
PNPLA3 Genotype and Dietary Fat Modify Concentrations of Plasma and Fecal Short Chain Fatty Acids and Plasma Branched-Chain Amino Acids
 , , , and
, , , and
Abstract
:1. Introduction
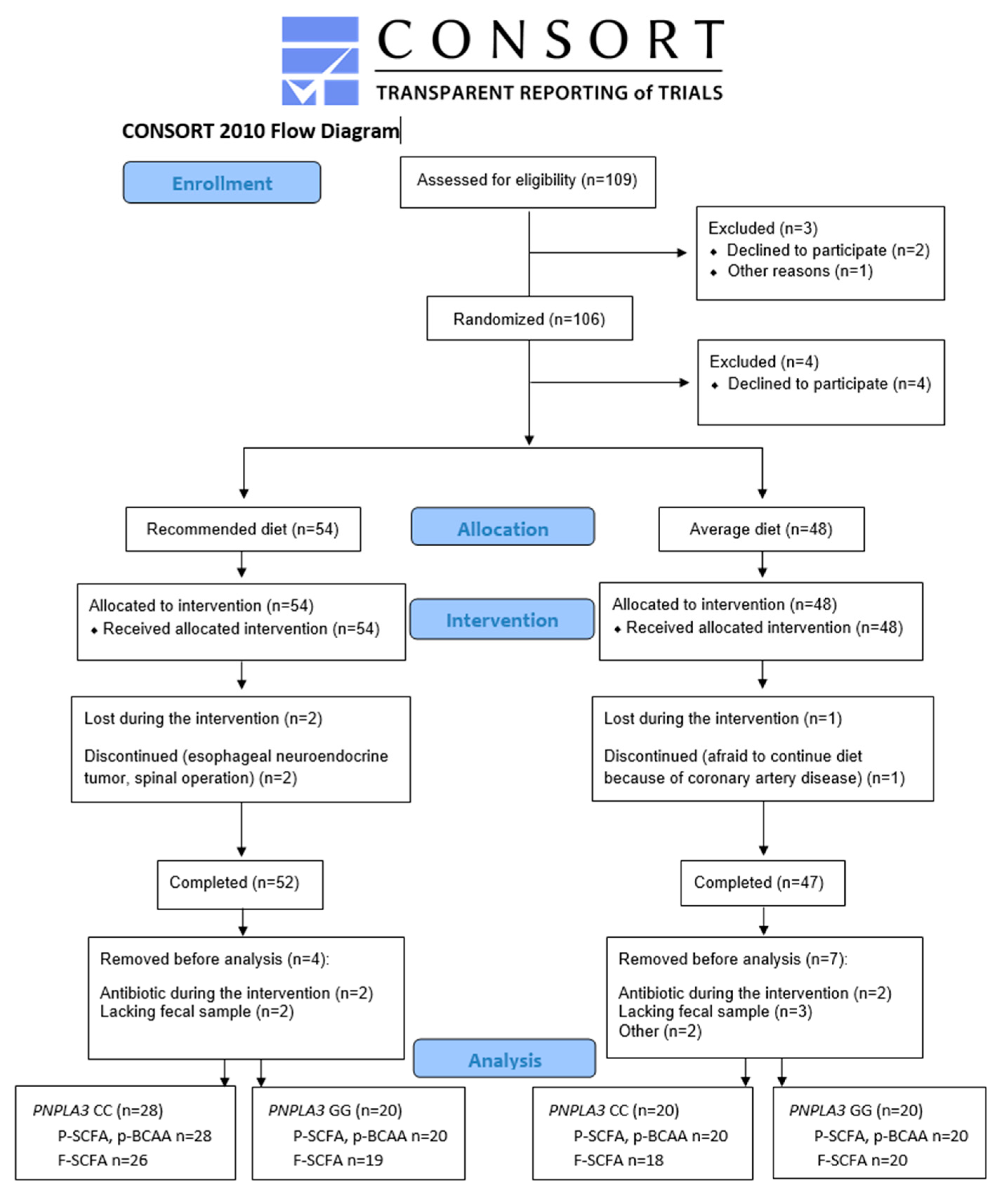
2. Materials and Methods
2.1. Study Participants
2.2. Genotyping for PNPLA3
2.3. Dietary Intervention and Food Records
2.4. Laboratory Analysis
2.5. Calculations for Glucose Metabolism and NAFLD Associated Scores
2.6. Plasma and Fecal SCFAs and Plasma BCAAs Quantification
2.7. Statistical Methods
2.8. Ethical Considerations
3. Results
3.1. Dietary Intervention: Recommended Diet and Average Diet
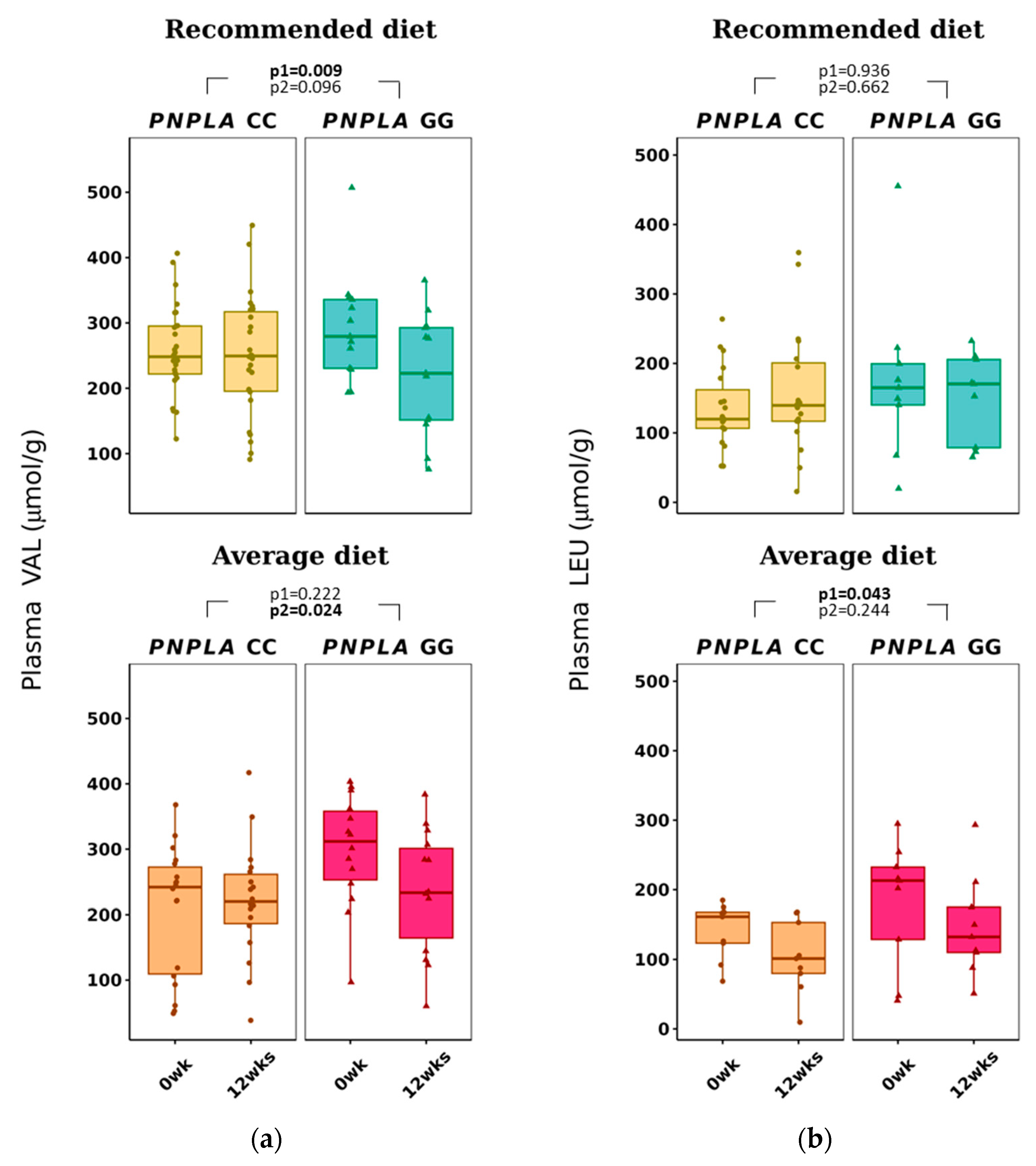
3.2. Changes in Dietary Fat Quality on Plasma SCFA and BCAA
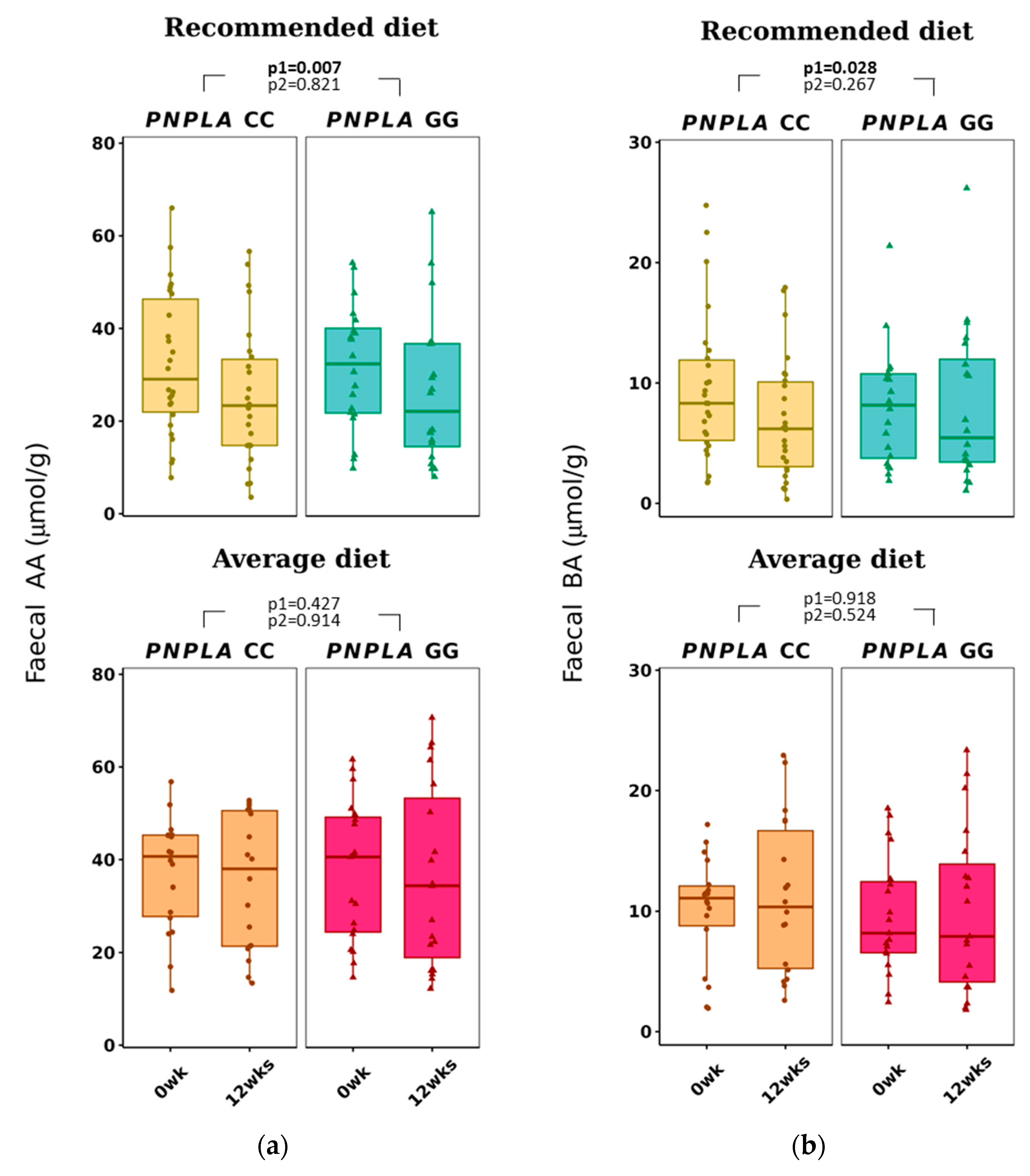
3.3. Changes in Dietary Fat Quality Modification on Fecal SCFA
3.4. Associations of Food Intake and Plasma SCFA and BCAA and Fecal SCFA
3.5. Plasma SCFA and BCAA Associate with Lipids, Glucose Metabolism and NAFLD Associated Scores
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic Variation in PNPLA3 Confers Susceptibility to Nonalcoholic Fatty Liver Disease. Nat. Genet. 2008, 40, 1461–1465.e24. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Waterworth, D.; Perry, J.R.B.; Lim, N.; Song, K.; Chambers, J.C.; Zhang, W.; Vollenweider, P.; Stirnadel, H.; Johnson, T.; et al. Population-Based Genome-Wide Association Studies Reveal Six Loci Influencing Plasma Levels of Liver Enzymes. Am. J. Hum. Genet. 2008, 83, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Trépo, E.; Valenti, L. Update on NAFLD Genetics: From New Variants to the Clinic. J. Hepatol. 2020, 72, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.C. PNPLA3-A Potential Therapeutic Target for Personalized Treatment of Chronic Liver Disease. Front. Med. 2019, 6, 304. [Google Scholar] [CrossRef] [PubMed]
- Kotronen, A.; Johansson, L.E.; Johansson, L.M.; Roos, C.; Westerbacka, J.; Hamsten, A.; Bergholm, R.; Arkkila, P.; Arola, J.; Kiviluoto, T.; et al. A Common Variant in PNPLA3, Which Encodes Adiponutrin, Is Associated with Liver Fat Content in Humans. Diabetologia 2009, 52, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Son, H.; Koo, B.K.; Joo, S.K.; Lee, D.H.; Jang, H.; Park, J.H.; Chang, M.S.; Kim, W. Innovative Target Exploration of NAFLD (ITEN) consortium PNPLA3 Genotypes Modify the Adverse Effect of the Total Energy Intake on High-Risk Nonalcoholic Steatohepatitis Development. Am. J. Clin. Nutr. 2023, 117, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.E.; Wong, V.W.-S.; Rinella, M. Non-Alcoholic Fatty Liver Disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Raizi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, P.M.G.; Congly, S.E.; Kaplan, P.G.G.; Shaheen, A.-A. The Prevalence and Incidence of NAFLD Worldwide: A Systematic Review and Meta-Analysis. Lancet. Gastroenterol. Hepatol. 2022, 7, 74–75. [Google Scholar] [CrossRef]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global Burden of NAFLD and NASH: Trends, Predictions, Risk Factors and Prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Yki-Järvinen, H. Non-Alcoholic Fatty Liver Disease as a Cause and a Consequence of Metabolic Syndrome. Lancet Diabetes Endocrinol. 2014, 2, 901–910. [Google Scholar] [CrossRef]
