCTTC | 55.0 | 242 | |
| R: TGTTGCCACCATGAAGACAC | |||||
| β-glucuronidase (3.2.1.31) | gusA, uidA (FC33_RS02420) | F: AATTCACCGCCCCGTTAAAC | 55.0 | 194 | |
| R: ATGAGTTTCCCTGAACCCGT | |||||
| Glycosyltransferase (2.4.-.-) | gtf3/csbB/epsF (BQ1177_RS07995) | F: AAGAAAATGGTGGGTTGTCTGA | 55.0 | 100 | |
| R: TGACTTTCAACGTAATCATCGGA | |||||
| Sugar transferase (2.7.8.-) | epsL (BQ1177_RS00420) | F: CAAGCTCGGAGACATGAAGT | 55.0 | 204 | |
| R: TGTTGCTGAACCTTCTTCTGA | |||||
| Bifunctional acetaldehyde- CoA/alcohol dehydrogenase (1.2.1.10/1.1.1.1) | adhE (BQ1177_RS09345) | F: GCATCTGACTTCACACGTCC | 55.0 | 219 | [50] |
| R: TCCAAATTCCCCACCAGTCT | |||||
| 3-hydroxyacyl-CoA dehydrogenase (1.1.1.35) | FadB/echA (BQ1177_RS04965) | F: AGGGGTTGCAGATCCAATGA | 55.0 | 205 | [50] |
| R: GCCAGCTTCAATGCCGTAAT | |||||
| Acyl-CoA thioesterase (3.1.2.-) | paaI (BQ1177_RS04090) | F: ACTAGCCATCGTGTTTTAGCT | 55.0 | 207 | [50] |
| R: GCCACTAACGTATGATTCGATAC | |||||
| Short-chain dehydrogenase(1.3.-.-) | DltE (BQ1177_RS00925) | F: GGAAGAAGTGGGACCTGTCT | 55.0 | 172 | [51] |
| R: ACCTACTTGTCTTTCGGCCA |
| Category | Enzyme | Ligilactobacillus salivarius Strain | |||||
|---|---|---|---|---|---|---|---|
| ES27 | ES34 | ES40 | ES41 | ES42 | ES43 | ||
| Conjugation/deconjugation-related enzymes | α-glucosidase | + | + | + | + | + | + |
| β-glucosidase | + | + | + | + | + | + | |
| α-glucuronidase | − | − | − | − | − | − | |
| β-glucuronidase | − | − | − | − | − | − | |
| Glycosyltransferase | + | + | + | + | + | + | |
| Sugar transferase | + | + | + | + | + | + | |
| Steroid core destructive enzyme | Bifunctional acetaldehyde-CoA/alcohol dehydrogenase | + | + | + | + | + | + |
| 3-hydroxyacyl-CoA dehydrogenase | + | + | + | + | + | + | |
| Steroid side-chain oxidase | Acyl-CoA thioesterase | + | + | + | + | + | + |
| Short chain dehydrogenase | + | + | + | + | + | + | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aragón, A.; Jurado, R.; Jara, J.; Rodríguez, J.M.; Orgaz, B. Investigating the Metabolism of Estrogens in Ligilactobacillus salivarius Strains Isolated from Human Milk and Vaginal Microbiota. Nutrients 2024, 16, 861. https://doi.org/10.3390/nu16060861
Aragón A, Jurado R, Jara J, Rodríguez JM, Orgaz B. Investigating the Metabolism of Estrogens in Ligilactobacillus salivarius Strains Isolated from Human Milk and Vaginal Microbiota. Nutrients. 2024; 16(6):861. https://doi.org/10.3390/nu16060861
Chicago/Turabian StyleAragón, Alberto, Rubén Jurado, Josué Jara, Juan Miguel Rodríguez, and Belén Orgaz. 2024. "Investigating the Metabolism of Estrogens in Ligilactobacillus salivarius Strains Isolated from Human Milk and Vaginal Microbiota" Nutrients 16, no. 6: 861. https://doi.org/10.3390/nu16060861