
Magnesium Improves Cardiac Function in Experimental Uremia by Altering Cardiac Elastin Protein Content
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
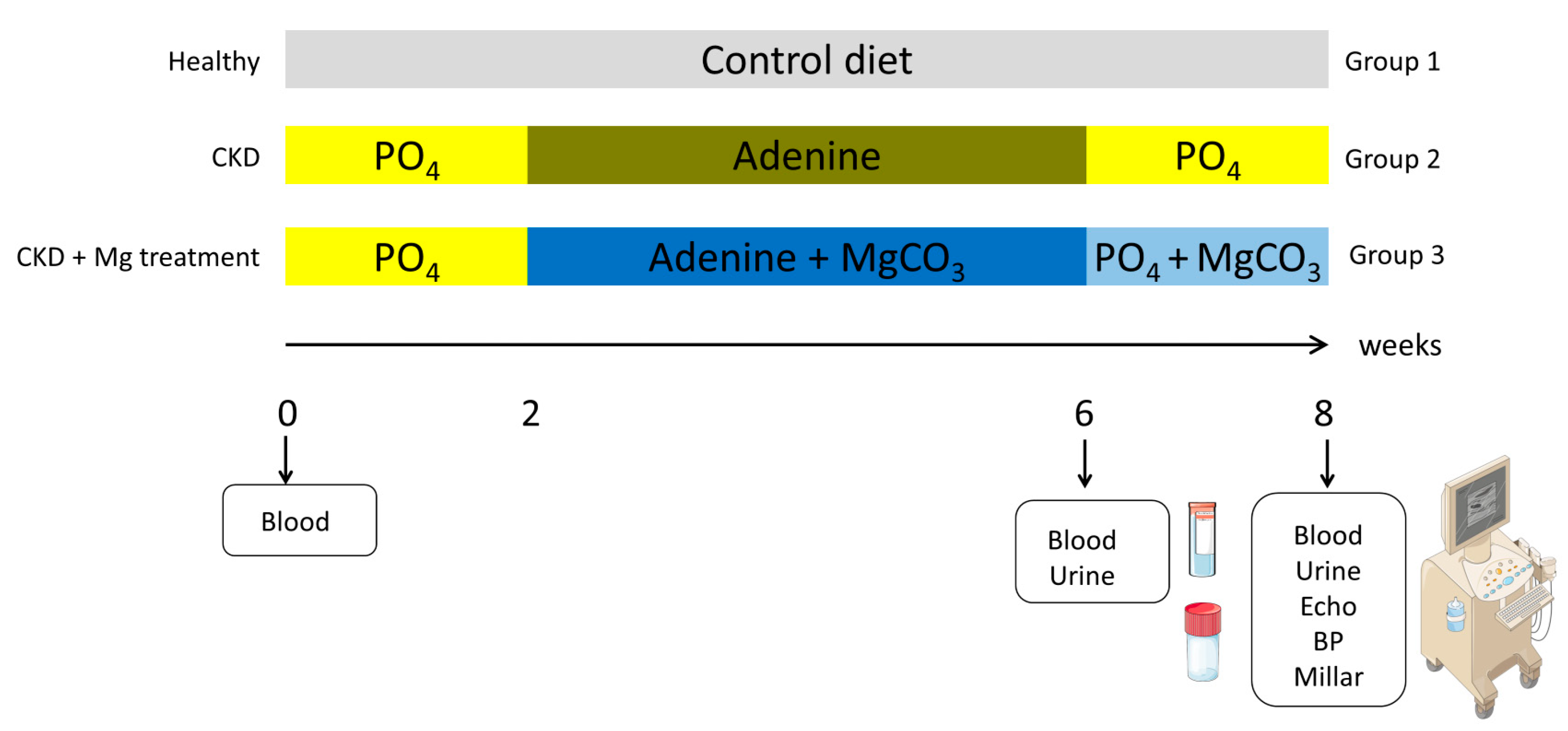
2.1. Animals
2.2. Functional Measurements
2.3. Biochemistry
2.4. RT-PCR
2.5. Tissue Stainings
2.6. Tissue Assays
2.7. Statistics
3. Results
3.1. Adenine-Receiving Rats Maintained Their Body Weights
3.2. Successful Induction of CKD by Adenine Diet
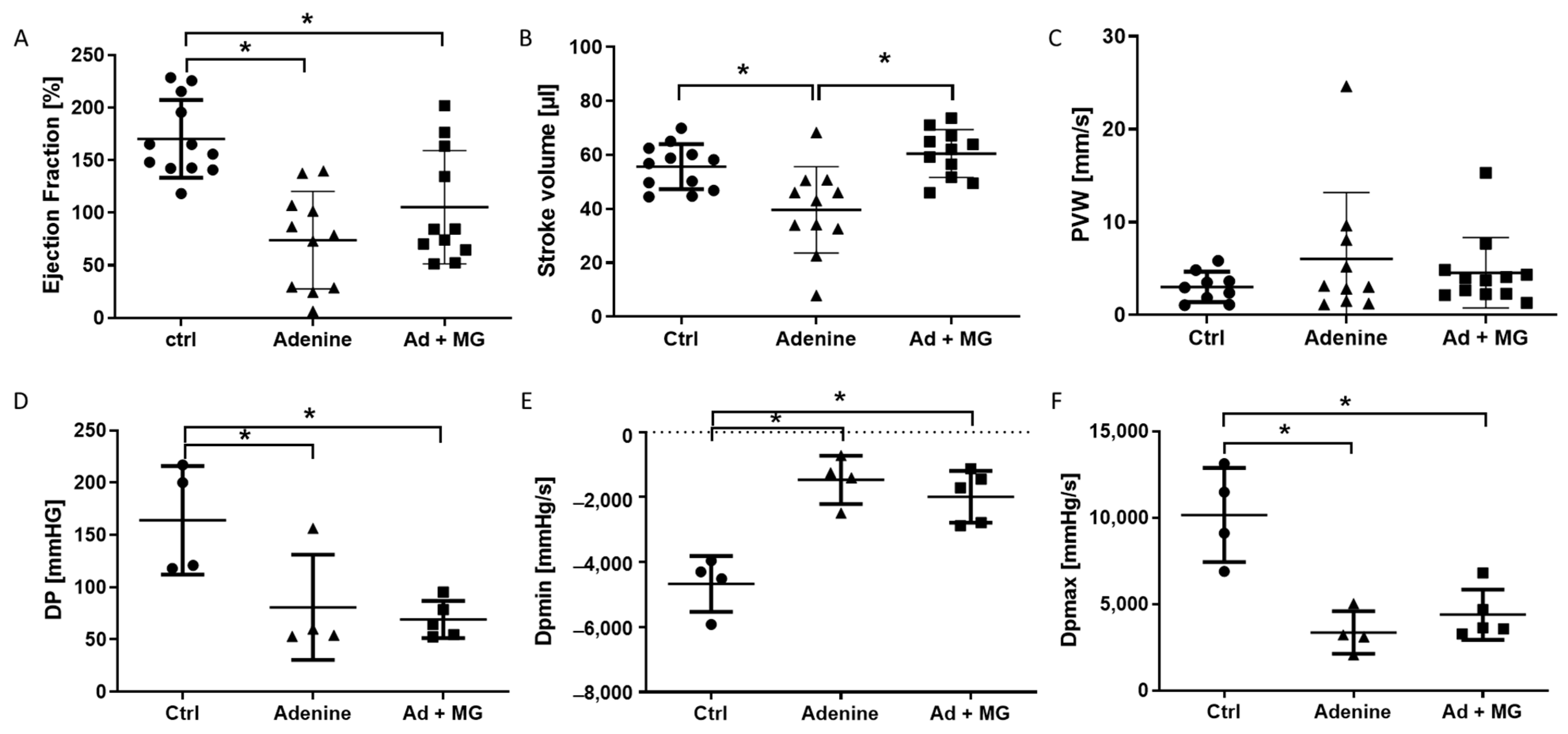
3.3. Preserved Left Ventricular Function by Magnesium
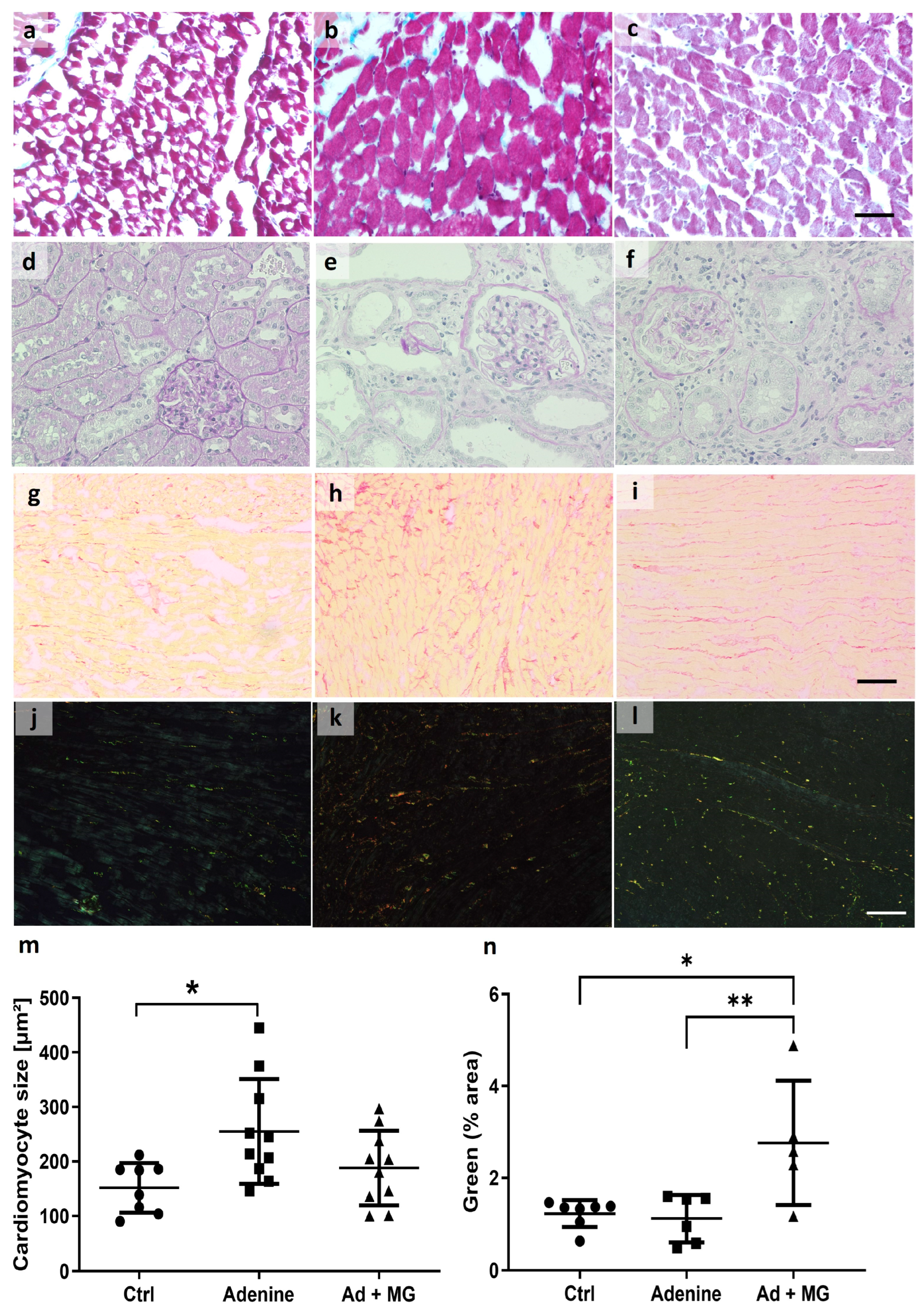
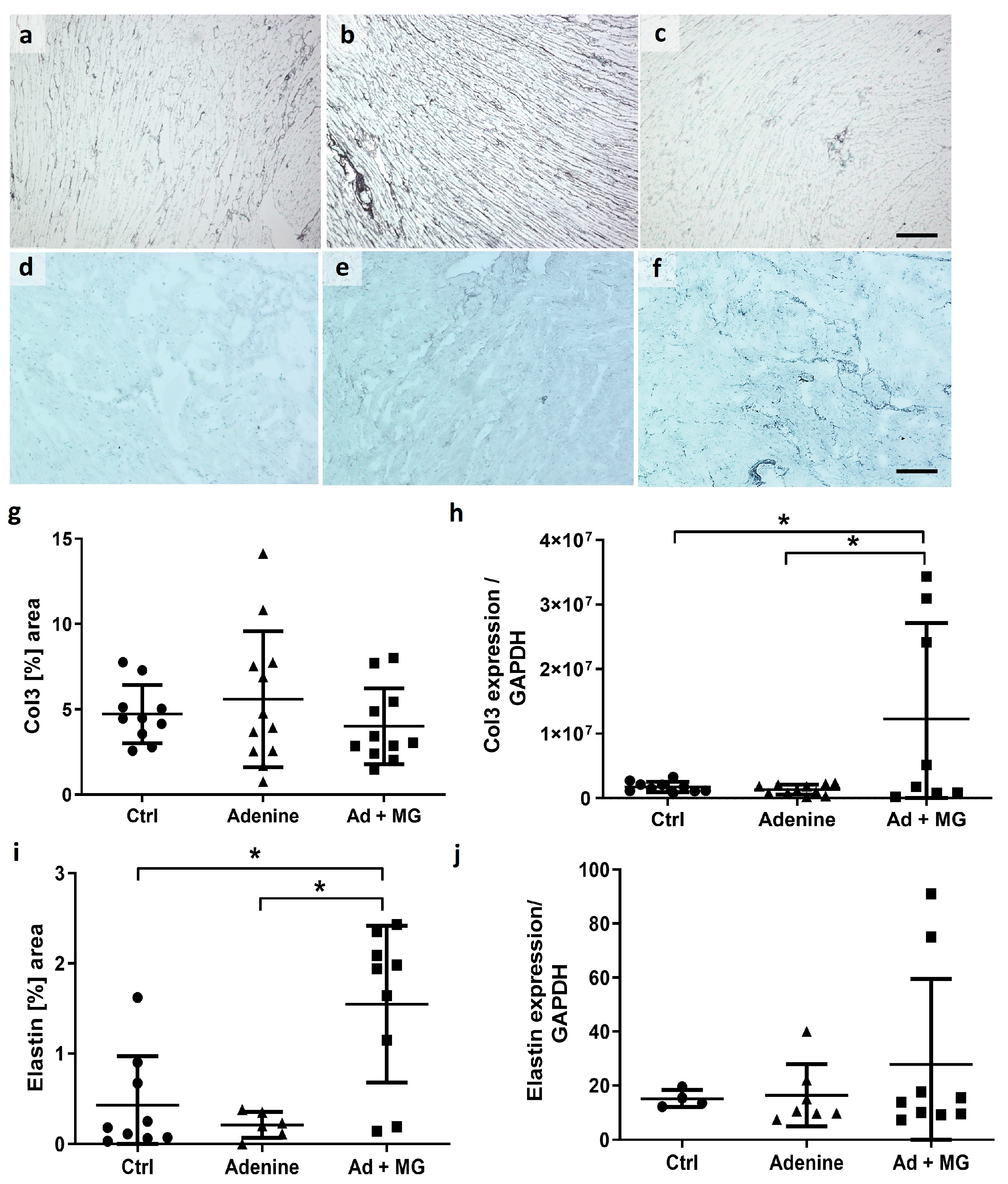
3.4. Magnesium Increased Elastin Protein and Limited Cardiomyocyte Hypertrophy, in the Absence of Fibrosis
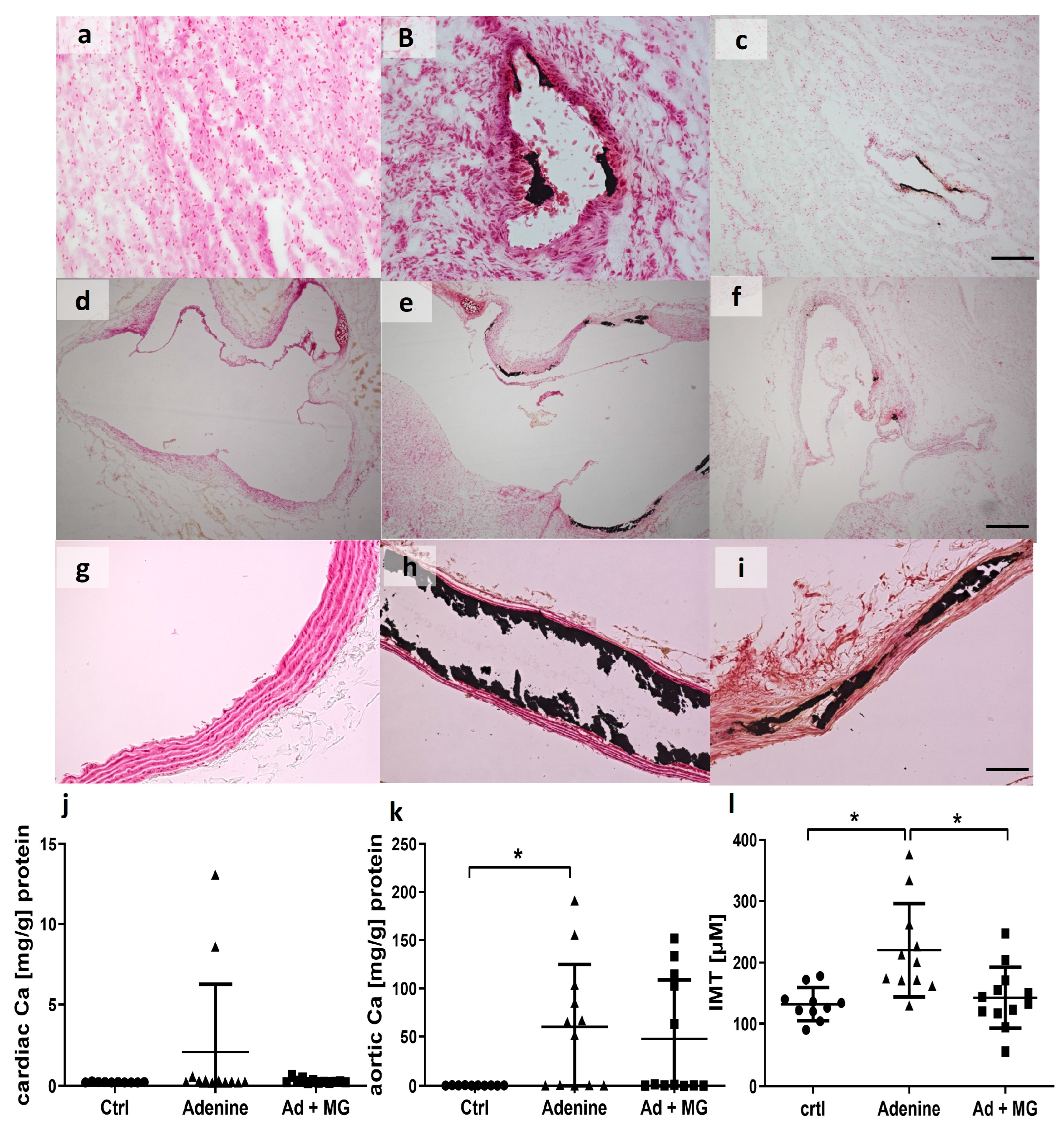
3.5. Adenine Diet Induced Vascular Calcification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azem, R.; Daou, R.; Bassil, E.; Anvari, E.M.; Taliercio, J.J.; Arrigain, S.; Schold, J.D.; Vachharajani, T.; Nally, J.; Na Khoul, G.N. Serum magnesium, mortality and disease progression in chronic kidney disease. BMC Nephrol. 2020, 21, 49. [Google Scholar] [CrossRef] [Green Version]
- Reffelmann, T.; Ittermann, T.; Dorr, M.; Volzke, H.; Reinthaler, M.; Petersmann, A.; Felix, S.B. Low serum magnesium concentrations predict cardiovascular and all-cause mortality. Atherosclerosis 2011, 219, 280–284. [Google Scholar] [CrossRef]
- Bouida, W.; Beltaief, K.; Msolli, M.A.; Azaiez, N.; Ben Soltane, H.; Sekma, A.; Trabelsi, I.; Boubaker, H.; Grissa, M.H.; Methemem, M.; et al. Low-dose Magnesium Sulfate Versus High Dose in the Early Management of Rapid Atrial Fibrillation: Randomized Controlled Double-blind Study (LOMAGHI