- Jarvis, H.; Craig, D.; Barker, R.; Spiers, G.; Stow, D.; Anstee, Q.M.; Hanratty, B. Metabolic Risk Factors and Incident Advanced Liver Disease in Non-Alcoholic Fatty Liver Disease (NAFLD): A Systematic Review and Meta-Analysis of Population-Based Observational Studies. PLoS Med. 2020, 17, e1003100. [Google Scholar] [CrossRef]
- Younossi, Z.M. Non-Alcoholic Fatty Liver Disease—A Global Public Health Perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef]
- Jonas, W.; Schürmann, A. Genetic and Epigenetic Factors Determining NAFLD Risk. Mol. Metab. 2021, 50, 101111. [Google Scholar] [CrossRef]
- Ramos Meyers, G.; Samouda, H.; Bohn, T. Short Chain Fatty Acid Metabolism in Relation to Gut Microbiota and Genetic Variability. Nutrients 2022, 14, 5361. [Google Scholar] [CrossRef]
- Lankinen, M.; Uusitupa, M.; Schwab, U. Genes and Dietary Fatty Acids in Regulation of Fatty Acid Composition of Plasma and Erythrocyte Membranes. Nutrients 2018, 10, 1785. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; De Michieli, F.; Cassader, M.; Rizzetto, M.; Durazzo, M.; Fagà, E.; Silli, B.; Pagano, G. Dietary Habits and Their Relations to Insulin Resistance and Postprandial Lipemia in Nonalcoholic Steatohepatitis. Hepatology 2003, 37, 909–916. [Google Scholar] [CrossRef]
- Tiikkainen, M.; Bergholm, R.; Vehkavaara, S.; Rissanen, A.; Häkkinen, A.-M.; Tamminen, M.; Teramo, K.; Yki-Järvinen, H. Effects of Identical Weight Loss on Body Composition and Features of Insulin Resistance in Obese Women with High and Low Liver Fat Content. Diabetes 2003, 52, 701–707. [Google Scholar] [CrossRef]
- Luukkonen, P.K.; Sädevirta, S.; Zhou, Y.; Kayser, B.; Ali, A.; Ahonen, L.; Lallukka, S.; Pelloux, V.; Gaggini, M.; Jian, C.; et al. Saturated Fat Is More Metabolically Harmful for the Human Liver Than Unsaturated Fat or Simple Sugars. Diabetes Care 2018, 41, 1732–1739. [Google Scholar] [CrossRef]
- Yki-Järvinen, H.; Luukkonen, P.K.; Hodson, L.; Moore, J.B. Dietary Carbohydrates and Fats in Nonalcoholic Fatty Liver Disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 770–786. [Google Scholar] [CrossRef]
- Rosqvist, F.; Iggman, D.; Kullberg, J.; Cedernaes, J.; Johansson, H.-E.; Larsson, A.; Johansson, L.; Ahlström, H.; Arner, P.; Dahlman, I.; et al. Overfeeding Polyunsaturated and Saturated Fat Causes Distinct Effects on Liver and Visceral Fat Accumulation in Humans. Diabetes 2014, 63, 2356–2368. [Google Scholar] [CrossRef]
- Valsta, L.; Kaartinen, N.; Tapanainen, H.; Männistö, S.; Sääksjärvi, K. Nutrition in Finland—The National FinDiet 2017 Survey; PunaMusta Oy, 2018; Finland Institute for Health and Welfare: Helsinki, Finland, 2018; pp. 74–75. [Google Scholar]
