
Astaxanthin Ameliorates Worsened Muscle Dysfunction of MDX Mice Fed with a High-Fat Diet through Reducing Lipotoxicity and Regulating Gut Microbiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experiment Design
2.2. Grip Strength Measurement
2.3. Treadmill Endurance Test
2.4. Determination of Serum Lipids and Glucose
2.5. Determination of Muscle TG and MDA
2.6. Hematoxylin and Eosin Staining
2.7. Immunofluorescence Staining
2.8. Transmission Electron Microscopy Analysis
2.9. Muscle Lipidomic Analysis
2.10. Gut Microbiota Analysis
2.11. Statistical Analysis
3. Results
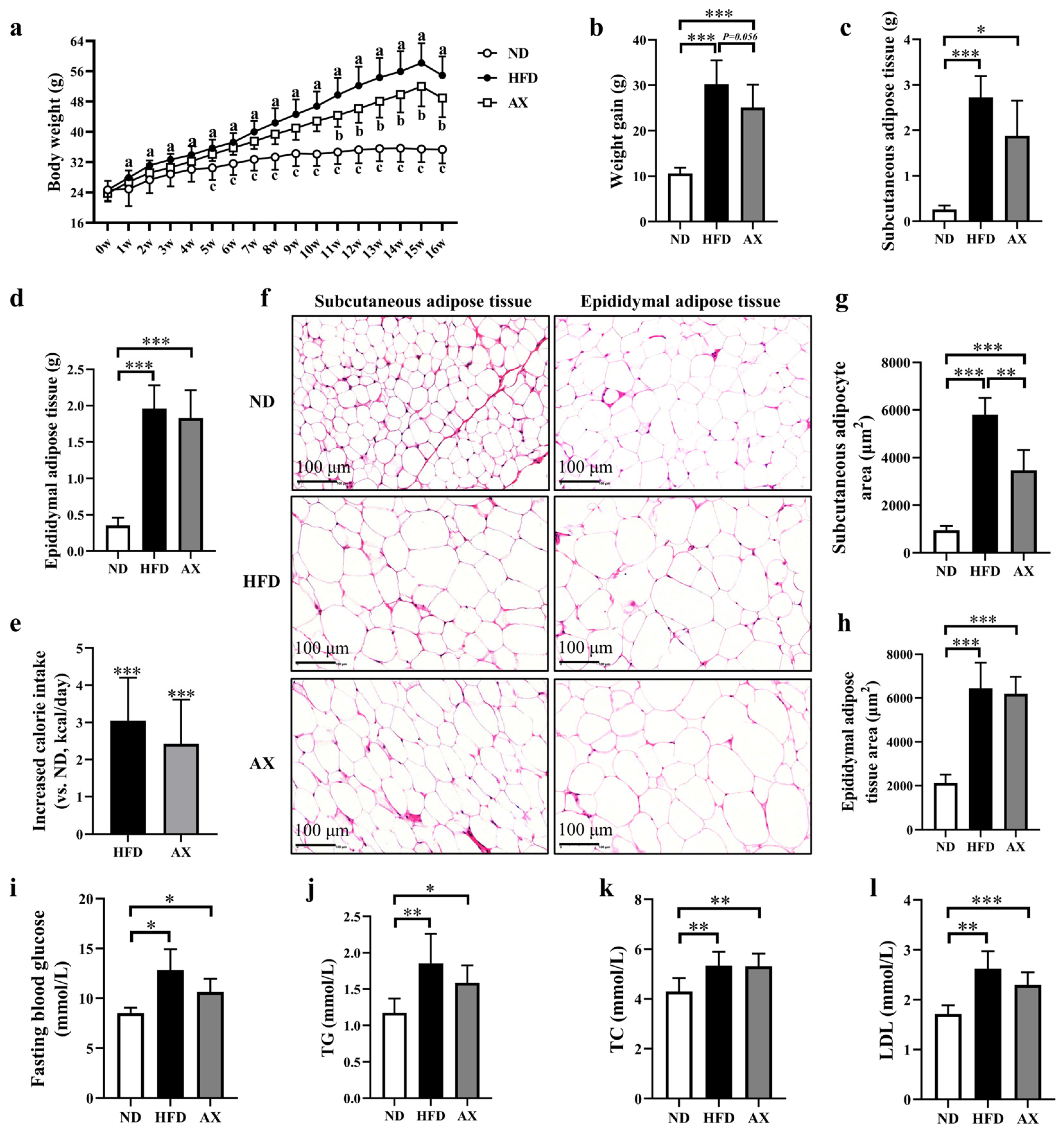
3.1. The Effects of AX on Body Weight, Body Fat, and Glucolipid Metabolism of MDX Mice Fed with HFD
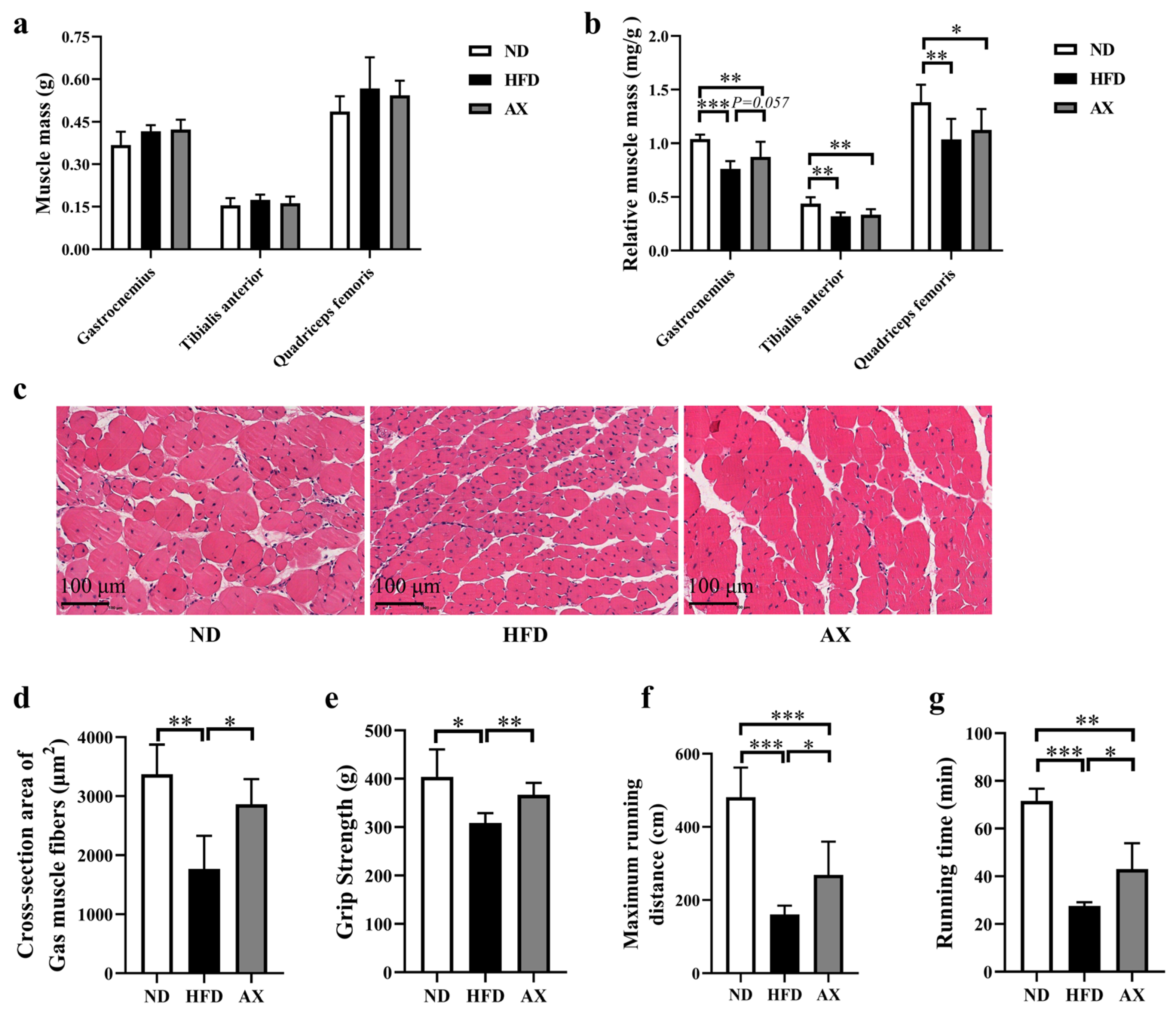
3.2. The Effects of AX on Muscle Mass, Morphology and Function of MDX Mice Fed with HFD
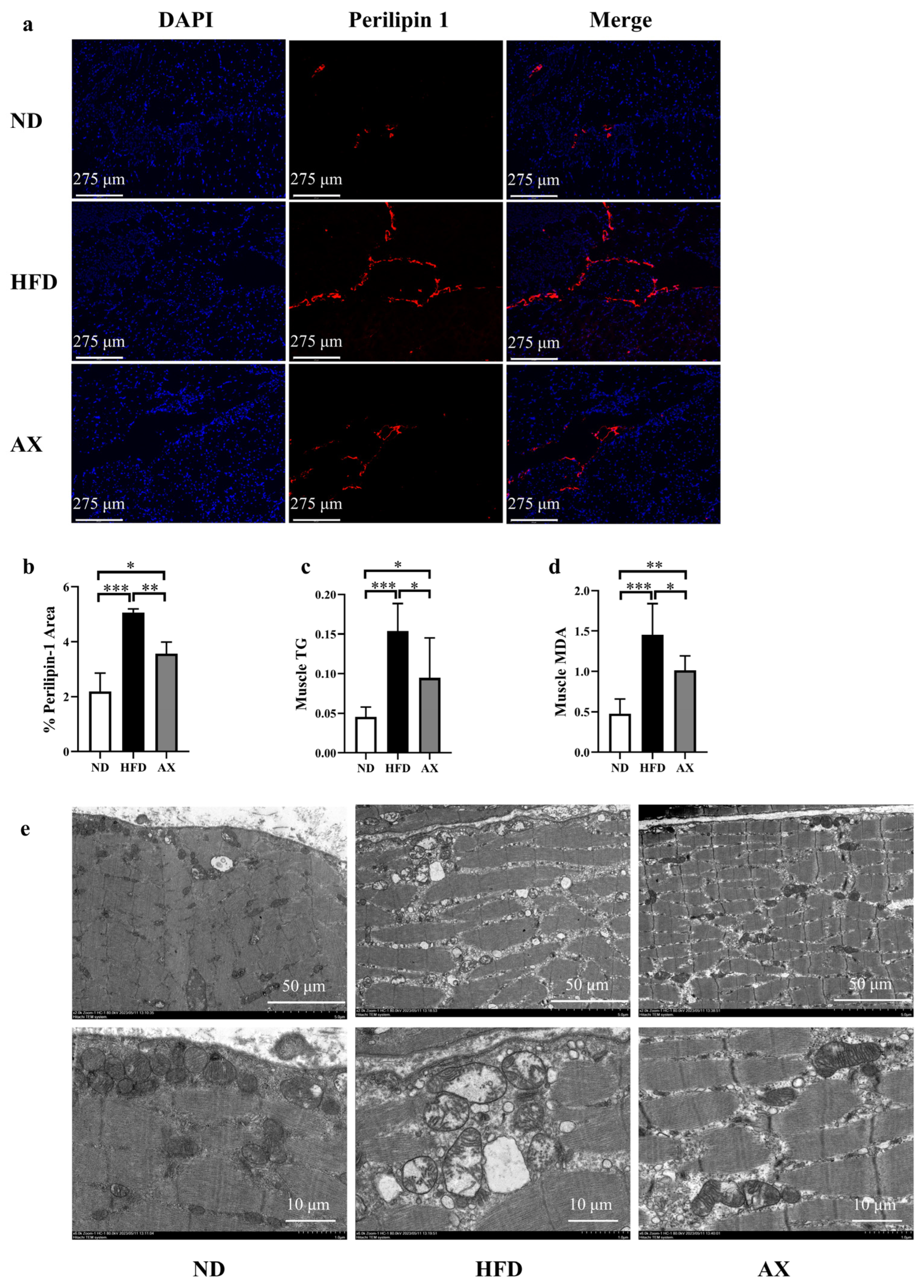
3.3. The Effects of AX on Muscle Lipid Deposition and Mitochondrial Damage of MDX Mice Fed with HFD
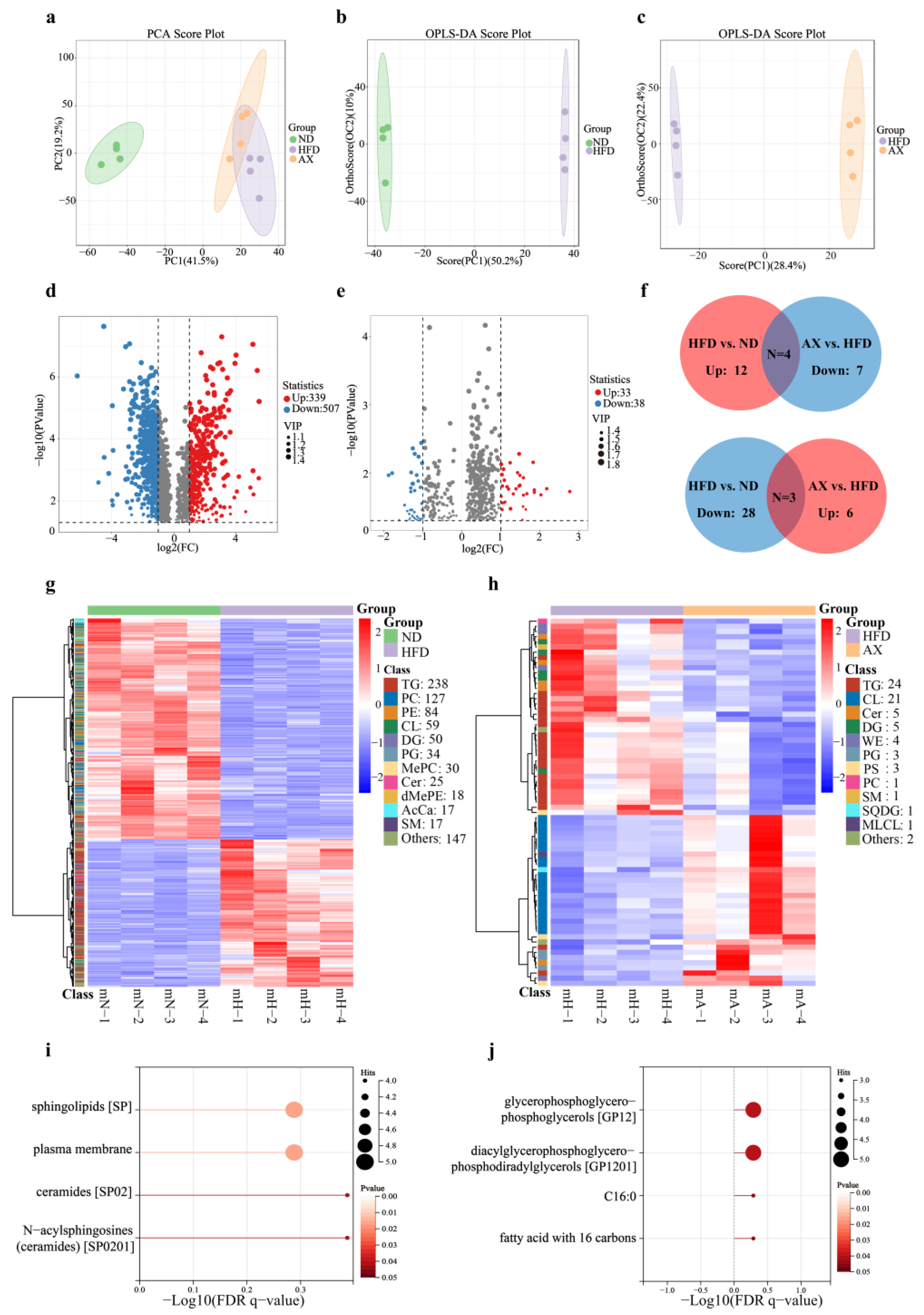
3.4. AX Supplementation Improved Muscle Lipid Metabolism by Lipidomic Analysis
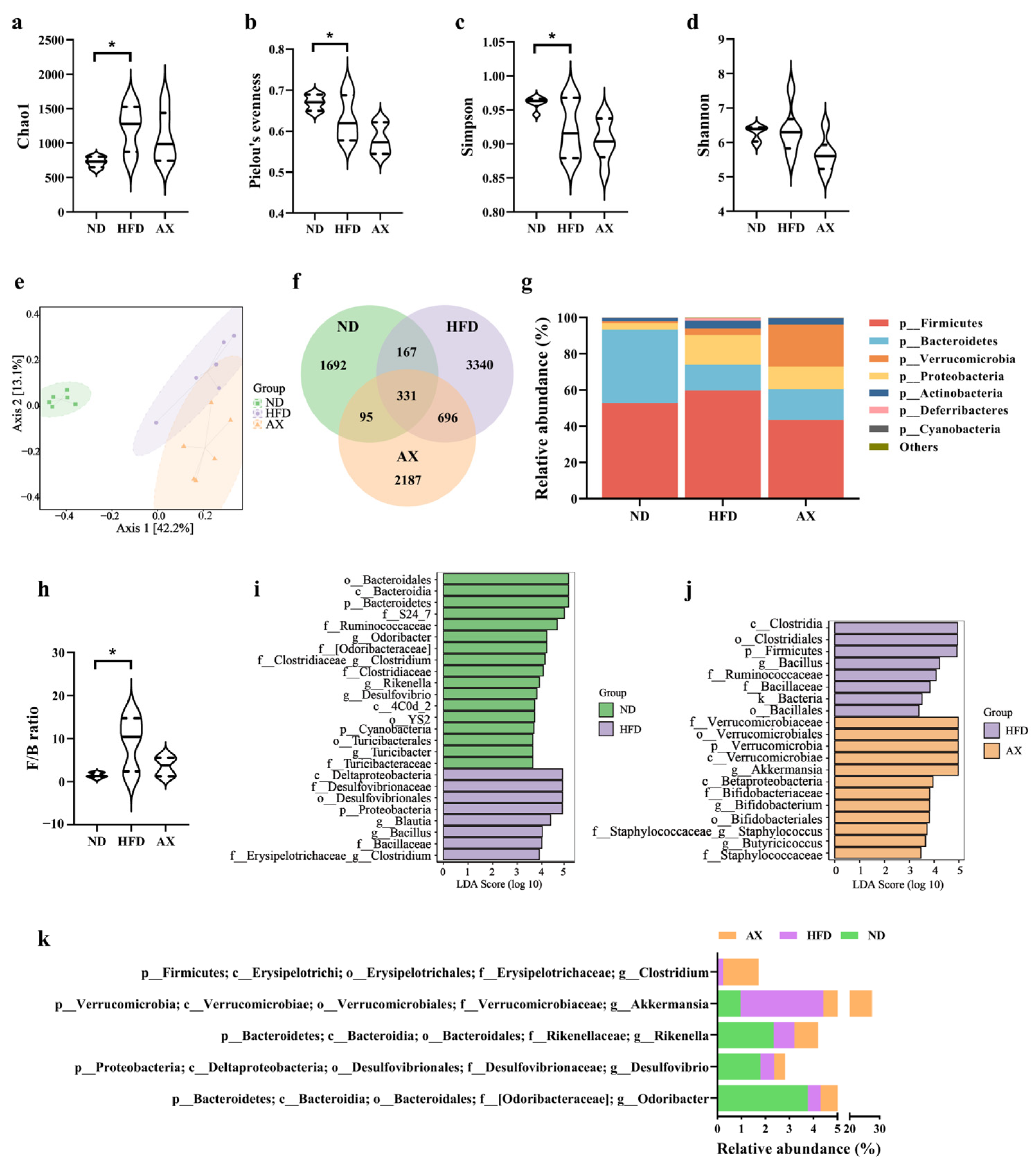
3.5. Gut Microbiota Changes in Diversity and Structure Caused by HFD and AX Intervention
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aartsma-Rus, A.; Ginjaar, I.B.; Bushby, K. The importance of genetic diagnosis for Duchenne muscular dystrophy. J. Med. Genet. 2016, 53, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Whitehead, N.P.; Froehner, S.C. Absence of Dystrophin Disrupts Skeletal Muscle Signaling: Roles of Ca2+, Reactive Oxygen Species, and Nitric Oxide in the Development of Muscular Dystrophy. Physiol. Rev. 2016, 96, 253–305. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Goemans, N.; Takeda, S.; Mercuri, E.; Aartsma-Rus, A. Duchenne muscular dystrophy. Nat. Rev. Dis. Primers 2021, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Bushby, K.; Finkel, R.; Birnkrant, D.J.; Case, L.E.; Clemens, P.R.; Cripe, L.; Kaul, A.; Kinnett, K.; McDonald, C.; Pandya, S.; et al. Diagnosis and management of Duchenne muscular dystrophy, part 1: Diagnosis, and pharmacological and psychosocial management. Lancet Neurol. 2010, 9, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qin, D.; Wu, L.; Li, M.; Song, L.; Wei, C.; Lu, C.; Zhang, X.; Hong, S.; Ma, M.; et al. Genotype characterization and delayed loss of ambulation by glucocorticoids in a large cohort of patients with Duchenne muscular dystrophy. Orphanet J. Rare Dis. 2021, 16, 188. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.M.; Henricson, E.K.; Abresch, R.T.; Duong, T.; Joyce, N.C.; Hu, F.; Clemens, P.R.; Hoffman, E.P.; Cnaan, A.; Gordish-Dressman, H. Long-term effects of glucocorticoids on function, quality of life, and survival in patients with Duchenne muscular dystrophy: A prospective cohort study. Lancet 2018, 391, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Barber, B.J.; Andrews, J.G.; Lu, Z.; West, N.A.; Meaney, F.J.; Price, E.T.; Gray, A.; Sheehan, D.W.; Pandya, S.; Yang, M.; et al. Oral corticosteroids and onset of cardiomyopathy in Duchenne muscular dystrophy. J. Pediatr. 2013, 163, 1080–1084.e1081. [Google Scholar] [CrossRef]
