
Structure, Biological Functions, Separation, Properties, and Potential Applications of Milk Fat Globule Membrane (MFGM): A Review
 , , and
, , and
Abstract
:1. Introduction
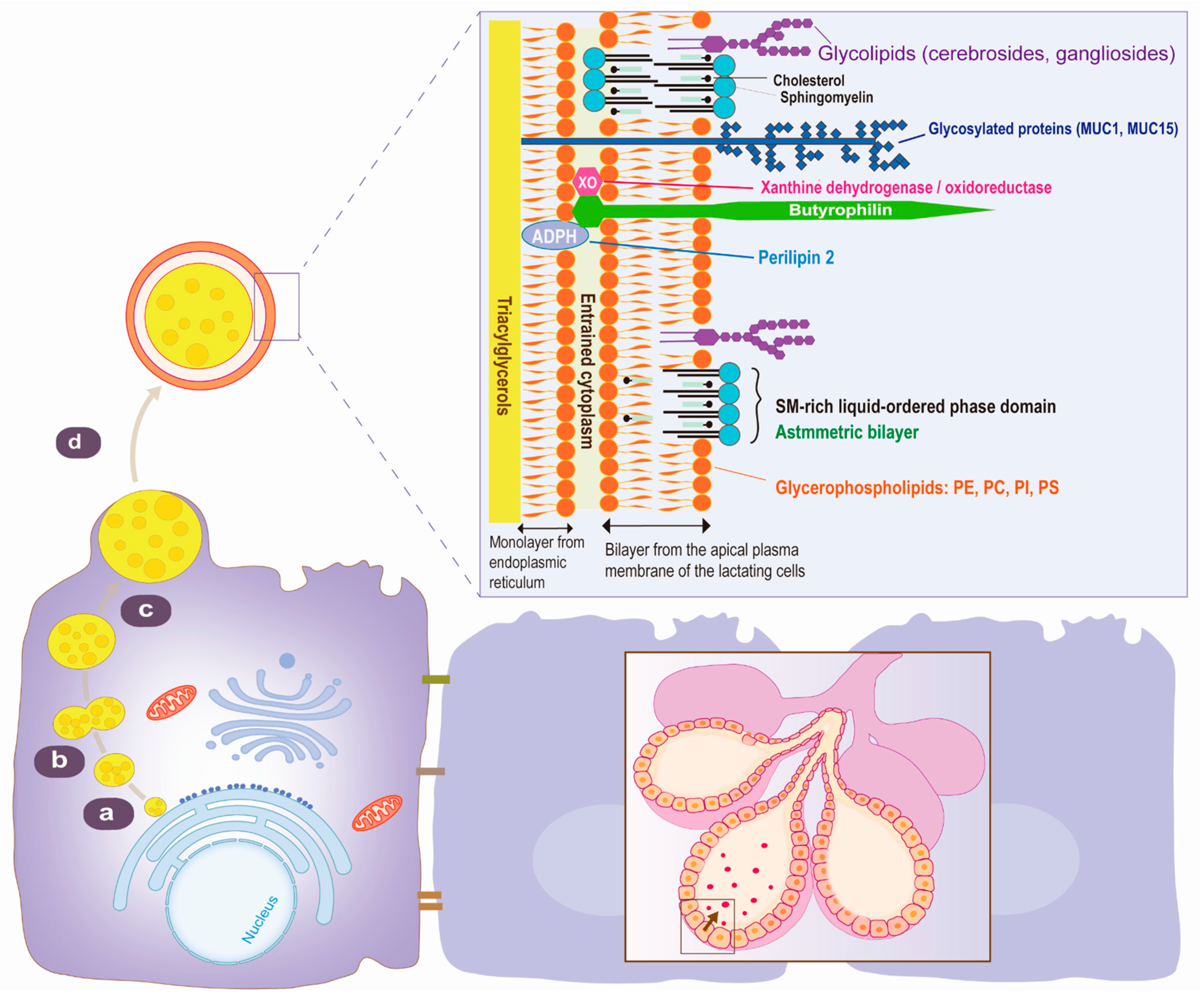
2. Structure and Composition of MFGM
2.1. Structure
2.2. Components
2.3. MFGM Protein Composition
2.4. MFGM Lipid Composition
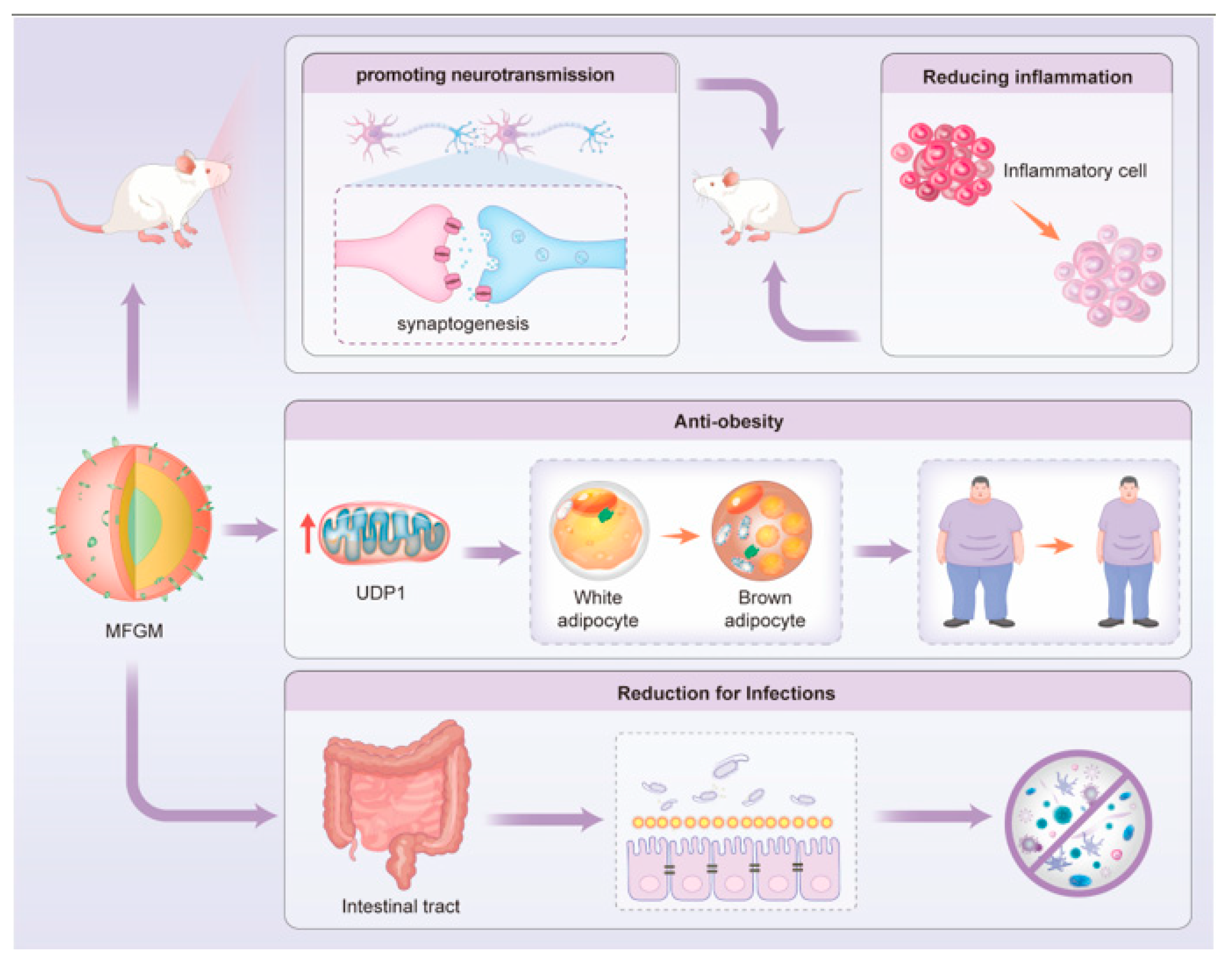
3. Health Benefits of MFGM as a Bioactive Ingredient
3.1. Improvement of the Neurodevelopmental Profile
3.2. Anti-Inflammatory Activity
3.3. Reduction in Infection Susceptibility
3.4. Other Biological Effects of MFGM
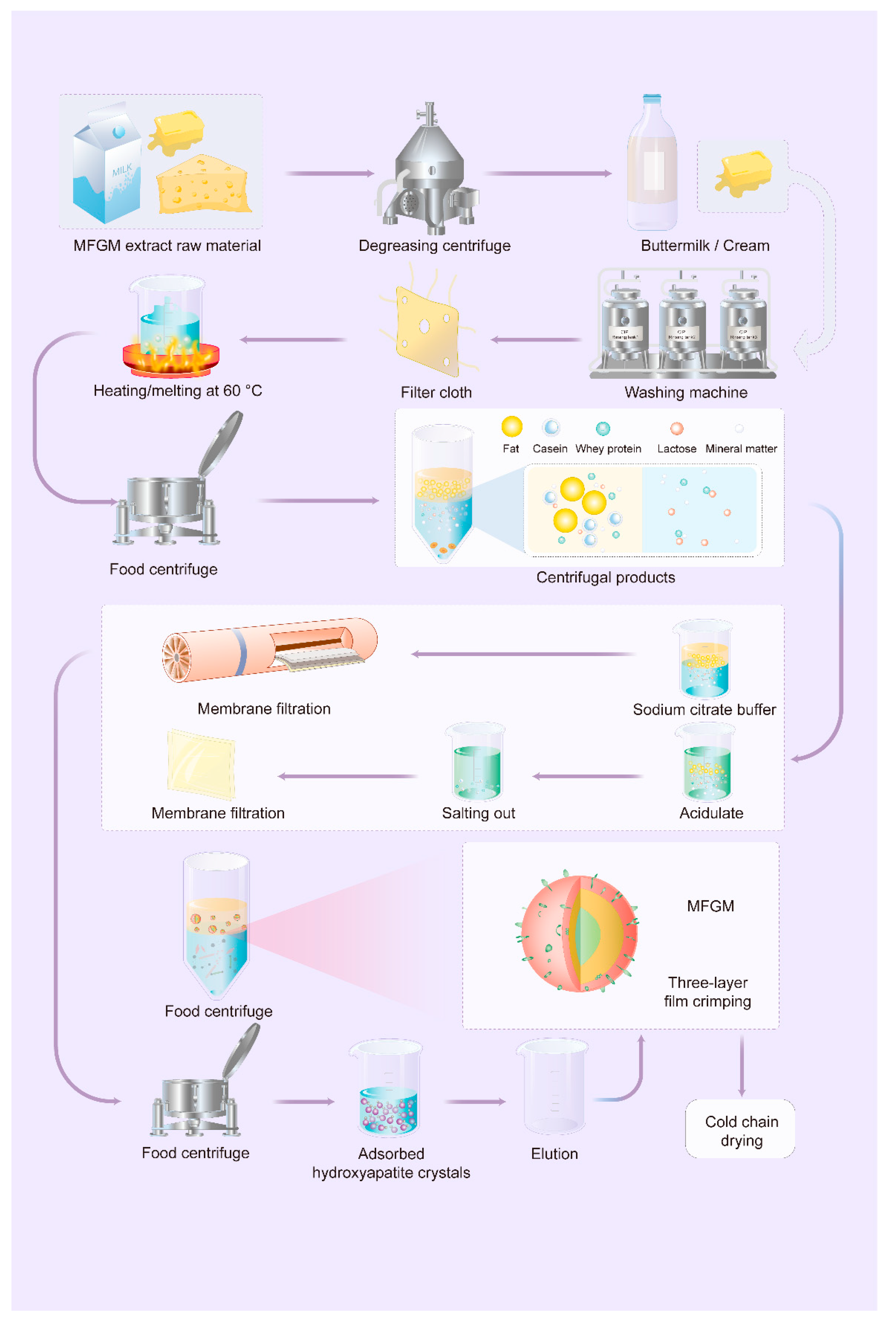
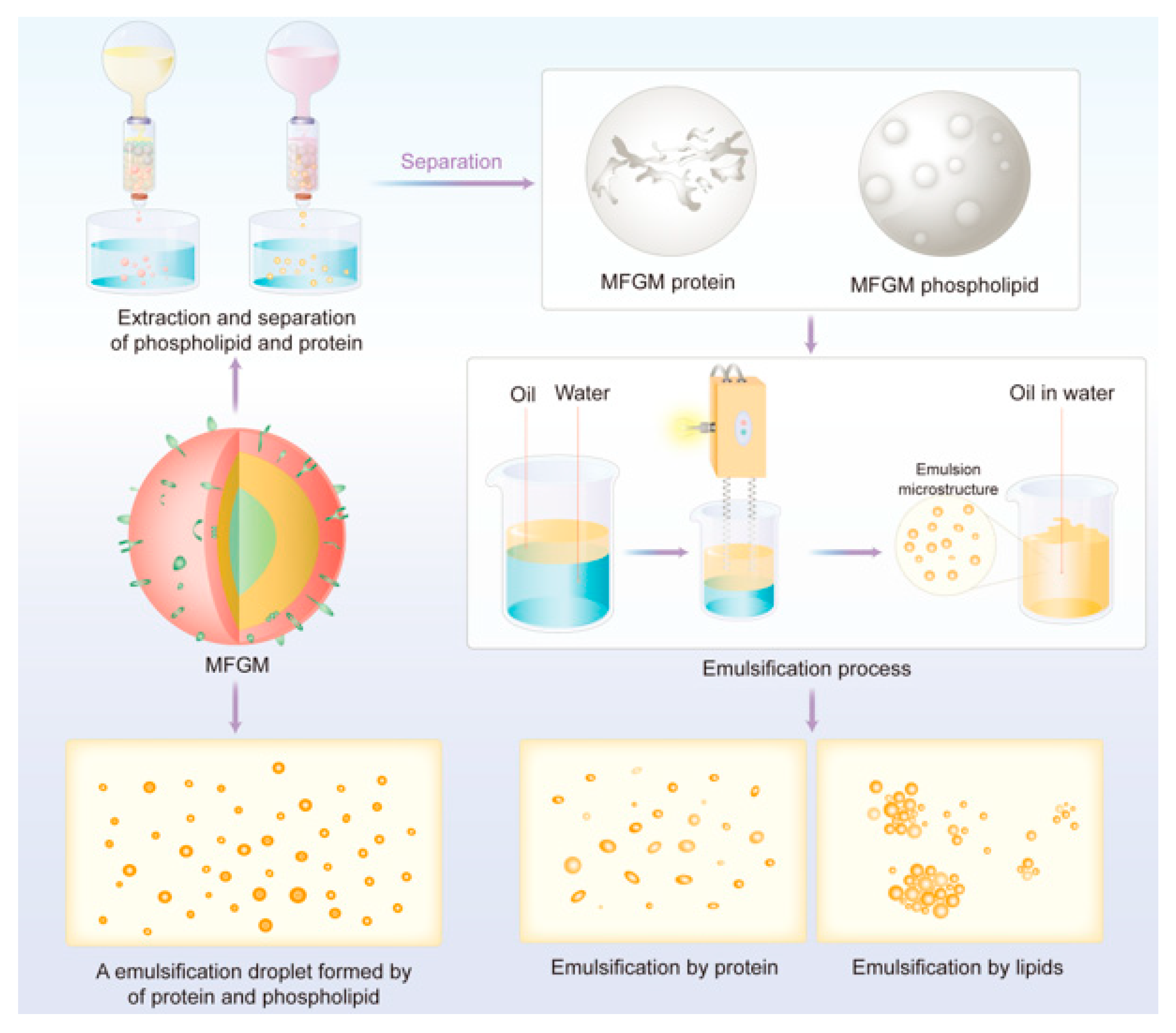
4. Separation and Purification of MFGM
4.1. Separation of Milk Fat Globule
4.2. Collection of MFGM
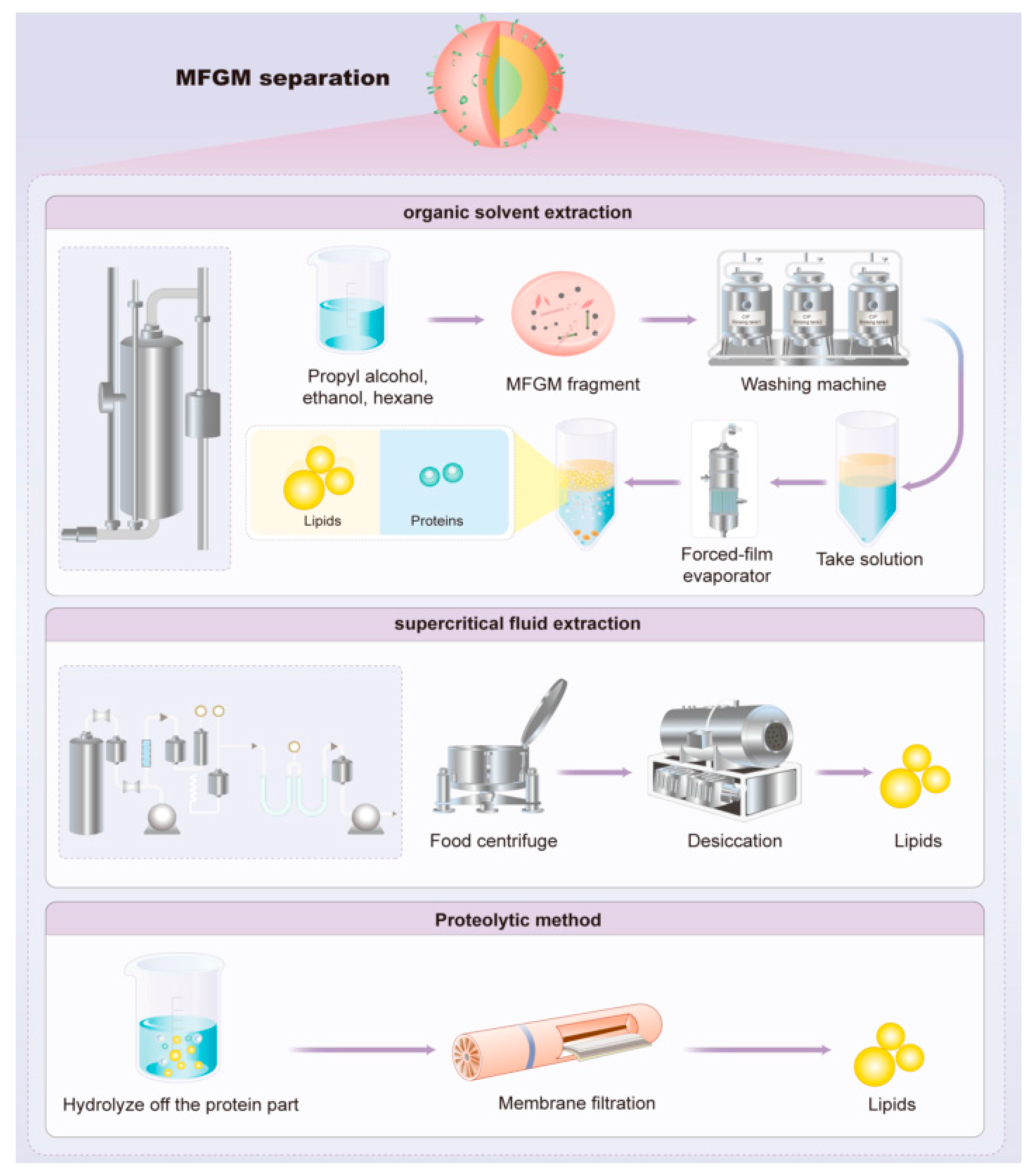
4.3. Extraction of Lipid and Protein from MFGM
- (1)
- Microfiltration and supercritical fluid extraction (SFE) were used to produce buttermilk-derived fractions with increased concentrations of polar MFGM lipids. A ceramic tubular membrane with a 0.8 μm pore size was employed to concentrate polar MFGM lipids, following a 2n factorial design. Subsequently, an SFE process utilizing supercritical carbon dioxide selectively removed non-polar lipid material from the microfiltered buttermilk product, resulting in a significant reduction in the non-polar lipid concentration along with a significant increase in the concentration of polar lipids from MFGM [71,72].
- (2)
- Polar lipid concentrates can be obtained from milk processing by-products using organic solvent extraction. A mixture of hydrophobic and hydrophilic solvents (such as propanol or methanol) can effectively extract amphiphilic polar lipids, while hydrophobic solvents (such as hexane or chloroform) are required to extract triglycerides [73,74].
5. Interfacial Properties of MFGM and Its Applications in Food Industry
5.1. Surface Activity of MFGM
5.1.1. Emulsifying Properties of MFGM
5.1.2. Effects of MFGM Proteins
5.1.3. Effects of MFGM Lipids
5.2. Applications of MFGM
5.2.1. Emulsifier
5.2.2. Infant Formula Milk Powder
5.2.3. Other Applications
6. Encapsulation Systems with Lactoferrin as a Functional Ingredient
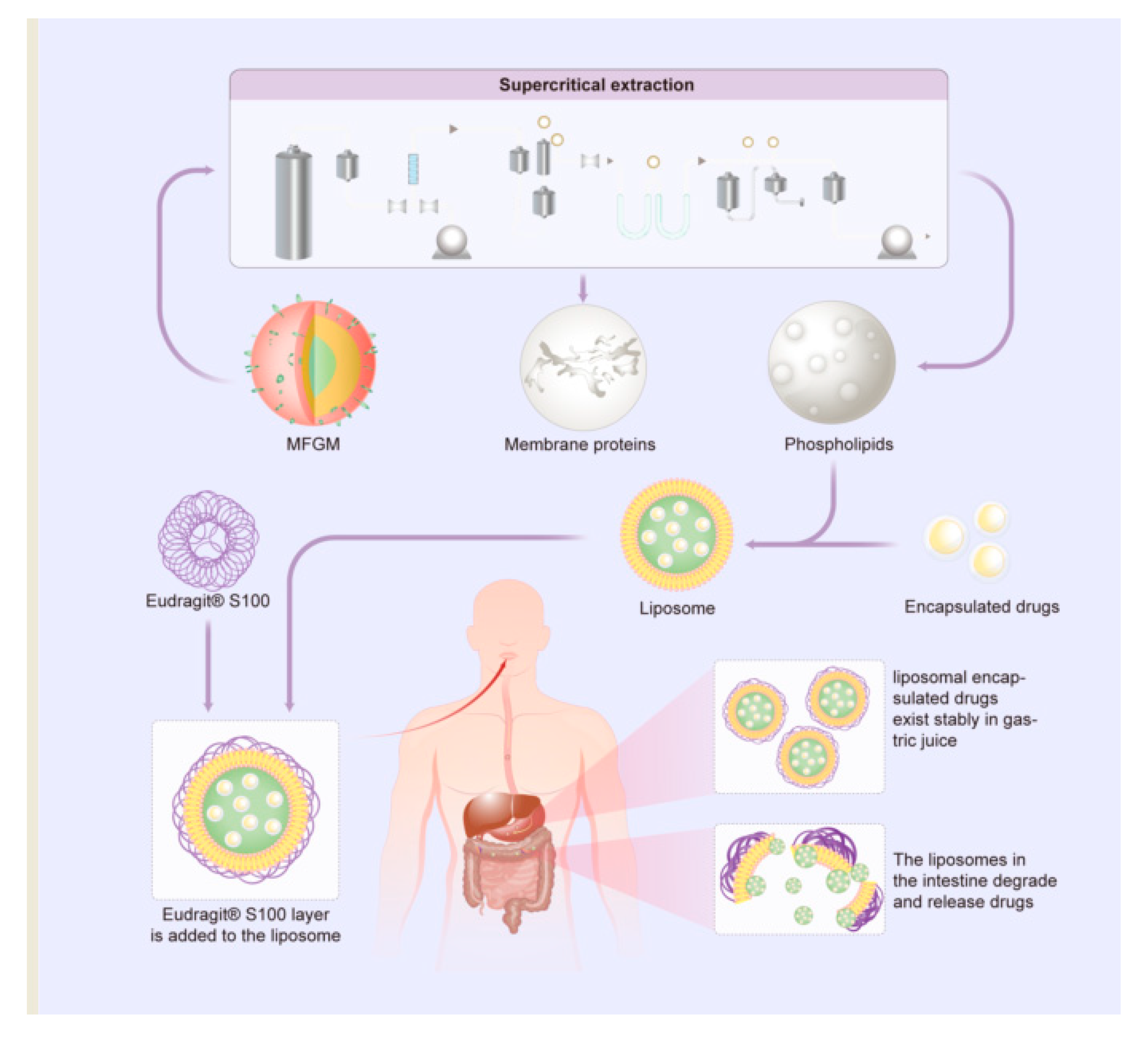
6.1. MFGM Liposomes
6.2. Advantages of MFMG Liposomes as Encapsulation Materials
6.3. Application of MFGM Liposomes in Medicine
6.4. Application of MFGM Liposomes in Food Industry
7. Conclusions and Future Trends
Funding
Conflicts of Interest
Abbreviations
| Milk fat globule membrane | MFGM |
| Mucin 1 | MUC1 |
| Xanthine dehydrogenase/oxidase | XDH/XO |
| Butyrophilin | BTN |
| Cluster of differentiation 36 | CD36 |
| Periodic acid–Schiff 6/7 | PAS 6/7 |
| Periodic acid–Schiff III, PAS III | also called MUC15 |
| Adipophilin | ADPH |
| Fatty acid-binding protein | FABP |
| Glycosylation-dependent cell adhesion molecule 1 | GlyCAM1 |
| Phosphatidylethanolamine | PE |
| Phosphatidylcholine | PC |
| Phosphatidylserine | PS |
| Phosphatidylinositol | PI |
| Lysophosphatidylcholine | LPC |
| Sphingomyelin | SM |
| Phospholipids | PLs |
| White adipose tissue | WAT |
| High-fat diet | HFD |
| Supercritical fluid extraction | SFE |
| High-pressure homogenisation | HPH |
References
- Lopez, C.; Menard, O. Human milk fat globules: Polar lipid composition and in situ structural investigations revealing the heterogeneous distribution of proteins and the lateral segregation of sphingomyelin in the biological membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef]
- Fong, B.Y.; Norris, C.S.; MacGibbon, A.K.H. Protein and lipid composition of bovine milk-fat-globule membrane. Int. Dairy J. 2007, 17, 275–288. [Google Scholar] [CrossRef]
- Nii, T.; Ishii, F. Dialkylphosphatidylcholine and egg yolk lecithin for emulsification of various triglycerides. Colloids Surf. B Biointerfaces 2005, 41, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ma, S.; Liu, Y.; Jia, Z.; Li, X.; Liu, L.; Ma, Q.; Kouame, K.J.E.-P.; Li, C.; Leng, Y.; et al. Changes in interfacial composition and structure of milk fat globules are crucial regulating lipid digestion in simulated in-vitro infant gastrointestinal digestion. Food Hydrocoll. 2023, 134, 108003. [Google Scholar] [CrossRef]
- Jukkola, A.; Partanen, R.; Xiang, W.; Heino, A.; Rojas, O.J. Food emulsifiers based on milk fat globule membranes and their interactions with calcium and casein phosphoproteins. Food Hydrocoll. 2019, 94, 30–37. [Google Scholar] [CrossRef]
- Phan, T.T.Q.; Moens, K.; Le, T.T.; Van der Meeren, P.; Dewettinck, K. Potential of milk fat globule membrane enriched materials to improve the whipping properties of recombined cream. Int. Dairy J. 2014, 39, 16–23. [Google Scholar] [CrossRef]
