

Polymorphisms of Fat Mass and Obesity-Associated Gene in the Pathogenesis of Child and Adolescent Metabolic Syndrome
 , ,
, ,
Abstract
:1. Background
1.1. Incidence of Pediatric Metabolic Syndrome
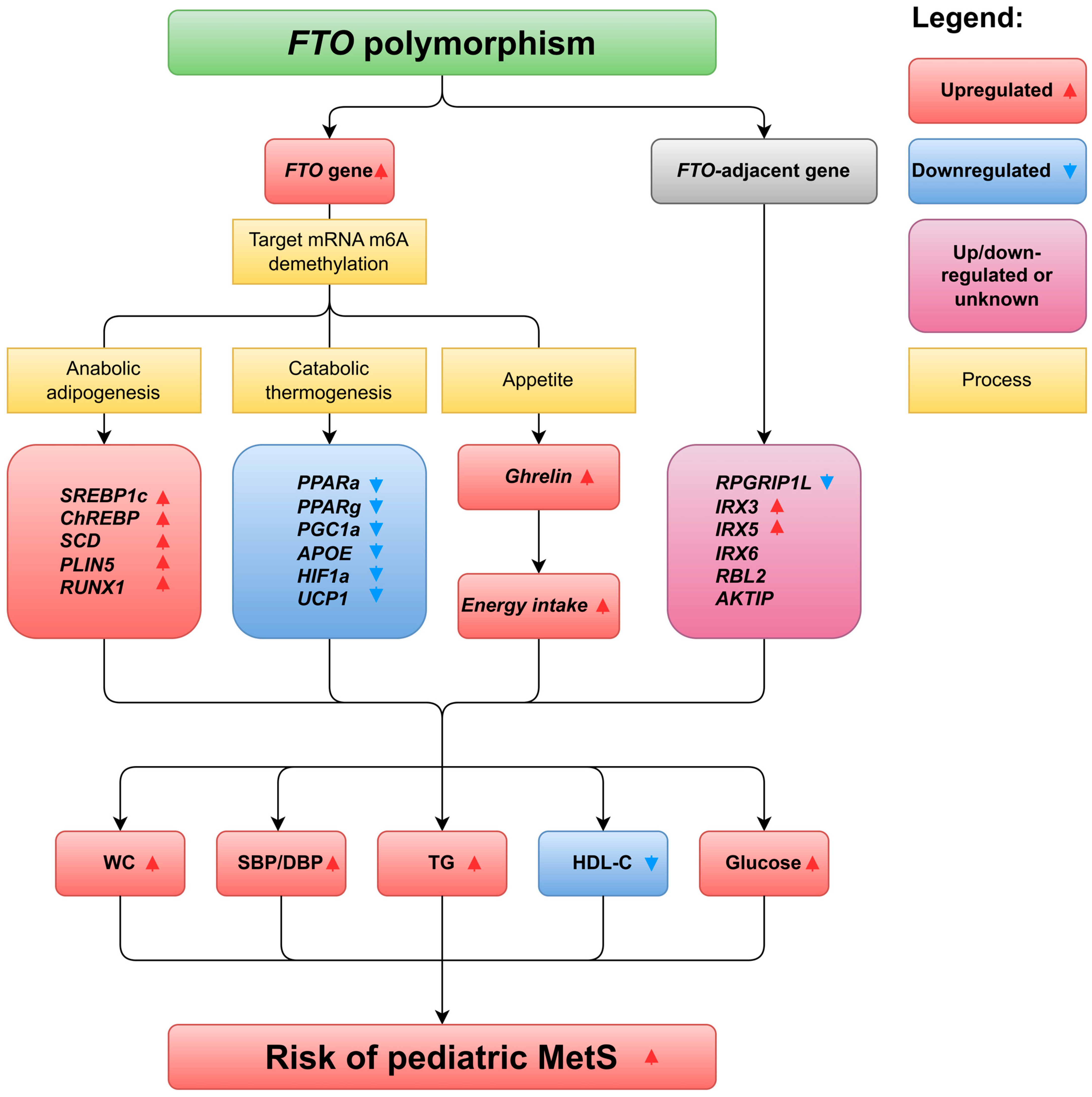
1.2. Fat Mass and Obesity-Associated Gene in Pediatric MetS
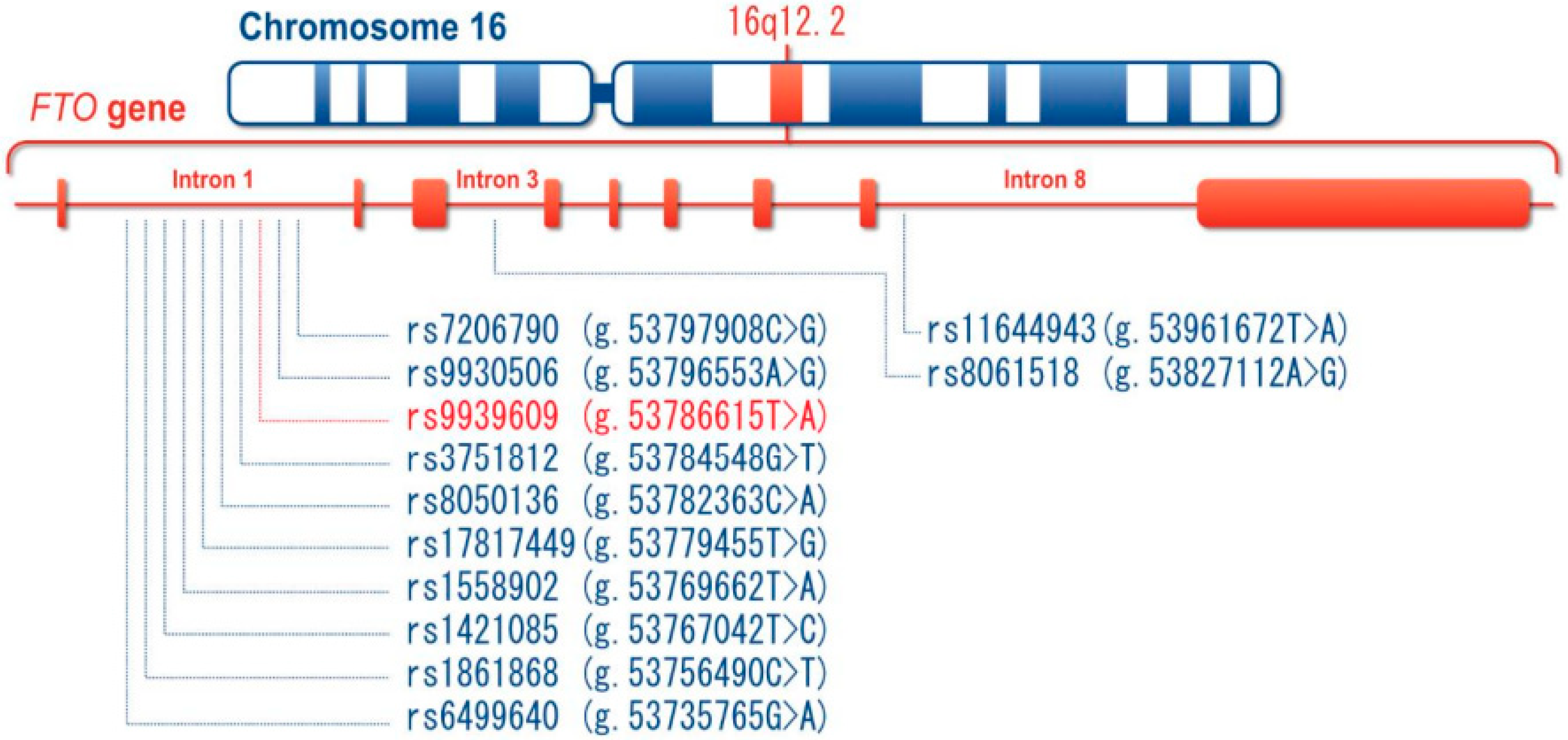
2. Some Genetic Polymorphisms of FTO Increase Susceptibility to MetS
2.1. The SNPs of FTO in MetS: rs9939609 and rs9930506 Polymorphisms
2.1.1. The rs9939609 Polymorphism
2.1.2. The rs9930506 Polymorphism
2.1.3. The Interaction between FTO Polymorphism and Adjacent Genes
2.2. The rs9939609 Polymorphism of FTO Is Related to Larger WC in Child and Adolescent Obesity
2.2.1. Mechanism of rs9939609 Polymorphism Affects Pediatric WC
2.2.2. Other FTO Polymorphisms Associated with Abnormal WC in Children and Adolescents
2.3. FTO Polymorphisms Regulate BP in Children and Adolescents
2.3.1. The Link between Hypertension and FTO rs9939609 Polymorphism in Children and Adolescents
2.3.2. Other FTO Polymorphisms in the Pathogenesis of Hypertension in Children and Adolescents
2.4. FTO Polymorphisms Regulate Lipid Metabolism in Children and Adolescents
2.4.1. The rs9939609 Polymorphism Increases Risk of Hypertriglyceridemia
2.4.2. FTO Polymorphisms Are Involved in HDL Metabolism in Children and Adolescents
2.5. FTO Polymorphisms Modulate Blood Glucose Levels in Children and Adolescents
2.5.1. The rs9939609 Polymorphism in Hyperglycemia and Diabetes
2.5.2. Other FTO Polymorphisms in Hyperglycemia
3. Advances in Prevention and Treatment of MetS in Children and Adolescents
4. Conclusions and Future Perspectives
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alanaeme, C.J.; Bittner, V.; Brown, T.M.; Colantonio, L.D.; Dhalwani, N.; Jones, J.; Kalich, B.; Exter, J.; Jackson, E.A.; Levitan, E.B.; et al. Estimated number and percentage of US adults with atherosclerotic cardiovascular disease recommended add-on lipid-lowering therapy by the 2018 AHA/ACC multi-society cholesterol guideline. Am. Heart J. Plus 2022, 21, 100201. [Google Scholar] [CrossRef] [PubMed]
- Pál, É.; Ungvári, Z.; Benyó, Z.; Várbíró, S. Role of Vitamin D Deficiency in the Pathogenesis of Cardiovascular and Cerebrovascular Diseases. Nutrients 2023, 15, 334. [Google Scholar] [CrossRef] [PubMed]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics-2023 Update: A Report From the American Heart Association. Circulation 2023, 147, e93–e621. [Google Scholar] [CrossRef] [PubMed]
- Nagrani, R.; Marron, M.; Bongaerts, E.; Nawrot, T.S.; Ameloot, M.; de Hoogh, K.; Vienneau, D.; Lequy, E.; Jacquemin, B.; Guenther, K.; et al. Association of urinary and ambient black carbon, and other ambient air pollutants with risk of prediabetes and metabolic syndrome in children and adolescents. Environ. Pollut. 2023, 317, 120773. [Google Scholar] [CrossRef]
- DeBoer, M.D.; Filipp, S.L.; Gurka, M.J. Geographical variation in the prevalence of obesity and metabolic syndrome among US adolescents. Pediatr. Obes. 2019, 14, e12483. [Google Scholar] [CrossRef]
- Lee, A.M.; Gurka, M.J.; DeBoer, M.D. Trends in Metabolic Syndrome Severity and Lifestyle Factors Among Adolescents. Pediatrics 2016, 137, e20153177. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zheng, H.; Zou, Z.; Jing, J.; Ma, Y.; Wang, H.; Luo, J.; Zhang, X.; Luo, C.; Wang, H.; et al. Metabolic Syndrome and Related Factors in Chinese Children and Adolescents: Analysis from a Chinese National Study. J. Atheroscler. Thromb. 2020, 27, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Andrabi, S.M.; Bhat, M.H.; Andrabi, S.R.; Kamili, M.M.; Imran, A.; Nisar, I.; Nisar, U. Prevalence of metabolic syndrome in 8-18-year-old school-going children of Srinagar city of Kashmir India. Indian J. Endocrinol. Metab. 2013, 17, 95–100. [Google Scholar] [CrossRef]
- Shi, J.; He, L.; Yu, D.; Ju, L.; Guo, Q.; Piao, W.; Xu, X.; Zhao, L.; Yuan, X.; Cao, Q.; et al. Prevalence and Correlates of Metabolic Syndrome and Its Components in Chinese Children and Adolescents Aged 7–17: The China National Nutrition and Health Survey of Children and Lactating Mothers from 2016–2017. Nutrients 2022, 14, 3348. [Google Scholar] [CrossRef]
- Laru, J.; Nedelec, R.; Koivuaho, E.; Ojaniemi, M.; Järvelin, M.R.; Tapanainen, J.S.; Franks, S.; Tolvanen, M.; Piltonen, T.T.; Sebert, S.; et al. BMI in childhood and adolescence is associated with impaired reproductive function-a population-based cohort study from birth to age 50 years. Hum. Reprod. 2021, 36, 2948–2961. [Google Scholar] [CrossRef]
- Ojanen, X.; Cheng, R.; Törmäkangas, T.; Rappaport, N.; Wilmanski, T.; Wu, N.; Fung, E.; Nedelec, R.; Sebert, S.; Vlachopoulos, D.; et al. Towards early risk biomarkers: Serum metabolic signature in childhood predicts cardio-metabolic risk in adulthood. EBioMedicine 2021, 72, 103611. [Google Scholar] [CrossRef]
- Weihe, P.; Spielmann, J.; Kielstein, H.; Henning-Klusmann, J.; Weihrauch-Blüher, S. Childhood Obesity and Cancer Risk in Adulthood. Curr. Obes. Rep. 2020, 9, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhao, Y.; Zhao, J.; Bian, M.; Qian, L.; Xue, T.; Zhang, J.J.; Duan, X. Physical activity attenuated the associations between ambient air pollutants and metabolic syndrome (MetS): A nationwide study across 28 provinces. Environ. Pollut. 2022, 315, 120348. [Google Scholar] [CrossRef] [PubMed]
- Seral-Cortes, M.; Larruy-García, A.; De Miguel-Etayo, P.; Labayen, I.; Moreno, L.A. Mediterranean Diet and Genetic Determinants of Obesity and Metabolic Syndrome in European Children and Adolescents. Genes 2022, 13, 420. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, L.; Anania, C.; Martino, F.; Poggiogalle, E.; Chiarelli, F.; Arca, M.; Chiesa, C. Management of metabolic syndrome in children and adolescents. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 455–466. [Google Scholar] [CrossRef]