Study). Acad. Emerg. Med. 2019, 26, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.J.; Teubner, D. A randomized controlled trial of magnesium sulfate, in addition to usual care, for rate control in atrial fibrillation. Ann. Emerg. Med. 2005, 45, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, A. Efficacy of class III antiarrhythmics and magnesium combination therapy for atrial fibrillation. Pharm. Pract. 2012, 10, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salaminia, S.; Sayehmiri, F.; Angha, P.; Sayehmiri, K.; Motedayen, M. Evaluating the effect of magnesium supplementation and cardiac arrhythmias after acute coronary syndrome: A systematic review and meta-analysis. BMC Cardiovasc. Disord. 2018, 18, 129. [Google Scholar] [CrossRef]
- Pokan, R.; Hofmann, P.; von Duvillard, S.P.; Smekal, G.; Wonisch, M.; Lettner, K.; Schmid, P.; Shechter, M.; Silver, B.; Bachl, N. Oral magnesium therapy, exercise heart rate, exercise tolerance, and myocardial function in coronary artery disease patients. Br. J. Sports Med. 2006, 40, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Almoznino-Sarafian, D.; Berman, S.; Mor, A.; Shteinshnaider, M.; Gorelik, O.; Tzur, I.; Alon, I.; Modai, D.; Cohen, N. Magnesium and C-reactive protein in heart failure: An anti-inflammatory effect of magnesium administration? Eur. J. Nutr. 2007, 46, 230–237. [Google Scholar] [CrossRef]
- Cunha, A.R.; D’El-Rei, J.; Medeiros, F.; Umbelino, B.; Oigman, W.; Touyz, R.M.; Neves, M.F. Oral magnesium supplementation improves endothelial function and attenuates subclinical atherosclerosis in thiazide-treated hypertensive women. J. Hypertens. 2017, 35, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Mathers, T.W.; Beckstrand, R.L. Oral magnesium supplementation in adults with coronary heart disease or coronary heart disease risk. J. Am. Acad. Nurse Pract. 2009, 21, 651–657. [Google Scholar] [CrossRef]