- Zhao, J.; Yi, W.; Liu, B.; Dai, Y.; Jiang, T.; Chen, S.; Wang, J.; Feng, B.; Qiao, W.; Liu, Y.; et al. MFGM components promote gut Bifidobacterium growth in infant and in vitro. Eur. J. Nutr. 2022, 61, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, Y.; Ye, H.; Feng, C.; Han, D.; Tao, S.; Pi, Y.; Zhao, J.; Chen, L.; Wang, J. Dietary milk fat globule membrane supplementation during late gestation increased the growth of neonatal piglets by improving their plasma parameters, intestinal barriers, and fecal microbiota. RSC Adv. 2020, 10, 16987–16998. [Google Scholar] [CrossRef] [PubMed]
- Chitchumroonchokchai, C.; Riedl, K.; García-Cano, I.; Chaves, F.; Walsh, K.R.; Jimenez-Flores, R.; Failla, M.L. Efficient in vitro digestion of lipids and proteins in bovine milk fat globule membrane ingredient (MFGMi) and whey-casein infant formula with added MFGMi. J. Dairy Sci. 2023, 106, 3086–3097. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, X.; Hussain, M.; Li, X.; Liu, L.; Liu, Y.; Ma, S.; Kouame, K.J.E.-P.; Li, C.; Leng, Y.; et al. Influence of milk fat globule membrane and milk protein concentrate treated by ultrasound on the structural and emulsifying stability of mimicking human fat emulsions. Ultrason. Sonochem. 2022, 82, 105881. [Google Scholar] [CrossRef]
- Peng, F.; He, S.; Yi, H.; Li, Q.; Xu, W.; Wang, R.; Ma, Y. Physical, textural, and rheological properties of whipped cream affected by milk fat globule membrane protein. Int. J. Food Prop. 2018, 21, 1190–1202. [Google Scholar] [CrossRef]
- Tang, H.; He, S.; Peng, F.; Wang, R.; Li, Q.; Ma, Y. The effects of milk fat globule membrane and its individual components on dough properties and bread quality. RSC Adv. 2016, 6, 102617–102625. [Google Scholar] [CrossRef]
- McManaman, J.L.; Russell, T.D.; Schaack, J.; Orlicky, D.J.; Robenek, H. Molecular determinants of milk lipid secretion. J. Mammary Gland. Biol. Neoplasia 2007, 12, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Argov-Argaman, N. Symposium review: Milk fat globule size: Practical implications and metabolic regulation. J. Dairy Sci. 2019, 102, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Heid, H.W.; Keenan, T.W. Intracellular origin and secretion of milk fat globules. Eur. J. Cell Biol. 2005, 84, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Guerin, J.; Burgain, J.; Gomand, F.; Scher, J.; Gaiani, C. Milk fat globule membrane glycoproteins: Valuable ingredients for lactic acid bacteria encapsulation? Crit. Rev. Food Sci. Nutr. 2019, 59, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Dewettinck, K.; Rombaut, R.; Thienpont, N.; Le, T.T.; Messens, K.; Van Camp, J. Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Kiełczewska, K.; Ambroziak, K.; Krzykowska, D.; Aljewicz, M. The effect of high-pressure homogenisation on the size of milk fat globules and MFGM composition in sweet buttermilk and milk. Int. Dairy J. 2021, 113, 104898. [Google Scholar] [CrossRef]
- Priyanka, M.; Sabine, D.; Aman, P.; Koen, D.; Christophe, B. The interfacial properties of various milk fat globule membrane components using Langmuir isotherms. Food Biosci. 2017, 20, 96–103. [Google Scholar] [CrossRef]
- Thum, C.; Roy, N.C.; Everett, D.W.; McNabb, W.C. Variation in milk fat globule size and composition: A source of bioactives for human health. Crit. Rev. Food Sci. Nutr. 2023, 63, 87–113. [Google Scholar] [CrossRef]
- Abeler-Dörner, L.; Swamy, M.; Williams, G.; Hayday, A.C.; Bas, A. Butyrophilins: An emerging family of immune regulators. Trends Immunol. 2012, 33, 34–41. [Google Scholar] [CrossRef]
- Zhang, L.; Yan, D.; Roy, M.C.; Huang, J.; Zhou, P. Variation in both proteome and N-glycoproteome of goat MFGM over lactation. J. Food Compos. Anal. 2022, 111, 104635. [Google Scholar] [CrossRef]
- Haddadian, Z.; Eyres, G.T.; Carne, A.; Everett, D.W.; Bremer, P. Impact of different milk fat globule membrane preparations on protein composition, xanthine oxidase activity, and redox potential. Int. Dairy J. 2017, 64, 14–21. [Google Scholar] [CrossRef]
- Valk-Weeber, R.L.; Nichols, K.; Dijkhuizen, L.; Bijl, E.; van Leeuwen, S.S. Variations in N-linked glycosylation of glycosylation-dependent cell adhesion molecule 1 (GlyCAM-1) whey protein: Intercow differences and dietary effects. J. Dairy Sci. 2021, 104, 5056–5068. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, M.; Yoshida, T.; Tamura, K.; Dbouk, M.; Canto, M.I.; Burkhart, R.; He, J.; Roberts, N.J.; Klein, A.P.; Goggins, M. Endoplasmic stress-inducing variants in carboxyl ester lipase and pancreatic cancer risk. Pancreatology 2022, 22, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Sudarshan, M.; Shree, V.D.; Jyothibai, P.; Kumar, B.G. N-Glycoprofiling of immunoglobulin G and lactoperoxidase from sheep milk using LC-MS/MS. Int. Dairy J. 2023, 140, 105582. [Google Scholar] [CrossRef]
- Iglesias-Figueroa, B.F.; Espinoza-Sánchez, E.A.; Siqueiros-Cendón, T.S.; Rascón-Cruz, Q. Lactoferrin as a nutraceutical protein from milk, an overview. Int. Dairy J. 2019, 89, 37–41. [Google Scholar] [CrossRef]
