

Unique Bioactives from Zombie Fungus (Cordyceps) as Promising Multitargeted Neuroprotective Agents

Department of Bionano Technology, Gachon Bionano Research Institute, Gachon University, 1342 Seongnam-daero, Sujeong-gu, Seongnam-si 461-701, Gyeonggi-do, Republic of Korea
*
Authors to whom correspondence should be addressed.
Nutrients 2024, 16(1), 102; https://doi.org/10.3390/nu16010102
Submission received: 17 November 2023
/
Revised: 8 December 2023
/
Accepted: 20 December 2023
/
Published: 27 December 2023
(This article belongs to the Special Issue Dietary Antioxidants and Oxidative Stress in Brain Health)
Abstract
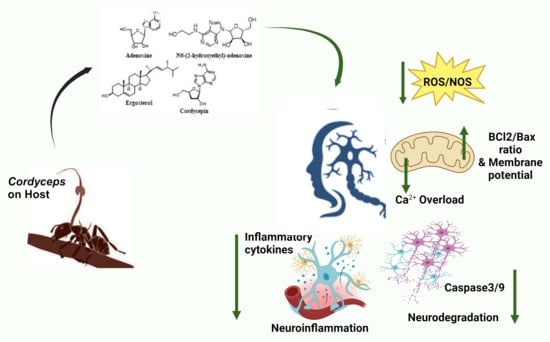
:Cordyceps, also known as “zombie fungus”, is a non-poisonous mushroom that parasitizes insects for growth and development by manipulating the host system in a way that makes the victim behave like a “zombie”. These species produce promising bioactive metabolites, like adenosine, β-glucans, cordycepin, and ergosterol. Cordyceps has been used in traditional medicine due to its immense health benefits, as it boosts stamina, appetite, immunity, longevity, libido, memory, and sleep. Neuronal loss is the typical feature of neurodegenerative diseases (NDs) (Alzheimer’s disease (AD), Parkinson’s disease (PD), multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS)) and neurotrauma. Both these conditions share common pathophysiological features, like oxidative stress, neuroinflammation, and glutamatergic excitotoxicity. Cordyceps bioactives (adenosine, N6-(2-hydroxyethyl)-adenosine, ergosta-7, 9 (11), 22-trien-3β-ol, active peptides, and polysaccharides) exert potential antioxidant, anti-inflammatory, and anti-apoptotic activities and display beneficial effects in the management and/or treatment of neurodegenerative disorders in vitro and in vivo. Although a considerable list of compounds is available from Cordyceps, only a few have been evaluated for their neuroprotective potential and still lack information for clinical trials. In this review, the neuroprotective mechanisms and safety profile of Cordyceps extracts/bioactives have been discussed, which might be helpful in the identification of novel potential therapeutic entities in the future.
1. Introduction
Neurodegenerative diseases (NDs) result from gradual loss of neuronal function, which ultimately causes cell death. These diseases remain incurable because of the complexity of brain function, leading to devastating neurological disorders. An increase in the incidence rate of NDs has been observed owing to rising life expectancy [1]. An imbalance between antioxidant and pro-oxidant species causes increased oxidative stress [2]. Increased reactive oxygen species (ROS)/reactive nitrogen species (RNS) in cells and tissues interfere with cellular mechanisms, such as mitochondrial dysfunction, leading to a decrease in energy production. Increased oxidative stress may lead to neuronal cell loss, a common feature of various NDs [2,3,4]. Oxidative stress, along with the production of cytokines, chemokines, and other secondary messengers, is the underlying cause of neuroinflammation. The expression levels of inflammatory cytokines positively correlate with the levels of neurotrophic factors [5]. Regulated inflammatory processes are important for maintaining tissue homeostasis and proper functioning, and extensive inflammation can lead to additional cell injury [6]. Neuroinflammation can be considered a double-edged sword, as it may cause damaging effects in NDs [7], while minor inflammation may be beneficial for recovery under some conditions [6,8,9,10]. Certain inflammatory inducers produce neurotoxic substances that amplify disease symptoms. These neurotoxic factors are associated with several NDs, like Alzheimer’s disease (AD), Parkinson’s disease (PD), multiple sclerosis (MS), and amyotrophic lateral sclerosis (ALS) [11,12]. Neuroinflammation is the major cause of ischemic stroke. M1-type microglial cells play a major role in prolonged inflammation, leading to brain tissue damage [13]. Although inflammation may not always be the initiating factor of these diseases, other sources of neuroinflammation include brain trauma, injury, and infection. Mitochondrial dysfunction, gliosis, and accumulation of abnormal protein aggregates also play critical roles in neurodegeneration [14]. Advanced glycation end products (AGEs) are formed by a non-enzymatic reaction (Maillard reaction) between the carbonyl group of reducing sugars and the amino group of amino acids. AGEs interact with the receptors of advanced glycation end products (RAGEs), initiating the activation of several pathways, including oxidative stress, and inflammation [15]. AGEs are also generated in food during thermal processing, like the pasteurization of dairy products [16,17] and immunomodulating gut bacteria. Nε-(carboxymethyl)lysine (CML) is an AGE that interacts with RAGEs and releases inflammatory cytokines, ultimately increasing oxidative stress [17]. Small-molecule therapeutics that interfere with AGE–RAGE interactions inhibit the inflammatory cascade and attenuate disease development [18]. Researchers worldwide have attempted to identify an effective cure for NDs. Despite several attempts to identify drugs that can reduce the symptoms of neurodegeneration, no permanent cure has been found. Hence, there is a critical need to identify bioactive compounds in nature and investigate their effects on neurodegeneration associated with NDs or neurotrauma.
Cordyceps, commonly known as the “caterpillar fungi”, “zombie fungus”, “Viagra of the Himalayas”, and “Yartsa gunbu”, belongs to the phylum Ascomycetes (Sac fungi). Cordyceps encompasses approximately 750 species that are distributed in different parts of the world, especially in temperate regions at altitudes over 3800 m. Cordyceps is the most expensive mushroom, with some species costing approximately USD 20,000/kg [19]. All Cordyceps species survive by invading their hosts with selective specificity. Most of these species parasitize insects and other arthropods, whereas a few invade Elaphomyces (Truffle genus) [20]. In brief, the life cycle of Cordyceps is initiated by spore germination, followed by the growth of hyphae by absorbing nutrients from the soil, invading the host, penetrating the host’s exoskeleton, and surviving in its tissues. After the host’s death, the fungus continues to grow, fruiting bodies sprout from the host’s head and release spores, and the cycle continues [21]. As this fungus manipulates the host system, the victim behaves like a “zombie”, with frequent convulsions; it is also called “zombie fungus” [22]. Due to these typical growth and survival tactics, these species synthesize promising bioactive metabolites, like anthraquinones, pyridines, cytochalasin, cyclic peptides, bioxanthracenes, polyketide, dihydrobenzofurans, glycans (β-glucans), alkaloids, phenols, flavonoids, terpenes, sterols (ergosterol), naphthoquinones, and nucleosides (adenosine, cordycepin) with broad therapeutic applications [23,24,25]. Cordyceps has been used in traditional Chinese medicine (TCM) health tonics [26] and for its medicinal properties, such as anticancer [27,28,29], antihyperglycemic [30,31], antifatigue [32], hepatoprotective [33], spermatogenic [34], hypolipidemic [35,36], antihypertensive [37,38], anti-inflammatory [39,40], nephroregulatory [41,42,43], antifibrosis [44], and immunomodulatory [45,46] properties. Although more than 750 Cordyceps species have been identified, only a few (C. militaris, C. ophioglossoides, C. sinensis, C. cicadae) have been studied for their neuroprotective activities.
Previous reviews of Cordyceps have focused on their nomenclature, structural elucidation, traditional use, and nutraceutical and pharmacological activities [23,34,46,47,48,49,50,51,52,53,54,55,56]. A bioactive compound (cordycepin) from this fungus has been a compound of choice among researchers, and most neuroprotective studies have been conducted using this compound, while information on other compounds is scarce. Hence, we present a comprehensive review of the neuroprotective mechanism and safety profile of the extracts and the active metabolites from Cordyceps investigated so far on neurotrauma and NDs.
2. Methods
This study presents a comprehensive review of the existing published scientific works from various databases (PubMed, Google Scholar, and Science Direct) until August 2023 on the key bioactive metabolites and neuroprotective potential of various species of Cordyceps to support its use in the treatment of searing diseases. The search terms used were “cordyceps” or “cordyceps bioactive” or “cordyceps metabolites” with the filter “neuroprotection”, “neurotrauma”, “neurodegenerative diseases”, “in vivo”, “in vitro” and “English”. Studies relevant to chemical analyses, medicinal applications, safety, toxicity, and neurodegeneration were selected.
3. Neuroprotective Potential of Cordyceps Extracts
Of all Cordyceps species, only 35 have been characterized [23], of which C. militaris and C. sinensis are the two most widely studied. C. sinensis is a rare and expensive species that is difficult to cultivate, whereas C. militaris is a successful commercially grown species and is considered an alternative to C. sinensis [57]. By changing the culture conditions, the concentrations of bioactive compounds can be manipulated.
Several extraction methods and solvents have been employed for the isolation of selective bioactive compounds [50,58], with each extract exhibiting specific activity. As polar molecules, the aqueous extract contains functional concentrations of nucleosides and polysaccharides. In contrast, alcoholic extracts are rich in nucleosides, polysaccharides, and proteins with a high antioxidant potential. Some of the bioactive components from Cordyceps species have been summarized in Table 1.
The neuroprotective mechanisms of the different types of Cordyceps extracts have been discussed below.
3.1. Cordyceps militaris
C. militaris is a valuable TCM that grows on moth larvae (Lepidoptera). This fungus has been reported to treat respiratory, renal, hepatic, and cardiovascular diseases and has antiaging, antiviral, anti-inflammatory, and antitumor potentials [59]. Recently, C. militaris has become an economical alternative to C. sinensis in TCM because it can be easily cultivated under artificial conditions using diverse media [57]. Analyses of the compositions revealed that the concentrations of cordycepin and polysaccharides in the media of cultured C. militaris were higher than those in C. sinensis from the natural site [60]. The major bioactive components of C. militaris are nucleosides (adenosine, uridine, and cordycepin), myriocin, ergosterol, polysaccharides, L-arginine, and L-proline [43,57,61]. Previous studies have shown the presence of GABA (γ-aminobutyric acid), ergothioneine, D-mannitol (cordysepic acid), glycolipids, glycoproteins, xanthophylls (like carotenoids), sterols, statins, phenolic compounds, vitamins, and biominerals in C. militaris [60,62]. A previous study reported differences in the concentrations of cordycepin, cordycepic acid, and ergothioneine between fruiting bodies and mycelial biomass. The concentrations of cordycepin, cordycepic acid, and carbohydrates are higher in mycelial biomass, whereas those of ergothioneine and total amino acids are higher in fruiting bodies [62]. The reported optimal drying temperature for C. militaris is 60 °C, over which, cordycepin and phenolic compounds are lost [63]. Pentostatin, used as an antileukemia drug, is also produced by C. militaris through the same biosynthetic gene cluster for cordycepin production [64]. Similar to other chemotherapeutic drugs, it also has side effects such as diarrhea, nausea, and neurological toxicities [65]. Cordymin is an antifungal peptide that inhibits the mycelial growth of various fungi, including Candida albicans, Bipolaris maydis, and Rhizoctonia solani [66]. Ergosta-7,9(11),22-trien-3β-ol isolated from C. militaris shows anti-inflammatory and antioxidative activity [67].
Selective deterioration of cholinergic neurons in AD diminishes acetylcholine (ACh) levels, contributing to cognitive decline [68]. In addition to acting as a neurotransmitter, ACh also induces neurite outgrowth [69,70]. The methanolic extract of C. militaris promoted neurite outgrowth and ACh expressions in Neuro 2A mouse neuroblastoma cells in a dose-dependent manner (5–20 µg/mL). It also reversed scopolamine-induced memory deficits in rats and increased central cholinergic function at a dose of 300 mg/kg [71]. The ethanolic extract has been known to promote neurite outgrowth in Neuro 2A cells [72], provide protection from amyloid beta (Aβ)-induced toxicity [73], reduce the expression of inflammatory markers (cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS)), and downregulate mitogen-activated protein kinases/c-Jun N-terminal kinase/extracellular signal-regulated kinase (MAPK/JNK/ERK) pathway in C6 glial cells [74], which helped to reduce stress, inflammation, and apoptosis [75]. In addition, the extract restores recognition and memory functions by inhibiting oxidative stress (nitric oxide (NO) and lipid peroxidation) caused by toxic peptides [76]. Moreover, it upregulated the dopaminergic system in vivo and in vitro by upregulating tyrosine hydroxylase, an enzyme that catalyzes the rate-limiting steps in the biosynthesis of dopamine and other catecholamines [77].
One of the most conspicuous age-related diseases is ischemia, which is a common form of neurodegeneration that leads to cognitive impairment in the elderly [78]. The post-ischemic brain induces hippocampal neuronal death, neuroinflammation, and neuropathy, similar to AD [79]. Post-ischemic treatment with the butanolic extract of the fungus (WIB-801C: 50 mg/kg) decreased the inflammatory cell infiltration into ischemic lesions by inhibiting chemotaxis through adenosine receptor A3 (A3AR), thus providing neuroprotection in the middle cerebral artery occlusion (MCAO) rat model [80]. Moreover, after spinal cord injury (SCI), it mitigated blood–spinal cord barrier (BSCB) disruption by inhibiting matrix metalloprotease-9 (MMP-9), downregulating the expression of chemokine and promoting that of pro-nerve growth factor (NGF) in microglia (MG) [81]. The fungus also improved memory impairment caused by global cerebral ischemia and memory deterioration by delaying neuronal death, decreasing MG expression in the CA1 region of the hippocampus in rats [82], and increasing the expression of brain-derived neurotrophic factor (BDNF) and tyrosine kinase B (TrkB) in gerbils [83].
C. militaris aqueous extract showed beneficial effects in a D-galactose (Gal)-induced aging mouse model by improving memory [84]. Extract supplementation improved the levels of antioxidants (superoxide dismutase (SOD), glutathione peroxidase (GPx), and glutathione (GSH)) and reduced malondialdehyde (MDA) and monoamine oxidase (MAO), which play important roles in the progression of aging. These results suggest a role for the antioxidant action of the fungus in recovering memory impairments in mice with D-Gal-induced aging [84].
In a recent study, nanoencapsulated C. militaris extract relieved neuronal pathology in SH-SY5Y cells (human neuroblastoma cells) by significantly improving dopamine secretion and the expression of dopaminergic-specific genes such as leucine-rich repeat kinase 2 gene (LRRK2), LIM homeobox transcription factor 1 beta (LMX1B), Forkhead Box (FOXA2), engrailed homeobox 1(EN1), and nuclear receptor-related 1 protein (NURR1) [85]. In line with this, C. militaris treatment enhanced the expression of neuronal protein paired box 6 (PAX6), a crucial player in brain development and function [86], and neuron-specific class III beta-tubulin (nestin), a marker of neuronal progenitor cells in the adult brain [87], indicating the role of C. militaris in enhancing neuronal maturation. Furthermore, it reduced amyloid precursor protein (APP) secretion by promoting autophagy [85]. As autophagy helps clear Aβ and tau aggregates in brain cells [88], C. militaris is considered important in AD treatment. The downregulated expression of AD-related genes presenilin 1 (PSEN1), presenilin 2 (PSEN2), and APP and the increased expression of the non-amyloidogenic pathway, ADAM metallopeptidase domain 10 (ADAM10), and sirtuin1 (SIRT1) by nanoencapsulated C. militaris extract suggest its potential in improving AD pathology at both the gene and protein levels [85].
These results suggest that the fungus is highly effective in protecting against memory-related neuronal degeneration in the brain and in retarding the progression of memory deficits associated with various NDs by its antioxidant, anti-inflammatory, and anti-apoptotic properties.
3.2. Cordyceps ophioglossoides
C. ophioglossoides, commonly known as the “golden thread Cordyceps”, is colonized on fruiting bodies of truffle-like Elaphomyces [89]. The fungus contains a variety of polysaccharides (antioxidant nature), ophiocordin (antibiotic), peptibiotics (antibiotic and antifungal properties), sesquiterpenes (antitumor activity), balanol (a protein kinase inhibitor with antitumor activity), and arsenocholine-O-sulfate (a nontoxic form of arsenic) [89,90,91,92,93].
Aβ(25–35) represents the biologically active region of Aβ, since it is the shortest fragment that displays large β-sheet aggregated structures, keeping the toxicity of the full-length peptide [94]; hence, it is often used as a model for inducing toxicity and memory deficits. The neuroprotective effect of C. ophioglossoides (methanolic extract) has been observed in vitro (extract: 100 μg/mL) and in vivo (extract:100 mg/kg) in Aβ(25–35) AD models, where the fungal extract protected SK-N-SH human neuroblastoma cells from cell death and helped in the restoration of spatial memory loss in induced memory deficit by Aβ(25–35) in rats probably by suppressing Aβ-induced oxidative stress [95].
3.3. Cordyceps sinensis
C. sinensis is the most popular Cordyceps, which parasitizes the larva of Hepialus armoricanus. This fungus has long been used in TCM to promote longevity and has anti-inflammatory and antitumor activities [24]. The major biochemical markers of nucleosides are adenosine and cordycepin [50,96], with immunomodulatory and antioxidant activities. In 2008, Yuan et al. reported the presence of other nucleosides (thymine, adenine, cytosine, uracil, uridine, hypoxanthine, ionosine, guanosine, and thymidine) in aqueous extracts of C. sinensis [97]. Polysaccharides are major contributors to the biological activities of C. sinensis. Guan et al. identified several monosaccharides (fructose, mannitol, galactose, arabinose, ribose, rhamnose, mannose, xylose, glucose, and sorbose) using GC-MS [98]. Ergosterol is the main identified sterol [99] and is present either as free or esterified ergosterol [50,100,101] with antitumor activity [102]. Other compounds, such as polyamines and free fatty acids, have also been identified in C. sinensis extracts [103]. Two peptides (cordymin and cordycedipeptide) and an ergosterol (H1-A) with biological activities were also isolated from the fungus [104].