- Titmuss, A.T.; Srinivasan, S. Metabolic syndrome in children and adolescents: Old concepts in a young population. J. Paediatr. Child Health 2016, 52, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Tavares Giannini, D.; Caetano Kuschnir, M.C.; Szklo, M. Metabolic syndrome in overweight and obese adolescents: A comparison of two different diagnostic criteria. Ann. Nutr. Metab. 2014, 64, 71–79. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Lappalainen, T.; Kolehmainen, M.; Schwab, U.; Pulkkinen, L.; de Mello, V.D.; Vaittinen, M.; Laaksonen, D.E.; Poutanen, K.; Uusitupa, M.; Gylling, H. Gene expression of FTO in human subcutaneous adipose tissue, peripheral blood mononuclear cells and adipocyte cell line. J. Nutr. Nutr. 2010, 3, 37–45. [Google Scholar] [CrossRef]
- Villalobos-Comparán, M.; Teresa Flores-Dorantes, M.; Teresa Villarreal-Molina, M.; Rodríguez-Cruz, M.; García-Ulloa, A.C.; Robles, L.; Huertas-Vázquez, A.; Saucedo-Villarreal, N.; López-Alarcón, M.; Sánchez-Muñoz, F.; et al. The FTO gene is associated with adulthood obesity in the Mexican population. Obesity 2008, 16, 2296–2301. [Google Scholar] [CrossRef]
- Church, C.; Moir, L.; McMurray, F.; Girard, C.; Banks, G.T.; Teboul, L.; Wells, S.; Brüning, J.C.; Nolan, P.M.; Ashcroft, F.M. Overexpression of FTO leads to increased food intake and results in obesity. Nat. Genet. 2010, 42, 1086–1092. [Google Scholar] [CrossRef]
- Fischer, J.; Koch, L.; Emmerling, C.; Vierkotten, J.; Peters, T.; Brüning, J.C.; Rüther, U. Inactivation of the FTO gene protects from obesity. Nature 2009, 458, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Guo, G.; Bi, Z.; Liu, Y.; Zhao, Y.; Chen, N.; Wang, F.; Wang, Y.; Wang, X. m6A methylation modulates adipogenesis through JAK2-STAT3-C/EBPβ signaling. Biochim. Biophys. Acta -Gene Regul. Mech. 2019, 1862, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Luo, Y.; Jia, G.; Liu, G.; Zhao, H.; Huang, Z. FTO promotes adipogenesis through inhibition of the Wnt/β-catenin signaling pathway in porcine intramuscular preadipocytes. Anim. Biotechnol. 2017, 28, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Laber, S.; McMurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.; Usher, S.; Sellayah, D.; Ashcroft, F.M. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6, 6792. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, Y.; Ma, J.; Guo, F.; Cao, Q.; Zhang, Y.; Zhou, B.; Chai, J.; Zhao, W.; Zhao, R. The demethylase activity of FTO (fat mass and obesity associated protein) is required for preadipocyte differentiation. PLoS ONE 2015, 10, e0133788. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kim, J.W.; Grønborg, M.; Urlaub, H.; Lane, M.D.; Tang, Q.-Q. Role of cdk2 in the sequential phosphorylation/activation of C/EBPβ during adipocyte differentiation. Proc. Natl. Acad. Sci. USA 2007, 104, 11597–11602. [Google Scholar] [CrossRef] [Green Version]
- Almén, M.S.; Jacobsson, J.A.; Moschonis, G.; Benedict, C.; Chrousos, G.P.; Fredriksson, R.; Schiöth, H.B. Genome wide analysis reveals association of a FTO gene variant with epigenetic changes. Genomics 2012, 99, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Skuladottir, G.V.; Oskarsdottir, H.; Pisanu, C.; Sjödin, M.; Lindberg, J.; Mwinyi, J.; Schiöth, H.B. Plasma stearoyl-CoA desaturase activity indices and bile acid concentrations after a low-fat meal: Association with a genetic variant in the FTO gene. Diabetes Metab. Syndr. Obes. 2018, 11, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Villalobos-Comparán, M.; Antuna-Puente, B.; Villarreal-Molina, M.T.; Canizales-Quinteros, S.; Velázquez-Cruz, R.; León-Mimila, P.; Villamil-Ramírez, H.; González-Barrios, J.A.; Merino-García, J.L.; Thompson-Bonilla, M.R.; et al. Interaction between FTO rs9939609 and the Native American-origin ABCA1 rs9282541 affects BMI in the admixed Mexican population. BMC Med. Genet. 2017, 18, 46. [Google Scholar] [CrossRef] [Green Version]
- Berulava, T.; Horsthemke, B. The obesity-associated SNPs in intron 1 of the FTO gene affect primary transcript levels. Eur. J. Hum. Genet. 2010, 18, 1054–1056. [Google Scholar] [CrossRef] [PubMed]
- Karra, E.; O’Daly, O.G.; Choudhury, A.I.; Yousseif, A.; Millership, S.; Neary, M.T.; Scott, W.R.; Chandarana, K.; Manning, S.; Hess, M.E.; et al. A link between FTO, ghrelin, and impaired brain food-cue responsivity. J. Clin. Investig. 2013, 123, 3539–3551. [Google Scholar] [CrossRef] [Green Version]
- Grunnet, L.G.; Brøns, C.; Jacobsen, S.; Nilsson, E.; Astrup, A.; Hansen, T.; Pedersen, O.; Poulsen, P.; Quistorff, B.; Vaag, A. Increased recovery rates of phosphocreatine and inorganic phosphate after isometric contraction in oxidative muscle fibers and elevated hepatic insulin resistance in homozygous carriers of the A-allele of FTO rs9939609. J. Clin. Endocrinol. Metab. 2009, 94, 596–602. [Google Scholar] [CrossRef] [Green Version]
- Doaei, S.; Kalantari, N.; Izadi, P.; Salonurmi, T.; Mosavi Jarrahi, A.; Rafieifar, S.; Azizi Tabesh, G.; Rahimzadeh, G.; Gholamalizadeh, M.; Goodarzi, M.O. Changes in FTO and IRX3 gene expression in obese and overweight male adolescents undergoing an intensive lifestyle intervention and the role of FTO genotype in this interaction. J. Transl. Med. 2019, 17, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javanrouh, N.; Daneshpour, M.S.; Soltanian, A.R.; Tapak, L. Kernel machine SNP set analysis provides new insight into the association between obesity and polymorphisms located on the chromosomal 16q.12.2 region: Tehran Lipid and Glucose Study. Gene 2018, 658, 146–151. [Google Scholar] [CrossRef]