- Tsompanaki, E.; Thanapirom, K.; Papatheodoridi, M.; Parikh, P.; Chotai de Lima, Y.; Tsochatzis, E.A. Systematic Review and Meta-Analysis: The Role of Diet in the Development of Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2023, 21, 1462–1474.e24. [Google Scholar] [CrossRef]
- Cronin, P.; Joyce, S.A.; O’Toole, P.W.; O’Connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655. [Google Scholar] [CrossRef]
- Bashiardes, S.; Shapiro, H.; Rozin, S.; Shibolet, O.; Elinav, E. Non-Alcoholic Fatty Liver and the Gut Microbiota. Mol. Metab. 2016, 5, 782–794. [Google Scholar] [CrossRef]
- Puertollano, E.; Kolida, S.; Yaqoob, P. Biological Significance of Short-Chain Fatty Acid Metabolism by the Intestinal Microbiome. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 139–144. [Google Scholar] [CrossRef]
- Jasirwan, C.O.M.; Lesmana, C.R.A.; Hasan, I.; Sulaiman, A.S.; Gani, R.A. The Role of Gut Microbiota in Non-Alcoholic Fatty Liver Disease: Pathways of Mechanisms. Biosci. Microbiota Food Health 2019, 38, 81–88. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]
- Rau, M.; Rehman, A.; Dittrich, M.; Groen, A.K.; Hermanns, H.M.; Seyfried, F.; Beyersdorf, N.; Dandekar, T.; Rosenstiel, P.; Geier, A. Fecal SCFAs and SCFA-Producing Bacteria in Gut Microbiome of Human NAFLD as a Putative Link to Systemic T-Cell Activation and Advanced Disease. United Eur. Gastroenterol. J. 2018, 6, 1496–1507. [Google Scholar] [CrossRef]
- Heimann, E.; Nyman, M.; Pålbrink, A.-K.; Lindkvist-Petersson, K.; Degerman, E. Branched Short-Chain Fatty Acids Modulate Glucose and Lipid Metabolism in Primary Adipocytes. Adipocyte 2016, 5, 359–368. [Google Scholar] [CrossRef]
- Sowah, S.A.; Riedl, L.; Damms-Machado, A.; Johnson, T.S.; Schübel, R.; Graf, M.; Kartal, E.; Zeller, G.; Schwingshackl, L.; Stangl, G.I.; et al. Effects of Weight-Loss Interventions on Short-Chain Fatty Acid Concentrations in Blood and Feces of Adults: A Systematic Review. Adv. Nutr. 2019, 10, 673–684. [Google Scholar] [CrossRef]
- Pagliai, G.; Russo, E.; Niccolai, E.; Dinu, M.; Di Pilato, V.; Magrini, A.; Bartolucci, G.; Baldi, S.; Menicatti, M.; Giusti, B.; et al. Influence of a 3-Month Low-Calorie Mediterranean Diet Compared to the Vegetarian Diet on Human Gut Microbiota and SCFA: The CARDIVEG Study. Eur. J. Nutr. 2020, 59, 2011–2024. [Google Scholar] [CrossRef]
- Vasconcellos, C.; Ferreira, O.; Lopes, M.F.; Ribeiro, A.F.; Vasques, J.; Guerreiro, C.S. Nutritional Genomics in Nonalcoholic Fatty Liver Disease. Biomedicines 2023, 11, 319. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Qi, Q.; Hruby, A.; Manson, J.E.; Willett, W.C.; Wolpin, B.M.; Hu, F.B.; Qi, L. Cumulative Consumption of Branched-Chain Amino Acids and Incidence of Type 2 Diabetes. Int. J. Epidemiol. 2016, 45, 1482–1492. [Google Scholar] [CrossRef]