Aqueous and different alcoholic extracts (CSEs) from the fungus revealed the presence of the antioxidants hesperidin, rutin, and ascorbic acid by high-performance thin-layer chromatography (HPTLC). Hesperidin, rutin, and ascorbic acid were present at high concentrations in the aqueous extract. However, the highest hesperidin content was observed in the 25% alcoholic extract in comparison to others [105]. Additionally, adenosine, adenine, and uracil are present at higher concentrations in the aqueous extract than in the other extracts [106]. The protective effects of the extracts against hypoxia-induced oxidative stress and inflammation were studied in mouse hippocampal (HT22) cells. CSEs (250 μg/mL) show neuroprotection by increasing the expression of endogenous antioxidants (GSH, GPx, and SOD), limiting lipid oxidation by decreasing MDA levels and reducing the level of inflammatory cytokines interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) as well as transcription factor nuclear factor-κB (NF-κB) to various extents. The aqueous extract is more effective as an antioxidant in hypoxia, whereas the alcoholic extract prevented oxidative stress and inflammation [105], owing to the presence of more phenolics and flavonoids [106].
The aqueous [107] and ethanolic [108] extracts of the fungus were also evaluated for anti-inflammatory effects in an experimental middle cerebral artery occlusion/reperfusion (MCAO/R) model, as ischemic brain injury is associated with inflammatory reactions. In addition, neuronal apoptosis is triggered by increased expression of Bcl2-associated X (Bax), an apoptosis regulator, which in turn activates caspase-3 and inhibits B-cell lymphoma-2 (Bcl-2) expression [109]. The extract provided neuroprotection by downregulating the expression of inflammatory cytokines and other inflammatory mediators (IL-1β, TNF-α, Myeloperoxidase (MPO), and adhesion molecules ICAM-1, COX-2, and iNOS), blocking polymorphonuclear cell (PMNC) infiltration, thereby subsiding neurological deficits and infarct volume. Moreover, the aqueous extract also revealed the decreased Bax, cytochrome c (Cyt c), and caspase-3 protein expressions, which in turn improved mitochondrial membrane potential (ψm), thus modulating the electron transport chain in the mitochondria in vivo and in vitro [110].
Multiple sclerosis (MS) is an autoimmune demyelinating disease of the CNS that involves a variety of immune cells [111]. Inflammation resulting from MS is mediated by the infiltration of autoreactive T cells into the CNS through the blood–brain barrier (BBB) [112]. Amongst the T cells, primarily interferon-gamma (IFN-γ)-producing T-helper 1 (Th1) cells and IL-17-producing Th17 cells had an important role in the pathogenesis of the disease [113]. Effector molecules secreted by Th1 cells directly affect the phenotype, function, and recruitment of MG, whereas Th17 cells upregulate chemokines during the inflammatory process [114]. C. sinensis extract was reported to reduce the number of Th1 cells in a mouse model of MS/experimental autoimmune encephalomyelitis (EAE), thus relieving EAE severity and the associated pathology [115].
The neuroprotective effect of fermented fungus powder (Cs-C-Q80 or ‘corbrin capsule’) was evaluated in subcortical ischemic vascular dementia induced in a mouse model of right unilateral common carotid artery occlusion (rUCCAO) [116], which damaged the white matter region in the brain, resulting in myelin loss, glial activation, neuroinflammation, and dementia [117]. However, both the prophylactic and therapeutic administration of corbrin (1 g/kg) significantly reduced white matter lesions and improved learning and memory loss through anti-inflammatory actions [116]. A lower dose of corbrin (1 mg/kg) was effective in reducing the pro-inflammatory cytokines (TNF-α, IL-1β, IL-6), improving the levels of oxidative stress parameters (SOD, MDA), increasing ATP concentration, and alleviating neurological deficits in an MCAO mice model [118].
3.4. Cordyceps cicadae
C. cicadae is the oldest known therapeutic fungus that feeds on Lepidoptera species larvae [119]. It has been used in TCM for the treatment of asthma, cancer, convulsions, dizziness, palpitations, and chronic renal disease. Natural C. cicadae is a slow-growing fungus in high demand, whereas its anamorph, Paecilomyces cicadae, can be cultured easily and used as a substitute for C. cicadae to accommodate market requirements [120]. Various bioactive compounds, such as cyclopeptides, myriocins, polysaccharides, nucleosides, and mannitol have been identified in C. cicadae [120,121]. LC-MS analyses have detected adenosine and adenosine analogs, N6-(2-hydroxyethyl)-adenosine (HEA), a Ca2+ antagonist, and an anti-inflammatory agent [122,123,124]. HEA is a major bioactive compound in C. cicadae that exhibits antidiabetic, sedative, analgesic, antitumor [125], and renoprotective activities [126,127]. Another isolated bioactive compound, ergosterol peroxide, exhibits immunomodulatory and anti-inflammatory effects [128,129].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Some important compounds from Cordyceps species and their biological activities.
| Bioactive Compound | Cordyceps Species | Chemical Class | Biological Activity | References |
|---|---|---|---|---|
| Adenosine | C. sinensis | Nucleoside | Prohibits cancer cell growth Anti-inflammatory effect | [103] |
| Cordycepin | C. sinensis C. militaris C. cicadae | Derivative of the nucleoside adenosine | Enhances immunity Anti-tumor activity Anti-inflammatory Antimicrobial activity | [57,103,121] |
| D-mannitol | C. militaris | Sugar alcohol | Diuretic effects | [57] |
| GABA | C. militaris | Primary amine | Neurotransmitter | [57] |
| Ergotheoneine | C. militaris C. cicadae | Thiourea derivative of histidine | Antioxidant | [57,121] |
| Lovastatin | C. militaris | Statin | Cholesterol-lowering agent | [57] |
| Uridine | C. militaris | Nucleoside | Maintenance of the cellular metabolism | [61] |
| N-(2-Hydroxyethyl) adenosine | C. cicadae | Derivative of the nucleoside adenosine | Anti-inflammatory activity | [121] |
| Cordycepic acid | C. cicadae | Sugar alcohol | Bacteriostatic activity | [103,121] |
| C. sinensis | Antioxidant | |||
| Beauvericin | C. cicadae | Cyclic hexadepsipeptide | Antimicrobial and antitumor activity | [121] |
| Methyl-2-(5-(3-Hydroxybutyl)furan-2-yl)acetate | C. cicadae | Furane methyl ester | Anti-AChE activity | [121] |
| α-furoic acid | C. cicadae | Carboxylic acid | Anti-AChE activity | [121] |
| 2-(5-(3-Oxobutyl)furan-2-yl) acetate | C. cicadae | Furane methyl ester | Anti-AChE activity | [121] |
| Hercynine | C. cicadae | Histadine derivative | Antioxidant | [121] |
| EPSF | C. sinensis | Polysaccharide | Antioxidant, antitumor | [103] |
| APS | C. sinensis | Polysaccharide | Antioxidant | [103] |
| CPS-1 | C. sinensis | Polysaccharide | Antioxidant | [103] |
| CPS-2 | C. sinensis | Polysaccharide | Inhibits cell proliferation | [103] |
| Ergosterol | C. sinensis | Phytosterol | Antimicrobial activity Cytotoxicity | [103] |
| Cordymin | C. sinensis | Peptide | Antidiabetic | [103] |
| Tryptophan | C. sinensis | Amino acid | Sedative effects | [103] |
Trauma to the CNS and NDs initiate a torrent of cellular and molecular reactions that result in neuronal loss and regenerative failure. To understand the associated mechanisms, the rodent optic nerve crush (ONC) model can be used and later extrapolated to NDs [130]. C. cicadae mycelium extract provided neuroprotection in the ONC rat model through anti-apoptotic and anti-inflammatory effects by improving retinal ganglion cell (RGC) density and P1-N2 amplitude [131], which intensified with visual–spatial attention in the visual cortex. The butanol fraction protected rat adrenal pheochromocytoma (PC12) cells against glutamate-induced oxidative damage. Additionally, the extract restored the mitochondrial function, suppressed ROS accumulation, upregulated the antioxidant enzymes (GPX and SOD), increased cell viability, decreased lactase dehydrogenase (LDH) release, and reduced apoptosis [132,133]. Subsequently, adenosine was identified as the main nucleoside responsible for this neuroprotective action [133]. The anti-inflammatory activities of three bioactive nucleosides (adenosine, cordycepin, and HEA) isolated from wild-type and artificially cultured C. cicadae were evaluated. Cordycepin was found to be more potent than other nucleosides in limiting the release of pro-inflammatory cytokines by lipopolysaccharide (LPS)-stimulated RAW 264.7; however, no synergistic effect of the three compounds was observed. LPS-induced pro-inflammatory responses were attenuated by HEA through the suppression of the toll-like receptor (TLR)-4-mediated NF-κB signaling pathway [134]. The effects of the hydroalcoholic fungal extract on cisplatin toxicity have also been evaluated. Cisplatin is an anticancer agent involved in multi-organ toxicity, including neurotoxicity. It accumulates in the dorsal root ganglion (DRG) and causes oxidative stress, neuronal apoptosis, and inflammation [135]. The nucleoside-rich extract of C. cicadae ameliorated memory impairment and neuropathy by reducing oxidative stress, acetylcholinesterase enzyme (AChE) levels, and inflammation in cisplatin-treated rats [136].
In a recent study, increased levels of bioactive compounds were obtained from cultured C. cicadae in deep ocean water (DOW) and minerals, thus increasing their therapeutic value [137]. The effect of DOW-cultured fungus (DCC) was investigated on D-Gal-induced brain damage and memory impairment in rats. DCC (100–500 mg/kg), in turn, improved cognition by alleviating the expressions of antioxidants and inflammatory genes (iNOS, TNF-α, IL-6, IL-1β, COX-2), along with reduced expressions of the aging-related proteins (glial fibrillary acidic protein (GFAP) and PS1) [137].
The neuroprotective activity of Cordyceps extracts has been summarized in Table 2.
Table 2.
Neuroprotective mechanism of Cordyceps extracts.
| Species | Extract | Study | Model | Study Outcome | Mechanism | Refs. |
|---|---|---|---|---|---|---|
| C. militaris | EtOH | in vitro, in vivo | Aβ(1–42)- induced toxicity in mice and C6 glial cells | Improved cognition, decreased NO and lipid peroxidation, downregulated COX-2 and iNOS, downregulated MAPK/JNK/ERK pathway | Antioxidant, Anti-inflammatory | [72,73,74,76] |
| C. militaris | MeOH | in vitro, in vivo | Neuro 2A, scopolamine-induced memory loss in rats | Promoted neurite outgrowth, increased ACh, improved memory | Increase ACh, neurogenesis | [71] |
| C. militaris | AQ | in vivo | Cerebral ischemia-induced short-term memory impairment in gerbils | Protected neuronal death Increased BDNF and TrkB expression | Anti-apoptotic Antioxidant | [83] |
| C. militaris | AQ | in vivo | D-Gal-induced aging mice | Increased SOD, GPx, GSH Decreased MDA Restoration of memory | Antioxidant | [84] |
| C. militaris | EtOH | in vitro, in vivo | PC12 cells and rat | Increased tyrosine hydroxylase | Upregulation of the dopaminergic system | [77] |
| C. militaris | BuOH | in vivo | MCAO-rat, scopolamine-induced memory loss in rats, spinal cord injury | Inhibited MMP-9, downregulated chemokines, delayed neuronal death | Anti-inflammatory | [80,81] |
| C. militaris | AQ | in vivo | Ischemia-induced death and cognitive impairment in rats | Decreased microglial expression Memory improvement | Anti-inflammatory | [82] |
| C. militaris | NP | in vitro | SH-SY5Y | Enhanced the expression of neuronal proteins Increased expression of dopaminergic-specific genes Decreased expression of PS1, PS2, APP Upregulated ADAM10 and SIRT1 Decreased Aβ secretion | Autophagy, neurogenesis, secretion of dopamine | [85] |
| C. ophioglossoides | MeOH | in vitro, in vivo | Aβ (25–35)-induced SK-N-SH and rats | Decreased oxidative stress Restored memory | Antioxidant | [95] |
| C. sinensis | AQ, EtOH | in vitro | Hypoxia-induced oxidative stress in HT22 | Increased SOD, GPx, GSH Decreased MDA, IL-6, TNF-α, NF-kB | Antioxidant Anti-inflammatory | [105] |
| C. sinensis | AQ, EtOH | in vivo | MCAO/R | Decreased IL-1β, TNF-α, MPO, ICAM-1, COX-2 and iNOS Suppressed PMNC infiltration | Anti-inflammatory | [107,108] |
| C. sinensis | AQ | in vitro, in vivo | MCAO/R | Decreased Bax, Cyt c, Caspase-3 | Anti-apoptotic | [110] |
| C. sinensis | - | in vivo | Mice mode of MS-EAE | Decreased Th1 | Immunoregulatory | [115] |
| C. sinensis | Fermented | in vivo | rUCCAO mice model | Reduced white matter lesion | Anti-inflammatory | [116] |
| C. sinensis | Fermented | in vivo | MCAO | Decreased TNF-α, IL-1β, IL-6 Increased SOD and ATP Decreased MDA Memory improvement | Antioxidant Anti-inflammatory | [118] |
| C. cicadae | - | in vivo | ONC rat model | Improved retinal ganglion cell density and P1-N2 amplitude | Antioxidant Anti-apoptotic | [131] |
| C. cicadae | BuOH | in vitro | Glutamate induced toxicity in PC12 cells | Increased GPx, SOD Increased cell viability, decreased LDH | Antioxidant Anti-apoptotic | [132,133] |
| C. cicadae | AQ, MeOH | in vitro | LPS-stimulated RAW 264.7 macrophages | Suppressed TLR-4-mediated NF-kB pathway | Anti-inflammatory | [134] |
| C. cicadae | HA | in vivo | Cisplatin-induced toxicity in mice | Reduced IL-6, TNF-α, and IL-1β; decreased AChE and oxidative stress | Antioxidant Anti-inflammatory | [136] |
| C. cicadae | DOW-cultured | in vivo | D-Gal-induced brain damage and memory impairment in rats | Decreased expression of GFAP, PS1 Decreased COX-2, TNF- α, IL-6, IL-1β | Antioxidant Anti-inflammatory | [137] |
Abbreviations: AQ: aqueous; MeOH: methanol; EtOH: ethanol; BuOH: butanol; HA: hydroalcoholic; NP: nanoparticle; DOW: deep Ocean water.
4. Neuroprotective Potential of Bioactive Compounds from Cordyceps
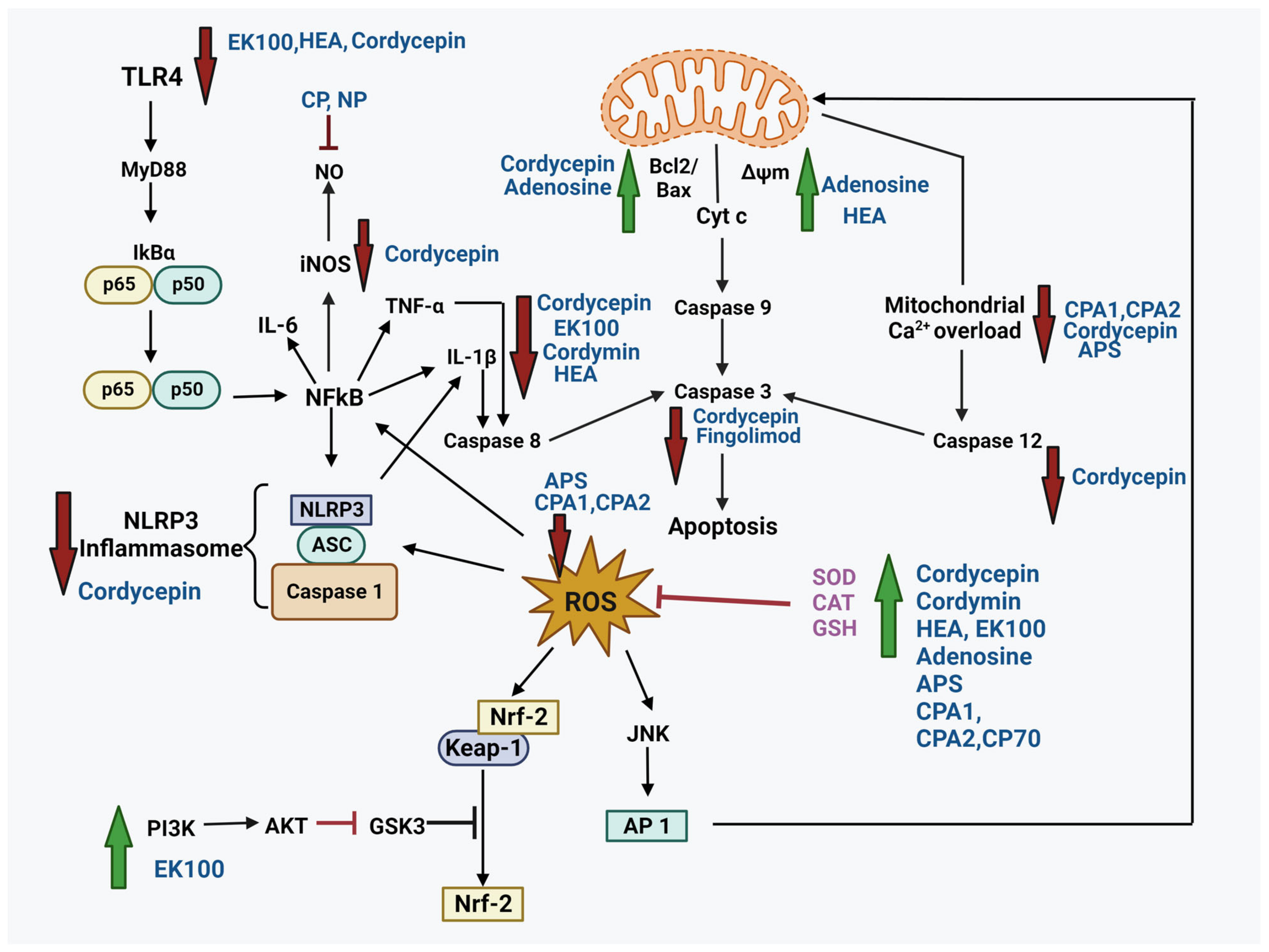
Cordyceps is a rich source of over 200 bioactive compounds, including nucleotides, nucleosides, polysaccharides, proteins, sterols, vitamins (Vit E, K, B1, B2, and B12), and trace elements [138]. These bioactive compounds are associated with a range of pharmacological activities, including antimicrobial, anti-allergic, antidiabetic, analgesic, anti-apoptotic, anticancer, anti-inflammatory, antioxidant, antiaging, and immunomodulatory effects [23,50,52]. However, few studies have investigated their neuroprotective activities. The mechanisms underlying the neuroprotective effects of these bioactive compounds (Figure 1) under various conditions are discussed below.