- Weber, D.R.; Hadjiyannakis, S.; McMillan, H.J.; Noritz, G.; Ward, L.M. Obesity and Endocrine Management of the Patient With Duchenne Muscular Dystrophy. Pediatrics 2018, 142, S43–S52. [Google Scholar] [CrossRef]
- Birnkrant, D.J.; Bushby, K.; Bann, C.M.; Alman, B.A.; Apkon, S.D.; Blackwell, A.; Case, L.E.; Cripe, L.; Hadjiyannakis, S.; Olson, A.K.; et al. Diagnosis and management of Duchenne muscular dystrophy, part 2: Respiratory, cardiac, bone health, and orthopaedic management. Lancet Neurol. 2018, 17, 347–361. [Google Scholar] [CrossRef]
- Ward, L.M.; Hadjiyannakis, S.; McMillan, H.J.; Noritz, G.; Weber, D.R. Bone Health and Osteoporosis Management of the Patient With Duchenne Muscular Dystrophy. Pediatrics 2018, 142, S34–S42. [Google Scholar] [CrossRef]
- Wood, C.L.; Straub, V.; Guglieri, M.; Bushby, K.; Cheetham, T. Short stature and pubertal delay in Duchenne muscular dystrophy. Arch. Dis. Child. 2016, 101, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.K.; Yadav, R.; Mukherjee, S.; Pal, L.; Sinha, N. Abnormal lipid metabolism in skeletal muscle tissue of patients with muscular dystrophy: In vitro, high-resolution NMR spectroscopy based observation in early phase of the disease. Magn. Reson. Imaging 2017, 38, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Cardone, N.; Taglietti, V.; Baratto, S.; Kefi, K.; Periou, B.; Gitiaux, C.; Barnerias, C.; Lafuste, P.; Pharm, F.L.; Pharm, J.N.; et al. Myopathologic trajectory in Duchenne muscular dystrophy (DMD) reveals lack of regeneration due to senescence in satellite cells. Acta Neuropathol. Commun. 2023, 11, 167. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.M.; Lin, A.J.; Strumwasser, A.R.; Cory, K.; Whitney, K.; Ho, T.; Ho, T.; Lee, J.L.; Rucker, D.H.; Nguyen, C.Q.; et al. Mitochondrial Dysfunction Is an Early Consequence of Partial or Complete Dystrophin Loss in mdx Mice. Front. Physiol. 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.V.; Winkler, K.; Wiedemann, F.R.; von Bossanyi, P.; Dietzmann, K.; Kunz, W.S. Impaired mitochondrial oxidative phosphorylation in skeletal muscle of the dystrophin-deficient mdx mouse. Mol. Cell. Biochem. 1998, 183, 87–96. [Google Scholar] [CrossRef]
- Kennedy, T.L.; Moir, L.; Hemming, S.; Edwards, B.; Squire, S.; Davies, K.; Guiraud, S. Utrophin influences mitochondrial pathology and oxidative stress in dystrophic muscle. Skelet. Muscle 2017, 7, 22. [Google Scholar] [CrossRef]
- Turner, N.; Kowalski, G.M.; Leslie, S.J.; Risis, S.; Yang, C.; Lee-Young, R.S.; Babb, J.R.; Meikle, P.J.; Lancaster, G.I.; Henstridge, D.C.; et al. Distinct patterns of tissue-specific lipid accumulation during the induction of insulin resistance in mice by high-fat feeding. Diabetologia 2013, 56, 1638–1648. [Google Scholar] [CrossRef]