- Javanrouh, N.; Soltanian, A.R.; Tapak, L.; Azizi, F.; Ott, J.; Daneshpour, M.S. A novel association of rs13334070 in the RPGRIP1L gene with adiposity factors discovered by joint linkage and linkage disequilibrium analysis in Iranian pedigrees: Tehran Cardiometabolic Genetic Study (TCGS). Genet. Epidemiol. 2019, 43, 342–351. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, G.; Yi, J.; Li, Q.; Tan, Z.; Fan, W.; Luo, X.; He, Z.; Si, Z.; Li, J. IRX3 plays an important role in the pathogenesis of metabolic-associated fatty liver disease by regulating hepatic lipid metabolism. Front. Endocrinol. 2022, 13, 895593. [Google Scholar] [CrossRef] [PubMed]
- Sobalska-Kwapis, M.; Suchanecka, A.; Słomka, M.; Siewierska-Górska, A.; Kępka, E.; Strapagiel, D. Genetic association of FTO/IRX region with obesity and overweight in the Polish population. PLoS ONE 2017, 12, e0180295. [Google Scholar] [CrossRef]
- Sung, Y.J.; Winkler, T.W.; de Las Fuentes, L.; Bentley, A.R.; Brown, M.R.; Kraja, A.T.; Schwander, K.; Ntalla, I.; Guo, X.; Franceschini, N.; et al. A Large-Scale Multi-ancestry Genome-wide Study Accounting for Smoking Behavior Identifies Multiple Significant Loci for Blood Pressure. Am. J. Hum. Genet. 2018, 102, 375–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Wu, D.; Zhang, C.; Yan, T.; Zhao, Y.; Shen, H.; Xue, K.; Huang, X.; Wang, Z.; Qiu, Y. Macrophage IRX3 promotes diet-induced obesity and metabolic inflammation. Nat. Immunol. 2021, 22, 1268–1279. [Google Scholar] [CrossRef]
- Yang, M.; Xu, Y.; Liang, L.; Fu, J.; Xiong, F.; Liu, G.; Gong, C.; Luo, F.; Chen, S.; Xu, C.; et al. The effects of genetic variation in FTO rs9939609 on obesity and dietary preferences in Chinese Han children and adolescents. PLoS ONE 2014, 9, e104574. [Google Scholar] [CrossRef]
- Albuquerque, D.; Nóbrega, C.; Manco, L. Association of FTO polymorphisms with obesity and obesity-related outcomes in Portuguese children. PLoS ONE 2013, 8, e54370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakanen, M.; Raitakari, O.T.; Lehtimäki, T.; Peltonen, N.; Pahkala, K.; Sillanmäki, L.; Lagström, H.; Viikari, J.; Simell, O.; Rönnemaa, T. FTO genotype is associated with body mass index after the age of seven years but not with energy intake or leisure-time physical activity. J. Clin. Endocrinol. Metab. 2009, 94, 1281–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labayen, I.; Ruiz, J.R.; Huybrechts, I.; Ortega, F.B.; Arenaza, L.; González-Gross, M.; Widhalm, K.; Molnar, D.; Manios, Y.; DeHenauw, S.; et al. Dietary fat intake modifies the influence of the FTO rs9939609 polymorphism on adiposity in adolescents: The HELENA cross-sectional study. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauria, F.; Siani, A.; Bammann, K.; Foraita, R.; Huybrechts, I.; Iacoviello, L.; Koni, A.C.; Kourides, Y.; Marild, S.; Molnar, D.; et al. Prospective analysis of the association of a common variant of FTO (rs9939609) with adiposity in children: Results of the IDEFICS study. PLoS ONE 2012, 7, e48876. [Google Scholar] [CrossRef]
- Mangge, H.; Renner, W.; Almer, G.; Weghuber, D.; Möller, R.; Horejsi, R. Rs9939609 variant of the fat mass and obesity-associated gene and trunk obesity in adolescents. J. Obes. 2011, 2011, 186368. [Google Scholar] [CrossRef]
- Olza, J.; Ruperez, A.I.; Gil-Campos, M.; Leis, R.; Fernandez-Orth, D.; Tojo, R.; Cañete, R.; Gil, A.; Aguilera, C.M. Influence of FTO variants on obesity, inflammation and cardiovascular disease risk biomarkers in Spanish children: A case-control multicentre study. BMC Med. Genet. 2013, 14, 123. [Google Scholar] [CrossRef] [Green Version]
- Wardle, J.; Carnell, S.; Haworth, C.M.; Farooqi, I.S.; O’Rahilly, S.; Plomin, R. Obesity associated genetic variation in FTO is associated with diminished satiety. J. Clin. Endocrinol. Metab. 2008, 93, 3640–3643. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Kim, I.K.; Kang, J.H.; Ahn, Y.; Han, B.G.; Lee, J.Y.; Song, J. Effects of common FTO gene variants associated with BMI on dietary intake and physical activity in Koreans. Clin. Chim. Acta 2010, 411, 1716–1722. [Google Scholar] [CrossRef]
- Li, M.; Zhang, T.; Xu, P.R. Study of the association between MC4R and FTO genotypes on childhood obesity. J. Xinjiang Med. Univ. 2020, 43, 6–9. [Google Scholar]
- Xi, B.; Cheng, H.; Shen, Y.; Chandak, G.R.; Zhao, X.; Hou, D.; Wu, L.; Wang, X.; Mi, J. Study of 11 BMI-associated loci identified in GWAS for associations with central obesity in the Chinese children. PLoS ONE 2013, 8, e56472. [Google Scholar] [CrossRef] [Green Version]
- Xi, B.; Shen, Y.; Zhang, M.; Liu, X.; Zhao, X.; Wu, L.; Cheng, H.; Hou, D.; Lindpaintner, K.; Liu, L.; et al. The common rs9939609 variant of the fat mass and obesity-associated gene is associated with obesity risk in children and adolescents of Beijing, China. BMC Med. Genet. 2010, 11, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todendi, P.F.; Klinger, E.I.; Geraldo, A.C.R.; Brixner, L.; Reuter, C.P.; Lindenau, J.D.R.; Valim, A.R.M.; Fiegenbaum, M. Genetic risk score based on fat mass and obesity-associated, transmembrane protein 18 and fibronectin type III domain containing 5 polymorphisms is associated with anthropometric characteristics in South Brazilian children and adolescents. Br. J. Nutr. 2019, 121, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todendi, P.F.; Martínez, J.A.; Reuter, C.P.; Klinger, E.I.; Fiegenbaum, M.; de Moura Valim, A.R. Influence of FTO (Fat mass and obesity) gene and parental obesity on Brazilian children and adolescents adiposity. J. Pediatr. Endocrinol. Metab. 2020, 33, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, N.; Villagrán, M.; Riffo, B.; Gleisner, A.; Petermann-Rocha, F.; Mardones, L.; Leiva, A.M.; Martínez-Sanguinetti, M.A.; Celis-Morales, C. Association between FTO gene rs9939609 and adiposity markers in Chilean children. Rev. Chil. Pediatr. 2020, 91, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhu, H.; Lagou, V.; Gutin, B.; Stallmann-Jorgensen, I.S.; Treiber, F.A.; Dong, Y.; Snieder, H. FTO variant rs9939609 is associated with body mass index and waist circumference, but not with energy intake or physical activity in European- and African-American youth. BMC Med. Genet. 2010, 11, 57. [Google Scholar] [CrossRef] [Green Version]