- Kulyar, M.F.-E.; Yao, W.; Ding, Y.; Li, K.; Zhang, L.; Li, A.; Waqas, M.; Huachun, P.; Quan, M.; Zeng, Z.; et al. Bioactive potential of yak’s milk and its products; pathophysiological and molecular role as an immune booster in antibiotic resistance. Food Biosci. 2021, 39, 100838. [Google Scholar] [CrossRef]
- Nafikov, R.; Schoonmaker, J.; Korn, K.; Noack, K.; Garrick, D.; Koehler, K.; Minick-Bormann, J.; Reecy, J.; Spurlock, D.; Beitz, D. Association of polymorphisms in solute carrier family 27, isoform A6 (SLC27A6) and fatty acid-binding protein-3 and fatty acid-binding protein-4 (FABP3 and FABP4) with fatty acid composition of bovine milk. J. Dairy Sci. 2013, 96, 6007–6021. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, S.; Wilson, M.R. The Dual Roles of Clusterin in Extracellular and Intracellular Proteostasis. Trends Biochem. Sci. 2021, 46, 652–660. [Google Scholar] [CrossRef]
- Chong, B.M.; Reigan, P.; Mayle-Combs, K.D.; Orlicky, D.J.; McManaman, J.L. Determinants of adipophilin function in milk lipid formation and secretion. Trends Endocrinol. Metab. 2011, 22, 211–217. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Yang, G.; Xu, K.; Yin, Y.; Brecchia, G.; Yin, J. CD36 favours fat sensing and transport to govern lipid metabolism. Prog. Lipid Res. 2022, 88, 101193. [Google Scholar] [CrossRef]
- Argov, N.; Lemay, D.G.; German, J.B. Milk Fat Globule structure & function: Nanosciece comes to milk production. Trends Food Sci. Technol. 2008, 19, 617–623. [Google Scholar]
- Sun, Y.; Tian, S.; Hussain, M.; Lin, S.; Pan, Y.; Li, X.; Liu, L.; Lu, X.; Li, C.; Leng, Y.; et al. Profiling of phospholipid classes and molecular species in human milk, bovine milk, and goat milk by UHPLC-Q-TOF-MS. Food Res. Int. 2022, 161, 111872. [Google Scholar] [CrossRef]
- Geng, M.; Zhao, F.; Lu, H.; Fang, L.; Wang, J.; Liu, C.; Min, W. Insights into the hippocampus proteome and phosphorylation modification alterations in C57BL/6 revealed the memory improvement mechanisms of a walnut-derived peptide. Food Res. Int. 2022, 156, 111311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zou, X.; Feng, R.; Zhan, X.; Hong, H.; Luo, Y.; Tan, Y. Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane. Nutrients 2023, 15, 534. [Google Scholar] [CrossRef]
- Unno, K.; Taguchi, K.; Hase, T.; Meguro, S.; Nakamura, Y. Coffee Polyphenol, Chlorogenic Acid, Suppresses Brain Aging and Its Effects Are Enhanced by Milk Fat Globule Membrane Components. Int. J. Mol. Sci. 2022, 23, 5832. [Google Scholar] [CrossRef]
- Yu, Z.; Huang, S.; Li, Y.; Niu, Y.; Chen, H.; Wu, J. Milk Fat Globule Membrane Alleviates Short Bowel Syndrome-Associated Liver Injury in Rats through Inhibiting Autophagy and NLRP3 Inflammasome Activation. Front. Nutr. 2022, 9, 758762. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Jacob, A.; Wu, R.; Zhou, M.; Aziz, M.; Wang, P. Milk fat globule--EGF factor VIII ameliorates liver injury after hepatic ischemia-reperfusion. J. Surg. Res. 2013, 180, e37–e46. [Google Scholar] [CrossRef]
- Douëllou, T.; Montel, M.; Sergentet, D.T. Thevenot Sergentet, Invited review: Anti-adhesive properties of bovine oligosaccharides and bovine milk fat globule membrane-associated glycoconjugates against bacterial food enteropathogens. J. Dairy Sci. 2017, 100, 3348–3359. [Google Scholar] [CrossRef] [PubMed]
- Guerin, J.; Soligot, C.; Burgain, J.; Huguet, M.; Francius, G.; El-Kirat-Chatel, S.; Gomand, F.; Lebeer, S.; Le Roux, Y.; Borges, F.; et al. Adhesive interactions between milk fat globule membrane and Lactobacillus rhamnosus GG inhibit bacterial attachment to Caco-2 TC7 intestinal cell. Colloids Surf. B Biointerfaces 2018, 167, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.; Kuhlenschmidt, T.; Kuhlenschmidt, M.; Jiménez-Flores, R.; Donovan, S. Milk fat globule membrane isolated from buttermilk or whey cream and their lipid components inhibit infectivity of rotavirus in vitro. J. Dairy Sci. 2013, 96, 3488–3497. [Google Scholar] [CrossRef] [PubMed]
- Monaco, M.H.; Gross, G.; Donovan, S.M. Whey Protein Lipid Concentrate High in Milk Fat Globule Membrane Components Inhibit Porcine and Human Rotavirus in vitro. Front. Pediatr. 2021, 9, 731005. [Google Scholar] [CrossRef] [PubMed]
- Tellez, A.; Corredig, M.; Guri, A.; Zanabria, R.; Griffiths, M.; Delcenserie, V. Bovine milk fat globule membrane affects virulence expression in Escherichia coli O157:H7. J. Dairy Sci. 2012, 95, 6313–6319. [Google Scholar] [CrossRef] [PubMed]
- Sprong, R.C.; Hulstein, M.F.; Lambers, T.T.; van der Meer, R. Sweet buttermilk intake reduces colonisation and translocation of Listeria monocytogenes in rats by inhibiting mucosal pathogen adherence. Br. J. Nutr. 2012, 108, 2026–2033. [Google Scholar] [CrossRef] [PubMed]