- Meex, R.C.R.; Blaak, E.E.; van Loon, L.J.C. Lipotoxicity plays a key role in the development of both insulin resistance and muscle atrophy in patients with type 2 diabetes. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2019, 20, 1205–1217. [Google Scholar] [CrossRef]
- Tsushima, K.; Bugger, H.; Wende, A.R.; Soto, J.; Jenson, G.A.; Tor, A.R.; McGlauflin, R.; Kenny, H.C.; Zhang, Y.; Souvenir, R.; et al. Mitochondrial Reactive Oxygen Species in Lipotoxic Hearts Induce Post-Translational Modifications of AKAP121, DRP1, and OPA1 That Promote Mitochondrial Fission. Circ. Res. 2018, 122, 58–73. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Matthews, E.; Brassington, R.; Kuntzer, T.; Jichi, F.; Manzur, A.Y. Corticosteroids for the treatment of Duchenne muscular dystrophy. Cochrane Database Syst. Rev. 2016, 2016, CD003725. [Google Scholar] [CrossRef] [PubMed]
- Saure, C.; Caminiti, C.; Weglinski, J.; de Castro Perez, F.; Monges, S. Energy expenditure, body composition, and prevalence of metabolic disorders in patients with Duchenne muscular dystrophy. Diabetes Metab. Syndr. 2018, 12, 81–85. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Cai, X.T.; Peng, W.T.; Song, J.; Zhou, H.M.; Li, X.R.; Wu, X.N. Cross-Sectional Study of Nutritional Status and Dietary Nutrient Intake in Children with Duchenne Muscular Dystrophy (DMD) in China. Sichuan Da Xue Xue Bao Yi Xue Ban J. Sichuan Univ. Med. Sci. Ed. 2022, 53, 1068–1073. [Google Scholar] [CrossRef]
- Davidson, Z.E.; Ryan, M.M.; Kornberg, A.J.; Sinclair, K.; Cairns, A.; Walker, K.Z.; Truby, H. Observations of body mass index in Duchenne muscular dystrophy: A longitudinal study. Eur. J. Clin. Nutr. 2014, 68, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Billich, N.; Adams, J.; Carroll, K.; Truby, H.; Evans, M.; Ryan, M.M.; Davidson, Z.E. The Relationship between Obesity and Clinical Outcomes in Young People with Duchenne Muscular Dystrophy. Nutrients 2022, 14, 3304. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.C.; Tsui, P.H.; Wang, C.Y.; Chien, Y.H.; Weng, H.L.; Yang, C.Y.; Weng, W.C. Hepatic Steatosis Assessment as a New Strategy for the Metabolic and Nutritional Management of Duchenne Muscular Dystrophy. Nutrients 2022, 14, 727. [Google Scholar] [CrossRef] [PubMed]
- Canapari, C.A.; Barrowman, N.; Hoey, L.; Walker, S.W.; Townsend, E.; Tseng, B.S.; Katz, S.L. Truncal fat distribution correlates with decreased vital capacity in Duchenne muscular dystrophy. Pediatr. Pulmonol. 2015, 50, 63–70. [Google Scholar] [CrossRef]
- Goemans, N.; Wong, B.; Van den Hauwe, M.; Signorovitch, J.; Sajeev, G.; Cox, D.; Landry, J.; Jenkins, M.; Dieye, I.; Yao, Z.; et al. Prognostic factors for changes in the timed 4-stair climb in patients with Duchenne muscular dystrophy, and implications for measuring drug efficacy: A multi-institutional collaboration. PLoS ONE 2020, 15, e0232870. [Google Scholar] [CrossRef]