- Reuter, É.M.; Reuter, C.P.; de Castro Silveira, J.F.; Carroll, S.; Hobkirk, J.P.; Todendi, P.F.; de Moura Valim, A.R.; de Mello, E.D. FTO gene polymorphism and longitudinal changes in nutritional/obesity status in children and adolescents: Schoolchildren’s health cohort study. Eur. J. Pediatr. 2021, 180, 3325–3333. [Google Scholar] [CrossRef]
- Liang, J.J.; Chen, R.S.; Xue, X.N. A case-control study of the association of a common variant of FTO (rs9939609) with adiposity in children in Guangdong. Clin. J. Chin. Med. 2015, 7, 112–114. [Google Scholar]
- Dušátková, L.; Zamrazilová, H.; Aldhoon-Hainerová, I.; Sedláčková, B.; Včelák, J.; Hlavatý, P.; Bendlová, B.; Kunešová, M.; Hainer, V. A common variant near BDNF is associated with dietary calcium intake in adolescents. Nutr. Res. 2015, 35, 766–773. [Google Scholar] [CrossRef]
- Lazopoulou, N.; Gkioka, E.; Ntalla, I.; Pervanidou, P.; Magiakou, A.M.; Roma-Giannikou, E.; Chrousos, G.P.; Papassotiriou, I.; Dedoussis, G.; Kanaka-Gantenbein, C. The combined effect of MC4R and FTO risk alleles on childhood obesity in Greece. Hormones 2015, 14, 126–133. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, H.; Dong, Y.; Podolsky, R.H.; Treiber, F.A.; Snieder, H. Influence of common variants in FTO and near INSIG2 and MC4R on growth curves for adiposity in African- and European-American youth. Eur. J. Epidemiol. 2011, 26, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, J.R.; Labayen, I.; Ortega, F.B.; Legry, V.; Moreno, L.A.; Dallongeville, J.; Martínez-Gómez, D.; Bokor, S.; Manios, Y.; Ciarapica, D.; et al. Attenuation of the effect of the FTO rs9939609 polymorphism on total and central body fat by physical activity in adolescents: The HELENA study. Arch. Pediatr. Adolesc. Med. 2010, 164, 328–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luczynski, W.; Zalewski, G.; Bossowski, A. The association of the FTO rs9939609 polymorphism with obesity and metabolic risk factors for cardiovascular diseases in Polish children. J. Physiol. Pharmacol. 2012, 63, 241–248. [Google Scholar]
- Łuczyński, W.; Szypowska, A.; Głowińska-Olszewska, B.; Szadkowska, A.; Bossowski, A. Disease associated clinical factors and FTO polymorphism: Effect on body mass in children with type 1 diabetes mellitus. Pediatr. Diabetes 2014, 15, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Tercjak-Rećko, M.; Luczyński, W.; Bernatowicz, P.; Zalewski, G.; Rembińska, M.; Lachowska, U.; Rećko, P.; Suchoń, P.; Czaban, M.; Sokal, J.; et al. Polymorphism rs9939609 of FTO gene is related to the body mass index in children from Podlaskie voievodship. Med. Wieku Rozw. 2012, 16, 53–60. [Google Scholar]
- Dwivedi, O.P.; Tabassum, R.; Chauhan, G.; Ghosh, S.; Marwaha, R.K.; Tandon, N.; Bharadwaj, D. Common variants of FTO are associated with childhood obesity in a cross-sectional study of 3126 urban Indian children. PLoS ONE 2012, 7, e47772. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhao, X.; Xi, B.; Shen, Y.; Wu, L.; Cheng, H.; Hou, D.; Mi, J. Impact of obesity-related gene polymorphism on risk of obesity and metabolic disorder in childhood. Chin. J. Prev. Med. 2014, 48, 776–783. [Google Scholar]
- Ferreira Todendi, P.; de Moura Valim, A.R.; Klinger, E.; Reuter, C.P.; Molina, S.; Martínez, J.A.; Fiegenbaum, M. The role of the genetic variants IRX3 rs3751723 and FTO rs9939609 in the obesity phenotypes of children and adolescents. Obes. Res. Clin. Pract. 2019, 13, 137–142. [Google Scholar] [CrossRef]
- Muñoz-Yáñez, C.; Pérez-Morales, R.; Moreno-Macías, H.; Calleros-Rincón, E.; Ballesteros, G.; González, R.A.; Espinosa, J. Polymorphisms FTO rs9939609, PPARG rs1801282 and ADIPOQ rs4632532 and rs182052 but not lifestyle are associated with obesity related-traits in Mexican children. Genet. Mol. Biol. 2016, 39, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Reuter, C.P.; Burgos, M.S.; Bernhard, J.C.; Tornquist, D.; Klinger, E.I.; Borges, T.S.; Renner, J.D.; de Moura Valim, A.R.; de Mello, E.D. Association between overweight and obesity in schoolchildren with rs9939609 polymorphism (FTO) and family history for obesity. J. Pediatr. 2016, 92, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Reuter, C.P.; Rosane De Moura Valim, A.; Gaya, A.R.; Borges, T.S.; Klinger, E.I.; Possuelo, L.G.; Franke, S.I.; Kmetzsch, L.; Vainstein, M.H.; Prá, D.; et al. FTO polymorphism, cardiorespiratory fitness, and obesity in Brazilian youth. Am. J. Hum. Biol. 2016, 28, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Klimentidis, Y.C.; Chen, G.B.; López-Alarcón, M.; Harris, J.J.; Duarte, C.W.; Fernández, J.R. Associations of obesity genes with obesity-related outcomes in multiethnic children. Arch. Med. Res. 2011, 42, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Luque, R.; Ulloa, N.; Romero-Saldaña, M.; Zilic, M.; Gleisner, A.; Lanuza, F.; Molina-Recio, G. Association between the FTO SNP rs9939609 and Metabolic Syndrome in Chilean Children. Nutrients 2021, 13, 2014. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.M.; Furtado, J.M.; Mascarenhas, P.; Ferraz, M.E.; Ferreira, J.C.; Monteiro, M.P.; Vilanova, M.; Ferraz, F.P. Association between LEPR, FTO, MC4R, and PPARG-2 polymorphisms with obesity traits and metabolic phenotypes in school-aged children. Endocrine 2018, 60, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iskandar, K.; Patria, S.Y.; Huriyati, E.; Luglio, H.F.; Julia, M.; Susilowati, R. Effect of FTO rs9939609 variant on insulin resistance in obese female adolescents. BMC Res. Notes 2018, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Reuter, É.M.; Reuter, C.P.; de Castro Silveira, J.F.; Sehn, A.P.; Todendi, P.F.; de Moura Valim, A.R.; Brazo-Sayavera, J.; de Mello, E.D. The genetic predisposition increases the chances of schoolchildren maintaining higher adiposity levels after three years. BMC Pediatr. 2023, 23, 57. [Google Scholar] [CrossRef]