- Bruggencate, S.J.T.; Frederiksen, P.D.; Pedersen, S.M.; Floris-Vollenbroek, E.G.; de Bos, E.L.-V.; van Hoffen, E.; Wejse, P.L. Dietary Milk-Fat-Globule Membrane Affects Resistance to Diarrheagenic Escherichia coli in Healthy Adults in a Randomized, Placebo-Controlled, Double-Blind Study. J. Nutr. 2016, 146, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.R.; Jimenez-Flores, R.; Ward, R.E.; Cambell, J.; Young, M.J.; Nemere, I.; Hintze, K.J. Dietary milk fat globule membrane reduces the incidence of aberrant crypt foci in Fischer-344 rats. J. Agric. Food Chem. 2010, 58, 2157–2163. [Google Scholar] [CrossRef]
- Li, T.; Gao, J.; Du, M.; Song, J.; Mao, X. Milk Fat Globule Membrane Attenuates High-Fat Diet-Induced Obesity by Inhibiting Adipogenesis and Increasing Uncoupling Protein 1 Expression in White Adipose Tissue of Mice. Nutrients 2018, 10, 331. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, X.; Huang, S.; Li, T.; Zhang, X.; Pang, J.; Zhao, J.; Chen, L.; Zhang, B.; Wang, J.; et al. Milk Fat Globule Membrane Attenuates Acute Colitis and Secondary Liver Injury by Improving the Mucus Barrier and Regulating the Gut Microbiota. Front. Immunol. 2022, 13, 865273. [Google Scholar] [CrossRef]
- Thompson, R.S.; Roller, R.; Mika, A.; Greenwood, B.N.; Knight, R.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Dietary Prebiotics and Bioactive Milk Fractions Improve NREM Sleep, Enhance REM Sleep Rebound and Attenuate the Stress-Induced Decrease in Diurnal Temperature and Gut Microbial Alpha Diversity. Front. Behav. Neurosci. 2016, 10, 240. [Google Scholar] [CrossRef]
- Ota, N.; Soga, S.; Hase, T.; Shimotoyodome, A. Daily consumption of milk fat globule membrane plus habitual exercise improves physical performance in healthy middle-aged adults. Springerplus 2015, 4, 120. [Google Scholar] [CrossRef] [PubMed]
- Minegishi, Y.; Ota, N.; Soga, S.; Shimotoyodome, A. Effects of Nutritional Supplementation with Milk Fat Globule Membrane on Physical and Muscle Function in Healthy Adults Aged 60 and Over with Semiweekly Light Exercise: A Randomized Double-Blind, Placebo-Controlled Pilot Trial. J. Nutr. Sci. Vitaminol. 2016, 62, 409–415. [Google Scholar] [CrossRef]
- Kokai, Y.; Mikami, N.; Tada, M.; Tomonobu, K.; Ochiai, R.; Osaki, N.; Katsuragi, Y.; Sohma, H.; Ito, Y.M. Effects of dietary supplementation with milk fat globule membrane on the physical performance of community-dwelling Japanese adults: A randomised, double-blind, placebo-controlled trial. J. Nutr. Sci. 2018, 7, e18. [Google Scholar] [CrossRef] [PubMed]
- Demmer, E.; Van Loan, M.D.; Rivera, N.; Rogers, T.S.; Gertz, E.R.; German, J.B.; Smilowitz, J.T.; Zivkovic, A.M. Addition of a dairy fraction rich in milk fat globule membrane to a high-saturated fat meal reduces the postprandial insulinaemic and inflammatory response in overweight and obese adults. J. Nutr. Sci. 2016, 5, e14. [Google Scholar] [CrossRef] [PubMed]
- Grip, T.; Dyrlund, T.S.; Ahonen, L.; Domellöf, M.; Hernell, O.; Hyötyläinen, T.; Knip, M.; Lönnerdal, B.; Orešič, M.; Timby, N. Serum, plasma and erythrocyte membrane lipidomes in infants fed formula supplemented with bovine milk fat globule membranes. Pediatr. Res. 2018, 84, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Colombo, J.; Harris, C.L.; Wampler, J.L.; Zhuang, W.; Shaddy, D.J.; Liu, B.Y.; Wu, S.S. Improved Neurodevelopmental Outcomes at 5.5 Years of Age in Children Who Received Bovine Milk Fat Globule Membrane and Lactoferrin in Infant Formula through 12 Months: A Randomized Controlled Trial. J. Pediatr. 2023, 261, 113483. [Google Scholar] [CrossRef] [PubMed]
- Palmano, K.P.; MacGibbon, A.K.H.; Gunn, C.A.; Schollum, L.M. In Vitro and In Vivo Anti-inflammatory Activity of Bovine Milkfat Globule (MFGM)-derived Complex Lipid Fractions. Nutrients 2020, 12, 2089. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.L.; Duncan, S.E.; Sumner, S.S.; Waterman, K.M.; Kaylegian, K.E. Influence of emulsifying component composition on creams formulated with fractionated milkfat. J. Agric. Food Chem. 2003, 51, 5933–5940. [Google Scholar] [CrossRef]
- Kanno, C.; Kim, D.-H. A simple procedure for the preparation of bovine milk fat globule membrane and a comparison of its composition, enzymatic activities, and electrophoretic properties with those prepared by other methods. Agric. Biol. Chem. 1990, 54, 2845–2854. [Google Scholar]
- Newman, R.A.; Harrison, R. Characterisation of the surface of bovine milk fat globule membrane using microelectrophoresis. Biochim. Biophys. Acta 1973, 298, 798–809. [Google Scholar] [CrossRef]
- Kathriarachchi, K.; Leus, M.; Everett, D.W. Oxidation of aldehydes by xanthine oxidase located on the surface of emulsions and washed milk fat globules. Int. Dairy J. 2014, 37, 117–126. [Google Scholar] [CrossRef]