- Jennings, A.; MacGregor, A.; Pallister, T.; Spector, T.; Cassidy, A. Associations between Branched Chain Amino Acid Intake and Biomarkers of Adiposity and Cardiometabolic Health Independent of Genetic Factors: A Twin Study. Int. J. Cardiol. 2016, 223, 992–998. [Google Scholar] [CrossRef]
- Branched-Chain Amino Acids Are Associated with Cardiometabolic Risk Profiles Found Already in Lean, Overweight and Obese Young—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/27142745/ (accessed on 16 March 2023).
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Harris, R.A. Exercise Promotes BCAA Catabolism: Effects of BCAA Supplementation on Skeletal Muscle during Exercise. J. Nutr. 2004, 134, 1583S–1587S. [Google Scholar] [CrossRef]
- Giacco, A.; Cioffi, F.; Cuomo, A.; Simiele, R.; Senese, R.; Silvestri, E.; Amoresano, A.; Fontanarosa, C.; Petito, G.; Moreno, M.; et al. Mild Endurance Exercise during Fasting Increases Gastrocnemius Muscle and Prefrontal Cortex Thyroid Hormone Levels through Differential BHB and BCAA-Mediated BDNF-MTOR Signaling in Rats. Nutrients 2022, 14, 1166. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature That Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell. Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef]
- Guasch-Ferre, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.G.; Reily, M.D.; Lehman-McKeeman, L.D.; Vaillancourt, R.R.; Cherrington, N.J. Branched Chain Amino Acid Metabolism Profiles in Progressive Human Nonalcoholic Fatty Liver Disease. Amino Acids 2015, 47, 603–615. [Google Scholar] [CrossRef]
- Zhao, X.; Han, Q.; Liu, Y.; Sun, C.; Gang, X.; Wang, G. The Relationship between Branched-Chain Amino Acid Related Metabolomic Signature and Insulin Resistance: A Systematic Review. J. Diabetes Res. 2016, 2016, 2794591. [Google Scholar] [CrossRef]
- Batch, B.C.; Hyland, K.; Svetkey, L.P. Branch Chain Amino Acids: Biomarkers of Health and Disease. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 86–89. [Google Scholar] [CrossRef]
- Fiehn, O.; Garvey, W.T.; Newman, J.W.; Lok, K.H.; Hoppel, C.L.; Adams, S.H. Plasma Metabolomic Profiles Reflective of Glucose Homeostasis in Non-Diabetic and Type 2 Diabetic Obese African-American Women. PLoS ONE 2010, 5, e15234. [Google Scholar] [CrossRef]
- Lackey, D.E.; Lynch, C.J.; Olson, K.C.; Mostaedi, R.; Ali, M.; Smith, W.H.; Karpe, F.; Humphreys, S.; Bedinger, D.H.; Dunn, T.N.; et al. Regulation of Adipose Branched-Chain Amino Acid Catabolism Enzyme Expression and Cross-Adipose Amino Acid Flux in Human Obesity. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1175–E1187. [Google Scholar] [CrossRef]
- Giesbertz, P.; Daniel, H. Branched-Chain Amino Acids as Biomarkers in Diabetes. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 48–54. [Google Scholar] [CrossRef]
- Clinical and Metabolic Characterization of Lean Caucasian Subjects with Non-Alcoholic Fatty Liver–PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/27527746/ (accessed on 16 March 2023).
