
2’-Hydroxychalcone Induces Autophagy and Apoptosis in Breast Cancer Cells via the Inhibition of the NF-κB Signaling Pathway: In Vitro and In Vivo Studies
 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Antibodies
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Wound Healing Assay
2.5. Transwell Assay
2.6. Detection of Acidic Vesicular Organelles
2.7. Autophagic Flux Analysis
2.8. Flow Cytometric Analysis of Apoptosis
2.9. Measurement of Intracellular ROS
2.10. Immunofluorescence Staining
2.11. Western Blot Analysis
2.12. Xenograft Mouse Model
2.13. H&E Staining
2.14. TUNEL Assay
2.15. RNA Sequencing Analysis and Prediction of the Potential Targets of 2’-HC
2.16. Statistical Analysis
3. Results
3.1. 2’-HC Inhibited the Proliferation of Breast Cancer In Vitro and In Vivo
3.2. 2’-HC Induced Autophagy and Promoted Autophagy Flux in Breast Cancer Cells
3.3. 2’-HC Induced Autophagy-Dependent Apoptosis of Breast Cancer Cells
3.4. Transcriptomic Changes in 2’-HC-Treated Breast Cancer Cells and Network Pharmacological Analysis of 2’-HC
3.5. 2’-HC Regulated MAPK/NF-κB Signaling Pathways and Inhibited Migration/Invasion of Breast Cancer Cells
3.6. 2’-HC Increased the Intracellular ROS Levels and Triggered ERS of Breast Cancer Cells
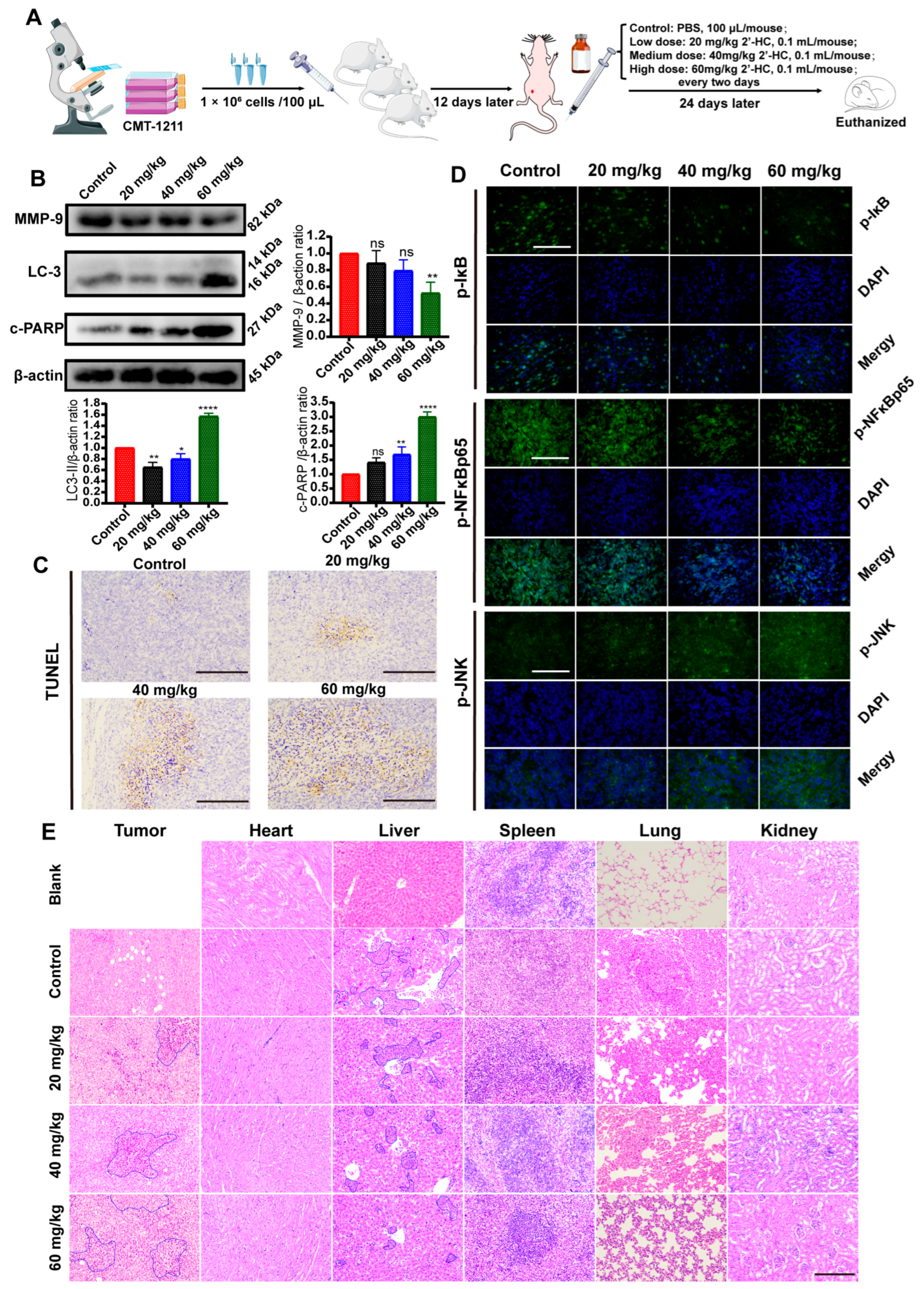
3.7. In Vivo Regulation of 2’-HC in Breast Cancer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2’-HC | (E)2’-hydroxychalone |
| ROS | reactive oxygen species |
| ERS | endoplasmic reticulum stress |
| HCQ | hydroxychloroquine |
| 3-MA | 3-methyladenine |
| NAC | N-acetylcysteine |
| Rap | rapamycin |
| DEGs | differentially expressed genes |
| GSEA | gene set enrichment analysis |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | gene ontology |
| STRING | Search Tool for the Retrieval of Interacting Genes |
| PI | propidium iodide |
| FITC | fluorescein isothiocyanate |
| TUNEL | terminal deoxynucleotidyl transferase dUTP nick end labeling |
| H&E | hematoxylin and eosin |
References
- Ueno, N.T.; Espinosa Fernandez, J.R.; Cristofanilli, M.; Overmoyer, B.; Rea, D.; Berdichevski, F.; El-Shinawi, M.; Bellon, J.; Le-Petross, H.T.; Lucci, A.; et al. International Consensus on the Clinical Management of Inflammatory Breast Cancer from the Morgan Welch Inflammatory Breast Cancer Research Program 10th Anniversary Conference. J. Cancer 2018, 9, 1437–1447. [Google Scholar] [CrossRef]
- Cancer Today. Available online: http://gco.iarc.fr/today/home (accessed on 1 November 2023).
- Orlikova, B.; Tasdemir, D.; Golais, F.; Dicato, M.; Diederich, M. Dietary Chalcones with Chemopreventive and Chemotherapeutic Potential. Genes. Nutr. 2011, 6, 125–147. [Google Scholar] [CrossRef]