- Dowben, R.M.; Brunner, J.R.; Philpott, D.E. Studies on milk fat globule membranes. Biochim. Biophys. Acta 1967, 135, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Khodaparast-Sharifi, S.H.; Snow, L.D. Levamisole inhibition of alkaline phosphatase and 5’-nucleotidase of bovine milk fat globule membranes. Int. J. Biochem. 1989, 21, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Innocente, N.; Blecker, C.; Deroanne, C.; Paquot, M. Langmuir Film Balance Study of the Surface Properties of a Soluble Fraction of Milk Fat-Globule Membrane. J. Agric. Food Chem. 1997, 45, 1559–1563. [Google Scholar] [CrossRef]
- Nejjar, Y.; Pâquet, D.; Godbillon, G.; Le Deaut, J. Immunological relationship between the hydrophobic fraction of proteose-peptone and the milk fat globule membrane of bovine milk. Int. J. Biochem. 1986, 18, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ruiz, M.E.; Richter, R.L. Effect of Homogenization Pressure on the Milk Fat Globule Membrane Proteins. J. Dairy Sci. 1997, 80, 2732–2739. [Google Scholar] [CrossRef]
- Rombaut, R.; Dejonckheere, V.; Dewettinck, K. Microfiltration of butter serum upon casein micelle destabilization. J. Dairy Sci. 2006, 89, 1915–1925. [Google Scholar] [CrossRef]
- Corredig, M.; Roesch, R.R.; Dalgleish, D.G. Production of a novel ingredient from buttermilk. J. Dairy Sci. 2003, 86, 2744–2750. [Google Scholar] [CrossRef]
- Holzmüller, W.; Kulozik, U. Technical difficulties and future challenges in isolating membrane material from milk fat globules in industrial settings—A critical review. J. Dairy Sci. 2016, 61, 51–66. [Google Scholar] [CrossRef]
- Iung, J.; Pouliot, Y.; Remondetto, G.; Jiménez-Flores, R.; Brisson, G. On the use on hydroxyapatite suspensions for the separation of milk fat globule membrane components from buttermilk. Food Chem. 2023, 404 Pt A, 134535. [Google Scholar] [CrossRef]
- Astaire, J.; Ward, R.; German, J.; Jiménez-Flores, R. Concentration of polar MFGM lipids from buttermilk by microfiltration and supercritical fluid extraction. J. Dairy Sci. 2003, 86, 2297–2307. [Google Scholar] [CrossRef]
- Roesch, R.R.; Rincon, A.; Corredig, M. Emulsifying properties of fractions prepared from commercial buttermilk by microfiltration. J. Dairy Sci. 2004, 87, 4080–4087. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M. Phospholipids in milk fat: Composition, biological and technological significance, and analytical strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef]
- Li, H.; Guan, K.; Liu, M.; Liu, D.; Wang, W.; Zhu, A. Novel antioxidant peptides from MFGM protein Hydrolysates: Separation, identification and effects on Dexamethasone-induced mitochondrial dysfunction. Food Chem. 2023, 403, 134473. [Google Scholar] [CrossRef]
- He, S.; Tang, H.; Yi, H.; Xu, W.; Ma, Y.; Wang, R. Properties of emulsions from milk fat globule membrane and its components. Int. J. Food Prop. 2017, 20, 1342–1353. [Google Scholar] [CrossRef]
- Gunning, P.A.; Mackie, A.R.; Gunning, A.P.; Woodward, N.C.; Wilde, P.J.; Morris, V.J. Effect of surfactant type on surfactant--protein interactions at the air-water interface. Biomacromolecules 2004, 5, 984–991. [Google Scholar] [CrossRef]
- Coke, M.; Wilde, P.J.; Russell, E.J.; Clark, D.C. The influence of surface composition and molecular diffusion on the stability of foams formed from protein/surfactant mixtures. J. Colloid Interface Sci. 1990, 138, 489–504. [Google Scholar] [CrossRef]
- Corredig, M.; Dalgleish, D.G. Isolates from Industrial Buttermilk: Emulsifying Properties of Materials Derived from the Milk Fat Globule Membrane. J. Agric. Food Chem. 1997, 45, 4595–4600. [Google Scholar] [CrossRef]
- Phan, T.T.Q.; Le, T.T.; Van de Walle, D.; Van der Meeren, P.; Dewettinck, K. Combined effects of milk fat globule membrane polar lipids and protein concentrate on the stability of oil-in-water emulsions. Int. Dairy J. 2016, 52, 42–49. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, Y.; Sun, M.; Liu, L.; Li, X.; Zhang, X.; Sun, Y.; Bora, A.F.M.; Li, C.; Leng, Y.; et al. Effects of egg yolk lecithin/milk fat globule membrane material ratio on the structure and stability of oil-in-water emulsions. LWT 2022, 168, 113891. [Google Scholar] [CrossRef]
- Liu, L.; Pan, Y.; Zhang, X.; Zhang, Y.; Li, X. Effect of Particle Size and Interface Composition on the Lipid Digestion of Droplets Covered with Membrane Phospholipids. J. Agric. Food Chem. 2021, 69, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Fani, A.; Singh, H. Biophysical insights into modulating lipid digestion in food emulsions. Prog. Lipid Res. 2022, 85, 101129. [Google Scholar] [CrossRef]
- Zhu, D.; Damodaran, S. Dairy Lecithin from Cheese Whey Fat Globule Membrane: Its Extraction, Composition, Oxidative Stability, and Emulsifying Properties. J. Am. Oil Chem. Soc. 2012, 90, 217–224. [Google Scholar] [CrossRef]