- Liu, M.; Dudley, S.C., Jr. Magnesium, Oxidative Stress, Inflammation, and Cardiovascular Disease. Antioxidants 2020, 9, 907. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.L. Treating arrhythmias with adjunctive magnesium: Identifying future research directions. Eur. Heart J. Cardiovasc. Pharmacother. 2017, 3, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Floege, J. Magnesium in CKD: More than a calcification inhibitor? J. Nephrol. 2015, 28, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Lioufas, N.M.; Pascoe, E.M.; Hawley, C.M.; Elder, G.J.; Badve, S.V.; Block, G.A.; Johnson, D.W.; Toussaint, N.D. Systematic Review and Meta-Analyses of the Effects of Phosphate-Lowering Agents in Nondialysis CKD. J. Am. Soc. Nephrol. 2022, 33, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Ruospo, M.; Palmer, S.C.; Natale, P.; Craig, J.C.; Vecchio, M.; Elder, G.J.; Strippoli, G.F. Phosphate binders for preventing and treating chronic kidney disease-mineral and bone disorder (CKD-MBD). Cochrane Database Syst. Rev. 2018, 8, CD006023. [Google Scholar] [CrossRef]
- Doshi, S.M.; Wish, J.B. Past, Present, and Future of Phosphate Management. Kidney Int. Rep. 2022, 7, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Leenders, N.H.J.; Vervloet, M.G. Magnesium: A Magic Bullet for Cardiovascular Disease in Chronic Kidney Disease? Nutrients 2019, 11, 455. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, Y. The emerging role of magnesium in CKD. Clin. Exp. Nephrol. 2022, 26, 379–384. [Google Scholar] [CrossRef]