4.1. Cordycepin
Cordycepin (C10H13N5O3, MW 251.24 g/mol) is a bioactive nucleoside (3′-deoxyadenosine) from Cordyceps. It has several pharmacological properties, including anti-aging, anti-inflammatory, anticancer, and antioxidant properties [139,140,141,142,143]. Cordycepin is the most extensively studied bioactive compound in Cordyceps and its neuroprotective activities in different diseases are discussed below.
4.1.1. Neuroprotection in PD
PD is a progressive neurological disorder primarily affecting dopaminergic neurons in the substantia nigra region and is characterized by intracellular α-synuclein aggregates in the form of Lewy bodies and Lewy neurites. The biochemical processes implicated in PD include neuroinflammation, mitochondrial dysfunction, and faulty protein clearance [144].
The neuroprotective effects of cordycepin in PD have been investigated in several neurotoxin-induced models, namely, 6-hydroxydopamine (6-OHDA)-, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), and rotenone-induced models, to reiterate disease pathology both in vitro and in vivo. Isolated cordycepin from C. cicadae protected the PC12 cells against 6-OHDA-induced neurotoxicity by reducing caspase-3 activity, improving ψm, and elevating the levels of antioxidant enzymes [145], and finally, through an anti-apoptotic mechanism via Bax downregulation [146]. Furthermore, cordycepin protected dopaminergic neurons from death and inflammation by inhibiting dynamin-related protein-1 (Drp-1)-mediated (NOD)-like receptor protein 3 (NLRP3) inflammasome activation by increasing AMP-activated protein kinase (AMPK) phosphorylation in a rotenone-induced PD rat model and cultured PC12 cells [147]. Cordycepin alleviated MPTP-induced PD through TLR/NF-κB inhibition and mitigated the cytotoxic effects of MG on LPS-induced PC12 cells [148]. The anti-inflammatory role of cordycepin, along with the neuroprotection, was reported in LPS-induced MG activation in hippocampal cultured neurons (BV2) by assisting neural growth and development in the hippocampal neurons and downregulating the levels of TNF-α, IL-1β, iNOS, and COX-2, leading to an anti-inflammatory effect [149]. As TNF-α and IL-1β had a role in the activation of NF-κB, COX-2 was in turn activated by it [150], suggesting the involvement of the NF-κB pathway in the anti-inflammatory action of cordycepin. Therefore, constraining the TLR-4/NF-κB pathway and NLRP3 inflammasome activation would be valuable therapeutic targets for controlling pyroptosis and consequently advancing neurodegeneration in PD [151].
Glutamate is a key excitatory neurotransmitter that maintains cognitive, motor, and sensory functions, while GABA is an inhibitory neurotransmitter that maintains neuronal function. An imbalance between the glutamate and GABA synaptic systems results in impaired neural function that affects memory and cognition [152]. The extra synaptic diffusion of glutamate is strongly associated with MG activation and neuroinflammation, which are considered common characteristics in many NDs, including PD [153]. Moreover, hyperactivations of postsynaptic glutamate receptors α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) and N-methyl-d-aspartic acid (NMDA) were implicated in various neurological dysfunctions. Hence, the antagonists of these receptors have several beneficial effects in reversing motor symptoms [154]. Cordycepin suppressed AMPA and NMDA-receptor-mediated responses through a reduction in the presynaptic mechanism [155] and reduced the frequency of glutamatergic and GABA-ergic postsynaptic transmission without affecting the amplitude through A1AR activation [156]. This mechanism might provide neuroprotection against Aβ toxicity, hypoxia, ischemia, and other excitotoxic disorders. Cordycepin also protects against glutamate-induced oxidative toxicity in HT22 cells by downregulating the endoplasmic reticulum (ER) stress-specific caspase-12, which is important for the initiation of ER stress-induced apoptosis. Additionally, cordycepin inhibited the expression of pro-apoptotic genes (C/EBP homologous protein (CHOP) and Bax) and genes involved in ER stress-induced apoptosis (JNK, protein kinase R (PKR)-like ER kinase (PERK), and mitogen-activated protein kinases (p38)) [157]. Cordycepin also reduced ROS and Ca2+ levels. A1AR activation has been shown to mediate the neuroprotective effect of cordycepin [157,158].
Promising results from various studies have indicated the potential of cordycepin as a drug for the treatment of PD.
4.1.2. Neuroprotection in AD
AD is a multifactorial ND characterized by neuronal loss, accumulation of Aβ plaques, and neurofibrillary tangles. Aβ toxicity is accompanied by increased ROS production in neurons, which in turn leads to a series of events, like AChE activation, rise in Ca2+ concentration, mitochondrial dysfunction, and increased neuronal apoptosis resulting in cognitive deficit [159,160]. AChE hydrolyzes ACh, a neurotransmitter required for synaptic transmission, and, in turn, increases AChE activity, adversely affecting neurotransmission. The neuroprotective effect of cordycepin has been established in Aβ-induced rat hippocampal cells in the AD model [158]. Cordycepin effectively reduced ROS production and Ca2+ levels, inhibited AChE, suppressed apoptosis, and downregulated p-tau expression. These results suggest the involvement of A1AR in the neuroprotective activity of cordycepin.
Communication between neurons and MG in healthy brains and NDs has been studied extensively [161]. Two forms of active MG with antagonistic actions have been identified: MG-M1 (pro-inflammatory) and MG-M2 (anti-inflammatory); the MG-M1 form is prevalent in AD, leading to Aβ and tau accumulations, neuronal damage, and synaptic dysfunction [162]. Since MG polarization determines the fate of MG neurons in NDs, it is an efficient approach to fight AD. Recently, cordycepin was reported to induce MG-M2 polarization through the activation of cAMP-response element binding protein (CREB) activation and upregulation of NGF. This mechanism improved cognitive deficits in an APP/PS1 mouse model [163].
4.1.3. Neuroprotection in Ischemic Stroke
Cerebral ischemia/reperfusion injury (CI/RI) is a general feature of ischemic stroke, involving an interval of limited blood flow to the brain, followed by the refurbishment of blood supply through clinical intervention [164]. CI/RI results in neuronal injury or death, and irreversible brain damage due to oxidative stress, amino acid toxicity, Ca2+ overload, BBB dysfunction, inflammation, and apoptosis [165]. Cordycepin has been reported to exert protective effects in vitro and in vivo against CI/RI by restoring the levels of oxidative stress markers (MDA and SOD), reducing the levels of excitatory amino acids (glutamate and aspartate), and suppressing the expression of the inflammatory enzyme MMP-3 [139]. A study showed that a lower dose (10 mg/kg) of cordycepin was more effective in multiple myeloma cells, indicating that a higher concentration might induce cell death by preventing RNA synthesis [166] in astrocytes, reducing the number of astrocytic glutamate transporters and elevating extracellular glutamate levels. In addition, both C. militaris (water extract, 500 mg/kg) and cordycepin (10 mg/kg) reduced the levels of 4-hydroxynonenal (a lipid peroxidation marker) by reducing oxidative stress [167], delaying membrane depolarization, adjusting the electrophysiological activity of hippocampal CA1 neurons [168,169], and improving learning and memory [170] through A1AR activation [171].
4.1.4. Neuroprotection in Multiple Sclerosis
MS is a prominent neuroinflammatory autoimmune disorder characterized by the intrusion of immune cells from the perivascular region into the CNS. It is a demyelinating disease in which myelin is lost from various regions, leaving a scar (sclerosis) and disrupting signal transmission to and from the brain [172]. Cordycepin promoted remyelination by suppressing neuroinflammation (IL-6 and IL-1β) and upregulation of BDNF and anti-inflammatory cytokines (IL-4, IL-10, and TGF-β) in the corpus callosum and hippocampus in a cuprizone (CPZ)-induced mice model [173]. Cordycepin suppressed LPS-induced dendritic cell activations by mitigating oxidative stress through alleviating the protein kinase B/extracellular signal-regulated kinase/NFκB (AKT/ERK/NF-kB) signaling pathways in vitro and decreased the levels of migration/adhesion molecules (integrin β1, integrin α4, c-type lectin, intermolecular adhesion molecule-1 (ICAM-1), CC motif chemokine receptor 7 (CCR7)) in vitro and in vivo. Furthermore, cordycepin treatment in the experimental autoimmune encephalomyelitis (EAE) mice model decreased the level of chemokines (CC chemokine ligand 6 (CCL6), PAR response elements 2 (PARRES-2), IL-16, C-X-C motif chemokine ligand 10 (CXCL10), and cc motif chemokine ligand 12 (CCL12)) in the CNS and spinal cord and inhibited the production of pro-inflammatory cytokines (IFN-γ, IL-6, TNF-α, and IL-17) in activated microglial cells, macrophages, and Th cells in vitro [174], thus potentially ameliorating MS progression.
In summary, cordycepin ameliorates motor dysfunction, improves remyelination, decreases the number of glial cells, suppresses pro-inflammatory cytokines, increases anti-inflammatory cytokines and neurotrophic factors, and reduces oxidative stress. Consequently, it is a potential candidate for treating demyelination-associated diseases such as MS.
4.1.5. Neuroprotection in Traumatic Brain Injury
Traumatic brain injury (TBI) leads to serious neurological dysfunctions that affect motor and cognitive functions. Additionally, white matter injury (WMI) resulting from secondary TBI is extremely vulnerable to neuroinflammation owing to a vicious cycle caused by the penetration of neuroinflammatory receptor immune cells. TLR-4 is expressed on astrocytes, microglia, and neurons and is activated upon infection, brain injury, BBB disruption, and many NDs, including AD, PD, and ALS [175,176]. Another important protein is MMP-9, which contributes to BBB disruption and edema after TBI [177]. Hence, therapies directed toward these targets are beneficial.
The administration of cordycepin in TBI mice (10 mg/kg) [178] and rats (20 mg/kg) [179] ameliorated long-term neurological deficits, as observed in behavioral tests. Immunohistochemical staining indicated that cordycepin secured the number and structure of nonmyelinated and myelinated axons, thereby enhancing their conductive abilities. Cordycepin administration also decreased the levels of pro-inflammatory markers: cluster of differentiation-16 (CD16), interleukin-17α (IL-17α), IL-1β, iNOS, and MPO, with an upregulation of anti-inflammatory ones (CD-206, IL-10, Arginase-1). Moreover, it elevated the expression of tight-junction proteins (zonula occludens-1 (ZO-1) and occludin) and reduced the activity of MMP-2 and MMP-9, preserving the integrity of the BBB [178,179]. Additionally, cordycepin inhibits NADPH oxidase (NOX1), which is the main contributor to ROS, by disrupting the BBB after TBI [179].
As the brain is sensitive to oxygen levels, hypobaric hypoxia (HH) could lead to neuronal death and neuropsychological dysfunction [180]. HH induces oxidative stress and neuroinflammation, which eventually disturb the integrity of the BBB. Early HH followed by TBI increases the severity of secondary brain injury [181]. Cordycepin (10 mg/kg) inhibits the hippocampus-dependent memory impairment caused by acute HH, relieves hyperactivation of astrocytes/microglia in the CA1 region, and mitigates HH-induced activation of the TLR-4/NF-κB neuroinflammation in the rat model. In addition, it conserved BBB integrity by repressing MMP-9 expression and increasing the levels of tight-junction proteins (claudin-5, occludin, and ZO-1) in the hippocampus [182].
From the above studies, it is evident that cordycepin provides neuroprotection through antioxidant and anti-inflammatory mechanisms via A1AR activation. Additionally, it enables the presynaptic suppression of excitatory synaptic transmission by limiting the release of excitatory neurotransmitters, a novel system for modulating CNS activity.
The mechanism of neuroprotection displayed by cordycepin in various conditions has been summarized in Table 3.
Table 3.
Neuroprotective mechanism of cordycepin.
| Disease | Study Model | Mechanism | MOA | Refs. |
|---|---|---|---|---|
| PD | 6-OHDA-induced neurotoxicity in PC12 cells | Decreased caspase-3, increased SOD and ψm | Antioxidant activity | [145] |
| Rotenone-induced toxicity in rat model | Decreased Bcl2 expression, increased ψm, decreased caspase-3 | Anti-apoptotic Antioxidant | [146,147] | |
| MPTP-induced PD in rats and PC12 cells | Suppressed TLR4/NF-κB pathway | Anti-inflammatory | [148] | |
| Glutamate-induced oxidative toxicity in HT22 cells | Downregulated caspase-12 Deceased expression of CHOP, Bax, JNK, PER, p38 Reduced ROS and Ca2+ | Anti-apoptotic, Antioxidant, A1AR activation | [157] | |
| LPS-induced BV2 cells | Neurogenesis Downregulated TNF-α, IL-1β, iNOS, Cox2 | Anti-inflammatory Neurogenesis | [149] | |
| LPS-treated C57BL/6J mice and BV2 cells | Suppressed TLR4/NF-κB-mediated NLRP3 inflammasome activation and GSDMD-related pyroptosis Inhibited pore formation in the plasma membrane Reduced the release of pro-inflammatory mediators | Anti-apoptotic Anti-inflammatory | [151] | |
| Hippocampal brain slice from rats | Reduced excitatory synaptic transmission | Synaptic transmission | [155] | |
| AD | Aβ-induced toxicity in primary hippocampal neurons | Downregulated pTau, anti-AChE, reduced ROS and Ca2+ | Anti-apoptotic Antioxidant Enzyme inhibition A1AR activation | [158] |
| APP/PS1 mice model | Microglia/macrophage polarization through CREB | Neurogenesis | [163] | |
| Ischemic Stroke | OGD model | Increased SOD Decreased MDA Suppressed Glu and Asp Decreased MMP3 | Antioxidant | [139] |
| Ischemic damage in gerbils; β-amyloid and ibotenic acid-induced hippocampal CA1 pyramidal neuronal hyperactivity | Reduced 4-hydroxynonenal, delayed membrane depolarization | Antioxidative A1AR activation | [167,168,169,170] | |
| Acute hypobaric hypoxia-induced BBB disruption and cognitive impairment in rats | Increased tight-junction proteins (claudin5, occluding, zonula occludens-1) Inhibited TLR-4/NF-κB/MMP-9 pathway | Anti-inflammatory Antioxidant | [182] | |
| MS | LPS-induced dendritic cells, MS-EAE mice model | Inhibited AKT/ERK/NF-kB pathway Decreased integrin (β1,α-4), c-type lectin, ICAM1, CCR7 Decreased chemokines Decreased INF-γ, IL-6, IL-17, TNF-α | Antioxidant, Anti-inflammatory | [174] |
| CPZ-induced demyelination in mice | Decreased IL-6, IL-1β Increased IL-4, IL-10, and TGF-β Upregulated BDNF Promoted remyelination | Anti-inflammatory | [173] | |
| TBI | TBI-mice, rats | Decreased MMP-2, MMP-9; CD-16, IL-17, NOX1, MPO, iNOS Increased ZO-1, CD-206, IL-10, IL-1β, Arginase-1 Suppressed neutrophil infiltration | Anti-inflammatory Antioxidant | [178,179] |
The neuroprotective mechanisms of other bioactive compounds from Cordyceps have been discussed below.
4.2. N6-(2-Hydroxyethyl)-Adenosine (HEA)
N6-(2-hydroxyethyl)-adenosine (HEA), a bioactive nucleoside (C12H17N5O5; MW 311.29 g/mol), was identified from the butanolic fraction of C. cicadae. It is a calcium antagonist and interacts with human serum albumin [125,127]. HEA is a potent antioxidant with glucose-lowering, hepatoprotective, cardioprotective, sedative, antitumor, eye-protective, and anti-inflammatory properties [126,134,165,183,184].
Limited literature is available on the neuroprotective activities of HEA. HEA (5–40 μM) protected against H2O2-induced oxidative stress in PC12 cells by increasing cell viability, decreasing LDH release, preventing ψm breakdown, limiting ROS generation, inhibiting lipid peroxidation, and reducing inflammatory cytokines (IL-6, IL-1β, TNF-α, and NF-κB) [185]. HEA alleviated the pro-inflammatory response in RAW264.7 macrophages by suppressing the TLR-4/NF-κB pathway in LPS-induced inflammation [134].
4.3. Adenosine
Adenosine is a pentose sugar bonded to adenine, that is, adenine riboside (C10H13N5O4; MW 267.24 g/mol). It is a neuromodulator of the CNS that primarily operates via the adenosine A1 receptor (A1R). A1R activation plays a neuroprotective role by modulating the Gα/cAMP/PCK pathway, enhancing synaptic plasticity, memory, and cognition [186].
Excessive glutamate triggers intracellular Ca2+ influx by activating NMDA receptors and enhancing mitochondrial oxidative stress, which in turn activates mitochondria-associated apoptotic proteins. Furthermore, the upregulation of MAPK during oxidative stress is associated with apoptosis in several NDs. However, adenosine isolated from C. cicadae protected PC12 cells from glutamate-induced apoptosis by increasing the Bcl-2/Bax ratio, decreasing oxidative stress (increasing GPX, SOD, ψm), and alleviating Ca2+ overload (via decreasing p38/JNK/ERK phosphorylation) [133]. Hence, adenosine is a promising pharmacophore for the treatment of NDs.