- Shah, S.H.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Crosslin, D.R.; Haynes, C.; Dungan, J.; Newby, L.K.; Hauser, E.R.; Ginsburg, G.S.; et al. Association of a Peripheral Blood Metabolic Profile with Coronary Artery Disease and Risk of Subsequent Cardiovascular Events. Circ. Cardiovasc. Genet. 2010, 3, 207–214. [Google Scholar] [CrossRef]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Mochizuki, S.; Sugahara, K.; Yoshizawa, F. Isoleucine, a Blood Glucose-Lowering Amino Acid, Increases Glucose Uptake in Rat Skeletal Muscle in the Absence of Increases in AMP-Activated Protein Kinase Activity. J. Nutr. 2005, 135, 2103–2108. [Google Scholar] [CrossRef]
- Nishimura, J.; Masaki, T.; Arakawa, M.; Seike, M.; Yoshimatsu, H. Isoleucine Prevents the Accumulation of Tissue Triglycerides and Upregulates the Expression of PPARalpha and Uncoupling Protein in Diet-Induced Obese Mice. J. Nutr. 2010, 140, 496–500. [Google Scholar] [CrossRef]
- Lange, O.; Proczko-Stepaniak, M.; Mika, A. Short-Chain Fatty Acids-A Product of the Microbiome and Its Participation in Two-Way Communication on the Microbiome-Host Mammal Line. Curr. Obes. Rep. 2023, 12, 108–126. [Google Scholar] [CrossRef]
- Laakso, M.; Kuusisto, J.; Stančáková, A.; Kuulasmaa, T.; Pajukanta, P.; Lusis, A.J.; Collins, F.S.; Mohlke, K.L.; Boehnke, M. The Metabolic Syndrome in Men Study: A Resource for Studies of Metabolic and Cardiovascular Diseases. J. Lipid Res. 2017, 58, 481–493. [Google Scholar] [CrossRef]
- Mithril, C.; Dragsted, L.O.; Meyer, C.; Blauert, E.; Holt, M.K.; Astrup, A. Guidelines for the New Nordic Diet. Public Health Nutr. 2012, 15, 1941–1947. [Google Scholar] [CrossRef]
- Lankinen, M.A.; de Mello, V.D.; Meuronen, T.; Sallinen, T.; Ågren, J.; Virtanen, K.A.; Laakso, M.; Pihlajamäki, J.; Schwab, U. The FADS1 Genotype Modifies Metabolic Responses to the Linoleic Acid and Alpha-Linolenic Acid Containing Plant Oils-Genotype Based Randomized Trial FADSDIET2. Mol. Nutr. Food Res. 2021, 65, e2001004. [Google Scholar] [CrossRef]
- Lankinen, M.A.; Fauland, A.; Shimizu, B.; Ågren, J.; Wheelock, C.E.; Laakso, M.; Schwab, U.; Pihlajamäki, J. Inflammatory Response to Dietary Linoleic Acid Depends on FADS1 Genotype. Am. J. Clin. Nutr. 2019, 109, 165–175. [Google Scholar] [CrossRef]
- Matsuda, M.; DeFronzo, R.A. Insulin Sensitivity Indices Obtained from Oral Glucose Tolerance Testing: Comparison with the Euglycemic Insulin Clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Ramdas Nayak, V.K.; Satheesh, P.; Shenoy, M.T.; Kalra, S. Triglyceride Glucose (TyG) Index: A Surrogate Biomarker of Insulin Resistance. J. Pak. Med. Assoc. 2022, 72, 986–988. [Google Scholar] [CrossRef]
- Wang, J.; Yan, S.; Cui, Y.; Chen, F.; Piao, M.; Cui, W. The Diagnostic and Prognostic Value of the Triglyceride-Glucose Index in Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD): A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 4969. [Google Scholar] [CrossRef]
- Bedogni, G.; Bellentani, S.; Miglioli, L.; Masutti, F.; Passalacqua, M.; Castiglione, A.; Tiribelli, C. The Fatty Liver Index: A Simple and Accurate Predictor of Hepatic Steatosis in the General Population. BMC Gastroenterol. 2006, 6, 33. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, D.; Kim, H.J.; Lee, C.H.; Yang, J.I.; Kim, W.; Kim, Y.J.; Yoon, J.H.; Cho, S.H.; Sung, M.W.; et al. Hepatic Steatosis Index: A Simple Screening Tool Reflecting Nonalcoholic Fatty Liver Disease. Dig. Liver Dis. 2010, 42, 503–508. [Google Scholar] [CrossRef]
- Kotronen, A.; Peltonen, M.; Hakkarainen, A.; Sevastianova, K.; Bergholm, R.; Johansson, L.M.; Lundbom, N.; Rissanen, A.; Ridderstrale, M.; Groop, L.; et al. Prediction of Non-Alcoholic Fatty Liver Disease and Liver Fat Using Metabolic and Genetic Factors. Gastroenterology 2009, 137, 865–872. [Google Scholar] [CrossRef]