- Diwan, V.; Brown, L.; Gobe, G.C. Adenine-induced chronic kidney disease in rats. Nephrology (Carlton) 2018, 23, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Klinkhammer, B.M.; Djudjaj, S.; Kunter, U.; Palsson, R.; Edvardsson, V.O.; Wiech, T.; Thorsteinsdottir, M.; Hardarson, S.; Foresto-Neto, O.; Mulay, S.R.; et al. Cellular and Molecular Mechanisms of Kidney Injury in 2,8-Dihydroxyadenine Nephropathy. J. Am. Soc. Nephrol. 2020, 31, 799–816. [Google Scholar] [CrossRef] [PubMed]
- Kaesler, N.; Magdeleyns, E.; Herfs, M.; Schettgen, T.; Brandenburg, V.; Fliser, D.; Vermeer, C.; Floege, J.; Schlieper, G.; Kruger, T. Impaired vitamin K recycling in uremia is rescued by vitamin K supplementation. Kidney Int. 2014, 86, 286–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, T.; Chen, J.; Fayad, Z.A.; Fuster, V.; Hajjar, R.J.; Chemaly, E.R. Comparison of echocardiographic measurements of left ventricular volumes to full volume magnetic resonance imaging in normal and diseased rats. J. Am. Soc. Echocardiogr. 2013, 26, 910–918. [Google Scholar] [CrossRef] [Green Version]
- Daugherty, A.; Rateri, D.; Hong, L.; Balakrishnan, A. Measuring blood pressure in mice using volume pressure recording, a tail-cuff method. J. Vis. Exp. 2009, 27, 1291. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Nagayama, T.; Mukhopadhyay, P.; Batkai, S.; Kass, D.A. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat. Protoc. 2008, 3, 1422–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Vlekkert, D.; Machado, E.; d’Azzo, A. Analysis of Generalized Fibrosis in Mouse Tissue Sections with Masson's Trichrome Staining. Bio. Protoc. 2020, 10, e3629. [Google Scholar] [CrossRef]