- Fan, L.; Strasser-Weippl, K.; Li, J.-J.; St Louis, J.; Finkelstein, D.M.; Yu, K.-D.; Chen, W.-Q.; Shao, Z.-M.; Goss, P.E. Breast Cancer in China. Lancet Oncol. 2014, 15, e279–e289. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Landau, J.M.; Huang, M.T.; Newmark, H.L. Inhibition of Carcinogenesis by Dietary Polyphenolic Compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [PubMed]
- Kar Mahapatra, D.; Asati, V.; Bharti, S.K. An Updated Patent Review of Therapeutic Applications of Chalcone Derivatives (2014–Present). Expert. Opin. Ther. Pat. 2019, 29, 385–406. [Google Scholar] [CrossRef]
- Nowakowska, Z. A Review of Anti-Infective and Anti-Inflammatory Chalcones. Eur. J. Med. Chem. 2007, 42, 125–137. [Google Scholar] [CrossRef]
- Cabrera, M.; Simoens, M.; Falchi, G.; Lavaggi, M.L.; Piro, O.E.; Castellano, E.E.; Vidal, A.; Azqueta, A.; Monge, A.; De Ceráin, A.L.; et al. Synthetic Chalcones, Flavanones, and Flavones as Antitumoral Agents: Biological Evaluation and Structure–Activity Relationships. Bioorganic Med. Chem. 2007, 15, 3356–3367. [Google Scholar] [CrossRef] [PubMed]
- Loa, J.; Chow, P.; Zhang, K. Studies of Structure–Activity Relationship on Plant Polyphenol-Induced Suppression of Human Liver Cancer Cells. Cancer Chemother. Pharmacol. 2009, 63, 1007–1016. [Google Scholar] [CrossRef]
- Wu, X.; Song, M.; Qiu, P.; Li, F.; Wang, M.; Zheng, J.; Wang, Q.; Xu, F.; Xiao, H. The Apple Dihydrochalcone Phloretin Suppresses Growth and Improves Chemosensitivity of Breast Cancer Cells via Inhibition of Cytoprotective Autophagy. Food Funct. 2018, 9, 87–95. [Google Scholar] [CrossRef]
- Chen, Q.; Lei, J.; Zhou, J.; Ma, S.; Huang, Q.; Ge, B. Chemopreventive Effect of 4′-hydroxychalcone on Intestinal Tumorigenesis in Mice. Oncol. Lett. 2021, 21, 213. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, W.-Q.; Zhang, S.-Q.; Bai, J.-X.; Liu, B.; Yung, K.K.-L.; Ko, J.K.-S. Isoliquiritigenin Inhibits Pancreatic Cancer Progression through Blockade of P38 MAPK-Regulated Autophagy. Phytomedicine 2022, 106, 154406. [Google Scholar] [CrossRef]
- Wang, F.-W.; Wang, S.-Q.; Zhao, B.-X.; Miao, J.-Y. Discovery of 2’-Hydroxychalcones as Autophagy Inducer in A549 Lung Cancer Cells. Org. Biomol. Chem. 2014, 12, 3062–3070. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, R.; Das, N.; Tan, C. Inhibitory Effects of 2-Hydroxy Chalcone and Other Flavonoids on Human Cancer Cell-Proliferation. Int. J. Oncol. 1993, 3, 115–119. [Google Scholar] [CrossRef]
- Pande, A.N.; Biswas, S.; Reddy, N.D.; Jayashree, B.S.; Kumar, N.; Rao, C.M. In Vitro and in Vivo Anticancer Studies of 2’-Hydroxy Chalcone Derivatives Exhibit Apoptosis in Colon Cancer Cells by HDAC Inhibition and Cell Cycle Arrest. EXCLI J. 2017, 16, 448–463. [Google Scholar]