- Rodrigues, L.D.S.; Santos, A.M.D.; Lima, M.I.S.; Simões, V.M.F.; Pereira, S.R. Association between the FTO gene polymorphism and obesity in Brazilian adolescents from the Northeast region. J. Pediatr. 2020, 96, 630–637. [Google Scholar] [CrossRef]
- Manco, L.; Pinho, S.; Albuquerque, D.; Machado-Rodrigues, A.M.; Padez, C. Physical activity and the association between the FTO rs9939609 polymorphism and obesity in Portuguese children aged 3 to 11 years. Am. J. Hum. Biol. 2019, 31, e23312. [Google Scholar] [CrossRef]
- Brand, C.; Sehn, A.P.; Todendi, P.F.; de Moura Valim, A.R.; Mattevi, V.S.; García-Hermoso, A.; Reis Gaya, A.; Reuter, C.P. The genetic predisposition to obesity has no influence on waist circumference when screen time and sleep duration are adequate in children and adolescents. Eur. J. Sport Sci. 2022, 22, 1757–1764. [Google Scholar] [CrossRef]
- Prats-Puig, A.; Grau-Cabrera, P.; Riera-Pérez, E.; Cortés-Marina, R.; Fortea, E.; Soriano-Rodríguez, P.; de Zegher, F.; Ibánez, L.; Bassols, J.; López-Bermejo, A. Variations in the obesity genes FTO, TMEM18 and NRXN3 influence the vulnerability of children to weight gain induced by short sleep duration. Int. J. Obes. 2013, 37, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Dedoussis, G.V.; Yannakoulia, M.; Timpson, N.J.; Manios, Y.; Kanoni, S.; Scott, R.A.; Papoutsakis, C.; Deloukas, P.; Pitsiladis, Y.P.; Davey-Smith, G.; et al. Does a short breastfeeding period protect from FTO-induced adiposity in children? Int. J. Pediatr. Obes. 2011, 6, e326–e335. [Google Scholar] [CrossRef] [PubMed]
- Quevedo Alves, F.; Reuter, C.P.; Neumann, I.; Todendi, P.F.; Brand, C.; Latosinski Matos, W.; Brazo-Sayavera, J.; Renner, J.D.P.; de Moura Valim, A.R. Relationship between rs9939609 FTO polymorphism with waist circumference and body fat is moderated by ponderal index at birth in youth. Am. J. Hum. Biol. 2022, 34, e23575. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Chen, X.; Cheng, S.; Shu, L.; Yan, M.; Yao, L.; Wang, B.; Huang, S.; Zhou, L.; Yang, Z.; et al. FTO promotes SREBP1c maturation and enhances CIDEC transcription during lipid accumulation in HepG2 cells. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2018, 1863, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Feng, Y.; Zhang, L.; Jia, Y.; Cai, D.; Qian, S.B.; Du, M.; Zhao, R. GR-mediated FTO transactivation induces lipid accumulation in hepatocytes via demethylation of m(6)A on lipogenic mRNAs. RNA Biol. 2020, 17, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Sun, C.; Yan, Y.; Niu, Z.; Li, Y.; Xu, X.; Zhang, J.; Wu, Y.; Li, Y.; Wang, L.; et al. Aberrant elevation of FTO levels promotes liver steatosis by decreasing the m6A methylation and increasing the stability of SREBF1 and ChREBP mRNAs. J. Mol. Cell Biol. 2023, 14, mjac061. [Google Scholar] [CrossRef]
- Wei, D.; Sun, Q.; Li, Y.; Li, C.; Li, X.; Sun, C. Leptin Reduces Plin5 m(6)A Methylation through FTO to Regulate Lipolysis in Piglets. Int. J. Mol. Sci. 2021, 22, 10610. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Sun, B.F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.J.; Ping, X.L.; Chen, Y.S.; Wang, W.J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, J.; Tang, M.; Wang, X.; Fan, N.; Peng, Y. Fat mass and obesity-associated protein promotes liver steatosis by targeting PPARα. Lipids Health Dis. 2022, 21, 29. [Google Scholar] [CrossRef]
- Chen, L.S.; Zhang, M.; Chen, P.; Xiong, X.F.; Liu, P.Q.; Wang, H.B.; Wang, J.J.; Shen, J. The m(6)A demethylase FTO promotes the osteogenesis of mesenchymal stem cells by downregulating PPARG. Acta Pharmacol. Sin. 2022, 43, 1311–1323. [Google Scholar] [CrossRef]
- Zhuang, C.; Zhuang, C.; Luo, X.; Huang, X.; Yao, L.; Li, J.; Li, Y.; Xiong, T.; Ye, J.; Zhang, F.; et al. N6-methyladenosine demethylase FTO suppresses clear cell renal cell carcinoma through a novel FTO-PGC-1α signalling axis. J. Cell. Mol. Med. 2019, 23, 2163–2173. [Google Scholar] [CrossRef]
- Huang, J.; Sun, W.; Wang, Z.; Lv, C.; Zhang, T.; Zhang, D.; Dong, W.; Shao, L.; He, L.; Ji, X.; et al. FTO suppresses glycolysis and growth of papillary thyroid cancer via decreasing stability of APOE mRNA in an N6-methyladenosine-dependent manner. J. Exp. Clin. Cancer Res. 2022, 41, 42. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Chen, Y.; Liu, Y.; Zhuang, L.; Chen, W.; Zeng, B.; Liao, X.; Guo, G.; Wang, Y.; Wang, X. m6A methylation promotes white-to-beige fat transition by facilitating Hif1a translation. EMBO Rep. 2021, 22, e52348. [Google Scholar] [CrossRef] [PubMed]
- Tews, D.; Fischer-Posovszky, P.; Fromme, T.; Klingenspor, M.; Fischer, J.; Rüther, U.; Marienfeld, R.; Barth, T.F.; Möller, P.; Debatin, K.M.; et al. FTO deficiency induces UCP-1 expression and mitochondrial uncoupling in adipocytes. Endocrinology 2013, 154, 3141–3151. [Google Scholar] [CrossRef] [Green Version]