- Lattouf, R.; Younes, R.; Lutomski, D.; Naaman, N.; Godeau, G.; Senni, K.; Changotade, S. Picrosirius red staining: A useful tool to appraise collagen networks in normal and pathological tissues. J. Histochem. Cytochem. 2014, 62, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Kaesler, N.; Goettsch, C.; Weis, D.; Schurgers, L.; Hellmann, B.; Floege, J.; Kramann, R. Magnesium but not nicotinamide prevents vascular calcification in experimental uraemia. Nephrol. Dial. Transplant. 2020, 35, 65–73. [Google Scholar] [CrossRef]
- Morin, L.G. Direct colorimetric determination of serum calcium with o-cresolphthalein complexon. Am. J. Clin. Pathol. 1974, 61, 114–117. [Google Scholar] [CrossRef]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. Methods Mol. Biol. 1994, 32, 5–8. [Google Scholar] [CrossRef]
- Reddy, G.K.; Enwemeka, C.S. A simplified method for the analysis of hydroxyproline in biological tissues. Clin. Biochem. 1996, 29, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Saritas, T.; Kjolby, M.; Hermann, J.; Speer, T.; Himmelsbach, A.; Mahr, K.; Heuschkel, M.A.; Schunk, S.J.; Thirup, S.; et al. Carbamylated sortilin associates with cardiovascular calcification in patients with chronic kidney disease. Kidney Int. 2022, 101, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Kooman, J.P.; Kotanko, P.; Schols, A.M.; Shiels, P.G.; Stenvinkel, P. Chronic kidney disease and premature ageing. Nat. Rev. Nephrol. 2014, 10, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Ebert, T.; Pawelzik, S.C.; Witasp, A.; Arefin, S.; Hobson, S.; Kublickiene, K.; Shiels, P.G.; Back, M.; Stenvinkel, P. Inflammation and Premature Ageing in Chronic Kidney Disease. Toxins 2020, 12, 227. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.; Floege, J.; Fliser, D.; Bohm, M.; Marx, N. Cardiovascular Disease in Chronic Kidney Disease Pathophysiological Insights and Therapeutic Options. Circulation 2021, 143, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Cocciolone, A.J.; Hawes, J.Z.; Staiculescu, M.C.; Johnson, E.O.; Murshed, M.; Wagenseil, J.E. Elastin, arterial mechanics, and cardiovascular disease. Am J. Physiol. Heart Circ. Physiol. 2018, 315, H189–H205. [Google Scholar] [CrossRef] [Green Version]
- Novitskaya, T.; Nishat, S.; Covarrubias, R.; Wheeler, D.G.; Chepurko, E.; Bermeo-Blanco, O.; Xu, Z.; Baer, B.; He, H.; Moore, S.N.; et al. Ectonucleoside triphosphate diphosphohydrolase-1 (CD39) impacts TGF-beta1 responses: Insights into cardiac fibrosis and function following myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2022, 323, H1244–H1261. [Google Scholar] [CrossRef]
- Muller, W.; Iffland, R.; Firsching, R. Relationship between magnesium and elastic fibres. Magnes. Res. 1993, 6, 215–222. [Google Scholar]
- Muller, W.; Firsching, R. Demonstration of elastic fibres with reagents for detection of magnesium. J. Anat. 1991, 175, 195–202. [Google Scholar]
- Basalyga, D.M.; Simionescu, D.T.; Xiong, W.; Baxter, B.T.; Starcher, B.C.; Vyavahare, N.R. Elastin degradation and calcification in an abdominal aorta injury model: Role of matrix metalloproteinases. Circulation 2004, 110, 3480–3487. [Google Scholar] [CrossRef]
- Smith, E.R.; Tomlinson, L.A.; Ford, M.L.; McMahon, L.P.; Rajkumar, C.; Holt, S.G. Elastin degradation is associated with progressive aortic stiffening and all-cause mortality in predialysis chronic kidney disease. Hypertension 2012, 59, 973–978. [Google Scholar] [CrossRef]