4.4. Polysaccharides
Polysaccharides obtained from Cordyceps are also important bioactive compounds with a wide range of activities, including immunomodulatory, antioxidant, and antitumor activities [187,188,189,190,191].
Polysaccharides from the fruiting bodies of C. militaris were evaluated for their protective effects in a D-Gal-induced aging mouse model [192], where they protected mitochondrial integrity by scavenging free radicals and increasing the activity of the antioxidant enzymes responsible for aging. Heterogeneous polysaccharides (CPA-1 and CPA-2), mainly composed of mannose, glucose, and galactose, isolated from C. cicadae provided neuroprotection against glutamate-induced toxicity in PC12 cells [193] by increasing cell viability and the levels of antioxidant enzymes (GSH-Px and SOD) and reducing LDH release, ROS, and Ca2+ levels through their antioxidant action. A similar neuroprotective effect was observed in H2O2-treated PC12 cells by acid polysaccharides (APS) from C. sinensis [194,195]. Furthermore, crude polysaccharides (CPs) and non-digestible polysaccharides (NPs) isolated from cultivated C. cicadae exhibited anti-inflammatory activities in LPS-induced RAW264.7 macrophages [196]. Compared to CPs, NPs displayed better inhibition of NO, TNF-α, and IL-1β production in LPS-stimulated cells. The different conformations and molecular weights of the two polysaccharides may be responsible for the variations in their activities. Additionally, CP70, a polysaccharide from C. cicadae, displayed anti-aging activity and extended the lifespan of Drosophila by upregulating the expression of antioxidant enzymes (catalase (CAT) and SOD) [197].
In summary, the polysaccharides from Cordyceps provide neuroprotection through antioxidant and anti-inflammatory mechanisms.
4.5. Ergosta-7, 9 (11), 22-Trien-3β-ol (EK100)
EK100 (C28H44O; MW 396.64 g/mol) is a derivative of fungal ergosterol. Ergosterols are the active components of Cordyceps and have important therapeutic activities, such as analgesic, antimicrobial, antitumor, antioxidant, anti-inflammatory, antidiabetic, antihyperlipidemic, and immunomodulatory activities [129,198,199,200,201,202,203].
In the AD model of Drosophila with the pan-neuronal overexpression of human Aβ, EK100 improved the life span, motor functions, and memory by modulating MG activation, only without having any effect on the oxidative stress markers [204]. MG-mediated innate immunity is a double-edged sword, particularly in AD. Activated MG clears Aβ but inevitably damages neurons in the microenvironment [205]. A decline in innate immunity (MG activation) reduces its impact on Aβ clearance, hence leading to Aβ deposition and generating oxidative stress.
During chronic inflammation, the MAPK/activator protein (AP-1) pathway plays a critical role in the release of pro-inflammatory cytokines. Hence, controlling LPS-induced TLR-4/NF-kB/MAPK may benefit cells. The anti-inflammatory potential of ergosterol EK100 isolated from C. militaris [206] was studied in LPS-induced RAW264.7 cells. EK100 (80 μM) significantly reduced the cytokine releases and the levels of pro-inflammatory mediator proteins, attenuated phosphatidylinositol-3-kinase (PI3K)/Akt phosphorylation, inhibited the TLR-4/myeloid differentiation factor 88 (MyD88)/IκB kinase (IKK) inflammatory signaling pathway, and suppressed the nuclear translocation of p65 and p50 in the treated cells. Molecular docking studies revealed that EK100 restricted docking of LPS to the LPS-binding protein (LBP), CD14, and TLR-4/myeloid differentiation-2 (MD-2) co-receptors and finally suppressed the TLR-4/NF-kB inflammatory pathway. Moreover, EK100 not only modulated the LPS/TLR-4-related MAPK/AP-1-induced inflammatory pathway but also activated nuclear factor erythroid 2-related factor (Nrf2)/heme oxygenase-1(HO-1) antioxidative signaling by increasing the levels of antioxidant enzymes (GPX, SOD, and CAT) [67].
At present, recombinant tissue plasminogen activator (rtPA) is the only standard treatment for ischemic stroke, which damages the brain tissue through brain infarction and inflammation, [207]. Recently in a study, EK100 (30–120 mg/kg) supplementation ameliorated ischemic stroke brain injury in mice [208]. The combination of EK100 (60 mg/kg) and r-tPA (10 mg/kg) enhanced the protective action compared to either of them alone. Reduced levels of inflammation and apoptosis markers (p65NF-κB and caspase-3) and upregulation of neurogenesis protein (doublecortin) by EK100 via PI3K/AKT activation, glycogen synthase kinase-3 (GSK-3) inhibition, and β-catenin upregulation were observed [208]. GSK-3 inhibition has already been reported as an effective neuroprotective approach to ischemic stroke [209].
Intracerebral hemorrhage (ICH) is a neurological disorder characterized by intensified excitotoxicity, neuroinflammation, and apoptosis in the damaged brain tissue. The upregulation of COX-2 induces an inflammatory cascade by activating specific prostaglandin receptors post-ICH [210]. Furthermore, oxidative stress and MMP-9 activation have deleterious effects on brain injury as they promote DNA damage, apoptosis, and edema [211]. Hence, treatments directed towards controlling the expression of COX-2 and MMP-9 may be beneficial for ICH. EK100 was reported to inhibit JNK/MAPK activation and COX-2 and MMP-9 expression in vitro (BV2 cells) and in vivo (collagenase-induced ICH mice) and improved brain edema and neurobehavioral defects in ICH mice [212].
Therefore, EK100 is a promising novel dual strategy for the treatment of inflammatory diseases that not only suppresses inflammatory transcription factor signaling but also activates the antioxidative transcription factor signaling pathway.
4.6. Cordymin
Cordymin is a 10.9 kDa weight fungal peptide with an N-terminal sequence of AMAPPYGYRTPDAAQ, isolated from Cordyceps [66,213]. It is reported to possess anticancer, antidiabetic, anti-inflammatory, and antinociceptive activities [214,215].
Ischemia-reperfusion (IR) injury is a common characteristic of ischemic stroke resulting from restoration of blood supply after ischemia. It results in the release of inflammatory cytokines and free radicals, resulting in apoptosis [216]. In the cerebral ischemia-reperfusion injury rat model, cordymin (1–4 mg/kg) protected the ischemic brain by elevating antioxidant activity (through increasing GSH, reducing lipid peroxidation), decreasing inflammation (downregulating IL-1β and TNF-α and C3 protein), and repressing infiltration of polymorphonuclear cells (PMNCs) in the lesion [213]. Consequently, cordymin can be used as a promising protective agent against IR injury.
4.7. Active Polypeptide
Active polypeptides are specific protein fragments that have a positive effect on health. Bioactive polypeptides can be directly absorbed into the system and serve as carriers. C. militaris polypeptides improve immunity and exhibit antioxidant properties [217,218].
The effect of Cordyceps polypeptide was studied in a scopolamine-induced memory-impaired mouse model, where it improved the condition through antioxidant action (increased SOD and decreased MDA) and retarded AChE activity in the mouse brain. It also elevated the activity of the sodium–potassium pump (Na+-K+-ATPase), which is involved in the energy supply, and upregulated the expression of GABA and glutamate, which are the central inhibitory and stimulatory neurotransmitters, respectively. Moreover, the polypeptide increased the gene expression of Slc18a2 (secretion of neurotransmitters) with a concomitant decrease in the expression of Pik3r5 (cell proliferation and apoptosis) and Il-1β (pro-inflammatory) [219].
4.8. Fingolimod
Fingolimod (FTY-720; C19H33NO2; MW 307.47 g/mol) is a synthetic analog of myriocin (C21H39NO6; MW 401.54 g/mol), which is a non-proteinogenic fungal amino acid.
Myriocin ((2S, 3R, 4R)-(E)-2-amino-3,4-dihydroxy-2-(hydroxymethyl)-14-oxoeicos-6-enoic acid) was isolated from Isaria sinclairii (imperfect stage of C. sinclairii) almost 30 years ago [220]; it shows strong in vitro immunosuppressive activity; however, it induces toxicity in vivo. Extensive chemical modification of the myriocin structure generated fingolimod [FTY-720; (2- amino-2-[2-(4-octylphenyl)ethyl]propane-1,3-diol)], which has stronger immunosuppressive activity and less toxicity than myriocin [221]. The structure of fingolimod is closely related to that of sphingosine; hence, fingolimod is phosphorylated by sphingosine kinases. Phosphorylated fingolimod depletes lymphocyte circulation by activating the sphingosine-1-phosphate receptor (S1PR) and displaying potent immunosuppressive activity [222,223,224]. As S1PR is located in the CNS, the neuroprotective properties of FTY720 have been reported in experimental models of AD and PD [225,226] and granted with US Food and Drug Administration (FDA) approval as the first oral drug to reduce MS relapse [227,228]. Recently, FTY720 was found to exert neuroprotection in CI/RI by reducing the protein levels of IL-17α in the glial cells and reducing inflammatory reactions in the brain. Moreover, it protected the entire neurovascular unit by reducing the infarct volume, protecting the BBB, improving neurological deficits, and reducing apoptosis in the neurons [229].
The mechanism of neuroprotection exerted by bioactive compounds from Cordyceps has been summarized in Table 4.
Table 4.
Neuroprotective potential of other bioactive components from Cordyceps.
| Name | Nature | Study | Model | Study Outcome | Mechanism | Refs. |
|---|---|---|---|---|---|---|
| N6-(2-hydroxyethyl)-adenosine | Nucleoside | in vitro | H2O2-induced oxidative stress in PC12 cells | Reduced IL-6, IL-1β, TNF-α and NF-kB Reduced LDH release, increased Ψm | Antioxidant Anti-inflammatory | [185] |
| in vitro | LPS-induced inflammation in RAW264.7 macrophages | Decreased pro-inflammatory cytokines by suppressing TLR-4/NF-kB pathway | Anti-inflammatory | [126] | ||
| Adenosine | Nucleoside | in vitro | Glutamate-induced toxicity in PC12 cells | Increased GSH-Px and SOD Increased Bcl-2/Bax ratio Reduced the expression of ERK, p38, and JNK, increased Ψm | Antioxidant Anti-inflammatory Anti-apoptotic | [133] |
| Mixture | Polysaccharide | in vivo | D-Gal-induced aging mice model | Decreased ROS Increased antioxidant enzymes Protected mitochondria | Antioxidant Anti-aging | [84] |
| CPA-1, CPA-2 | Polysaccharide | in vitro | Glutamate-induced toxicity in PC12 cells | Increased cell viability; ncreased GSH-Px, and SOD Reduced LDH release, ROS, and Ca2+ levels | Antioxidant | [193] |
| CP, NP | Polysaccharide | in vitro | LPS-induced inflammation in RAW264.7 macrophages | Inhibited NO, IL-1β, TNF-α | Anti-inflammatory | [196] |
| CP70 | Polysaccharide | in vivo | Drosophila | Increased CAT, SOD expression | Antioxidant Anti-aging | [197] |
| APS | Polysaccharide | in vitro | H2O2-induced stress in PC12 | Increased cell viability; increased GSH-Px, and SOD Reduced LDH release, ROS, and Ca2+ levels | Antioxidant | [194] |
| Ergosta-7, 9 (11), 22-trien-3β-ol | Ergosterol | in vivo | Drosophila AD model | Reduced microglia activation and inflammatory markers | Anti-inflammatory | [204] |
| in vitro | LPS-induced RAW264.7 and BV2 cells | Reduced the cytokine release and pro-inflammatory markers Suppressed TLR4/NF-kB pathway, activated Nrf2/HO-1 pathway | Antioxidant Anti-inflammatory | [67,206] | ||
| in vivo | Ischemic stroke brain injury in mice | Increased neurogenesis, upregulated PI3K/AKT pathway | Anti-inflammatory Anti-apoptotic | [208] | ||
| in vivo, in vitro | Collagenase-induced ICH in mice, BV2 cells | Downregulated MMP-9, COX-2 | Anti-inflammatory | [212] | ||
| Cordymin | Peptide | in vivo | Ischemic stroke brain injury in mice | Elevated GSH Reduced MDA, IL-1β, TNF-α Reduced infiltration of PMNCs | Antioxidant Anti-inflammatory | [213] |
| Active polypeptide | Peptide | in vivo | Scopolamine-induced memory impairment in mice | Increased SOD, Na-K-ATPase Decreased MDA and AChE Increased secretion of neurotransmitters | Antioxidant Anti-inflammatory Anti-apoptotic | [219] |
| Fingolimod | Myriocin synthetic analog | in vivo, in vitro | Focal CI/RI in the rat, mice PD model 6-OHDG Rotenone-induced SH-SY5Y Cells | Protected BBB Improved neurological deficits Reduced IL-17 Reduced caspase-3 expression | Immunosuppressant Anti-inflammatory Anti-apoptotic | [225,226,229] |
As the common molecular mechanism in NDs and neurotrauma involves the interplay of oxidative stress, neuroinflammation, and programmed cell death, a combination therapy has become utterly important. Henceforward, the bioactive compounds from the zombie fungus Cordyceps are the potential candidates in the treatment of such pathological conditions. They affect various pathways (Figure 2) involved in neuroprotection by alleviating ROS, reducing inflammation, restoring mitochondrial dysfunction, limiting apoptosis, and improving levels of antioxidant enzymes.
5. Safety and Toxicity
Cordyceps is a medicinal mushroom widely used as a health supplement. Like all other supplements, the US FDA has not approved it for safety concerns, but Cordyceps and its two dietary supplements have been approved by the Chinese National Medical Products Administration (NMPA) [230]. Cordyceps is relatively safe but may cause allergies, nausea, and stomach aches in some cases. Certain medications, such as antidiabetic, antithrombotic, and anticancer drugs [31,231,232], may interact with Cordyceps; hence, it is advised to consult a doctor before consuming them. The most prominent concern is the accumulation of heavy metals, especially arsenic, in naturally growing mushrooms in contaminated soils. The recommended dose of C. sinensis is 4 g/day for no more than 5 months [233]. Moreover, it is not recommended for patients with chronic kidney disease because it affects renal function [26]. As per the reports, cordycepin resulted in gastrointestinal and bone marrow toxicity in dogs [234,235], oosporein caused gout in avian species [234,236], and beauvericin might induce apoptosis and cell cytotoxicity [237]. Clinical trials have been conducted to investigate Cordyceps for immunomodulation [238,239,240,241], anticancer activity [242], respiratory function enhancement [243], insomnia management [244], prostate problem management [245], liver dysfunction alleviation [246], and increased tolerance to high-intensity exercise [32,247,248]. However, no clinical studies have investigated its neuroprotective activity.
6. Conclusions and Future Directions
Similarities in the pathophysiological mechanisms have been implicated in neurotrauma and neurodegenerative diseases. Both conditions are associated with oxidative stress, neuroinflammation, and glutamatergic excitotoxicity. The cascade of reactions can lead to apoptosis, necrosis, Ca2+ overload, protein aggregation, and mitochondrial dysfunction, affecting various signaling pathways (PI3K/AKT, TLR-4/NF-kB, Nrf/HO-1, ERK/p38/JNK] and eventually causing amnesia and brain damage.
Recent studies have demonstrated the role of dietary interventions in the control of oxidative stress, a key regulator of the pathogenesis of several diseases. Dietary components are known to boost mood, memory, and several other brain processes by maintaining a healthy oxidative state and improving synaptic function and neuronal plasticity. In this context, various studies validated the effectiveness of Cordyceps bioactives in protection against memory-related neuronal degeneration by antioxidant, anti-inflammatory, and anti-apoptotic properties. They exhibit anti-AChE activity, stimulate neurite outgrowth and neurotrophic growth factors, promote remyelination and dopamine release, and improve motor and cognitive function. These properties are important for maintaining synaptic plasticity and promoting neuron recovery. However, the pharmacodynamics of Cordyceps reveal a short half-life and low bioavailability, which limits therapeutic effect; hence, a system to increase bioavailability and enhance efficacy with curative practicality must be developed. Although Cordyceps contain several important compounds, only a few have been evaluated for their neuroprotective potential. Furthermore, no clinical studies have been conducted on the neuroprotective efficacy of the fungus. Thus, there is a need to investigate new bioactive compounds of Cordyceps to develop new drugs for preventing and treating neurodegeneration, in combination with preclinical and clinical safety assessments. Such studies will help in the design and development of safe and effective neuroprotective drugs.