- White, Z.; Hakim, C.H.; Theret, M.; Yang, N.N.; Rossi, F.; Cox, D.; Francis, G.A.; Straub, V.; Selby, K.; Panagiotopoulos, C.; et al. High prevalence of plasma lipid abnormalities in human and canine Duchenne and Becker muscular dystrophies depicts a new type of primary genetic dyslipidemia. J. Clin. Lipidol. 2020, 14, 459–469.e450. [Google Scholar] [CrossRef]
- Tamilarasan, K.P.; Temmel, H.; Das, S.K.; Al Zoughbi, W.; Schauer, S.; Vesely, P.W.; Hoefler, G. Skeletal muscle damage and impaired regeneration due to LPL-mediated lipotoxicity. Cell Death Dis. 2012, 3, e354. [Google Scholar] [CrossRef]
- Radice, R.P.; Limongi, A.R.; Viviano, E.; Padula, M.C.; Martelli, G.; Bermano, G. Effects of astaxanthin in animal models of obesity-associated diseases: A systematic review and meta-analysis. Free Radic. Biol. Med. 2021, 171, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Moulin, M.; Ferreiro, A. Muscle redox disturbances and oxidative stress as pathomechanisms and therapeutic targets in early-onset myopathies. Semin. Cell Dev. Biol. 2017, 64, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M.; et al. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. J. Cachexia Sarcopenia Muscle 2020, 11, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Ima-Nirwana, S.; Chin, K.Y. Effects of astaxanthin on the protection of muscle health (Review). Exp. Ther. Med. 2020, 20, 2941–2952. [Google Scholar] [CrossRef] [PubMed]
- Farini, A.; Tripodi, L.; Villa, C.; Strati, F.; Facoetti, A.; Baselli, G.; Troisi, J.; Landolfi, A.; Lonati, C.; Molinaro, D.; et al. Microbiota dysbiosis influences immune system and muscle pathophysiology of dystrophin-deficient mice. EMBO Mol. Med. 2023, 15, e16244. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, H.; Pagano, E.; Paris, D.; Panza, E.; Cuozzo, M.; Moriello, C.; Piscitelli, F.; Abolghasemi, A.; Gazzerro, E.; Silvestri, C.; et al. Targeting gut dysbiosis against inflammation and impaired autophagy in Duchenne muscular dystrophy. EMBO Mol. Med. 2023, 15, e16225. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Wang, H.; Xiao, S.; Li, C.; Li, Y.; Liu, B. Xanthophyllomyces dendrorhous-Derived Astaxanthin Regulates Lipid Metabolism and Gut Microbiota in Obese Mice Induced by A High-Fat Diet. Mar. Drugs 2019, 17, 337. [Google Scholar] [CrossRef]
- Liu, D.; Ji, Y.; Cheng, Q.; Zhu, Y.; Zhang, H.; Guo, Y.; Cao, X.; Wang, H. Dietary astaxanthin-rich extract ameliorates atherosclerosis/retinopathy and restructures gut microbiome in apolipoprotein E-deficient mice fed on a high-fat diet. Food Funct. 2022, 13, 10461–10475. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, L.; Chin, Y.; Liu, F.; Li, R.W.; Yuan, S.; Xue, C.; Xu, J.; Tang, Q. Astaxanthin n-Octanoic Acid Diester Ameliorates Insulin Resistance and Modulates Gut Microbiota in High-Fat and High-Sucrose Diet-Fed Mice. Int. J. Mol. Sci. 2020, 21, 2149. [Google Scholar] [CrossRef]