- Li, M.M.; Madara, J.C.; Steger, J.S.; Krashes, M.J.; Balthasar, N.; Campbell, J.N.; Resch, J.M.; Conley, N.J.; Garfield, A.S.; Lowell, B.B. The Paraventricular Hypothalamus Regulates Satiety and Prevents Obesity via Two Genetically Distinct Circuits. Neuron 2019, 102, 653–667.e656. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Simpson, K.A.; Minnion, J.S.; Shillito, J.C.; Bloom, S.R. The role of gut hormones and the hypothalamus in appetite regulation. Endocr. J. 2010, 57, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardle, J.; Llewellyn, C.; Sanderson, S.; Plomin, R. The FTO gene and measured food intake in children. Int. J. Obes. 2009, 33, 42–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert-Diamond, D.; Emond, J.A.; Lansigan, R.K.; Rapuano, K.M.; Kelley, W.M.; Heatherton, T.F.; Sargent, J.D. Television food advertisement exposure and FTO rs9939609 genotype in relation to excess consumption in children. Int. J. Obes. 2017, 41, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Tanofsky-Kraff, M.; Han, J.C.; Anandalingam, K.; Shomaker, L.B.; Columbo, K.M.; Wolkoff, L.E.; Kozlosky, M.; Elliott, C.; Ranzenhofer, L.M.; Roza, C.A.; et al. The FTO gene rs9939609 obesity-risk allele and loss of control over eating. Am. J. Clin. Nutr. 2009, 90, 1483–1488. [Google Scholar] [CrossRef] [Green Version]
- Emond, J.A.; Tovar, A.; Li, Z.; Lansigan, R.K.; Gilbert-Diamond, D. FTO genotype and weight status among preadolescents: Assessing the mediating effects of obesogenic appetitive traits. Appetite 2017, 117, 321–329. [Google Scholar] [CrossRef]
- Obregón Rivas, A.M.; Santos, J.L.; Valladares, M.A.; Cameron, J.; Goldfield, G. Association of the FTO fat mass and obesity-associated gene rs9939609 polymorphism with rewarding value of food and eating behavior in Chilean children. Nutrition 2018, 54, 105–110. [Google Scholar] [CrossRef]
- Rapuano, K.M.; Zieselman, A.L.; Kelley, W.M.; Sargent, J.D.; Heatherton, T.F.; Gilbert-Diamond, D. Genetic risk for obesity predicts nucleus accumbens size and responsivity to real-world food cues. Proc. Natl. Acad. Sci. USA 2017, 114, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Velders, F.P.; De Wit, J.E.; Jansen, P.W.; Jaddoe, V.W.; Hofman, A.; Verhulst, F.C.; Tiemeier, H. FTO at rs9939609, food responsiveness, emotional control and symptoms of ADHD in preschool children. PLoS ONE 2012, 7, e49131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micali, N.; Field, A.E.; Treasure, J.L.; Evans, D.M. Are obesity risk genes associated with binge eating in adolescence? Obesity 2015, 23, 1729–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.; Downer, M.K.; Kilpeläinen, T.O.; Taal, H.R.; Barton, S.J.; Ntalla, I.; Standl, M.; Boraska, V.; Huikari, V.; Kiefte-de Jong, J.C.; et al. Dietary Intake, FTO Genetic Variants, and Adiposity: A Combined Analysis of Over 16,000 Children and Adolescents. Diabetes 2015, 64, 2467–2476. [Google Scholar] [CrossRef] [Green Version]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef] [PubMed]
- Katus, U.; Villa, I.; Ringmets, I.; Vaht, M.; Mäestu, E.; Mäestu, J.; Veidebaum, T.; Harro, J. Association of FTO rs1421085 with obesity, diet, physical activity, and socioeconomic status: A longitudinal birth cohort study. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 948–959. [Google Scholar] [CrossRef]
- Timpson, N.J.; Emmett, P.M.; Frayling, T.M.; Rogers, I.; Hattersley, A.T.; McCarthy, M.I.; Davey Smith, G. The fat mass- and obesity-associated locus and dietary intake in children. Am. J. Clin. Nutr. 2008, 88, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Wiedeman, A.M.; Ngai, Y.F.; Henderson, A.M.; Panagiotopoulos, C.; Devlin, A.M. The FTO rs9939609 Variant Is Associated with Cardiometabolic Disease Risk and Dietary Energy Intakes in Children with Mental Health Disorders. Curr. Dev. Nutr. 2022, 6, nzac014. [Google Scholar] [CrossRef]
- Mehrdad, M.; Doaei, S.; Gholamalizadeh, M.; Fardaei, M.; Fararouei, M.; Eftekhari, M.H. Association of FTO rs9939609 polymorphism with serum leptin, insulin, adiponectin, and lipid profile in overweight adults. Adipocyte 2020, 9, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Jalili, V.; Mokhtari, Z.; Rastgoo, S.; Hajipour, A.; Bourbour, F.; Gholamalizadeh, M.; Mosavi Jarrahi, A.; JavadiKooshesh, S.; Moslem, A.; Abdollahi, M.; et al. The association between FTO rs9939609 polymorphism and serum lipid profile in adult women. Diabetol. Metab. Syndr. 2021, 13, 138. [Google Scholar] [CrossRef]
- Parthasarthy, L.S.; Phadke, N.; Chiplonkar, S.; Khadilkar, A.; Khatod, K.; Ekbote, V.; Shah, S.; Khadilkar, V. Association of Fat Mass and Obesity-associated Gene Variant with Lifestyle Factors and Body Fat in Indian Children. Indian J. Endocrinol. Metab. 2017, 21, 297–301. [Google Scholar] [PubMed]
- Sehn, A.P.; Brand, C.; de Castro Silveira, J.F.; Andersen, L.B.; Gaya, A.R.; Todendi, P.F.; de Moura Valim, A.R.; Reuter, C.P. What is the role of cardiorespiratory fitness and sedentary behavior in relationship between the genetic predisposition to obesity and cardiometabolic risk score? BMC Cardiovasc. Disord. 2022, 22, 92. [Google Scholar] [CrossRef]
- Wang, L.; De Solis, A.J.; Goffer, Y.; Birkenbach, K.E.; Engle, S.E.; Tanis, R.; Levenson, J.M.; Li, X.; Rausch, R.; Purohit, M.; et al. Ciliary gene RPGRIP1L is required for hypothalamic arcuate neuron development. JCI Insight 2019, 4, e123337. [Google Scholar] [CrossRef] [Green Version]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Stratigopoulos, G.; Padilla, S.L.; LeDuc, C.A.; Watson, E.; Hattersley, A.T.; McCarthy, M.I.; Zeltser, L.M.; Chung, W.K.; Leibel, R.L. Regulation of FTO/FTM gene expression in mice and humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1185–R1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jowett, J.B.; Curran, J.E.; Johnson, M.P.; Carless, M.A.; Göring, H.H.; Dyer, T.D.; Cole, S.A.; Comuzzie, A.G.; MacCluer, J.W.; Moses, E.K.; et al. Genetic variation at the FTO locus influences RBL2 gene expression. Diabetes 2010, 59, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Barseem, N.F.; El Ella, S.S.A.; Tawfik, M.A.; El-Nehrawy, R.R. The Potential Implication of FTO rs17817449 Gene Polymorphism on BMI Mediated Risk for Type2 Diabetes Among Obese Egyptian Children and Adolescents. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 697–704. [Google Scholar] [CrossRef] [PubMed]