Author Contributions
Conceptualization, N.S. and S.S.A.A.; writing—original draft preparation, N.S.; writing—review and editing, N.S., S.S.A.A. and H.S.; funding acquisition, S.S.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number: RS-2023-00251396 and 2021R1A6A1A03038996).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, D.; Ying, J.; Wang, X.; Zhao, T.; Yoon, S.; Fang, Y.; Zheng, Q.; Liu, X.; Yu, W.; Hua, F. Mitochondrial Dynamics: A Key Role in Neurodegeneration and a Potential Target for Neurodegenerative Disease. Front. Neurosci. 2021, 15, 654785. [Google Scholar] [CrossRef]
- Olufunmilayo, E.O.; Gerke-Duncan, M.B.; Holsinger, R.M.D. Oxidative Stress and Antioxidants in Neurodegenerative Disorders. Antioxidants 2023, 12, 517. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; O, W.; Li, W.; Jiang, Z.-G.; Ghanbari, H.A. Oxidative stress and neurodegenerative disorders. Int. J. Mol. Sci. 2013, 14, 24438–24475. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, A.; Jose Egea-Guerrero, J.; Murillo-Cabezas, F.; Carrillo-Vico, A. Oxidative stress in traumatic brain injury. Curr. Med. Chem. 2014, 21, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Keeler, J.L.; Patsalos, O.; Chung, R.; Schmidt, U.; Breen, G.; Treasure, J.; Himmerich, H.; Dalton, B. Short communication: Serum levels of brain-derived neurotrophic factor and association with pro-inflammatory cytokines in acute and recovered anorexia nervosa. J. Psychiatr. Res. 2022, 150, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.M.; Hong, J.S. Why neurodegenerative diseases are progressive: Uncontrolled inflammation drives disease progression. Trends Immunol. 2008, 29, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.M.; Liu, B.; Zhang, W.; Hong, J.S. Novel anti-inflammatory therapy for Parkinson’s disease. Trends Pharmacol. Sci. 2003, 24, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T.; Mucke, L. Inflammation in neurodegenerative disease--a double-edged sword. Neuron 2002, 35, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Ziv, Y. Immunity to self and self-maintenance: A unified theory of brain pathologies. Trends Immunol. 2008, 29, 211–219. [Google Scholar] [CrossRef]
- Liu, B.; Hong, J.S. Role of microglia in inflammation-mediated neurodegenerative diseases: Mechanisms and strategies for therapeutic intervention. J. Pharmacol. Exp. Ther. 2003, 304, 1–7. [Google Scholar] [CrossRef]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed]
- Das Sarma, J. Microglia-mediated neuroinflammation is an amplifier of virus-induced neuropathology. J. Neurovirol. 2014, 20, 122–136. [Google Scholar] [CrossRef]
- Lian, L.; Zhang, Y.; Liu, L.; Yang, L.; Cai, Y.; Zhang, J.; Xu, S. Neuroinflammation in Ischemic Stroke: Focus on MicroRNA-mediated Polarization of Microglia. Front. Mol. Neurosci. 2020, 13, 612439. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Srivastava, A.K.; Arnold, W.D.; Singh, M.P.; Zhang, Y. Neurodegeneration: Etiologies and new therapies. Biomed. Res. Int. 2015, 2015, 272630. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.; Shen, Y.; Zhang, Y.; Liu, L.; Yang, X. Dietary polyphenols: Regulate the advanced glycation end products-RAGE axis and the microbiota-gut-brain axis to prevent neurodegenerative diseases. Crit. Rev. Food Sci. Nutr. 2023, 63, 9816–9842. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, Y.; Chen, Q.; Liu, Y.; Qiao, Z.; Sang, S.; Zhang, J.; Zhan, S.; Wu, Z.; Liu, L. Research advances of advanced glycation end products in milk and dairy products: Formation, determination, control strategy and immunometabolism via gut microbiota. Food Chem. 2023, 417, 135861. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, Y.; Dong, L.; Shi, R.; Wu, Z.; Liu, L.; Zhang, J.; Wu, Z.; Pan, D. Quantitative determination of Nε-(carboxymethyl) lysine in sterilized milk by isotope dilution UPLC-MS/MS method without derivatization and ion pair reagents. Food Chem. 2022, 385, 132697. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P.; Aryal, P.; Soni, P. RAGE Inhibitors in Neurodegenerative Diseases. Biomedicines 2023, 11, 1131. [Google Scholar] [CrossRef]
- Mushrooms, R. Cordyceps Sinensis vs Militaris: What’s the Best Cordyceps Supplement? 2023. Available online: https://www.realmushrooms.com/cordyceps-sinensis-vs-militaris/ (accessed on 20 August 2023).
- Kobayasi, Y. The genus Cordyceps and its allies. Sci. Rep. Tokyo Bunrika Daigaku Sec. B 1941, 84, 53–260. [Google Scholar]
- Zhang, Y.; Li, E.; Wang, C.; Li, Y.; Liu, X. Ophiocordyceps sinensis, the flagship fungus of China: Terminology, life strategy and ecology. Mycology 2012, 3, 2–10. [Google Scholar]
- Lin, W.-J.; Lee, Y.-I.; Liu, S.-L.; Lin, C.-C.; Chung, T.-Y.; Chou, J.-Y. Evaluating the tradeoffs of a generalist parasitoid fungus, Ophiocordyceps unilateralis, on different sympatric ant hosts. Sci. Rep. 2020, 10, 6428. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Tang, J.; Tola, A.; Auberon, F.; Oluwaniyi, O.; Ouyang, Z. The genus Cordyceps: An extensive review of its traditional uses, phytochemistry and pharmacology. Fitoterapia 2018, 129, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R. Cordyceps: A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Xia, Q.; Liu, L.; Wu, Z.; Pan, D. Recent advances of cereal β-glucan on immunity with gut microbiota regulation functions and its intelligent gelling application. Crit. Rev. Food Sci. Nutr. 2023, 63, 3895–3911. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of a precious ancient Chinese herbal regimen: Cordyceps sinensis Part II. J. Altern. Complement. Med. 1998, 4, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.Y.; Park, S.J.; Park, Y.J. The Anticancer Properties of Cordycepin and Their Underlying Mechanisms. Int. J. Mol. Sci. 2018, 19, 3027. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Lee, S.; Lee, K.; Shin, Y.S.; Kang, H.; Cho, H. Anti-cancer effect of Cordyceps militaris in human colorectal carcinoma RKO cells via cell cycle arrest and mitochondrial apoptosis. Daru 2015, 23, 35. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.; Jang, H.J.; Shen, L.; Yang, K.E.; Jang, M.S.; Huh, Y.H.; Yoo, H.S.; Park, J.; Jang, I.S.; Park, S.J. Cordyceps militaris Exerts Anticancer Effect on Non-Small Cell Lung Cancer by Inhibiting Hedgehog Signaling via Suppression of TCTN3. Integr. Cancer Ther. 2020, 19, 1534735420923756. [Google Scholar] [CrossRef]
- Liu, X.; Dun, M.; Jian, T.; Sun, Y.; Wang, M.; Zhang, G.; Ling, J. Cordyceps militaris extracts and cordycepin ameliorate type 2 diabetes mellitus by modulating the gut microbiota and metabolites. Front. Pharmacol. 2023, 14, 1134429. [Google Scholar] [CrossRef]
- Dong, Y.; Jing, T.; Meng, Q.; Liu, C.; Hu, S.; Ma, Y.; Liu, Y.; Lu, J.; Cheng, Y.; Wang, D.; et al. Studies on the antidiabetic activities of Cordyceps militaris extract in diet-streptozotocin-induced diabetic Sprague-Dawley rats. Biomed. Res. Int. 2014, 2014, 160980. [Google Scholar] [CrossRef]
- Hirsch, K.R.; Smith-Ryan, A.E.; Roelofs, E.J.; Trexler, E.T.; Mock, M.G. Cordyceps militaris Improves Tolerance to High-Intensity Exercise After Acute and Chronic Supplementation. J. Diet. Suppl. 2017, 14, 42–53. [Google Scholar] [CrossRef]
- Nguyen, Q.-V.; Vu, T.-T.; Tran, M.-T.; Ho Thi, P.T.; Thu, H.; Le Thi, T.H.; Chuyen, H.V.; Dinh, M.-H. Antioxidant activity and hepatoprotective effect of exopolysaccharides from cultivated ophiocordyceps sinensis against CCl4-induced liver damages. Nat. Prod. Commun. 2021, 16, 1934578X21997670. [Google Scholar] [CrossRef]
- Chang, Y.; Jeng, K.-C.; Huang, K.-F.; Lee, Y.-C.; Hou, C.-W.; Chen, K.-H.; Cheng, F.-Y.; Liao, J.-W.; Chen, Y.-S. Effect of Cordyceps militaris supplementation on sperm production, sperm motility and hormones in Sprague-Dawley rats. Am. J. Chin. Med. 2008, 36, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.Q.; Yin, F.; Shen, N.; Lin, P.; Xia, B.; Li, Y.J.; Guo, S.D. Polysaccharide CM1 from Cordyceps militaris hinders adipocyte differentiation and alleviates hyperlipidemia in LDLR((+/−)) hamsters. Lipids Health Dis. 2021, 20, 178. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Kai, Q.; Gao, J.; Lian, Z.Q.; Wu, C.M.; Wu, C.A.; Zhu, H.B. Cordycepin prevents hyperlipidemia in hamsters fed a high-fat diet via activation of AMP-activated protein kinase. J. Pharmacol. Sci. 2010, 113, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Solana, M.A.; Corral-Guerrero, I.A.; Téllez-Valencia, A.; Avitia-Domínguez, C.; Meza-Velázquez, J.A.; de Casa, A.G.; Sierra-Campos, E. Cordyceps militaris Inhibited Angiotensin-Converting Enzyme through Molecular Interaction between Cordycepin and ACE C-Domain. Life 2022, 12, 1450. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, H.; Yang, M.; Xu, Y.; Hou, L.; Yu, H.; Wang, X.; Zhang, Z.; Han, J. Bidirectional regulatory effects of Cordyceps on arrhythmia: Clinical evaluations and network pharmacology. Front. Pharmacol. 2022, 13, 948173. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C.; Liang, H.; Liu, L.; Li, S.; Chen, J.; Xie, Y. Transcriptomic analysis of the anti-inflammatory effect of Cordyceps militaris extract on acute gouty arthritis. Front. Pharmacol. 2022, 13, 1035101. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Baggio, C.H.; Borato, D.G.; Santana-Filho, A.P.; Sassaki, G.L.; Iacomini, M.; Van Griensven, L.J. Anti-inflammatory properties of the medicinal mushroom Cordyceps militaris might be related to its linear (1→3)-β-D-glucan. PLoS ONE 2014, 9, e110266. [Google Scholar] [CrossRef]
- Tan, W.; Wang, Y.; Dai, H.; Deng, J.; Wu, Z.; Lin, L.; Yang, J. Potential Therapeutic Strategies for Renal Fibrosis: Cordyceps and Related Products. Front. Pharmacol. 2022, 13, 932172. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, L.; Lu, Y.; Zhang, J.; Yang, M.; Tian, Y.; Dong, J.; Liao, L. Protective effect of Cordyceps sinensis against diabetic kidney disease through promoting proliferation and inhibiting apoptosis of renal proximal tubular cells. BMC Complement. Med. Ther. 2023, 23, 109. [Google Scholar] [CrossRef]
- Sun, T.; Dong, W.; Jiang, G.; Yang, J.; Liu, J.; Zhao, L.; Ma, P. Cordyceps militaris Improves Chronic Kidney Disease by Affecting TLR4/NF-κB Redox Signaling Pathway. Oxid. Med. Cell Longev. 2019, 2019, 7850863. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cheung, F.W.; Chan, M.H.; Hui, P.K.; Ip, S.P.; Ling, Y.H.; Che, C.T.; Liu, W.K. Protective roles of Cordyceps on lung fibrosis in cellular and rat models. J. Ethnopharmacol. 2012, 143, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Huang, K.S.; Shaw, J.F.; Chen, J.R.; Kuo, W.S.; Shen, G.; Grumezescu, A.M.; Holban, A.M.; Wang, Y.T.; Wang, J.S.; et al. Trends in the Immunomodulatory Effects of Cordyceps militaris: Total Extracts, Polysaccharides and Cordycepin. Front. Pharmacol. 2020, 11, 575704. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Shin, H.S.; Leyva-Gómez, G.; Prado-Audelo, M.L.D.; Cortes, H.; Singh, Y.D.; Panda, M.K.; Mishra, A.P.; Nigam, M.; Saklani, S.; et al. Cordyceps spp.: A Review on Its Immune-Stimulatory and Other Biological Potentials. Front. Pharmacol. 2020, 11, 602364. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Gong, Z.; Su, Y.; Lin, J.; Tang, K. Cordyceps fungi: Natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xie, J.; Wang, L.; Li, S. Advanced development in chemical analysis of Cordyceps. J. Pharm. Biomed. Anal. 2014, 87, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gao, Y.; Zhou, Y.; Yu, F.; Li, X.; Zhang, N. Mechanism of Cordyceps sinensis and its Extracts in the Treatment of Diabetic Kidney Disease: A Review. Front. Pharmacol. 2022, 13, 881835. [Google Scholar] [CrossRef] [PubMed]
- Shashidhar, M.G.; Giridhar, P.; Udaya Sankar, K.; Manohar, B. Bioactive principles from Cordyceps sinensis: A potent food supplement—A review. J. Funct. Foods 2013, 5, 1013–1030. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of Cordyceps with special reference to Cordycepin. 3 Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef]
- Shweta; Abdullah, S.; Komal; Kumar, A. A brief review on the medicinal uses of Cordyceps militaris. Pharmacol. Res. Mod. Chin. Med. 2023, 7, 100228. [Google Scholar] [CrossRef]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2012, 65, 474–493. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.-R.; Ahmed, M.; Park, H.-J. Cordyceps militaris as a Bio Functional Food Source: Pharmacological Potential, Anti-Inflammatory Actions and Related Molecular Mechanisms. Microorganisms 2022, 10, 405. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Cordyceps Medicinal Fungus and Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef] [PubMed]
- Elkhateeb, W.A.; Daba, G. Cordyceps more than edible mushroom—A rich source of diverse bioactive metabolites with huge medicinal benefits. J. Biomed. Res. Environ. Sci. 2022, 3, 566–574. [Google Scholar] [CrossRef]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.X.; Wang, S.; Nie, S.; Marcone, M. Properties of Cordyceps sinensis: A review. J. Funct. Foods 2013, 5, 550–569. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wen, C.; Duan, Y.; Zhang, H.; Ma, H. Advance in Cordyceps militaris (Linn) Link polysaccharides: Isolation, structure, and bioactivities: A review. Int. J. Biol. Macromol. 2019, 132, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical composition and nutritional and medicinal value of fruit bodies and submerged cultured mycelia of culinary-medicinal higher Basidiomycetes mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef]
- Deacon, J.W. Fungal Biology; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Chan, J.S.; Barseghyan, G.S.; Asatiani, M.D.; Wasser, S.P. Chemical Composition and Medicinal Value of Fruiting Bodies and Submerged Cultured Mycelia of Caterpillar Medicinal Fungus Cordyceps militaris CBS-132098 (Ascomycetes). Int. J. Med. Mushrooms 2015, 17, 649–659. [Google Scholar] [CrossRef]
- Wu, X.-F.; Zhang, M.; Li, Z. Influence of infrared drying on the drying kinetics, bioactive compounds and flavor of Cordyceps militaris. LWT 2019, 111, 790–798. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489.e4. [Google Scholar] [CrossRef] [PubMed]
- Margolis, J.; Grever, M.R. Pentostatin (Nipent): A review of potential toxicity and its management. Semin Oncol. 2000, 27, 9–14. [Google Scholar] [PubMed]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine 2011, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-P.; Chen, D.-R.; Lin, W.-J.; Lin, Y.-H.; Chen, J.-Y.; Kuo, Y.-H.; Chung, J.-G.; Hsia, T.-C.; Hsieh, W.-T. Ergosta-7, 9 (11), 22-trien-3β-ol attenuates inflammatory responses via inhibiting MAPK/AP-1 induced IL-6/JAK/STAT pathways and activating Nrf2/HO-1 signaling in LPS-stimulated macrophage-like cells. Antioxidants 2021, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.V.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther. 2003, 306, 821–827. [Google Scholar] [CrossRef] [PubMed]
- De Jaco, A.; Augusti-Tocco, G.; Biagioni, S. Muscarinic acetylcholine receptors induce neurite outgrowth and activate the synapsin I gene promoter in neuroblastoma clones. Neuroscience 2002, 113, 331–338. [Google Scholar] [CrossRef]
- Anelli, T.; Mannello, F.; Salani, M.; Tonti, G.A.; Poiana, G.; Biagioni, S. Acetylcholine induces neurite outgrowth and modulates matrix metalloproteinase 2 and 9. Biochem. Biophys. Res. Commun. 2007, 362, 269–274. [Google Scholar] [CrossRef]
- Lee, B.; Park, J.; Park, J.; Shin, H.-J.; Kwon, S.; Yeom, M.; Sur, B.; Kim, S.; Kim, M.; Lee, H. Cordyceps militaris improves neurite outgrowth in Neuro2a cells and reverses memory impairment in rats. Food Sci. Biotechnol. 2011, 20, 1599–1608. [Google Scholar] [CrossRef]
- Phan, C.-W.; David, P.; Naidu, M.; Wong, K.-H.; Sabaratnam, V. Neurite outgrowth stimulatory effects of culinary-medicinal mushrooms and their toxicity assessment using differentiating Neuro-2a and embryonic fibroblast BALB/3T3. BMC Complement. Altern. Med. 2013, 13, 261. [Google Scholar] [CrossRef]