- Tallis, J.; Shelley, S.; Degens, H.; Hill, C. Age-Related Skeletal Muscle Dysfunction Is Aggravated by Obesity: An Investigation of Contractile Function, Implications and Treatment. Biomolecules 2021, 11, 372. [Google Scholar] [CrossRef]
- Pérez-Díaz, S.; Koumaiha, Z.; Borok, M.J.; Aurade, F.; Pini, M.; Periou, B.; Rouault, C.; Baba-Amer, Y.; Clément, K.; Derumeaux, G.; et al. Obesity impairs skeletal muscle repair through NID-1 mediated extracellular matrix remodeling by mesenchymal progenitors. Matrix Biol. J. Int. Soc. Matrix Biol. 2022, 112, 90–115. [Google Scholar] [CrossRef] [PubMed]
- Tallis, J.; James, R.S.; Seebacher, F. The effects of obesity on skeletal muscle contractile function. J. Exp. Biol. 2018, 221, jeb163840. [Google Scholar] [CrossRef] [PubMed]
- Akhmedov, D.; Berdeaux, R. The effects of obesity on skeletal muscle regeneration. Front. Physiol. 2013, 4, 371. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.; James, R.S.; Cox, V.M.; Hill, C.; Tallis, J. Investigating a dose-response relationship between high-fat diet consumption and the contractile performance of isolated mouse soleus, EDL and diaphragm muscles. Eur. J. Appl. Physiol. 2019, 119, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Radley-Crabb, H.G.; Fiorotto, M.L.; Grounds, M.D. The different impact of a high fat diet on dystrophic mdx and control C57Bl/10 mice. PLoS Curr. 2011, 3, RRN1276. [Google Scholar] [CrossRef] [PubMed]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-inflammatory action of astaxanthin and its use in the treatment of various diseases. Biomed. Pharmacother. 2022, 145, 112179. [Google Scholar] [CrossRef] [PubMed]
- Tulangekar, A.; Sztal, T.E. Inflammation in Duchenne Muscular Dystrophy-Exploring the Role of Neutrophils in Muscle Damage and Regeneration. Biomedicines 2021, 9, 1366. [Google Scholar] [CrossRef]
- Grounds, M.D.; Terrill, J.R.; Al-Mshhdani, B.A.; Duong, M.N.; Radley-Crabb, H.G.; Arthur, P.G. Biomarkers for Duchenne muscular dystrophy: Myonecrosis, inflammation and oxidative stress. Dis. Models Mech. 2020, 13, dmm043638. [Google Scholar] [CrossRef]
- Kawamura, A.; Aoi, W.; Abe, R.; Kobayashi, Y.; Wada, S.; Kuwahata, M.; Higashi, A. Combined intake of astaxanthin, β-carotene, and resveratrol elevates protein synthesis during muscle hypertrophy in mice. Nutrition 2020, 69, 110561. [Google Scholar] [CrossRef]
- Shibaguchi, T.; Yamaguchi, Y.; Miyaji, N.; Yoshihara, T.; Naito, H.; Goto, K.; Ohmori, D.; Yoshioka, T.; Sugiura, T. Astaxanthin intake attenuates muscle atrophy caused by immobilization in rats. Physiol. Rep. 2016, 4, e12885. [Google Scholar] [CrossRef]
- Ren, P.; Yu, X.; Tang, Q.; Huan, Y.; Xu, J.; Wang, Y.; Xue, C. Astaxanthin Supplementation Assists Sorafenib in Slowing Skeletal Muscle Atrophy in H22 Tumor-Bearing Mice via Reversing Abnormal Glucose Metabolism. Mol. Nutr. Food Res. 2023, 67, e2300076. [Google Scholar] [CrossRef] [PubMed]