- Timby, N.; Adamsson, M.; Domellöf, E.; Grip, T.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment and growth until 6.5 years of infants who consumed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2021, 113, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Ambrożej, D.; Dumycz, K.; Dziechciarz, P.; Ruszczyński, M. Milk Fat Globule Membrane Supplementation in Children: Systematic Review with Meta-Analysis. Nutrients 2021, 13, 714. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Zhao, J.; Qiao, W.; Liu, B.; Yang, B.; Chen, L. Comparison of phospholipid composition and microstructure of milk fat globules contained in human milk and infant formulae. Food Chem. 2023, 415, 135762. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; Zhang, Z.; Liu, X.; Zhao, S.; Bai, X.; Luo, X.; Feng, D.; Yang, L.; Cao, X.; Yue, X. Quantitative label-free site-specific glycoproteomic analysis of the milk fat globule membrane protein in human colostrum and mature milk. Carbohydr. Polym. 2023, 306, 120588. [Google Scholar] [CrossRef]
- Timby, N.; Hernell, O.; Vaarala, O.; Melin, M.; Lönnerdal, B.; Domellöf, M. Infections in infants fed formula supplemented with bovine milk fat globule membranes. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 384–389. [Google Scholar] [CrossRef]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Stenlund, H.; Lönnerdal, B.; Hernell, O. Feeding Infants Formula With Probiotics or Milk Fat Globule Membrane: A Double-Blind, Randomized Controlled Trial. Front. Pediatr. 2019, 7, 347. [Google Scholar] [CrossRef]
- Chen, M.; Sagis, L.M.C. The influence of protein/phospholipid ratio on the physicochemical and interfacial properties of biomimetic milk fat globules. Food Hydrocoll. 2019, 97, 105179. [Google Scholar] [CrossRef]
- Singh, H. The milk fat globule membrane—A biophysical system for food applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 154–163. [Google Scholar] [CrossRef]
- Arranz, E.; Corredig, M. Invited review: Milk phospholipid vesicles, their colloidal properties, and potential as delivery vehicles for bioactive molecules. J. Dairy Sci. 2017, 100, 4213–4222. [Google Scholar] [CrossRef] [PubMed]
- Farhang, B.; Kakuda, Y.; Corredig, M. Encapsulation of ascorbic acid in liposomes prepared with milk fat globule membrane-derived phospholipids. Dairy Sci. Technol. 2012, 92, 353–366. [Google Scholar] [CrossRef]
- Paramsothy, S.; Kamm, M.A.; Kaakoush, N.O.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; Leong, R.W.L.; Connor, S.; Ng, W.; Paramsothy, R.; et al. Multidonor intensive faecal microbiota transplantation for active ulcerative colitis: A randomised placebo-controlled trial. Lancet 2017, 389, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- da Silva Malheiros, P.; Daroit, D.J.; Brandelli, A. Food applications of liposome-encapsulated antimicrobial peptides. Trends Food Sci. Technol. 2010, 21, 284–292. [Google Scholar] [CrossRef]
- Thompson, A.K.; Couchoud, A.; Singh, H. Comparison of hydrophobic and hydrophilic encapsulation using liposomes prepared from milk fat globule-derived phospholipids and soya phospholipids. Dairy Sci. Technol. 2009, 89, 99–113. [Google Scholar] [CrossRef]
- Liu, W.; Ye, A.; Liu, C.; Liu, W.; Singh, H. Structure and integrity of liposomes prepared from milk- or soybean-derived phospholipids during in vitro digestion. Food Res. Int. 2012, 48, 499–506. [Google Scholar] [CrossRef]
- Gülseren, İ.; Guri, A.; Corredig, M. Encapsulation of Tea Polyphenols in Nanoliposomes Prepared with Milk Phospholipids and Their Effect on the Viability of HT-29 Human Carcinoma Cells. Food Dig. 2012, 3, 36–45. [Google Scholar] [CrossRef]
- Gulseren, I.; Corredig, M. Storage stability and physical characteristics of tea-polyphenol-bearing nanoliposomes prepared with milk fat globule membrane phospholipids. J. Agric. Food Chem. 2013, 61, 3242–3251. [Google Scholar] [CrossRef]
- Moreira, T.G.; Horta, L.S.; Gomes-Santos, A.C.; Oliveira, R.P.; Queiroz, N.M.G.P.; Mangani, D.; Daniel, B.; Vieira, A.T.; Liu, S.; Faria, A.M.C.; et al. CLA-supplemented diet accelerates experimental colorectal cancer by inducing TGF-beta-producing macrophages and T cells. Mucosal Immunol. 2019, 12, 188–199. [Google Scholar] [CrossRef]
- Thompson, A.K.; Haisman, D.; Singh, H. Physical stability of liposomes prepared from milk fat globule membrane and soya phospholipids. J. Agric. Food Chem. 2006, 54, 6390–6397. [Google Scholar] [CrossRef]
- Mathiassen, J.H.; Nejrup, R.G.; Frøkiær, H.; Nilsson, C.; Ohlsson, L.; Hellgren, L.I. Emulsifying triglycerides with dairy phospholipids instead of soy lecithin modulates gut lipase activity. Eur. J. Lipid Sci. Technol. 2015, 117, 1522–1539. [Google Scholar] [CrossRef]
- Li, Z.; Paulson, A.T.; Gill, T.A. Encapsulation of bioactive salmon protein hydrolysates with chitosan-coated liposomes. J. Funct. Foods 2015, 19, 733–743. [Google Scholar] [CrossRef]
- Jash, A.; Rizvi, S.S.H. Heat-stable liposomes from milk fat globule membrane phospholipids for pH-triggered delivery of hydrophilic and lipophilic bioactives. Innov. Food Sci. Emerg. Technol. 2022, 79, 103030. [Google Scholar] [CrossRef]