- He, M.T.; Park, C.H.; Cho, E.J. Caterpillar Medicinal: Mushroom, Cordyceps militaris (Ascomycota), Attenuates Aβ 1− 42− Induced Amyloidogenesis and Inflammatory Response by Suppressing Amyloid Precursor Protein Progression and p38 MAPK/JNK Activation. Int. J. Med. Mushrooms 2021, 23, 2634. [Google Scholar] [CrossRef]
- He, M.T.; Lee, A.Y.; Park, C.H.; Cho, E.J. Protective effect of Cordyceps militaris against hydrogen peroxide-induced oxidative stress in vitro. Nutr. Res. Pr. 2019, 13, 279–285. [Google Scholar] [CrossRef]
- Bogoyevitch, M.A.; Kobe, B. Uses for JNK: The many and varied substrates of the c-Jun N-terminal kinases. Microbiol. Mol. Biol. Rev. 2006, 70, 1061–1095. [Google Scholar] [CrossRef] [PubMed]
- He, M.T.; Lee, A.Y.; Kim, J.H.; Park, C.H.; Shin, Y.S.; Cho, E.J. Protective role of Cordyceps militaris in Aβ(1-42)-induced Alzheimer’s disease in vivo. Food Sci. Biotechnol. 2019, 28, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, K.; Kim, S.; Park, Y.L.; Choi, B.-S.; Park, S.-E.; Kim, S.-J. Enhancement of tyrosine hydroxylase expression by Cordyceps militaris. Cent. Eur. J. Biol. 2010, 5, 214–223. [Google Scholar] [CrossRef]
- Lucke-Wold, B.P.; Logsdon, A.F.; Turner, R.C.; Rosen, C.L.; Huber, J.D. Chapter Fourteen—Aging, the Metabolic Syndrome, and Ischemic Stroke: Redefining the Approach for Studying the Blood–Brain Barrier in a Complex Neurological Disease. In Advances in Pharmacology; Davis, T.P., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 71, pp. 411–449. [Google Scholar]
- Pluta, R.; Furmaga-Jabłońska, W.; Januszewski, S.; Czuczwar, S.J. Post-ischemic brain neurodegeneration in the form of Alzheimer’s disease proteinopathy: Possible therapeutic role of curcumin. Nutrients 2022, 14, 248. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Cho, G.-S.; Ryu, S.; Kim, H.J.; Song, H.Y.; Yune, T.Y.; Ju, C.; Kim, W.-K. Post-ischemic treatment of WIB801C, standardized Cordyceps extract, reduces cerebral ischemic injury via inhibition of inflammatory cell migration. J. Ethnopharmacol. 2016, 186, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Choi, H.Y.; Baik, H.H.; Ju, B.G.; Kim, W.K.; Yune, T.Y. Cordycepin-enriched WIB-801C from Cordyceps militaris improves functional recovery by attenuating blood-spinal cord barrier disruption after spinal cord injury. J. Ethnopharmacol. 2017, 203, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.O.; Kim, H.J.; Abu-Taweel, G.M.; Oh, J.; Sung, G.H. Neuroprotective and therapeutic effect of Cordyceps militaris on ischemia-induced neuronal death and cognitive impairments. Saudi J. Biol. Sci. 2019, 26, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Ko, I.-G.; Kim, S.-E.; Hwang, L.; Jin, J.-J.; Choi, H.-H.; Kim, C.-J. Aqueous extract of Cordyceps alleviates cerebral ischemia-induced short-term memory impairment in gerbils. J. Exerc. Rehabil. 2016, 12, 69. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; Zhang, J.; Jia, J.; Ding, J.; Luo, R.; Liu, Z. Cordyceps militaris extract attenuates D-galactose-induced memory impairment in mice. J. Med. Food 2012, 15, 1057–1063. [Google Scholar] [CrossRef]
- Kaokaen, P.; Sorraksa, N.; Phonchai, R.; Chaicharoenaudomrung, N.; Kunhorm, P.; Noisa, P. Enhancing Neurological Competence of Nanoencapsulated Cordyceps/Turmeric Extracts in Human Neuroblastoma SH-SY5Y Cells. Cell Mol. Bioeng. 2023, 16, 81–93. [Google Scholar] [CrossRef]
- Kikkawa, T.; Casingal, C.R.; Chun, S.H.; Shinohara, H.; Hiraoka, K.; Osumi, N. The role of Pax6 in brain development and its impact on pathogenesis of autism spectrum disorder. Brain Res. 2019, 1705, 95–103. [Google Scholar] [CrossRef]
- Krishnasamy, S.; Weng, Y.-C.; Thammisetty, S.S.; Phaneuf, D.; Lalancette-Hebert, M.; Kriz, J. Molecular imaging of nestin in neuroinflammatory conditions reveals marked signal induction in activated microglia. J. Neuroinflamm. 2017, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rahman, M.H.; Mamun-Or-Rashid, A.; Hwang, H.; Chung, S.; Kim, B.; Rhim, H. Autophagy modulation in aggresome formation: Emerging implications and treatments of Alzheimer’s disease. Biomedicines 2022, 10, 1027. [Google Scholar] [CrossRef] [PubMed]
- Kneifel, H.; Konig, W.A.; Loeffler, W.; Müller, R. Ophiocordin, an antifungal antibiotic of Cordyceps ophioglossoides. Arch. Microbiol. 1977, 113, 121–130. [Google Scholar] [CrossRef]
- Braeuer, S.; Borovička, J.; Glabonjat, R.A.; Steiner, L.; Goessler, W. Arsenocholine-O-sulfate: A novel compound as major arsenic species in the parasitic mushroom Tolypocladium ophioglossoides. Chemosphere 2021, 265, 128886. [Google Scholar] [CrossRef] [PubMed]
- Quandt, C.A.; Bushley, K.E.; Spatafora, J.W. The genome of the truffle-parasite Tolypocladium ophioglossoides and the evolution of antifungal peptaibiotics. BMC Genom. 2015, 16, 553. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhang, M.; Guo, Y.-Y.; Mao, X.-M.; Chen, X.-A.; Li, Y.-Q. Revelation of the balanol biosynthetic pathway in Tolypocladium ophioglossoides. Org. Lett. 2018, 20, 6323–6326. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, Z.; Sun, Y.; Ding, Z.; LÜ, L.; Li, Y. Optimization for Production of Intracellular Polysaccharide from Cordyceps ophioglossoides L2 in Submerged Culture and Its Antioxidant Activities in vitro. Chin. J. Chem. Eng. 2012, 20, 294–301. [Google Scholar] [CrossRef]
- Sato, K.; Wakamiya, A.; Maeda, T.; Noguchi, K.; Takashima, A.; Imahori, K. Correlation among secondary structure, amyloid precursor protein accumulation, and neurotoxicity of amyloid β (25–35) peptide as analyzed by single alanine substitution. J. Biochem. 1995, 118, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.-Q.; Park, B.-C.; Lee, J.-S.; Choi, H.-D.; Lee, Y.-S.; Yang, J.-H.; Kim, J.-A. Mycelial extract of Cordyceps ophioglossoides prevents neuronal cell death and ameliorates β-amyloid peptide-induced memory deficits in rats. Biol. Pharm. Bull. 2004, 27, 1126–1129. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, F.; Tsim, K.W. Quality control of Cordyceps sinensis, a valued traditional Chinese medicine. J. Pharm. Biomed. Anal. 2006, 41, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Zhao, S.Y.; Wang, J.H.; Kuang, H.C.; Liu, X. Distribution of nucleosides and nucleobases in edible fungi. J. Agric. Food Chem. 2008, 56, 809–815. [Google Scholar] [CrossRef]
- Guan, J.; Yang, F.Q.; Li, S.P. Evaluation of carbohydrates in natural and cultured Cordyceps by pressurized liquid extraction and gas chromatography coupled with mass spectrometry. Molecules 2010, 15, 4227–4241. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Li, X.L. Determination of ergosterol in Cordyceps sinensis and Cordyceps black-bone chicken capsules by HPLC. Yao Xue Xue Bao 1991, 26, 768–771. [Google Scholar]
- Li, S.P.; Li, P.; Lai, C.M.; Gong, Y.X.; Kan, K.K.; Dong, T.T.; Tsim, K.W.; Wang, Y.T. Simultaneous determination of ergosterol, nucleosides and their bases from natural and cultured Cordyceps by pressurised liquid extraction and high-performance liquid chromatography. J. Chromatogr. A 2004, 1036, 239–243. [Google Scholar] [CrossRef]
- Yuan, J.-P.; Wang, J.-H.; Liu, X.; Kuang, H.-C.; Zhao, S.-Y. Simultaneous determination of free ergosterol and ergosteryl esters in Cordyceps sinensis by HPLC. Food Chem. 2007, 105, 1755–1759. [Google Scholar] [CrossRef]
- Bok, J.W.; Lermer, L.; Chilton, J.; Klingeman, H.G.; Towers, G.H. Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 1999, 51, 891–898. [Google Scholar] [CrossRef]
- Mishra, R.; Upadhyay, Y. Cordicepssinensis: The Chinese Rasayan-current research scenario. Int. J. Res. Pharm. Biomed. Sci. 2011, 2, 1503–1519. [Google Scholar]
- Liu, Y.; Wang, J.; Wang, W.; Zhang, H.; Zhang, X.; Han, C. The Chemical Constituents and Pharmacological Actions of Cordyceps sinensis. Evid.-Based Complement. Altern. Med. 2015, 2015, 575063. [Google Scholar]
- Pal, M.; Bhardwaj, A.; Manickam, M.; Tulsawani, R.; Srivastava, M.; Sugadev, R.; Misra, K. Protective efficacy of the caterpillar mushroom, Ophiocordyceps sinensis (Ascomycetes), from India in neuronal hippocampal cells against hypoxia. Int. J. Med. Mushrooms 2015, 17, 829–840. [Google Scholar] [CrossRef]
- Mehrotra, S.; Kirar, V.; Vats, P.; Nandi, S.P.; Negi, P.; Misra, K. Phytochemical and Antimicrobial Activities of Himalayan Cordyceps Sinensis (Berk.) Sacc; NISCAIR-CSIR: New Delhi, India, 2015. [Google Scholar]
- Liu, Z.; Li, P.; Zhao, D.; Tang, H.; Guo, J. Anti-inflammation Effects of Cordyceps sinensis Mycelium in Focal Cerebral Ischemic Injury Rats. Inflammation 2011, 34, 639–644. [Google Scholar] [CrossRef]
- Kong, R.; Zhang, Y.; Zhang, S.; Liu, M.; Sun, W.; Xing, Y.; Guan, Y.; Han, C.; Liu, Z. Protective Effect of Ethanol Extracts of the Chinese Caterpillar Mushroom, Ophiocordyceps sinensis (Ascomycetes), on the Experimental Middle Cerebral Artery Occlusion/Reperfusion (MCAO/R) Model. Int. J. Med. Mushrooms 2015, 17, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, T.; Wang, T.; Song, J.; Zhou, Z. Effects of apoptosis-related proteins caspase-3, Bax and Bcl-2 on cerebral ischemia rats. Biomed. Rep. 2013, 1, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Tan, T.-Y.; Li, Y.-X.; Li, Y.; Chen, Y.-F.; Ma, R.; Wang, S.-Y.; Li, Q.; Liu, Z.-Q. The protective effect of cordyceps sinensis extract on cerebral ischemic injury via modulating the mitochondrial respiratory chain and inhibiting the mitochondrial apoptotic pathway. Biomed. Pharmacother. 2020, 124, 109834. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Arima, Y.; Kamimura, D.; Sabharwal, L.; Yamada, M.; Bando, H.; Ogura, H.; Atsumi, T.; Murakami, M. Regulation of Immune Cell Infiltration into the CNS by Regional Neural Inputs Explained by the Gate Theory. Mediat. Inflamm. 2013, 2013, 898165. [Google Scholar] [CrossRef] [PubMed]
- Goverman, J. Autoimmune T cell responses in the central nervous system. Nat. Rev. Immunol. 2009, 9, 393–407. [Google Scholar] [CrossRef]
- Prajeeth, C.K.; Kronisch, J.; Khorooshi, R.; Knier, B.; Toft-Hansen, H.; Gudi, V.; Floess, S.; Huehn, J.; Owens, T.; Korn, T.; et al. Effectors of Th1 and Th17 cells act on astrocytes and augment their neuroinflammatory properties. J. Neuroinflamm. 2017, 14, 204. [Google Scholar] [CrossRef]
- Zhong, S.S.; Xiang, Y.J.; Liu, P.J.; He, Y.; Yang, T.T.; Wang, Y.Y.; Rong, A.; Zhang, J.; Liu, G.Z. Effect of Cordyceps sinensis on the Treatment of Experimental Autoimmune Encephalomyelitis: A Pilot Study on Mice Model. Chin. Med. J. 2017, 130, 2296–2301. [Google Scholar] [PubMed]
- Chen, Y.; Fu, L.; Han, M.; Jin, M.; Wu, J.; Tan, L.; Chen, Z.; Zhang, X. The Prophylactic and Therapeutic Effects of Fermented Cordyceps sinensis Powder, Cs-C-Q80, on Subcortical Ischemic Vascular Dementia in Mice. Evid.-Based Complement. Altern. Med. 2018, 2018, 4362715. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, K.; Adachi, K.; Kataoka, S.; Watanabe, A.; Tabira, T.; Takahashi, K.; Wakita, H. Chronic cerebral hypoperfusion induced by right unilateral common carotid artery occlusion causes delayed white matter lesions and cognitive impairment in adult mice. Exp. Neurol. 2008, 210, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yan, W.; Wu, X.; Hong, D.; Lu, X.; Rao, Y. Protective effects of Corbrin Capsule against permanent cerebral ischemia in mice. Biomed. Pharmacother. 2020, 121, 109646. [Google Scholar] [CrossRef] [PubMed]
- Li, I.C.; Lin, S.; Tsai, Y.T.; Hsu, J.H.; Chen, Y.L.; Lin, W.H.; Chen, C.C. Cordyceps cicadae mycelia and its active compound HEA exert beneficial effects on blood glucose in type 2 diabetic db/db mice. J. Sci. Food Agric. 2019, 99, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, T.; Li, C.; Xie, L.; Li, N.; Hou, T.; Wang, Y.; Wang, B. Potential therapeutic effects of Cordyceps cicadae and Paecilomyces cicadae on adenine-induced chronic renal failure in rats and their phytochemical analysis. Drug Des. Devel Ther. 2019, 13, 103–117. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, Q.; Weng, Q. Secondary metabolites (SMs) of Isaria cicadae and Isaria tenuipes. RSC Adv. 2019, 9, 172–184. [Google Scholar] [CrossRef]
- Lu, Y.; Luo, F.; Cen, K.; Xiao, G.; Yin, Y.; Li, C.; Li, Z.; Zhan, S.; Zhang, H.; Wang, C. Omics data reveal the unusual asexual-fruiting nature and secondary metabolic potentials of the medicinal fungus Cordyceps cicadae. BMC Genom. 2017, 18, 668. [Google Scholar] [CrossRef]
- Chyau, C.-C.; Wu, H.-L.; Peng, C.-C.; Huang, S.-H.; Chen, C.-C.; Chen, C.-H.; Peng, R.Y. Potential protection effect of ER homeostasis of N6-(2-hydroxyethyl) adenosine isolated from Cordyceps cicadae in nonsteroidal anti-inflammatory drug-stimulated human proximal tubular cells. Int. J. Mol. Sci. 2021, 22, 1577. [Google Scholar] [CrossRef]
- Latini, S.; Pedata, F. Adenosine in the central nervous system: Release mechanisms and extracellular concentrations. J. Neurochem. 2001, 79, 463–484. [Google Scholar] [CrossRef]
- Furuya, T.; Hirotani, M.; Matsuzawa, M. N6-(2-hydroxyethyl) adenosine, a biologically active compound from cultured mycelia of Cordyceps and Isaria species. Phytochemistry 1983, 22, 2509–2512. [Google Scholar] [CrossRef]
- Wang, X.; Qin, A.; Xiao, F.; Olatunji, O.J.; Zhang, S.; Pan, D.; Han, W.; Wang, D.; Ni, Y. N(6) -(2-hydroxyethyl)-adenosine from Cordyceps cicadae protects against diabetic kidney disease via alleviation of oxidative stress and inflammation. J. Food Biochem. 2019, 43, e12727. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Kang, J.; Wen, T.; Lei, B.; Hyde, K.D. Cordycepin and N6-(2-hydroxyethyl)-adenosine from Cordyceps pruinosa and their interaction with human serum albumin. PLoS ONE 2015, 10, e0121669. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zheng, R.; Deng, Y.; Chen, Y.; Zhang, S. Ergosterol peroxide from Cordyceps cicadae ameliorates TGF-beta1-induced activation of kidney fibroblasts. Phytomedicine 2014, 21, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyama, M.; Shinmoto, H. Ergosterol peroxide from an edible mushroom suppresses inflammatory responses in RAW264.7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharmacol. 2007, 150, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.P.; McDowell, C.M.; Liu, Y.; Wagner, A.H.; Thole, D.; Faga, B.P.; Wordinger, R.J.; Braun, T.A.; Clark, A.F. Optic nerve crush induces spatial and temporal gene expression patterns in retina and optic nerve of BALB/cJ mice. Mol. Neurodegener. 2014, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.T.; Jhou, B.Y.; Hsu, J.H.; Fu, H.I.; Chen, Y.L.; Shih, Y.C.; Chen, C.C.; Tsai, R.K. Neuroprotective Effects of Cordyceps cicadae (Ascomycetes) Mycelium Extract in the Rat Model of Optic Nerve Crush. Int. J. Med. Mushrooms 2022, 24, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, J.; Wang, D.; Yu, X.; Olatunji, O.J.; Ouyang, Z.; Wei, Y. Neuroprotective effects of butanol fraction of Cordyceps cicadae on glutamate-induced damage in pc12 cells involving oxidative toxicity. Chem. Biodivers. 2018, 15, e1700385. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z.; Wang, D.; Yu, X. Neuroprotective effects of adenosine isolated from Cordyceps cicadae against oxidative and ER stress damages induced by glutamate in PC12 cells. Environ. Toxicol. Pharmacol. 2016, 44, 53–61. [Google Scholar] [CrossRef]
- Lu, M.Y.; Chen, C.C.; Lee, L.Y.; Lin, T.W.; Kuo, C.F. N(6)-(2-Hydroxyethyl)adenosine in the Medicinal Mushroom Cordyceps cicadae Attenuates Lipopolysaccharide-Stimulated Pro-inflammatory Responses by Suppressing TLR4-Mediated NF-κB Signaling Pathways. J. Nat. Prod. 2015, 78, 2452–2460. [Google Scholar] [CrossRef]