- Bandet, C.L.; Tan-Chen, S.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism: New Insight into Ceramide-Induced Lipotoxicity in Muscle Cells. Int. J. Mol. Sci. 2019, 20, 479. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.I.; Laurila, P.P.; Wohlwend, M.; Morel, J.D.; Goeminne, L.J.E.; Li, H.; Romani, M.; Li, X.; Oh, C.M.; Park, D.; et al. Inhibiting de novo ceramide synthesis restores mitochondrial and protein homeostasis in muscle aging. Sci. Transl. Med. 2023, 15, eade6509. [Google Scholar] [CrossRef] [PubMed]
- Bielawiec, P.; Harasim-Symbor, E.; Konstantynowicz-Nowicka, K.; Sztolsztener, K.; Chabowski, A. Chronic Cannabidiol Administration Attenuates Skeletal Muscle De Novo Ceramide Synthesis Pathway and Related Metabolic Effects in a Rat Model of High-Fat Diet-Induced Obesity. Biomolecules 2020, 10, 1241. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.P.; Shin, J.H.; Seo, S.H.; Kim, S.G.; Lee, S.H.; Shin, E.H. Effects of Antioxidants in Reducing Accumulation of Fat in Hepatocyte. Int. J. Mol. Sci. 2018, 19, 2563. [Google Scholar] [CrossRef]
- Ni, Y.; Nagashimada, M.; Zhuge, F.; Zhan, L.; Nagata, N.; Tsutsui, A.; Nakanuma, Y.; Kaneko, S.; Ota, T. Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E. Sci. Rep. 2015, 5, 17192. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-oxidation of saturated fatty acids in humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Starinets, V.S.; Belosludtseva, N.V.; Mikheeva, I.B.; Chelyadnikova, Y.A.; Penkina, D.K.; Vedernikov, A.A.; Belosludtsev, K.N. The Effect of Uridine on the State of Skeletal Muscles and the Functioning of Mitochondria in Duchenne Dystrophy. Int. J. Mol. Sci. 2022, 23, 660. [Google Scholar] [CrossRef]
- Wu, L.; Mo, W.; Feng, J.; Li, J.; Yu, Q.; Li, S.; Zhang, J.; Chen, K.; Ji, J.; Dai, W.; et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br. J. Pharmacol. 2020, 177, 3760–3777. [Google Scholar] [CrossRef]
- Van Hul, M.; Cani, P.D. The gut microbiota in obesity and weight management: Microbes as friends or foe? Nat. Rev. Endocrinol. 2023, 19, 258–271. [Google Scholar] [CrossRef]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Yu, X.; Yue, H.; Tang, Q.; Wang, Y.; Xue, C. Dietary supplementation with astaxanthin enhances anti-tumor immune response and aids the enhancement of molecularly targeted therapy for hepatocellular carcinoma. Food Funct. 2023, 14, 8309–8320. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Bose, S.; Lim, S.; Seo, J.; Shin, J.; Lee, D.; Chung, W.H.; Song, E.J.; Nam, Y.D.; Kim, H. Beneficial Effects of Newly Isolated Akkermansia muciniphila Strains from the Human Gut on Obesity and Metabolic Dysregulation. Microorganisms 2020, 8, 1413. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Noh, J.R.; Choe, D.; Lee, N.; Song, Y.; Cho, S.; Kang, E.J.; Go, M.J.; Ha, S.K.; Chang, D.H.; et al. Ageing and rejuvenation models reveal changes in key microbial communities associated with healthy ageing. Microbiome 2021, 9, 240. [Google Scholar] [CrossRef] [PubMed]