- Zheng, X.; Qiu, Y.; Zhong, W.; Baxter, S.; Su, M.; Li, Q.; Xie, G.; Ore, B.M.; Qiao, S.; Spencer, M.D.; et al. A Targeted Metabolomic Protocol for Short-Chain Fatty Acids and Branched-Chain Amino Acids. Metabolomics 2013, 9, 818–827. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, J.; Tian, Y.; Zhang, L.; Hatzakis, E.; Krausz, K.W.; Smith, P.B.; Gonzalez, F.J.; Patterson, A.D. Orthogonal Comparison of GC-MS and 1H NMR Spectroscopy for Short Chain Fatty Acid Quantitation. Anal. Chem. 2017, 89, 7900–7906. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review Article: The Role of Butyrate on Colonic Function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Maldonado-Contreras, A.; Noel, S.E.; Ward, D.V.; Velez, M.; Mangano, K.M. Associations between Diet, the Gut Microbiome, and Short-Chain Fatty Acid Production among Older Caribbean Latino Adults. J. Acad. Nutr. Diet 2020, 120, 2047–2060.e6. [Google Scholar] [CrossRef]
- Wang, B.; Kong, Q.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. A High-Fat Diet Increases Gut Microbiota Biodiversity and Energy Expenditure Due to Nutrient Difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef]
- Ziętek, M.; Celewicz, Z.; Kikut, J.; Szczuko, M. Implications of SCFAs on the Parameters of the Lipid and Hepatic Profile in Pregnant Women. Nutrients 2021, 13, 1749. [Google Scholar] [CrossRef]
- Zhao, G.; Yang, L.; Zhong, W.; Hu, Y.; Tan, Y.; Ren, Z.; Ban, Q.; Yang, C.S.; Wang, Y.; Wang, Z. Polydatin, A Glycoside of Resveratrol, Is Better Than Resveratrol in Alleviating Non-Alcoholic Fatty Liver Disease in Mice Fed a High-Fructose Diet. Front. Nutr. 2022, 9, 857879. [Google Scholar] [CrossRef]
- Vogt, J.A.; Wolever, T.M.S. Fecal Acetate Is Inversely Related to Acetate Absorption from the Human Rectum and Distal Colon. J. Nutr. 2003, 133, 3145–3148. [Google Scholar] [CrossRef]
- Deng, K.; Xu, J.; Shen, L.; Zhao, H.; Gou, W.; Xu, F.; Fu, Y.; Jiang, Z.; Shuai, M.; Li, B.; et al. Comparison of Fecal and Blood Metabolome Reveals Inconsistent Associations of the Gut Microbiota with Cardiometabolic Diseases. Nat. Commun. 2023, 14, 571. [Google Scholar] [CrossRef]
- Trefflich, I.; Dietrich, S.; Braune, A.; Abraham, K.; Weikert, C. Short- and Branched-Chain Fatty Acids as Fecal Markers for Microbiota Activity in Vegans and Omnivores. Nutrients 2021, 13, 1808. [Google Scholar] [CrossRef]
- McOrist, A.L.; Abell, G.C.J.; Cooke, C.; Nyland, K. Bacterial Population Dynamics and Faecal Short-Chain Fatty Acid (SCFA) Concentrations in Healthy Humans. Br. J. Nutr. 2008, 100, 138–146. [Google Scholar] [CrossRef]
- Zhou, M.; Shao, J.; Wu, C.-Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA Catabolism to Treat Obesity-Associated Insulin Resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef]
- Vanweert, F.; Schrauwen, P.; Phielix, E. Role of Branched-Chain Amino Acid Metabolism in the Pathogenesis of Obesity and Type 2 Diabetes-Related Metabolic Disturbances BCAA Metabolism in Type 2 Diabetes. Nutr. Diabetes 2022, 12, 35. [Google Scholar] [CrossRef]
- Tai, E.S.; Tan, M.L.S.; Stevens, R.D.; Low, Y.L.; Muehlbauer, M.J.; Goh, D.L.M.; Ilkayeva, O.R.; Wenner, B.R.; Bain, J.R.; Lee, J.J.M.; et al. Insulin Resistance Is Associated with a Metabolic Profile of Altered Protein Metabolism in Chinese and Asian-Indian Men. Diabetologia 2010, 53, 757–767. [Google Scholar] [CrossRef]
- McCormack, S.E.; Shaham, O.; McCarthy, M.A.; Deik, A.A.; Wang, T.J.; Gerszten, R.E.; Clish, C.B.; Mootha, V.K.; Grinspoon, S.K.; Fleischman, A. Circulating Branched-Chain Amino Acid Concentrations Are Associated with Obesity and Future Insulin Resistance in Children and Adolescents. Pediatr. Obes. 2013, 8, 52–61. [Google Scholar] [CrossRef]
- Gleason, C.E.; Lu, D.; Witters, L.A.; Newgard, C.B.; Birnbaum, M.J. The Role of AMPK and MTOR in Nutrient Sensing in Pancreatic Beta-Cells. J. Biol. Chem. 2007, 282, 10341–10351. [Google Scholar] [CrossRef]
- Tajiri, K.; Shimizu, Y. Branched-Chain Amino Acids in Liver Diseases. World J. Gastroenterol. 2013, 19, 7620–7629. [Google Scholar] [CrossRef]
- Yoon, M.-S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human Gut Microbes Impact Host Serum Metabolome and Insulin Sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Shi, X.; Yin, H.; Li, J.; Huang, C.; Chen, Y.; Chen, Z.; Liu, W.; Su, W.; Zhang, Y.; Lin, M.; et al. Circulating Branch Chain Amino Acids and Improvement in Liver Fat Content in Response to Exercise Interventions in NAFLD. Sci. Rep. 2021, 11, 13415. [Google Scholar] [CrossRef]