- Santos, N.A.G.d.; Ferreira, R.S.; Santos, A.C.d. Overview of cisplatin-induced neurotoxicity and ototoxicity, and the protective agents. Food Chem. Toxicol. 2020, 136, 111079. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, S.; Olatunji, O.J.; Wu, Y. Nucleosides rich extract from Cordyceps cicadae alleviated cisplatin-induced neurotoxicity in rats: A behavioral, biochemical and histopathological study. Arab. J. Chem. 2022, 15, 103476. [Google Scholar] [CrossRef]
- Chang, C.Y.; Yang, P.X.; Yu, T.L.; Lee, C.L. Cordyceps cicadae NTTU 868 Mycelia Fermented with Deep Ocean Water Minerals Prevents D-Galactose-Induced Memory Deficits by Inhibiting Oxidative Inflammatory Factors and Aging-Related Risk Factors. Nutrients 2023, 15, 1968. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis Part I. J. Altern. Complement. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; He, W.; Zhou, X.; Lv, Q.; Xu, X.; Yang, S.; Zhao, C.; Guo, L. Cordycepin protects against cerebral ischemia/reperfusion injury in vivo and in vitro. Eur. J. Pharmacol. 2011, 664, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Ge, Y.; Sun, L.; Xu, X.; Xie, P.; Zhan, M.; Wang, M.; Dong, Z.; Li, J.; Duan, S. Cordycepin inhibits albumin-induced epithelial-mesenchymal transition of renal tubular epithelial cells by reducing reactive oxygen species production. Free Radic. Res. 2012, 46, 174–183. [Google Scholar] [CrossRef]
- He, Y.T.; Zhang, X.L.; Xie, Y.M.; Xu, Y.X.; Li, J.R. Extraction and antioxidant property in vitro of cordycepin in artificially cultivated Cordyceps militaris. Adv. Mater. Res. 2013, 750, 1593–1596. [Google Scholar] [CrossRef]
- Ramesh, T.; Yoo, S.-K.; Kim, S.-W.; Hwang, S.-Y.; Sohn, S.-H.; Kim, I.-W.; Kim, S.-K. Cordycepin (3′-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol. 2012, 47, 979–987. [Google Scholar] [CrossRef]
- Zhou, Y.; Guo, Z.; Meng, Q.; Lu, J.; Wang, N.; Liu, H.; Liang, Q.; Quan, Y.; Wang, D.; Xie, J. Cordycepin affects multiple apoptotic pathways to mediate hepatocellular carcinoma cell death. Anti-Cancer Agents Med. Chem. 2017, 17, 143–149. [Google Scholar] [CrossRef]
- Kouli, A.; Torsney, K.M.; Kuan, W.-L. Parkinson’s Disease: Etiology, Neuropathology, and Pathogenesis; Exon Publications: Brisbane City, Australia, 2018; pp. 3–26. [Google Scholar]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z. Cordycepin protects PC12 cells against 6-hydroxydopamine induced neurotoxicity via its antioxidant properties. Biomed. Pharmacother. 2016, 81, 7–14. [Google Scholar] [CrossRef]
- Jiang, X.; Tang, P.-C.; Chen, Q.; Zhang, X.; Fan, Y.-Y.; Yu, B.-C.; Gu, X.-X.; Sun, Y.; Ge, X.-Q.; Zhang, X.-L. Cordycepin exerts neuroprotective effects via an anti-apoptotic mechanism based on the mitochondrial pathway in a rotenone-induced parkinsonism rat model. CNS Neurol. Disord. Drug Targets 2019, 18, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Huang, W.M.; Tang, P.C.; Sun, Y.; Zhang, X.; Qiu, L.; Yu, B.C.; Zhang, X.Y.; Hong, Y.X.; He, Y.; et al. Anti-inflammatory and neuroprotective effects of natural cordycepin in rotenone-induced PD models through inhibiting Drp1-mediated mitochondrial fission. Neurotoxicology 2021, 84, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Zhu, X. Cordycepin mitigates MPTP-induced Parkinson’s disease through inhibiting TLR/NF-κB signaling pathway. Life Sci 2019, 223, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, P.; Ge, H.; Qu, X.; Jin, X. Effects of cordycepin on the microglia-overactivation-induced impairments of growth and development of hippocampal cultured neurons. PLoS ONE 2015, 10, e0125902. [Google Scholar] [CrossRef]
- Berg, A. Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of IκB: A mechanism for NK-κB acitivation. Mol. Cell Biol. 1993, 13, 3301. [Google Scholar]
- Sun, Y.; Huang, W.-m.; Tang, P.-c.; Zhang, X.; Zhang, X.-y.; Yu, B.-c.; Fan, Y.-Y.; Ge, X.-q.; Zhang, X.-L. Neuroprotective effects of natural cordycepin on LPS-induced Parkinson’s disease through suppressing TLR4/NF-κB/NLRP3-mediated pyroptosis. J. Funct. Foods 2020, 75, 104274. [Google Scholar] [CrossRef]
- Zou, R.-X.; Gu, X.; Ding, J.-J.; Wang, T.; Bi, N.; Niu, K.; Ge, M.; Chen, X.-T.; Wang, H.-L. Pb exposure induces an imbalance of excitatory and inhibitory synaptic transmission in cultured rat hippocampal neurons. Toxicol. Vitr. 2020, 63, 104742. [Google Scholar] [CrossRef] [PubMed]
- Iovino, L.; Tremblay, M.E.; Civiero, L. Glutamate-induced excitotoxicity in Parkinson’s disease: The role of glial cells. J. Pharmacol. Sci. 2020, 144, 151–164. [Google Scholar] [CrossRef]
- Ahmed, I.; Bose, S.K.; Pavese, N.; Ramlackhansingh, A.; Turkheimer, F.; Hotton, G.; Hammers, A.; Brooks, D.J. Glutamate NMDA receptor dysregulation in Parkinson’s disease with dyskinesias. Brain 2011, 134, 979–986. [Google Scholar] [CrossRef]
- Yao, L.H.; Huang, J.N.; Li, C.H.; Li, H.H.; Yan, W.W.; Cai, Z.L.; Liu, W.X.; Xiao, P. Cordycepin suppresses excitatory synaptic transmission in rat hippocampal slices via a presynaptic mechanism. CNS Neurosci. Ther. 2013, 19, 216–221. [Google Scholar] [CrossRef]
- Wang, J.; Gong, Y.; Tan, H.; Li, W.; Yan, B.; Cheng, C.; Wan, J.; Sun, W.; Yuan, C.; Yao, L.-H. Cordycepin suppresses glutamatergic and GABAergic synaptic transmission through activation of A1 adenosine receptor in rat hippocampal CA1 pyramidal neurons. Biomed. Pharmacother. 2022, 145, 112446. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Oh, J.-I.; Lee, S.J.; Park, G. The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells. Neurotoxicology 2014, 41, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Huang, L.-P.; Li, Y.; Liu, C.; Wang, S.; Meng, W.; Wei, S.; Liu, X.-P.; Gong, Y.; Yao, L.-H. Neuroprotective effects of cordycepin inhibit Aβ-induced apoptosis in hippocampal neurons. Neurotoxicology 2018, 68, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Arbel-Ornath, M.; Hudry, E.; Boivin, J.R.; Hashimoto, T.; Takeda, S.; Kuchibhotla, K.V.; Hou, S.; Lattarulo, C.R.; Belcher, A.M.; Shakerdge, N. Soluble oligomeric amyloid-β induces calcium dyshomeostasis that precedes synapse loss in the living mouse brain. Mol. Neurodegener. 2017, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Sayer, R.; Law, E.; Connelly, P.J.; Breen, K.C. Association of a salivary acetylcholinesterase with Alzheimer’s disease and response to cholinesterase inhibitors. Clin. Biochem. 2004, 37, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Pósfai, B.; Cserép, C.; Orsolits, B.; Dénes, Á. New insights into microglia–neuron interactions: A neuron’s perspective. Neuroscience 2019, 405, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Sarlus, H.; Heneka, M.T. Microglia in Alzheimer’s disease. J. Clin. Investig. 2017, 127, 3240–3249. [Google Scholar] [CrossRef]
- Jiao, L.; Yu, Z.; Zhong, X.; Yao, W.; Xing, L.; Ma, G.; Shen, J.; Wu, Y.; Du, K.; Liu, J.; et al. Cordycepin improved neuronal synaptic plasticity through CREB-induced NGF upregulation driven by MG-M2 polarization: A microglia-neuron symphony in AD. Biomed. Pharmacother. 2023, 157, 114054. [Google Scholar] [CrossRef]
- L, L.; X, W.; Z, Y. Ischemia-reperfusion Injury in the Brain: Mechanisms and Potential Therapeutic Strategies. Biochem. Pharmacol. 2016, 5, 213. [Google Scholar] [CrossRef]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting oxidative stress and inflammation to prevent ischemia-reperfusion injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef]
- Chen, L.S.; Stellrecht, C.M.; Gandhi, V. RNA-directed agent, cordycepin, induces cell death in multiple myeloma cells. Br. J. Haematol. 2008, 140, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.K.; Lim, S.S.; Yoo, K.-Y.; Lee, Y.S.; Kim, H.G.; Kang, I.-J.; Kwon, H.J.; Park, J.; Choi, S.Y.; Won, M.-H. A phytochemically characterized extract of Cordyceps militaris and cordycepin protect hippocampal neurons from ischemic injury in gerbils. Planta Medica 2008, 74, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, X.-P.; Jiang, W.; Zeng, B.; Meng, W.; Huang, L.-P.; Li, Y.-P.; Sun, W.; Yuan, C.-H.; Yao, L.-H. Anti-effects of cordycepin to hypoxia-induced membrane depolarization on hippocampal CA1 pyramidal neuron. Eur. J. Pharmacol. 2017, 796, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.-H.; Wang, J.; Liu, C.; Wei, S.; Li, G.; Wang, S.; Meng, W.; Liu, Z.-B.; Huang, L.-P. Cordycepin protects against β-amyloid and ibotenic acid-induced hippocampal CA1 pyramidal neuronal hyperactivity. Korean J. Physiol. Pharmacol. 2019, 23, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.-L.; Wang, C.-Y.; Jiang, Z.-J.; Li, H.-H.; Liu, W.-X.; Gong, L.-W.; Xiao, P.; Li, C.-H. Effects of cordycepin on Y-maze learning task in mice. Eur. J. Pharmacol. 2013, 714, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.S.; Cao, Z.P.; Shang, Y.J.; Liu, Q.Y.; Wu, B.Y.; Liu, W.X.; Li, C.H. Neuroprotection of cordycepin in NMDA-induced excitotoxicity by modulating adenosine A(1) receptors. Eur. J. Pharmacol. 2019, 853, 325–335. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, H.; Bao, H.; Zhang, D.; Feng, L.; Xiao, Y.; Zhu, K.; Hou, Y.; Luo, S.; Zhang, Y. Cordycepin (3′-deoxyadenosine) promotes remyelination via suppression of neuroinflammation in a cuprizone-induced mouse model of demyelination. Int. Immunopharmacol. 2019, 75, 105777. [Google Scholar] [CrossRef]
- Song, Y.-C.; Liu, C.-T.; Lee, H.-J.; Yen, H.-R. Cordycepin prevents and ameliorates experimental autoimmune encephalomyelitis by inhibiting leukocyte infiltration and reducing neuroinflammation. Biochem. Pharmacol. 2022, 197, 114918. [Google Scholar] [CrossRef]
- Paschon, V.; Takada, S.H.; Ikebara, J.M.; Sousa, E.; Raeisossadati, R.; Ulrich, H.; Kihara, A.H. Interplay between exosomes, microRNAs and toll-like receptors in brain disorders. Mol. Neurobiol. 2016, 53, 2016–2028. [Google Scholar] [CrossRef]
- Lee, J.; Hamanaka, G.; Lo, E.H.; Arai, K. Heterogeneity of microglia and their differential roles in white matter pathology. CNS Neurosci. Ther. 2019, 25, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Shigemori, Y.; Katayama, Y.; Mori, T.; Maeda, T.; Kawamata, T. Matrix metalloproteinase-9 is associated with blood-brain barrier opening and brain edema formation after cortical contusion in rats. Acta Neurochir. Suppl. 2006, 96, 130–133. [Google Scholar] [PubMed]
- Wei, P.; Wang, K.; Luo, C.; Huang, Y.; Misilimu, D.; Wen, H.; Jin, P.; Li, C.; Gong, Y.; Gao, Y. Cordycepin confers long-term neuroprotection via inhibiting neutrophil infiltration and neuroinflammation after traumatic brain injury. J. Neuroinflamm. 2021, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Wang, A.; He, Y.; Si, Z.; Xu, S.; Zhang, S.; Wang, K.; Wang, D.; Liu, Y. Cordycepin attenuates traumatic brain injury-induced impairments of blood-brain barrier integrity in rats. Brain Res. Bull. 2016, 127, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.M.; Brugniaux, J.V.; Filipponi, T.; Marley, C.J.; Stacey, B.; Soria, R.; Rimoldi, S.F.; Cerny, D.; Rexhaj, E.; Pratali, L. Exaggerated systemic oxidative-inflammatory-nitrosative stress in chronic mountain sickness is associated with cognitive decline and depression. J. Physiol. 2019, 597, 611–629. [Google Scholar] [CrossRef]
- Goodman, M.D.; Makley, A.T.; Huber, N.L.; Clarke, C.N.; Friend, L.A.W.; Schuster, R.M.; Bailey, S.R.; Barnes, S.L.; Dorlac, W.C.; Johannigman, J.A. Hypobaric hypoxia exacerbates the neuroinflammatory response to traumatic brain injury. J. Surg. Res. 2011, 165, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Pan, L.; Cui, L.; Li, T.; Zhao, S.; Hu, Y.; Tao, X.; Deng, H.; Jiang, J.; Zhao, B.; et al. Cordycepin ameliorates acute hypobaric hypoxia induced blood-brain barrier disruption, and cognitive impairment partly by suppressing the TLR4/NF-κB/MMP-9 pathway in the adult rats. Eur. J. Pharmacol. 2022, 924, 174952. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Li, X.; Yang, W.; Yu, L.; Jiang, X.; Chen, Y.; Shen, Z.; Li, C.; Gu, M.; Shi, L. N6-(2-hydroxyethyl)-Adenosine Induces Apoptosis via ER Stress and Autophagy of Gastric Carcinoma Cells In Vitro and In Vivo. Int. J. Mol. Sci. 2020, 21, 5815. [Google Scholar] [CrossRef]
- Lu, T.-H.; Chang, J.-W.; Jhou, B.-Y.; Hsu, J.-H.; Li, T.-J.; Lee, L.-Y.; Chen, Y.-L.; Chang, H.-H.; Chen, C.-C.; Wu, P.-S.; et al. Preventative Effects of Cordyceps cicadae Mycelial Extracts on the Early-Stage Development of Cataracts in UVB-Induced Mice Cataract Model. Nutrients 2023, 15, 3103. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, T.; Olatunji, O.J.; Tang, J.; Wei, Y.; Ouyang, Z. N(6)-(2-hydroxyethyl)-adenosine from Cordyceps cicadae attenuates hydrogen peroxide induced oxidative toxicity in PC12 cells. Metab. Brain Dis. 2019, 34, 1325–1334. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, H.; Qiu, X.; Xu, D.; Chen, Y.; Barnes, G.N.; Tu, Y.; Gyabaah, A.T.; Gharbal, A.H.A.A.; Peng, C. Neuroprotective effects of adenosine A1 receptor signaling on cognitive impairment induced by chronic intermittent hypoxia in mice. Front. Cell Neurosci. 2020, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.K.; Li, J.; Cheung, A.W.; Zhu, Y.; Zheng, K.Y.; Bi, C.W.; Duan, R.; Choi, R.C.; Lau, D.T.; Dong, T.T. Cordysinocan, a polysaccharide isolated from cultured Cordyceps, activates immune responses in cultured T-lymphocytes and macrophages: Signaling cascade and induction of cytokines. J. Ethnopharmacol. 2009, 124, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yan, H.; Ma, X.; Jia, J.; Zhang, G.; Guo, X.; Gui, Z. Comparison of the structural characterization and biological activity of acidic polysaccharides from Cordyceps militaris cultured with different media. World J. Microbiol. Biotechnol. 2012, 28, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Song, L.; Zhao, Y.; Bin, W.; Wang, L.; Zhang, H.; Wu, Y.; Ye, W.; Yao, X. Isolation and biological properties of polysaccharide CPS-1 from cultured Cordyceps militaris. Fitoterapia 2004, 75, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cui, S.W.; Cheung, P.C.; Wang, Q. Antitumor polysaccharides from mushrooms: A review on their isolation process, structural characteristics and antitumor activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Liu, J.-y.; Feng, C.-p.; Li, X.; Chang, M.-c.; Meng, J.-l.; Xu, L.-j. Immunomodulatory and antioxidative activity of Cordyceps militaris polysaccharides in mice. Int. J. Biol. Macromol. 2016, 86, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-T.; Li, H.-C.; Li, C.-B.; Dou, D.-Q.; Gao, M.-B. Protective effects on mitochondria and anti-aging activity of polysaccharides from cultivated fruiting bodies of Cordyceps militaris. Am. J. Chin. Med. 2010, 38, 1093–1106. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Wei, Y.; Ouyang, Z.; Su, Z. Polysaccharides purified from Cordyceps cicadae protects PC12 cells against glutamate-induced oxidative damage. Carbohydr. Polym. 2016, 153, 187–195. [Google Scholar] [CrossRef]
- Shen, W.; Song, D.; Wu, J.; Zhang, W. Protective effect of a polysaccharide isolated from a cultivated Cordyceps mycelia on hydrogen peroxide-induced oxidative damage in PC12 cells. Phytother. Res. 2011, 25, 675–680. [Google Scholar] [CrossRef]
- Li, S.P.; Zhao, K.J.; Ji, Z.N.; Song, Z.H.; Dong, T.T.X.; Lo, C.K.; Cheung, J.K.H.; Zhu, S.Q.; Tsim, K.W.K. A polysaccharide isolated from Cordyceps sinensis, a traditional Chinese medicine, protects PC12 cells against hydrogen peroxide-induced injury. Life Sci. 2003, 73, 2503–2513. [Google Scholar] [CrossRef]
- Yang, C.H.; Su, C.H.; Liu, S.C.; Ng, L.T. Isolation, Anti-Inflammatory Activity and Physico-chemical Properties of Bioactive Polysaccharides from Fruiting Bodies of Cultivated Cordyceps cicadae (Ascomycetes). Int. J. Med. Mushrooms 2019, 21, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, X.; Ge, Q.; Li, J.; Wang, D.; Wei, Y.; Ouyang, Z. Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, H.; Nakamura, E.; Okuyama, E.; Ishibashi, M. Six immunosuppressive features from an ascomycete, Zopfiella longicaudata, found in a screening study monitored by immunomodulatory activity. Chem. Pharm. Bull. 2004, 52, 1005–1008. [Google Scholar] [CrossRef]