- Khalid, A.S.; Niaz, T.; Zarif, B.; Shabbir, S.; Noor, T.; Shahid, R.; Imran, M. Milk phospholipids-based nanostructures functionalized with rhamnolipids and bacteriocin: Intrinsic and synergistic antimicrobial activity for cheese preservation. Food Biosci. 2022, 47, 101442. [Google Scholar] [CrossRef]
- Sarabandi, K.; Rafiee, Z.; Khodaei, D.; Jafari, S.M. Encapsulation of food ingredients by nanoliposomes. In Lipid-Based Nanostructures for Food Encapsulation Purposes; Academic Press: Cambridge, MA, USA, 2019; pp. 347–404. [Google Scholar]
- Ünal, N.G.; Kozak, A.; Karakaya, S.; Oruç, N.; Barutçuoğlu, B.; Aktan, Ç.; Sezak, M.; Özütemiz, A. Anti-Inflammatory Effect of Crude Momordica charantia L. Extract on 2,4,6-Trinitrobenzene Sulfonic Acid-Induced Colitis Model in Rat and the Bioaccessibility of its Carotenoid Content. J. Med. Food 2020, 23, 641–648. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Valdés, L.; Altmäe, S.; Ruíz, A.; Suárez, A.; Campoy, C. Role of microbiota function during early life on child’s neurodevelopment. Trends Food Sci. Technol. 2016, 57, 273–288. [Google Scholar] [CrossRef]
- Morin, P.; Jiménez-Flores, R.; Pouliot, Y. Effect of processing on the composition and microstructure of buttermilk and its milk fat globule membranes. Int. Dairy J. 2007, 17, 1179–1187. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Function | Purification Method | Molecular Weight (kDa) | References |
|---|---|---|---|---|
| Butyrophilin | Modulate MFG production and secretion; Might support the infant immune system. | Reversed-phase chromatography | 66–67 | [21] |
| Glycoprotein 2 | Anti-adhesion and immunoprotective | Sepharose CL-4B gel filtration and cesium chloride density-gradient centrifugation | 160–200 | [22] |
| Xanthine oxidase | Redox reaction; anti-inflammatory | Sulfate fractionation and affinity chromatography | 146–155 | [23] |
| Glycosylation-dependent cell adhesion molecule 1 | Mucin-like antibacterial component | Not be established | 18–34 | [24] |
| Carboxyl ester lipase | Hydrolyzes triglycerides, diglycerides, monoglycerides, vitamin A esters, cholesterol, and lysophospholipids in the gut lumen | Reverse-phase high-performance liquid chromatography | 70–80 | [25] |
| Lactoperoxidase | Catalyze oxidation of certain molecules and antimicrobial activity | Ammonium sulfate precipitation and ion-exchange chromatography | 70–80 | [26] |
| Lactoferrin | Promote probiotic growth and antimicrobial activity | Ion-exchange chromatography | 70–80 | [27] |
| Milk fat globule-EGF factor 8 | Maintenance of intestinal epithelial homeostasis and the promotion of mucosal healing and phospholipid binding | Ammonium sulfate precipitation and immunoaffinity chromatography | 47–59 | [28] |
| Fatty acid-binding protein | Lipid transport and regulation of lipid metabolism | Not be established | 13–15 | [29] |
| Clusterin/ apolipoprotein J | Regulate apoptosis and protect other proteins from damage | Not be established | 40–60 | [30] |
| Perilipin 2 | Protect lipid droplets from enzymatic attack by lipases | ion-exchange chromatography | 45–55 | [31] |
| Cluster of differentiation | Mediates innate immunity | immunoaffinity chromatography | 76–78 | [32] |
| Ingredient Description | Model | Design | Primary Finding | Function or Activity | Reference |
|---|---|---|---|---|---|
| MFGM | Male BALB/c mice (7–8 weeks) | Intragastric gavage once a day for 18 weeks | Dietary supplementation by MFGM significantly improved memory and cognitive impairment | Neurodevelopment | [36] |
| Supplementation of formula with MFGM | Infants | Infants were randomized to receive either the experimental formula or SF from inclusion until 6 months of age | Higher concentrations of many phosphatidylcholines and altered concentrations of sphingomyelins | Neurodevelopment | [55] |
| Similar formula (milk fat globule membrane plus lactoferrin [MFGM + LF]) | 5.5 years of age in children | Added sources of bovine MFGM and bovine lactoferrin (bLF) through 12 months of age. | MFGM + LF improved cognitive outcomes in multiple domains at 5.5 years of age, including measures of intelligence and executive function | Neurodevelopment | [56] |
| MFGM complex lipid products | Rats, LPS-stimulated monocytes | MFGM preparations at 38.3 mg/g of diet (treatment groups) for two weeks, anti-arthritic drug via oral gavage (0.12 mg meloxicam/kg body weight (Metacam®, Boehringer-Ingelheim, St. Joseph, MO, USA)) | MFGM to influence the activity and recruitment of inflammatory mediators | Anti-inflammatory | [57] |
| MFGM-10 Lacprodan | Five-week-old male Sprague Dawley (SD) rats | 1.5 g/kg/d of MFGM for 15 days | Inhibition of NLRP3 inflammasome activation | Anti-inflammatory | [38] |