- Merz, B.; Frommherz, L.; Rist, M.J.; Kulling, S.E.; Bub, A.; Watzl, B. Dietary Pattern and Plasma BCAA-Variations in Healthy Men and Women-Results from the KarMeN Study. Nutrients 2018, 10, 623. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial Ecology: Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef]
- Ecklu-Mensah, G.; Choo-Kang, C.; Gjerstad Maseng, M.; Donato, S.; Bovet, P.; Bedu-Addo, K.; Plange-Rhule, J.; Forrester, T.E.; Lambert, E.V.; Rae, D.; et al. Gut Microbiota and Fecal Short Chain Fatty Acids Differ with Adiposity and Country of Origin: The METS-Microbiome Study. Nat. Commun. 2023, 14, 5160. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CC Genotype of PNPLA3 | GG Genotype of PNPLA3 | ||||
|---|---|---|---|---|---|
| RD n = 28 | AD n = 20 | RD n = 20 | AD n = 20 | p Value | |
| Age (years) | 68.9 ± 4.5 | 66.2 ± 3.7 | 68.4 ± 4.5 | 67.2 ± 4.2 | 0.151 |
| BMI (kg/m2) | 27.8 ± 2.5 | 28.1 ± 2.4 | 25.8 ± 2.0 | 26.4 ± 2.3 | 0.004 *,2,3 |
| Waist (cm) | 101.5 ± 8.7 | 104.4 ± 8.0 | 94.6 ± 7.6 | 95.8 ± 7.5 | 0.00036 *,2,3 |
| GGT (U/L) | 32.8 ± 19.3 | 32.8 ± 21.1 | 20.1 ± 5.6 | 26.1 ± 9.9 | 0.026 *,2,4 |
| ALT (U/L) | 26.3 ± 13.3 | 27.7 ± 9.1 | 23.5 ± 12.9 | 27.7 ± 10.6 | 0.633 |
| AST (U/L) | 28.2 ± 7.4 | 27.6 ± 5.0 | 27.0 ± 6.9 | 27.0 ± 7.5 | 0.902 |
| Albumin (g/dL) | 38.3 ± 2.8 | 38.9 ± 2.2 | 38.0 ± 2.4 | 38.5 ± 3.2 | 0.763 |
| Total cholesterol (mmol/L) | 4.21 ± 0.97 | 4.66 ± 0.96 | 4.51 ± 0.80 | 4.58 ± 1.01 | 0.355 |
| HDL cholesterol (mmol/L) | 1.47 ± 0.54 | 1.29 ± 0.25 | 1.49 ± 0.37 | 1.41 ± 0.37 | 0.230 |
| LDL cholesterol (mmol/L) | 2.53 ± 0.77 | 3.04 ± 0.86 | 2.84 ± 0.78 | 2.93 ± 0.89 | 0.167 |
| Triglycerides (mmol/L) | 0.97 ± 0.37 | 1.28 ± 0.42 | 1.21 ± 0.79 | 1.12 ± 0.49 | 0.193 |
| Fasting glucose (mmol/L) | 5.71 ± 0.45 | 5.78 ± 0.35 | 5.58 ± 0.35 | 5.83 ± 0.41 | 0.229 |
| 120 min glucose (mmol/L) | 6.23 ± 1.46 | 6.31 ± 1.67 | 5.81 ± 1.46 | 5.96 ± 1.16 | 0.648 |
| Fasting insulin (mU/L) | 9.1 ± 5.4 | 14.0 ± 7.6 | 7.4 ± 3.5 | 9.1 ± 5.4 | 0.0031 *,1,4 |
| 120 min insulin (mU/L) | 63.2 ± 58.6 | 59.8 ± 40.6 | 40.9 ± 27.2 | 52.1 ± 46.3 | 0.387 |
| Hs-CRP (mg/L) | 1.01 ± 1.04 | 1.65 ± 1.45 | 0.71 ± 0.31 | 1.02 ± 0.87 | 0.032 * |
| PNPLA3 Genotype | Recommended Diet | p1 | p2 | Average Diet | p1 | p2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | GG | CC | GG | |||||||||
| Total, n | 28 | 20 | 20 | 20 | ||||||||
| Study Time | 0 | Inter. | 0 | Inter. | 0 | Inter. | 0 | Inter. | ||||
| Energy (kcal/day) | 2220 ± 425 | 2277 ± 444 | 2195 ± 402 | 2139 ± 423 | 0.893 | 0.209 | 2429 ± 406 | 2580 ± 454 | 2342 ± 510 | 2284 ± 399 | 0.683 | 0.057 |
| Protein (E%) | 16.4 ± 2.8 | 17.1 ± 2.3 | 17.5 ± 2.5 | 17.4 ± 1.6 | 0.313 | 0.209 | 16.3 ± 2.7 | 15.6 ± 1.8 | 16.8 ± 3.1 | 16.0 ± 2.3 | 0.039 * | 0.914 |
| Carbohydrate (E%) | 40.8 ± 5.5 | 41.3 ± 5.6 | 42.0 ± 6.1 | 43.0 ± 5.5 | 0.218 | 0.880 | 43.2 ± 4.8 | 40.9 ± 5.0 | 43.6 ± 4.8 | 40.7 ± 4.7 | 0.001 * | 0.327 |
| Fat (E%) | 38.3 ± 4.5 | 36.5 ± 3.5 | 36.2 ± 5.5 | 35.1 ± 4.8 | 0.032 * | 0.606 | 36.8 ± 4.8 | 38.9 ± 4.4 | 35.0 ± 5.1 | 38.0 ± 4.7 | 0.001 * | 0.327 |
| SFA (E%) | 13.2 ± 2.6 | 10.9 ± 1.9 | 12.3 ± 2.6 | 10.3 ± 1.8 | 2.47−8 * | 0.616 | 13.0 ± 1.5 | 16.6 ± 2.5 | 12.3 ± 3.0 | 15.7 ± 2.0 | 4.06−8 * | 0.778 |
| MUFA (E%) | 11.1 ± 2.4 | 14.6 ± 1.8 | 13.5 ± 2.6 | 14.2 ± 2.4 | 0.048 * | 0.750 | 13.3 ± 1.9 | 12.9 ± 1.4 | 12.2 ± 2.1 | 12.8 ± 2.1 | 0.673 | 0.175 |
| PUFA (E%) | 7.3 ± 1.7 | 7.7 ± 1.3 | 7.1 ± 2.0 | 7.5 ± 1.4 | 0.028 * | 0.798 | 7.2 ± 2.2 | 5.4 ± 0.8 | 6.9 ± 2.2 | 5.3 ± 0.7 | 1.20−6 * | 0.720 |
| Omega 3 (E%) | 1.84 ± 0.61 | 2.16 ± 0.44 | 1.83 ± 0.73 | 2.18 ± 0.49 | 0.000018 * | 0.671 | 1.78 ± 0.48 | 1.40 ± 0.23 | 1.7 ± 0.7 | 1.4 ± 0.2 | 0.004 * | 0.241 |
| Omega 6 (E%) | 5.0 ± 1.2 | 5.3 ± 1.0 | 5.1 ± 1.8 | 5.0 ± 1.0 | 0.210 | 0.522 | 5.0 ± 1.5 | 3.9 ± 0.5 | 4.8 ± 1.4 | 3.7 ± 0.5 | 0.000013 * | 0.980 |
| EPA (E%) | 0.056 ± 0.069 | 0.070 ± 0.045 | 0.064 ± 0.084 | 0.083 ± 0.056 | 0.00026 * | 0.996 | 0.047 ± 0.060 | 0.023 ± 0.021 | 0.068 ± 0.071 | 0.035 ± 0.030 | 0.178 | 0.830 |