- González-Herrera, L.; Zavala-Castro, J.; Ayala-Cáceres, C.; Pérez-Mendoza, G.; López-González, M.J.; Pinto-Escalante, D.; Canto-Cetina, T.; García-Escalante, M.G.; Rubi-Castellanos, R.; Contreras-Capetillo, S.; et al. Genetic variation of FTO: rs1421085 T>C, rs8057044 G>A, rs9939609 T>A, and copy number (CNV) in Mexican Mayan school-aged children with obesity/overweight and with normal weight. Am. J. Hum. Biol. 2019, 31, e23192. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Li, G.; Li, L.; Yin, J.; Cheng, H.; Han, L.; Zhang, Q.; Li, N.; Xiao, X.; Grant, S.F.A.; et al. The role of established East Asian obesity-related loci on pediatric leptin levels highlights a neuronal influence on body weight regulation in Chinese children and adolescents: The BCAMS study. Oncotarget 2017, 8, 93593–93607. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ling, J.; Yang, M.; Wang, H.; Zhang, S.; Zhang, X.; Zhu, Y. Rs7206790 and rs11644943 in FTO gene are associated with risk of obesity in Chinese school-age population. PLoS ONE 2014, 9, e108050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Solís, P.; Reyes-Bastidas, M.; Flores, K.; García, O.P.; Rosado, J.L.; Méndez-Villa, L.; Garcia, G.C.; García-Gutiérrez, D.; Kuri-García, A.; Hernández-Montiel, H.L.; et al. Fat mass obesity-associated (FTO) (rs9939609) and melanocortin 4 receptor (MC4R) (rs17782313) SNP are positively associated with obesity and blood pressure in Mexican school-aged children. Br. J. Nutr. 2016, 116, 1834–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wang, K.N.; Tu, L.X.; Zhu, W.F.; Wang, C.L.; Liang, L. FTO polymorphisms associated with metabolic components in Chinese obese children. China Mod. Dr. 2014, 52, 24–27. [Google Scholar]
- Bal, C.; Öztürk, A.; Çiçek, B.; Özdemir, A.; Zararsız, G.; Ünalan, D.; Ertürk Zararsız, G.; Korkmaz, S.; Göksülük, D.; Eldem, V.; et al. The Relationship Between Blood Pressure and Sleep Duration in Turkish Children: A Cross-Sectional Study. J. Clin. Res. Pediatr. Endocrinol. 2018, 10, 51–58. [Google Scholar] [CrossRef]
- Santos, E.; Souza, O.F. Evidence of the Association between Sleep Duration and Blood Pressure in Adolescents: A Systematic Review. Rev. Paul. Pediatr. 2021, 39, e2019225. [Google Scholar] [CrossRef]
- López-Rodríguez, G.; Estrada-Neria, A.; Suárez-Diéguez, T.; Tejero, M.E.; Fernández, J.C.; Galván, M. Common polymorphisms in MC4R and FTO genes are associated with BMI and metabolic indicators in Mexican children: Differences by sex and genetic ancestry. Gene 2020, 754, 144840. [Google Scholar] [CrossRef]
- Inandiklioğlu, N.; Yaşar, A. Association between rs1421085 and rs9939609 Polymorphisms of Fat Mass and Obesity-Associated Gene with High-Density Lipoprotein Cholesterol and Triglyceride in Obese Turkish Children and Adolescents. J. Pediatr. Genet. 2021, 10, 9–15. [Google Scholar] [CrossRef]
- Cao, L.F.; Luo, F.H.; Zhi, D.J.; Cheng, R.Q.; Shen, S.X.; Yang, Y. Association of FTO gene rs9939609, rs1421085 single nucleotide polymorphisms with obesity and metabolic parameters in Chinese Han children and adolescents. Chin. J. Evid.-Based Pediatr. 2010, 5, 46–50. [Google Scholar]
- Duicu, C.; Mărginean, C.O.; Voidăzan, S.; Tripon, F.; Bănescu, C. FTO rs 9939609 SNP Is Associated With Adiponectin and Leptin Levels and the Risk of Obesity in a Cohort of Romanian Children Population. Medicine 2016, 95, e3709. [Google Scholar] [CrossRef]
- Gao, L.; Wu, L.; Zhang, M.; Zhao, X.; Cheng, H.; Mi, J. Gender-specific association of the rs6499640 polymorphism in the FTO gene with plasma lipid levels in Chinese children. Genet. Mol. Biol. 2018, 41, 397–402. [Google Scholar] [CrossRef]
- Wang, L.; Yu, Q.; Xiong, Y.; Liu, L.; Zhang, X.; Zhang, Z.; Wu, J.; Wang, B. Variant rs1421085 in the FTO gene contribute childhood obesity in Chinese children aged 3–6 years. Obes. Res. Clin. Pract. 2013, 7, e14–e22. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y.; Xu, P.; Ye, M.; Liu, Y. Association of the rs9939609 polymorphism of FTO gene with overweight or obesity in Hazakh children. Chin. J. Med. Genet. 2010, 27, 678–681. [Google Scholar]
- do Nascimento, G.A.; Leite, N.; Furtado-Alle, L.; Teixeira, M.D.; de Souza, R.L.R.; Milano, G.E.; da Silva, L.R.; Pizzi, J.; Lopes, W.A.; Lopes, M.F.A.; et al. FTO rs9939609 Does Not Interact with Physical Exercise but Influences Basal Insulin Metabolism in Brazilian Overweight and Obese Adolescents. J. Obes. 2018, 2018, 3134026. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, J.A.; Danielsson, P.; Svensson, V.; Klovins, J.; Gyllensten, U.; Marcus, C.; Schiöth, H.B.; Fredriksson, R. Major gender difference in association of FTO gene variant among severely obese children with obesity and obesity related phenotypes. Biochem. Biophys. Res. Commun. 2008, 368, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Gesteiro, E.; Sánchez-Muniz, F.J.; Ortega-Azorín, C.; Guillén, M.; Corella, D.; Bastida, S. Maternal and neonatal FTO rs9939609 polymorphism affect insulin sensitivity markers and lipoprotein profile at birth in appropriate-for-gestational-age term neonates. J. Physiol. Biochem. 2016, 72, 169–181. [Google Scholar] [CrossRef]
- Cho, Y.; Kim, T.; Lim, S.; Choi, S.; Shin, H.; Lee, H.; Park, K.; Jang, H. Type 2 diabetes-associated genetic variants discovered in the recent genome-wide association studies are related to gestational diabetes mellitus in the Korean population. Diabetologia 2009, 52, 253–261. [Google Scholar] [CrossRef] [Green Version]
- de Melo, S.F.; Frigeri, H.R.; dos Santos-Weiss, I.C.R.; Rea, R.R.; de Souza, E.M.; Alberton, D.; de Moraes Rego, F.G.; Picheth, G. Polymorphisms in FTO and TCF7L2 genes of Euro-Brazilian women with gestational diabetes. Clin. Biochem. 2015, 48, 1064–1067. [Google Scholar] [CrossRef]