| Milk fat globule-EGF factor VIII (MFG-E8) | Male Sprague-Dawley rats (275–325 g) | rhMFG-E8; 160 μg/kg in 1 mL normal saline | Hepatic I/R induces significant accumulation of apoptotic cells in the live, modulation of PPARγ/NF-κB pathway | Anti-inflammatory | [39] |
| Buttermilk MFGM | MA-104 cells | 125 μL containing the desired amount of MFGM fraction in MEM or MEM alone was mixed with an equal volume of RV containing approximately 1000 FFU of infectious activity. | MFGM inhibited the infectivity of the neuraminidase-sensitive OSU-RV strain in a dose-dependent manner | Anti-RV infection | [42]. |
| MFGM | E. coli O157:H7 | The resulting cell suspensions were diluted 1:100 in fresh medium with and without supplementation with MFGM. The MFGM was tested at 3 protein concentrations: 10, 100, and 1000 μg/mL | MFGM decreased the expression of E. coli O157:H7 virulence gene | Against the E. coli O157:H7 | [44] |
| MFGM | Fischer-344 rats | a combination of AMF and MFGM. After a 7-day acclimation period on standard chow diets | MFGM intake can affect carcinogenesis | Against colon cancer | [47] |
| Whey-derived MFGM (MFGM-10 Lacprodan) | Male C57BL/6J mice (5-week-old) | High-fat diet with MFGM at 100 mg/kg BW, 200 mg/kg BW and 400 mg/kg BW for another 8-week | MFGM suppressed the protein and mRNA expression of PPARγ, C/EBPα and SREBP-1c and activated the AMPK pathway, correlating with the suppression of adipogenic differentiation. MFGM also induced the formation of brown-like adipocytes, as indicated by the upregulation of the protein expression of UCP1. | Anti-obesity | [48] |
| MFGM tablets (1 g MFGM/day) | Middle-aged Japanese adults | During the 10-week study period, the participants took either MFGM (1 g/day) or placebo (1 g/day of whole milk powder) tablets daily and participated in a bi-weekly exercise training program | Daily consumption of MFGM combined with regular exercise improve physical performance in healthy middle-aged adults | Improved physical performance, such as agility | [51] |
| MFGM tablets (1 g MFGM/day) | 22 Japanese participants aged 60–73 years old | One group received MFGM tablets, while the other received placebo tablets daily for a duration of 10 weeks. Both groups participated in a light exercise program twice a week. | Daily supplementation with MFGM led to enhanced motor unit action potential conduction, resulting in improved muscle strength and physical performance in healthy Japanese adults aged over 60 years, who were also involved in bi-weekly light exercise. | Improved physical performance | [52] |
| MFGM tablets | 115 middle-aged subjects (range 50–70 years old) | Six MGFM or Placebo tablets were taken by the participants per d for 24 weeks after breakfast | MFGM exhibited positive effects on the physical performance of community-dwelling Japanese adults | Improved physical performance | [53] |
| Dairy fraction rich in MFGM | Overweight and obese men and women (between 18 and 65 years of age) | Two isoenergetic high-fat meals. One meal includes a smoothie enriched with PO, while the other contains the same smoothie but without a cream-derived complex milk lipid fraction, which is a dairy fraction rich in MFGM | Addition of a dairy fraction rich in MFGM to a high-saturated fat meal reduced the postprandial insulinaemic and inflammatory response | Reduces the postprandial insulinaemic and inflammatory response | [54] |
| Type of Washing Solution | Number of Washing Steps | Cream Washing Solution | Temperature | References |
|---|---|---|---|---|
| Distilled or deionized water | 8 | 1: 3 | 40 °C | [60] |
| 0.25 M sucrose solution containing 2 mM MgCl2 | 4 | 1: 3 | 35 °C | [62] |
| 0.25 M sucrose and 10 mM imidaxole (pH 7.1) | 2 | 1: 1 | 15 °C | [63] |
| Phosphate buffer (NaH2PO4/Na2HPO4 0.01 M, pH 7.2, 0.09% NaCl) | 3 | None | 40 °C | [64] |
| Sucrose–saline solution (WS 1, WS 2, and WS 3) | 3 | None | None | [65] |
| Simulated milk ultrafiltrate containing 6 mol·L−1 urea and 50 mmol·L−1 EDTA | 2 | 1: 10 | 20–25 °C | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, C.; Zhao, Y.; Wang, X.; Li, Y.; Fang, B.; Wang, R.; Wang, X.; Liao, H.; Li, G.; Wang, P.; et al. Structure, Biological Functions, Separation, Properties, and Potential Applications of Milk Fat Globule Membrane (MFGM): A Review. Nutrients 2024, 16, 587. https://doi.org/10.3390/nu16050587
Nie C, Zhao Y, Wang X, Li Y, Fang B, Wang R, Wang X, Liao H, Li G, Wang P, et al. Structure, Biological Functions, Separation, Properties, and Potential Applications of Milk Fat Globule Membrane (MFGM): A Review. Nutrients. 2024; 16(5):587. https://doi.org/10.3390/nu16050587
Chicago/Turabian StyleNie, Chao, Yunyi Zhao, Xifan Wang, Yixuan Li, Bing Fang, Ran Wang, Xiaoyu Wang, Haiping Liao, Gengsheng Li, Pengjie Wang, and et al. 2024. "Structure, Biological Functions, Separation, Properties, and Potential Applications of Milk Fat Globule Membrane (MFGM): A Review" Nutrients 16, no. 5: 587. https://doi.org/10.3390/nu16050587