| DHA (E%) | 0.146 ± 0.203 | 0.178 ± 0.137 | 0.170 ± 0.245 | 0.227 ± 0.162 | 0.00022 * | 0.957 | 0.127 ± 0.167 | 0.587 ± 0.542 | 0.175 ± 0.202 | 0.100 ± 0.793 | 0.324 | 0.886 |
| Fiber (g/day) | 29.3 ± 12.3 | 30.3 ± 10.2 | 31.8 ± 11.7 | 31.0 ± 10.8 | 0.709 | 0.224 | 31.2 ± 8.3 | 30.1 ± 9.3 | 29.6 ± 8.0 | 27.5 ± 7.2 | 0.064 | 0.717 |
| Recommended Diet | p1 | p2 | Average Diet | p1 | p2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | GG | CC | GG | |||||||||
| Week | 0 | 12 | 0 | 12 | 0 | 12 | 0 | 12 | ||||
| Plasma SCFA (µmol/g) | n = 28 | n = 20 | n = 20 | n = 20 | ||||||||
| Total SCFA | 245 ± 130 | 227 ± 54 | 232 ± 68 | 209 ± 85 | 0.041 * | 0.315 | 228 ± 48 | 235 ± 78 | 219 ± 63 | 208 ± 61 | 0.888 | 0.934 |
| Acetic acid (AA) | 179 ± 121 | 145 ± 31 | 162 ± 63 | 143 ± 41 | 0.120 | 0.879 | 155 ± 33 | 167 ± 59 | 161 ± 60 | 142 ± 61 | 0.420 | 0.211 |
| Propionic acid (PA) | 48 ± 27 | 64 ± 52 | 43 ± 31 | 42 ± 48 | 0.700 | 0.354 | 52 ± 31 | 50 ± 44 | 36 ± 27 | 29 ± 34 | 0.077 | 0.983 |
| Iso-butyric acid (IBA) | 7.3 ± 1.4 | 6.6 ± 2.0 | 7.3 ± 1.3 | 5.7 ± 2.3 | 0.002 * | 0.184 | 5.6 ±3.0 | 5.4 ± 2.5 | 7.3 ± 1.6 | 5.9 ± 2.5 | 0.613 | 0.399 |
| Butyric acid (BA) | 7.4 ± 1.0 | 7.3 ± 1.0 | 7.3 ± 1.3 | 5.7 ± 2.3 | 0.160 | 0.635 | 7.0 ± 0.7 | 7.2 ± 1.0 | 7.3 ± 0.7 | 6.9 ± 0.7 | 0.411 | 0.065 |
| Valeric acid (VA) | 3.7 ± 0.4 | 4.2 ± 1.7 | 3.6 ± 0.6 | 7.0 ± 0.6 | 0.0003 * | 0.004 * | 3.5 ± 0.6 | 4.3 ± 2.1 | 3.8 ± 0.3 | 9.7 ± 8.5 | 0.004 * | 0.079 |
| Plasma BCAA (µmol/g) | ||||||||||||
| Total BCAA | 572 ± 128 | 581 ± 182 | 599 ± 202 | 634 ± 154 | 0.688 | 0.280 | 612 ± 184 | 532 ± 149 | 587 ± 182 | 590 ± 130 | 0.015 * | 0.376 |
| Valine (VAL) | 258 ± 67 | 252 ± 90 | 266 ± 92 | 212 ± 95 | 0.009 * | 0.096 | 196 ± 100 | 214 ± 89 | 290 ± 75 | 211 ± 108 | 0.222 | 0.024 * |
| Leucine (LEU) | 136 ± 55 | 145 ± 85 | 141 ± 96 | 142 ± 59 | 0.936 | 0.662 | 141 ± 49 | 124 ± 71 | 162 ± 75 | 148 ± 61 | 0.043 * | 0.244 |
| Isoleucine (ILE) | 160 ± 23 | 153 ± 33 | 169 ± 65 | 157 ± 90 | 0.133 | 0.657 | 164 ±112 | 155 ± 31 | 167 ± 26 | 139 ± 37 | 0.238 | 0.081 |
| Fecal SCFA (µmol/g) | n = 26 | n = 20 | n = 18 | n = 19 | ||||||||
| Total SCFA | 52 ± 25 | 40 ± 24 | 48 ± 21 | 42 ± 26 | 0.010 * | 0.461 | 59 ± 22 | 57 ± 27 | 58 ± 25 | 60 ± 35 | 0.659 | 0.964 |
| Acetic acid (AA) | 32 ± 15 | 26 ± 15 | 32 ± 13 | 26 ± 16 | 0.007 * | 0.821 | 37 ± 12 | 35 ± 15 | 37 ± 15 | 36 ± 20 | 0.427 | 0.914 |
| Propionic acid (PA) | 8.7 ± 5.3 | 6.5 ± 3.6 | 6.9 ± 4.3 | 6.9 ± 4.6 | 0.304 | 0.145 | 9.6 ± 6.5 | 8.9 ± 7.4 | 9.2 ± 6.2 | 11.8 ± 3.4 | 0.882 | 0.340 |
| Iso-butyric acid (IBA) | 0.60±0.39 | 0.61±0.45 | 0.62±0.45 | 0.55±0.57 | 0.461 | 0.861 | 0.62±0.55 | 0.60±0.55 | 0.66±0.55 | 0.67±0.52 | 0.621 | 0.660 |
| Butyric acid (BA) | 9.5 ± 6.0 | 6.9 ± 5.0 | 8.0 ± 4.9 | 8.0 ± 6.4 | 0.028 * | 0.267 | 10.1 ± 4.5 | 11.2 ± 6.5 | 9.7 ± 4.9 | 10.1 ± 6.8 | 0.918 | 0.524 |
| Valeric acid (VA) | 1.04±0.59 | 1.00±0.53 | 0.91±0.53 | 0.81±0.66 | 0.339 | 0.894 | 1.32±0.99 | 1.29±0.71 | 1.28±0.95 | 1.35±1.18 | 0.725 | 0.808 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tauriainen, M.-M.; Csader, S.; Lankinen, M.; Lo, K.K.; Chen, C.; Lahtinen, O.; El-Nezamy, H.; Laakso, M.; Schwab, U. PNPLA3 Genotype and Dietary Fat Modify Concentrations of Plasma and Fecal Short Chain Fatty Acids and Plasma Branched-Chain Amino Acids. Nutrients 2024, 16, 261. https://doi.org/10.3390/nu16020261
Tauriainen M-M, Csader S, Lankinen M, Lo KK, Chen C, Lahtinen O, El-Nezamy H, Laakso M, Schwab U. PNPLA3 Genotype and Dietary Fat Modify Concentrations of Plasma and Fecal Short Chain Fatty Acids and Plasma Branched-Chain Amino Acids. Nutrients. 2024; 16(2):261. https://doi.org/10.3390/nu16020261
Chicago/Turabian StyleTauriainen, Milla-Maria, Susanne Csader, Maria Lankinen, Kwun Kwan Lo, Congjia Chen, Olli Lahtinen, Hani El-Nezamy, Markku Laakso, and Ursula Schwab. 2024. "PNPLA3 Genotype and Dietary Fat Modify Concentrations of Plasma and Fecal Short Chain Fatty Acids and Plasma Branched-Chain Amino Acids" Nutrients 16, no. 2: 261. https://doi.org/10.3390/nu16020261