- Junqueira, L.C.; Montes, G.S.; Sanchez, E.M. The influence of tissue section thickness on the study of collagen by the Picrosirius-polarization method. Histochemistry 1982, 74, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Debessa, C.R.G.; Maifrino, L.B.M.; de Souza, R.R. Age related changes of the collagen network of the human heart. Mech. Ageing Dev. 2001, 122, 1049–1058. [Google Scholar] [CrossRef]
- Uchinaka, A.; Yoshida, M.; Tanaka, K.; Hamada, Y.; Mori, S.; Maeno, Y.; Miyagawa, S.; Sawa, Y.; Nagata, K.; Yamamoto, H.; et al. Overexpression of collagen type III in injured myocardium prevents cardiac systolic dysfunction by changing the balance of collagen distribution. J. Thorac. Cardiovasc. Surg. 2018, 156, 217–226.e213. [Google Scholar] [CrossRef]
- Kaesler, N.; Babler, A.; Floege, J.; Kramann, R. Cardiac Remodeling in Chronic Kidney Disease. Toxins 2020, 12, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balafa, O.; Dounousi, E.; Giannikouris, I.; Petrakis, I.; Georgoulidou, A.; Karassavidou, D.; Kokalis, A.; Stauroulopoulos, A.; Theodoridis, M.; Oikonomidis, I.; et al. Lower serum magnesium is a predictor of left ventricular hypertrophy in patients on dialysis. Int. Urol. Nephrol. 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Ma, D.; Zhang, J.; Zhang, Y.; Zhang, X.; Han, X.; Song, T.; Zhang, Y.; Chu, L. Inhibition of myocardial hypertrophy by magnesium isoglycyrrhizinate through the TLR4/NF-kappaB signaling pathway in mice. Int. Immunopharmacol. 2018, 55, 237–244. [Google Scholar] [CrossRef]