- Siqueira, E.D.S.; Concato, V.M.; Tomiotto-Pellissier, F.; Silva, T.F.; Bortoleti, B.T.D.S.; Gonçalves, M.D.; Costa, I.N.; Junior, W.A.V.; Pavanelli, W.R.; Panis, C.; et al. Trans-Chalcone Induces Death by Autophagy Mediated by P53 up-Regulation and β-Catenin down-Regulation on Human Hepatocellular Carcinoma HuH7.5 Cell Line. Phytomedicine 2021, 80, 153373. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Kuo, P.L.; Tzeng, W.S.; Lin, C.C. Chalcone Inhibits the Proliferation of Human Breast Cancer Cell by Blocking Cell Cycle Progression and Inducing Apoptosis. Food Chem. Toxicol. 2006, 44, 704–713. [Google Scholar] [CrossRef]
- Shen, K.-H.; Chang, J.-K.; Hsu, Y.-L.; Kuo, P.-L. Chalcone Arrests Cell Cycle Progression and Induces Apoptosis through Induction of Mitochondrial Pathway and Inhibition of Nuclear Factor Kappa B Signalling in Human Bladder Cancer Cells. Basic Clin. Pharmacol. Toxicol. 2007, 101, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Di, S.; Fan, C.; Ma, Z.; Li, M.; Guo, K.; Han, D.; Li, X.; Mu, D.; Yan, X. PERK/eIF-2α/CHOP Pathway Dependent ROS Generation Mediates Butein-Induced Non-Small-Cell Lung Cancer Apoptosis and G2/M Phase Arrest. Int. J. Biol. Sci. 2019, 15, 1637–1653. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Park, S.J.; Yun, K.-J.; Cho, Y.-W.; Park, H.-J.; Lee, K.-T. Isoliquiritigenin Isolated from the Roots of Glycyrrhiza Uralensis Inhibits LPS-Induced iNOS and COX-2 Expression via the Attenuation of NF-kappaB in RAW 264.7 Macrophages. Eur. J. Pharmacol. 2008, 584, 175–184. [Google Scholar] [CrossRef]
- Pandey, M.K.; Sandur, S.K.; Sung, B.; Sethi, G.; Kunnumakkara, A.B.; Aggarwal, B.B. Butein, a Tetrahydroxychalcone, Inhibits Nuclear Factor (NF)-kappaB and NF-kappaB-Regulated Gene Expression through Direct Inhibition of IkappaBalpha Kinase Beta on Cysteine 179 Residue. J. Biol. Chem. 2007, 282, 17340–17350. [Google Scholar] [CrossRef]
- Srinivasan, B.; Johnson, T.E.; Lad, R.; Xing, C. Structure-Activity Relationship Studies of Chalcone Leading to 3-Hydroxy-4,3′,4′,5′-Tetramethoxychalcone and Its Analogues as Potent Nuclear Factor kappaB Inhibitors and Their Anticancer Activities. J. Med. Chem. 2009, 52, 7228–7235. [Google Scholar] [CrossRef]
- Verzella, D.; Pescatore, A.; Capece, D.; Vecchiotti, D.; Ursini, M.V.; Franzoso, G.; Alesse, E.; Zazzeroni, F. Life, Death, and Autophagy in Cancer: NF-κB Turns up Everywhere. Cell Death Dis. 2020, 11, 210. [Google Scholar] [CrossRef]
- Liu, G.; Ge, Z.; Zhao, M.; Zhou, Y. Design, Synthesis and Cytotoxic Activities of Novel Aliphatic Amino-Substituted Flavonoids. Molecules 2013, 18, 14070–14084. [Google Scholar] [CrossRef]
- Kim, T.-M.; Yang, I.S.; Seung, B.-J.; Lee, S.; Kim, D.; Ha, Y.-J.; Seo, M.-K.; Kim, K.-K.; Kim, H.S.; Cheong, J.-H.; et al. Cross-Species Oncogenic Signatures of Breast Cancer in Canine Mammary Tumors. Nat. Commun. 2020, 11, 3616. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the Pathogenesis of Disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Rybstein, M.D.; Bravo-San Pedro, J.M.; Kroemer, G.; Galluzzi, L. The Autophagic Network and Cancer. Nat. Cell Biol. 2018, 20, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lam, G.Y.; Brumell, J.H. Autophagy Signaling Through Reactive Oxygen Species. Antioxid. Redox Signal. 2011, 14, 2215–2231. [Google Scholar] [CrossRef] [PubMed]