- Sakakida, T.; Ishikawa, T.; Doi, T.; Morita, R.; Endo, Y.; Matsumura, S.; Ota, T.; Yoshida, J.; Hirai, Y.; Mizushima, K.; et al. Water-soluble dietary fiber alleviates cancer-induced muscle wasting through changes in gut microenvironment in mice. Cancer Sci. 2022, 113, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zheng, T.; Hui, H.; Xie, G. Soybean isoflavones modulate gut microbiota to benefit the health weight and metabolism. Front. Cell. Infect. Microbiol. 2022, 12, 1004765. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin in obese mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2007, 71, 893–899. [Google Scholar] [CrossRef]
- Shatoor, A.S.; Al Humayed, S.; Almohiy, H.M. Astaxanthin attenuates hepatic steatosis in high-fat diet-fed rats by suppressing microRNA-21 via transactivation of nuclear factor erythroid 2-related factor 2. J. Physiol. Biochem. 2022, 78, 151–168. [Google Scholar] [CrossRef]
- Wang, M.; Ma, H.; Guan, S.; Luo, T.; Zhao, C.; Cai, G.; Zheng, Y.; Jia, X.; Di, J.; Li, R.; et al. Astaxanthin from Haematococcus pluvialis alleviates obesity by modulating lipid metabolism and gut microbiota in mice fed a high-fat diet. Food Funct. 2021, 12, 9719–9738. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | ND | HFD | AX |
|---|---|---|---|
| Casein | 200 | 258 | 258 |
| L-Cystine | 3 | 4 | 4 |
| Sucrose | 100 | 89 | 89 |
| Cornstarch | 397.5 | - | - |
| Dyetrose | 132 | 161 | 161 |
| Lard | - | 317 | 317 |
| Soybean Oil | 70 | 32 | 32 |
| Cellulose | 50 | 65 | 65 |
| Mineral Mix | 35 | 58 | 57.2 |
| Vitamin Mix | 10 | 13 | 13 |
| Choline Bitartrate | 2.5 | 3 | 3 |
| Astaxanthin | - | - | 0.2 |
| Total | 1000 | 1000 | 1000 |
| Protein (%) | 20.3 | 20.0 | 20.0 |
| Carbohydrate (%) | 63.9 | 20.0 | 20.0 |
| Fat (%) | 15.8 | 60.0 | 60.0 |
| Energy density (kcal/g) | 4.00 | 5.26 | 5.26 |
| Lipid Species | Class | Subclass | HFD vs. ND | AX vs. HFD | ||||
|---|---|---|---|---|---|---|---|---|
| FC | VIP | Trend | FC | VIP | Trend | |||
| TG (16:0_11:3_14:1) | GL | TG | 9.93 | 1.33 | Up | 0.39 | 1.38 | Down |
| TG (16:0_11:3_16:0) | GL | TG | 8.05 | 1.38 | Up | 0.49 | 1.61 | Down |
| TG (16:0_11:4_16:0) | GL | TG | 6.17 | 1.31 | Up | 0.29 | 1.57 | Down |
| TG (16:0_11:4_18:1) | GL | TG | 2.47 | 1.10 | Up | 0.43 | 1.49 | Down |
| TG (16:1_11:2_14:0) | GL | TG | 6.52 | 1.37 | Up | 0.41 | 1.57 | Down |
| TG (18:0e_16:0_18:1) | GL | TG | 5.67 | 1.39 | Up | 0.41 | 1.40 | Down |
| TG (18:1_18:1_23:0) | GL | TG | 4.06 | 1.35 | Up | 0.44 | 1.34 | Down |
| TG (18:1_18:1_24:0) | GL | TG | 4.39 | 1.34 | Up | 0.43 | 1.36 | Down |
| TG (20:1e_16:0_18:1) | GL | TG | 3.6 | 1.34 | Up | 0.39 | 1.35 | Down |
| TG (25:0_18:1_18:1) | GL | TG | 3.75 | 1.34 | Up | 0.45 | 1.34 | Down |
| TG (26:0_18:1_24:2) | GL | TG | 4.55 | 1.36 | Up | 0.5 | 1.36 | Down |