- Saucedo, R.; Valencia, J.; Gutierrez, C.; Basurto, L.; Hernandez, M.; Puello, E.; Rico, G.; Vega, G.; Zarate, A. Gene variants in the FTO gene are associated with adiponectin and TNF-alpha levels in gestational diabetes mellitus. Diabetol. Metab. Syndr. 2017, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhang, T.; Liu, Y.; Xu, P.R. The research of association between gene rs9930506 polymorphism and Hazakh children with overweight or obesity in Xinjiang. Chin. J. Prev. Med. 2010, 44, 1106–1110. [Google Scholar]
- Hartiala, O.; Magnussen, C.G.; Kajander, S.; Knuuti, J.; Ukkonen, H.; Saraste, A.; Rinta-Kiikka, I.; Kainulainen, S.; Kähönen, M.; Hutri-Kähönen, N. Adolescence risk factors are predictive of coronary artery calcification at middle age: The cardiovascular risk in young Finns study. J. Am. Coll. Cardiol. 2012, 60, 1364–1370. [Google Scholar] [CrossRef] [Green Version]
- Magnussen, C.G.; Venn, A.; Thomson, R.; Juonala, M.; Srinivasan, S.R.; Viikari, J.S.; Berenson, G.S.; Dwyer, T.; Raitakari, O.T. The association of pediatric low-and high-density lipoprotein cholesterol dyslipidemia classifications and change in dyslipidemia status with carotid intima-media thickness in adulthood: Evidence from the Cardiovascular Risk in Young Finns Study, the Bogalusa Heart Study, and the CDAH (Childhood Determinants of Adult Health) Study. J. Am. Coll. Cardiol. 2009, 53, 860–869. [Google Scholar] [PubMed] [Green Version]
- Dorgan, J.F.; Liu, L.; Barton, B.A.; Deshmukh, S.; Snetselaar, L.G.; Van Horn, L.; Stevens, V.J.; Robson, A.M.; Lasser, N.L.; Himes, J.H. Adolescent diet and metabolic syndrome in young women: Results of the Dietary Intervention Study in Children (DISC) follow-up study. J. Clin. Endocrinol. Metab. 2011, 96, E1999–E2008. [Google Scholar] [CrossRef]
- Elizondo-Montemayor, L.; Gutierrez, N.G.; Moreno, D.M.; Martínez, U.; Tamargo, D.; Treviño, M. School-based individualised lifestyle intervention decreases obesity and the metabolic syndrome in Mexican children. J. Hum. Nutr. Diet. 2013, 26 (Suppl. 1), 82–89. [Google Scholar] [CrossRef] [PubMed]
- García Flores, S.A.; Ninatanta-Ortiz, J.A.; Abanto Villar, M.V.; Pérez Cieza, K.M.; Chávez Farro, R.R.; Palacios Sánchez, S.E.; Romaní Romaní, F.R. Lifestyle school-based intervention to increase the proportion of adolescents free of components of the metabolic syndrome in an andean region of Peru. Rev. Peru. Med. Exp. Salud Pública 2022, 39, 36–46. [Google Scholar] [CrossRef]
- Rabbani, B.; Chiti, H.; Sharifi, F.; Mazloomzadeh, S. Effect of lifestyle modification for two years on obesity and metabolic syndrome components in elementary students: A community- based trial. Casp. J. Intern. Med. 2022, 13, 555–566. [Google Scholar]
- Cifuentes, L.; Ghusn, W.; Feris, F.; Campos, A.; Sacoto, D.; De la Rosa, A.; McRae, A.; Rieck, T.; Mansfield, S.; Ewoldt, J.; et al. Phenotype tailored lifestyle intervention on weight loss and cardiometabolic risk factors in adults with obesity: A single-centre, non-randomised, proof-of-concept study. EClinicalMedicine 2023, 58, 101923. [Google Scholar] [CrossRef]
- Foster, B.A.; Farragher, J.; Parker, P.; Sosa, E.T. Treatment Interventions for Early Childhood Obesity: A Systematic Review. Acad. Pediatr. 2015, 15, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, E.; Atkinson, G.; Richter, B.; Metzendorf, M.I.; Baur, L.; Finer, N.; Corpeleijn, E.; O’Malley, C.; Ells, L.J. Drug interventions for the treatment of obesity in children and adolescents. Cochrane Database Syst. Rev. 2016, 11, Cd012436. [Google Scholar]
- Torbahn, G.; Brauchmann, J.; Axon, E.; Clare, K.; Metzendorf, M.I.; Wiegand, S.; Pratt, J.S.; Ells, L.J. Surgery for the treatment of obesity in children and adolescents. Cochrane Database Syst. Rev. 2022, 9, Cd011740. [Google Scholar]
- Mühlig, Y.; Wabitsch, M.; Moss, A.; Hebebrand, J. Weight loss in children and adolescents. Dtsch. Arztebl. Int. 2014, 111, 818–824. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| MetS Components | IDF Criteria | NCEP ATP III Criteria | ||
|---|---|---|---|---|
| 6–9 Years | 10–15 Years | ≥16 Years | ND | |
| Abdominal obesity | WC ≥ 90th percentile for age [15] | WC ≥ 90th percentile [16] | WC ≥ 94 cm in boys and ≥ 80 cm in girls [15] or WC ≥ 90 cm in boys and ≥ 80 cm in girls [17] | WC ≥ 102 cm in boys and ≥ 88 cm in girls [16] or WC ≥ 75th percentile for age and sex [17] |
| Hypertension | ND | SBP ≥ 130 or DBP ≥ 85 mmHg [17] | SBP ≥ 130 or DBP ≥ 85 mmHg [17] | SBP ≥ 130 mmHg [16] or SBP ≥ 90th percentile for age and sex [17] |
| Hypertriglyceridemia | ND | TG ≥ 1.7 mmol/L [16] | TG ≥ 1.7 mmol/L [17] | TG ≥ 1.7 mmol/L [17] |
| Hypo-high-density lipoproteinemia | ND | HDL-C ≤ 1.03 mmol/L [16] | HDL-C ≤ 1.03 mmol/L in boys and ≤ 1.29 mmol/L in girls [17] | HDL-C ≤ 1.03 mmol/L [16] |
| Hyperglycemia | ND | FBG ≥ 5.6 mmol/L [16] | FBG ≥ 5.6 mmol/L [17] | FBG ≥ 6.1 mmol/L [16] |
| Diagnostic criteria | Abdominal obesity and any two of the other components must be present | At least three components are present | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Wade, H.; Zhang, B.; Xu, W.; Wu, R.; Li, S.; Su, Q. Polymorphisms of Fat Mass and Obesity-Associated Gene in the Pathogenesis of Child and Adolescent Metabolic Syndrome. Nutrients 2023, 15, 2643. https://doi.org/10.3390/nu15122643
Song Y, Wade H, Zhang B, Xu W, Wu R, Li S, Su Q. Polymorphisms of Fat Mass and Obesity-Associated Gene in the Pathogenesis of Child and Adolescent Metabolic Syndrome. Nutrients. 2023; 15(12):2643. https://doi.org/10.3390/nu15122643
Chicago/Turabian StyleSong, Yongyan, Henry Wade, Bingrui Zhang, Wenhao Xu, Rongxue Wu, Shujin Li, and Qiaozhu Su. 2023. "Polymorphisms of Fat Mass and Obesity-Associated Gene in the Pathogenesis of Child and Adolescent Metabolic Syndrome" Nutrients 15, no. 12: 2643. https://doi.org/10.3390/nu15122643