- Clarke, H.J.; Chambers, J.E.; Liniker, E.; Marciniak, S.J. Endoplasmic Reticulum Stress in Malignancy. Cancer Cell 2014, 25, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Wilde, L.; Tanson, K.; Curry, J.; Martinez-Outschoorn, U. Autophagy in Cancer: A Complex Relationship. Biochem. J. 2018, 475, 1939–1954. [Google Scholar] [CrossRef]
- Carbone, D.; Vestuto, V.; Ferraro, M.R.; Ciaglia, T.; Pecoraro, C.; Sommella, E.; Cascioferro, S.; Salviati, E.; Novi, S.; Tecce, M.F.; et al. Metabolomics-assisted Discovery of a New Anticancer GLS-1 Inhibitor Chemotype from a Nortopsentin-inspired Library: From Phenotype Screening to Target Identification. Eur. J. Med. Chem. 2022, 234, 114233. [Google Scholar] [CrossRef]
- Russell, R.C.; Guan, K.L. The multifaceted role of autophagy in cancer. EMBO J. 2022, 41, e110031. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in Mammalian Autophagy Research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef]
- Fan, L. Role of P62 in the Regulation of Cell Death Induction. Apoptosis 2018, 23, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.; Liu, T.; You, Q.; Jiang, Z. P62 as a Therapeutic Target for Tumor. Eur. J. Med. Chem. 2020, 193, 112231. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, A.R. Signal Integration by JNK and P38 MAPK Pathways in Cancer Development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Selim, K.A.; Abdelrasoul, H.; Aboelmagd, M.; Tawila, A.M. The Role of the MAPK Signaling, Topoisomerase and Dietary Bioactives in Controlling Cancer Incidence. Diseases 2017, 5, 13. [Google Scholar] [CrossRef]
- Stefanelli, C.; Tantini, B.; Fattori, M.; Stanic’, I.; Pignatti, C.; Clo, C.; Guarnieri, C.; Caldarera, C.M.; Mackintosh, C.A.; Pegg, A.E.; et al. Caspase Activation in Etoposide-Treated Fibroblasts Is Correlated to ERK Phosphorylation and Both Events Are Blocked by Polyamine Depletion. FEBS Lett. 2002, 527, 223–228. [Google Scholar] [CrossRef]
- Gomez-Sarosi, L.A.; Strasberg-Rieber, M.; Rieber, M. ERK Activation Increases Nitroprusside Induced Apoptosis in Human Melanoma Cells Irrespective of P53 Status: Role of Superoxide Dismutases. Cancer Biol. Ther. 2009, 8, 1173–1182. [Google Scholar] [CrossRef]
- Zhang, G.; He, J.; Ye, X.; Zhu, J.; Hu, X.; Shen, M.; Ma, Y.; Mao, Z.; Song, H.; Chen, F. β-Thujaplicin Induces Autophagic Cell Death, Apoptosis, and Cell Cycle Arrest through ROS-Mediated Akt and P38/ERK MAPK Signaling in Human Hepatocellular Carcinoma. Cell Death Dis. 2019, 10, 255. [Google Scholar] [CrossRef]