| TG (30:0_18:1_24:2) | GL | TG | 5.75 | 1.36 | Up | 0.5 | 1.35 | Down |
| TG (61:2e) | GL | TG | 3.57 | 1.33 | Up | 0.45 | 1.37 | Down |
| TG (70:3) | GL | TG | 10.89 | 1.40 | Up | 0.43 | 1.40 | Down |
| TG (72:4) | GL | TG | 6.61 | 1.38 | Up | 0.41 | 1.37 | Down |
| TG (16:0_16:0_22:6) | GL | TG | 0.32 | 1.31 | Down | 2.04 | 1.55 | Up |
| DG (50:0) | GL | DG | 2.39 | 1.27 | Up | 0.46 | 1.61 | Down |
| DG (54:2) | GL | DG | 3.6 | 1.34 | Up | 0.39 | 1.35 | Down |
| PG (20:5_22:6) | GP | PG | 0.23 | 1.37 | Down | 2.04 | 1.43 | Up |
| PG (38:4) | GP | PG | 0.46 | 1.20 | Down | 2.12 | 1.44 | Up |
| PG (42:7) | GP | PG | 0.43 | 1.27 | Down | 2.37 | 1.54 | Up |
| PC (28:1_18:2) | GP | PC | 3.09 | 1.02 | Up | 0.38 | 1.38 | Down |
| CL (18:1_16:0_16:0_18:1) | GP | CL | 0.29 | 1.27 | Down | 4.52 | 1.49 | Up |
| CL (18:2_18:1_20:4_16:0) | GP | CL | 0.4 | 1.23 | Down | 2.99 | 1.54 | Up |
| CL (18:2_18:1_22:4_18:2) | GP | CL | 0.37 | 1.08 | Down | 4.09 | 1.47 | Up |
| CL (18:2_20:4_18:1_18:1) | GP | CL | 0.33 | 1.30 | Down | 3.55 | 1.59 | Up |
| CL (69:5) | GP | CL | 0.36 | 1.27 | Down | 2.5 | 1.46 | Up |
| SM (t18:0_24:3) | SP | SM | 2.92 | 1.22 | Up | 0.46 | 1.52 | Down |
| Cer (d18:2_21:2) | SP | Cer | 0.2 | 1.28 | Down | 2.94 | 1.34 | Up |
| Cer (m18:1_18:0) | SP | Cer | 8.73 | 1.38 | Up | 0.42 | 1.68 | Down |
| Cer (m18:1_20:0) | SP | Cer | 5.86 | 1.34 | Up | 0.48 | 1.50 | Down |
| Cer (m18:1_21:0) | SP | Cer | 4.07 | 1.27 | Up | 0.42 | 1.64 | Down |
| Cer (t42:3) | SP | Cer | 4.51 | 1.12 | Up | 0.27 | 1.54 | Down |
| CerG2GNAc1 (d42:1 + O) | SL | CerG2GNAc1 | 0.21 | 1.25 | Down | 2.8 | 1.52 | Up |
| WE (20:1_16:0) | WE | WE | 6.15 | 1.39 | Up | 0.49 | 1.65 | Down |
| WE (21:1_16:0) | WE | WE | 4.28 | 1.33 | Up | 0.4 | 1.47 | Down |
| WE (22:1_16:0) | WE | WE | 5.06 | 1.33 | Up | 0.43 | 1.61 | Down |
| ZyE (33:0) | ZyE | ZyE | 2.5 | 1.16 | Up | 0.4 | 1.35 | Down |
| DG (18:2_18:2) | GL | DG | 0.46 | 1.03 | Down | 0.33 | 1.33 | Down |
| SQDG (51:12) | GL | SQDG | 6.74 | 1.17 | Up | 2.01 | 1.59 | Up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Ling, C.; Chen, M.; Yu, L.; Yang, J.; Fang, Q. Astaxanthin Ameliorates Worsened Muscle Dysfunction of MDX Mice Fed with a High-Fat Diet through Reducing Lipotoxicity and Regulating Gut Microbiota. Nutrients 2024, 16, 33. https://doi.org/10.3390/nu16010033
Chen Y, Ling C, Chen M, Yu L, Yang J, Fang Q. Astaxanthin Ameliorates Worsened Muscle Dysfunction of MDX Mice Fed with a High-Fat Diet through Reducing Lipotoxicity and Regulating Gut Microbiota. Nutrients. 2024; 16(1):33. https://doi.org/10.3390/nu16010033
Chicago/Turabian StyleChen, Ying, Chenjie Ling, Mengting Chen, Liqiang Yu, Jing Yang, and Qi Fang. 2024. "Astaxanthin Ameliorates Worsened Muscle Dysfunction of MDX Mice Fed with a High-Fat Diet through Reducing Lipotoxicity and Regulating Gut Microbiota" Nutrients 16, no. 1: 33. https://doi.org/10.3390/nu16010033