- Yoneyama, T.; Takahashi, H.; Grudniewska, A.; Ban, S.; Umeyama, A.; Noji, M. Ergostane-Type Sterols from Several Cordyceps Strains. Nat. Prod. Commun. 2022, 17, 1934578X221105363. [Google Scholar] [CrossRef]
- Zhabinskii, V.N.; Drasar, P.; Khripach, V.A. Structure and biological activity of ergostane-type steroids from fungi. Molecules 2022, 27, 2103. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.; Wang, H. Pharmacological actions of Cordyceps, a prized folk medicine. J. Pharm. Pharmacol. 2005, 57, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.-H.; Lin, C.-H.; Shih, C.-C. Ergostatrien-3β-ol from Antrodia camphorata inhibits diabetes and hyperlipidemia in high-fat-diet treated mice via regulation of hepatic related genes, glucose transporter 4, and AMP-activated protein kinase phosphorylation. J. Agric. Food Chem. 2015, 63, 2479–2489. [Google Scholar] [CrossRef]
- Huang, G.-J.; Huang, S.-S.; Lin, S.-S.; Shao, Y.-Y.; Chen, C.-C.; Hou, W.-C.; Kuo, Y.-H. Analgesic effects and the mechanisms of anti-inflammation of ergostatrien-3β-ol from Antrodia camphorata submerged whole broth in mice. J. Agric. Food Chem. 2010, 58, 7445–7452. [Google Scholar] [CrossRef]
- Liu, H.-P.; Kuo, Y.-H.; Cheng, J.; Chang, L.-Z.; Chang, M.-S.; Su, L.-W.; Chuang, T.-N.; Lin, W.-Y. Ergosta-7, 9 (11), 22-trien-3β-ol Rescues AD Deficits by Modulating Microglia Activation but Not Oxidative Stress. Molecules 2021, 26, 5338. [Google Scholar] [CrossRef]
- Konishi, H.; Kiyama, H. Microglial TREM2/DAP12 Signaling: A Double-Edged Sword in Neural Diseases. Front. Cell Neurosci. 2018, 12, 206. [Google Scholar] [CrossRef]
- Hsieh, W.-T.; Hsu, M.-H.; Lin, W.-J.; Xiao, Y.-C.; Lyu, P.-C.; Liu, Y.-C.; Lin, W.-Y.; Kuo, Y.-H.; Chung, J.-G. Ergosta-7, 9 (11), 22-trien-3β-ol Interferes with LPS Docking to LBP, CD14, and TLR4/MD-2 Co-Receptors to Attenuate the NF-κB Inflammatory Pathway In Vitro and Drosophila. Int. J. Mol. Sci. 2021, 22, 6511. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.M. Recombinant tissue plasminogen activator for the treatment of acute ischemic stroke. In Baylor University Medical Center Proceedings; Taylor & Francis: Abingdon, UK, 2011; Volume 24, pp. 257–259. [Google Scholar]
- Wang, Y.-H.; Chern, C.-M.; Liou, K.-T.; Kuo, Y.-H.; Shen, Y.-C. Ergostatrien-7, 9 (11), 22-trien-3β-ol from Antrodia camphorata ameliorates ischemic stroke brain injury via downregulation of p65NF-κ-B and caspase 3, and activation of Akt/GSK3/catenin-associated neurogenesis. Food Funct. 2019, 10, 4725–4738. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Zorov, D.B.; Kim, S.-H.; Pepe, S.; Fu, Q.; Fishbein, K.W.; Ziman, B.D.; Wang, S.; Ytrehus, K.; Antos, C.L. Glycogen synthase kinase-3β mediates convergence of protection signaling to inhibit the mitochondrial permeability transition pore. J. Clin. Investig. 2004, 113, 1535–1549. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.; Jeong, S.-W.; Jung, K.-H.; Han, S.-Y.; Lee, S.-T.; Kim, M.; Roh, J.-K. Celecoxib induces functional recovery after intracerebral hemorrhage with reduction of brain edema and perihematomal cell death. J. Cereb. Blood Flow Metab. 2004, 24, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.; Poddar, R.; Thompson, J.; Rosenberg, G.; Yang, Y. Intranuclear matrix metalloproteinases promote DNA damage and apoptosis induced by oxygen–glucose deprivation in neurons. Neuroscience 2012, 220, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, P.-J.; Wang, M.-H.; Hsiao, C.-J.; Chen, C.-K.; Lin, F.-L.; Huang, S.-H.; Yen, J.-L.; Tsai, P.-H.; Kuo, Y.-H.; Hsiao, G. Ergosta-7,9(11),22-trien-3β-ol Alleviates Intracerebral Hemorrhage-Induced Brain Injury and BV-2 Microglial Activation. Molecules 2021, 26, 2970. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.M.; Cao, W.; Yao, K.W.; Liu, Z.Q.; Guo, J.Y. Anti-inflammation and antioxidant effect of Cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis, in middle cerebral artery occlusion-induced focal cerebral ischemia in rats. Metab. Brain Dis. 2012, 27, 159–165. [Google Scholar] [CrossRef]
- Qi, W.; Zhang, Y.; Yan, Y.-b.; Lei, W.; Wu, Z.-x.; Liu, N.; Liu, S.; Shi, L.; Fan, Y. The protective effect of cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis, on diabetic osteopenia in alloxan-induced diabetic rats. Evid.-Based Complement. Altern. Med. 2013, 2013, 985636. [Google Scholar] [CrossRef]
- Qian, G.M.; Pan, G.F.; Guo, J.Y. Anti-inflammatory and antinociceptive effects of cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis. Nat. Prod. Res. 2012, 26, 2358–2362. [Google Scholar] [CrossRef]
- Kuroda, S.; Siesjö, B.K. Reperfusion damage following focal ischemia: Pathophysiology and therapeutic windows. Clin. Neurosci. 1997, 4, 199–212. [Google Scholar]
- Xu, G.; Du, P.; An, L.; Lv, G.; Miao, L.; Guangxin, Y. Research progress of active polypeptides of Cordyceps militaris. In Proceedings of the 40th World Congress of Vine and Wine BIO Web of Conferences, Sofia, Bulgaria, 29 May–2 June 2017; p. 01002. [Google Scholar]
- Xie, L.; An, L.; Du, P. Study on the enzymolysis technology of Cordyceps militaris peptides and its effect on immune function in mice. Chin. Tradit. Pat. Med. 2016, 38, 2048–2050. [Google Scholar]
- Yuan, G.; An, L.; Sun, Y.; Xu, G.; Du, P. Improvement of Learning and Memory Induced by Cordyceps Polypeptide Treatment and the Underlying Mechanism. Evid.-Based Complement. Altern. Med. 2018, 2018, 9419264. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Inoue, K.; Yamamoto, S.; Ikumoto, T.; Sasaki, S.; Toyama, R.; Chiba, K.; Hoshino, Y.; Okumoto, T. Fungal metabolites. Part 11. A potent immunosuppressive activity found in Isaria sinclairii metabolite. J. Antibiot. 1994, 47, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Kohara, T.; Nakao, N.; Arita, M.; Chiba, K.; Mishina, T.; Sasaki, S.; Fujita, T. Design, synthesis, and structure-activity relationships of 2-substituted-2-amino-1, 3-propanediols: Discovery of a novel immunosuppressant, FTY720. Bioorg. Med. Chem. Lett. 1995, 5, 853–856. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Suzuki, H.; Sozen, T.; Rolland, W.; Zhang, J.H. Activation of sphingosine 1-phosphate receptor-1 by FTY720 is neuroprotective after ischemic stroke in rats. Stroke 2010, 41, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, Y.; Sugahara, K.; Kataoka, H.; Kawaguchi, T.; Masubuchi, Y.; Chiba, K. FTY720, a novel immunosuppressant, induces sequestration of circulating mature lymphocytes by acceleration of lymphocyte homing in rats. II. FTY720 prolongs skin allograft survival by decreasing T cell infiltration into grafts but not cytokine production in vivo. J. Immunol. 1998, 160, 5493–5499. [Google Scholar] [PubMed]
- Chiba, K. Discovery of fingolimod based on the chemical modification of a natural product from the fungus, Isaria sinclairii. J. Antibiot. 2020, 73, 666–678. [Google Scholar] [CrossRef]
- Jęśko, H.; Wencel, P.L.; Wójtowicz, S.; Strosznajder, J.; Lukiw, W.J.; Strosznajder, R.P. Fingolimod affects transcription of genes encoding enzymes of ceramide metabolism in animal model of Alzheimer’s disease. Mol. Neurobiol. 2020, 57, 2799–2811. [Google Scholar] [CrossRef]
- Zhao, P.; Yang, X.; Yang, L.; Li, M.; Wood, K.; Liu, Q.; Zhu, X. Neuroprotective effects of fingolimod in mouse models of Parkinson’s disease. FASEB J. 2017, 31, 172–179. [Google Scholar] [CrossRef]
- Kappos, L.; Antel, J.; Comi, G.; Montalban, X.; O’Connor, P.; Polman, C.H.; Haas, T.; Korn, A.A.; Karlsson, G.; Radue, E.W. Oral fingolimod (FTY720) for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 355, 1124–1140. [Google Scholar] [CrossRef]
- FDA. FDA Approves First Oral Drug to Reduce MS Relapses; Press Release; FDA: Silver Spring, MD, USA, 2010. [Google Scholar]
- Zhu, X.Y.; Ma, T.T.; Li, Y.; Zhang, M.Q.; Zhao, L.; Liang, J.; Min, L.Q. Fingolimod protects against neurovascular unit injury in a rat model of focal cerebral ischemia/reperfusion injury. Neural Regen. Res. 2023, 18, 869–874. [Google Scholar] [PubMed]
- Lin, B.-Q.; Li, S.-P. Cordyceps as an herbal drug. In Herbal Medicine: Biomolecular and Clinical Aspects; CRC Press: Boca Raton, FL, USA, 2011; Volume 5. [Google Scholar]
- Kwon, H.-W.; Shin, J.-H.; Lim, D.H.; Ok, W.J.; Nam, G.S.; Kim, M.J.; Kwon, H.-K.; Noh, J.-H.; Lee, J.-Y.; Kim, H.-H.; et al. Antiplatelet and antithrombotic effects of cordycepin-enriched WIB-801CE from Cordyceps militaris ex vivo, in vivo, and in vitro. BMC Complement. Altern. Med. 2016, 16, 508. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.C.; Wong, J.W.; Suen, Y.-K.; Kwok, T.; Fung, K.; Kong, S. Cordycepin induced eryptosis in mouse erythrocytes through a Ca 2+-dependent pathway without caspase-3 activation. Arch. Toxicol. 2007, 81, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Han, X.; Jin, H.; Ma, S. Arsenic species in Cordyceps sinensis and its potential health risks. Front. Pharmacol. 2019, 10, 1471. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Sun, Y.; Luo, F.; Wang, C. Bioactive Metabolites and Potential Mycotoxins Produced by Cordyceps Fungi: A Review of Safety. Toxins 2020, 12, 410. [Google Scholar] [CrossRef] [PubMed]
- Rodman, L.E.; Farnell, D.R.; Coyne, J.M.; Allan, P.W.; Hill, D.L.; Duncan, K.L.; Tomaszewski, J.E.; Smith, A.C.; Page, J.G. Toxicity of cordycepin in combination with the adenosine deaminase inhibitor 2’-deoxycoformycin in beagle dogs. Toxicol. Appl. Pharmacol. 1997, 147, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Pegram, R.A.; Wyatt, R.D. Avian gout caused by oosporein, a mycotoxin produced by Caetomium trilaterale. Poult Sci. 1981, 60, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Mallebrera, B.; Juan-Garcia, A.; Font, G.; Ruiz, M.J. Mechanisms of beauvericin toxicity and antioxidant cellular defense. Toxicol. Lett. 2016, 246, 28–34. [Google Scholar] [CrossRef]
- Jung, S.J.; Jung, E.S.; Choi, E.K.; Sin, H.S.; Ha, K.C.; Chae, S.W. Immunomodulatory effects of a mycelium extract of Cordyceps (Paecilomyces hepiali; CBG-CS-2): A randomized and double-blind clinical trial. BMC Complement. Altern. Med. 2019, 19, 77. [Google Scholar] [CrossRef]
- Chae, S.-W. Efficacy and Safety of Cordyceps Sinensis Mycelium Culture Extract(Paecilomyces Hepiali, CBG-CS-2) on Promotion of Immunity. 2019. Available online: https://clinicaltrials.gov/show/NCT02814617 (accessed on 25 August 2023).
- Jung, S.J.; Hwang, J.H.; Oh, M.R.; Chae, S.W. Effects of Cordyceps militaris supplementation on the immune response and upper respiratory infection in healthy adults: A randomized, double-blind, placebo-controlled study. J. Nutr. Health 2019, 52, 258–267. [Google Scholar] [CrossRef]
- Kang, H.J.; Baik, H.W.; Kim, S.J.; Lee, S.G.; Ahn, H.Y.; Park, J.S.; Park, S.J.; Jang, E.J.; Park, S.W.; Choi, J.Y. Cordyceps militaris enhances cell-mediated immunity in healthy Korean men. J. Med. Food 2015, 18, 1164–1172. [Google Scholar] [CrossRef]
- University of Oxford. Anti-Cancer Drug Derived from Fungus Shows Promise in Clinical Trials; University of Oxford: Oxford, UK, 2021. [Google Scholar]
- Xiao, Y.; Huang, X.-Z.; Zhu, J.-S. Randomized double-blind placebo-controlled clinical trial and assessment of fermentation product of Cordyceps sinensis (Cs-4) in enhancing aerobic capacity and respiratory function of the healthy elderly volunteers. Chin. J. Integr. Med. 2004, 10, 187–192. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, X.; Xiao, L.; Zhou, J.; Feng, L.; Wang, G. Efficacy and Safety of Cordyceps militaris as an Adjuvant to Duloxetine in the Treatment of Insomnia in Patients with Depression: A 6-Week Double-Blind, Randomized, Placebo-Controlled Trial. Front. Psychiatry 2021, 12, 754921. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.-A.; Lin, T.-H.; Wang, J.-S.; Chen, J.-J.; Hsu, W.-K.; Ying, L.-C.; Liang, Z.-C. The effects of Cordyceps militaris fruiting bodies in micturition and prostate size in benign prostatic hyperplasia patients: A pilot study. Pharmacol. Res. Mod. Chin. Med. 2022, 4, 100143. [Google Scholar] [CrossRef]
- Heo, J.Y.; Baik, H.W.; Kim, H.J.; Lee, J.M.; Kim, H.W.; Choi, Y.S.; Won, J.H.; Hyun Mi Kim, W.I.P.; Kim, C.Y. The efficacy and safety of Cordyceps militaris in Korean adults who have mild liver dysfunction. J. Korean Soc. Parenter. Enter. Nutr. 2015, 7, 81–86. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Krochmal, R.; Abrazado, M.; Kim, W.; Cooper, C.B. Effect of Cs-4®(Cordyceps sinensis) on exercise performance in healthy older subjects: A double-blind, placebo-controlled trial. J. Altern. Complement. Med. 2010, 16, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, K.R.; Mock, M.G.; Roelofs, E.J.; Trexler, E.T.; Smith-Ryan, A.E. Chronic supplementation of a mushroom blend on oxygen kinetics, peak power, and time to exhaustion. J. Int. Soc. Sports Nutr. 2015, 12, 1. [Google Scholar] [CrossRef]
Figure 1.
Some of the bioactive compounds from Cordyceps.

Figure 2.
Summary of neuroprotective mechanisms exerted by various bioactive compounds from Cordyceps. Cordycepin, HEA, and EK100 suppressed the TLR-4/NF-kB pathway, suppressing inflammatory cytokines (TNF-α, IL-1β, IL-6). Additionally, cordycepin suppressed NLRP3 inflammasome activation and iNOS levels. Cordymin also inhibited pro-inflammatory cytokines in addition to elevating the levels of antioxidant enzymes. CP and NP inhibited NO levels. Mitochondrial membrane potential was improved by adenosine and HEA. Cordycepin and adenosine improved Bcl2/Bax ratio. CPA1, CPA2, APS, and cordycepin reduced Ca2+ overload with cordycepin further decreasing caspase-12 and caspase-3 activity. Fingolimod also reduced caspase-3 indicating an anti-apoptotic mechanism. CPA1, CPA2, and APS decreased ROS. EK100 activated the PI3K/AKT pathway. Cordycepin, HEA, adenosine, APS, CPA1, CPA2, and CP70 increased the antioxidant levels. (red arrows represent downregulation/decrease, while green arrows indicate upregulation/increase by the compounds).
Figure 2.
Summary of neuroprotective mechanisms exerted by various bioactive compounds from Cordyceps. Cordycepin, HEA, and EK100 suppressed the TLR-4/NF-kB pathway, suppressing inflammatory cytokines (TNF-α, IL-1β, IL-6). Additionally, cordycepin suppressed NLRP3 inflammasome activation and iNOS levels. Cordymin also inhibited pro-inflammatory cytokines in addition to elevating the levels of antioxidant enzymes. CP and NP inhibited NO levels. Mitochondrial membrane potential was improved by adenosine and HEA. Cordycepin and adenosine improved Bcl2/Bax ratio. CPA1, CPA2, APS, and cordycepin reduced Ca2+ overload with cordycepin further decreasing caspase-12 and caspase-3 activity. Fingolimod also reduced caspase-3 indicating an anti-apoptotic mechanism. CPA1, CPA2, and APS decreased ROS. EK100 activated the PI3K/AKT pathway. Cordycepin, HEA, adenosine, APS, CPA1, CPA2, and CP70 increased the antioxidant levels. (red arrows represent downregulation/decrease, while green arrows indicate upregulation/increase by the compounds).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharma, H.; Sharma, N.; An, S.S.A. Unique Bioactives from Zombie Fungus (Cordyceps) as Promising Multitargeted Neuroprotective Agents. Nutrients 2024, 16, 102. https://doi.org/10.3390/nu16010102
AMA Style
Sharma H, Sharma N, An SSA. Unique Bioactives from Zombie Fungus (Cordyceps) as Promising Multitargeted Neuroprotective Agents. Nutrients. 2024; 16(1):102. https://doi.org/10.3390/nu16010102
Chicago/Turabian StyleSharma, Himadri, Niti Sharma, and Seong Soo A. An. 2024. "Unique Bioactives from Zombie Fungus (Cordyceps) as Promising Multitargeted Neuroprotective Agents" Nutrients 16, no. 1: 102. https://doi.org/10.3390/nu16010102
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.