- Luo, J.-L.; Kamata, H.; Karin, M. IKK/NF-kappaB Signaling: Balancing Life and Death-a New Approach to Cancer Therapy. J. Clin. Investig. 2005, 115, 2625–2632. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, J.Y.; Seo, G.S.; Kim, Y.-C.; Sohn, D.H. Isoliquiritigenin, from Dalbergia Odorifera, up-Regulates Anti-Inflammatory Heme Oxygenase-1 Expression in RAW264.7 Macrophages. Inflamm. Res. 2009, 58, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kim, M.-O.; Choi, Y.H.; Kim, G.-Y. Butein Sensitizes Human Hepatoma Cells to TRAIL-Induced Apoptosis via Extracellular Signal-Regulated Kinase/Sp1–Dependent DR5 Upregulation and NF-κB Inactivation. Mol. Cancer Ther. 2010, 9, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Orlikova, B.; Tasdemir, D.; Golais, F.; Dicato, M.; Diederich, M. The Aromatic Ketone 4′-Hydroxychalcone Inhibits TNFα-Induced NF-κB Activation via Proteasome Inhibition. Biochem. Pharmacol. 2011, 82, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Kiningham, K.K.; Xu, Y.; Daosukho, C.; Popova, B.; St Clair, D.K. Nuclear Factor kappaB-Dependent Mechanisms Coordinate the Synergistic Effect of PMA and Cytokines on the Induction of Superoxide Dismutase 2. Biochem. J. 2001, 353, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Papa, S.; Jones, J.; Alvarez, K.; Jayawardena, S.; De Smaele, E.; Cong, R.; Beaumont, C.; et al. Ferritin Heavy Chain Upregulation by NF-kappaB Inhibits TNFalpha-Induced Apoptosis by Suppressing Reactive Oxygen Species. Cell 2004, 119, 529–542. [Google Scholar] [CrossRef]
- Kamata, H.; Honda, S.-I.; Maeda, S.; Chang, L.; Hirata, H.; Karin, M. Reactive Oxygen Species Promote TNFalpha-Induced Death and Sustained JNK Activation by Inhibiting MAP Kinase Phosphatases. Cell 2005, 120, 649–661. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-κB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Liang, Y.; Zhang, B.; He, L.; Li, W.; Zhang, W.; Li, C.; Luo, L.; Umar, T.; Feng, H.; et al. 2’-Hydroxychalcone Induces Autophagy and Apoptosis in Breast Cancer Cells via the Inhibition of the NF-κB Signaling Pathway: In Vitro and In Vivo Studies. Nutrients 2024, 16, 514. https://doi.org/10.3390/nu16040514
Wang X, Liang Y, Zhang B, He L, Li W, Zhang W, Li C, Luo L, Umar T, Feng H, et al. 2’-Hydroxychalcone Induces Autophagy and Apoptosis in Breast Cancer Cells via the Inhibition of the NF-κB Signaling Pathway: In Vitro and In Vivo Studies. Nutrients. 2024; 16(4):514. https://doi.org/10.3390/nu16040514
Chicago/Turabian StyleWang, Xiao, Yongjie Liang, Bohan Zhang, Lixin He, Wenxuan Li, Wenwen Zhang, Chengzong Li, Lihong Luo, Talha Umar, Huili Feng, and et al. 2024. "2’-Hydroxychalcone Induces Autophagy and Apoptosis in Breast Cancer Cells via the Inhibition of the NF-κB Signaling Pathway: In Vitro and In Vivo Studies" Nutrients 16, no. 4: 514. https://doi.org/10.3390/nu16040514