- Cunningham, J.; Rodriguez, M.; Messa, P. Magnesium in chronic kidney disease Stages 3 and 4 and in dialysis patients. Clin. Kidney J. 2012, 5, i39–i51. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Mohammadi, F.; Behmanesh, F.; Samani, S.M.; Borzouee, M.; Amoozgar, H.; Haghpanah, S. Effect of combination therapy of hydroxyurea with l-carnitine and magnesium chloride on hematologic parameters and cardiac function of patients with beta-thalassemia intermedia. Eur. J. Haematol. 2010, 84, 52–58. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Adenine | Adenine +MgCO3 | |
|---|---|---|---|
| Creatinine, mg/dL, median (IQR) | 0.4 (0.32–0.49) | 1.7 (1.3–5.1) * | 1.9 (1.1–5.4) * |
| Urea, mg/dL, median (IQR) | 1.5 (13.2–18.5) | 66.1 (51.8–212.9) * | 49.6 (35.6–172.5) * |
| Phosphate, mg/dL, median (IQR) | 6.6 (6.2–8.1) | 15.4 (10.6–23.8) * | 10.9 (8.3–23.8) * |
| Calcium, mg/dL, median (IQR) | 11.4 (11.1–11.7) | 11.8 (11–12.7) | 11.7 (11.2–12) |
| Ca × P, mg2/dL2, median (IQR) | 74 (70.6–97.3) | 191.8 (121–260.9) * | 127.7 (91.1–300.5) * |
| Magnesium, mg/dL, median (IQR) | 2.9 (2.7–2.9) | 2.9 (2.7–3.4) | 2.9 (2.7–4.8) |
| ALP, UI/L, mean (SD) | 91.7 ± 16.4 | 97.54 ± 34.72 | 107.17 ± 43.25 |
| PTH, pg/mL, mean (SD) | 1700 ± 918 | 5338 ± 4525 * | 4133 ± 1396 * |
| FGF23, ng/mL, median (IQR) | 0.34 (0.28–0.41) | 9.84 (2.72–30.95) * | 24.1 (1.15–286.75) * |
| Protein, g/dL, median (IQR) | 5.8 (5.6–6.02) | 5.4 (5–5.8) * | 5.2 (5–5.5) * |
| Albumin, g/dL, median (IQR) | 2.9 (2.8–3.2) | 2.6 (2.4–2.8) * | 2.5 (2.3–2.8) * |
| Control | Adenine | Adenine + MgCO3 | |
|---|---|---|---|
| Phosphate mg/24 h, mean (SD) | 12.89 ± 4.15 | 48.10 ± 12.65 * | 43.03 ± 12.47 * |
| FEP %, median (IQR) | 5.1 (4.6–7.8) | 67 (60.6–80.7) * | 69.4 (52.3–85.5) * |
| Magnesium mg/24 h, median (IQR) | 2.9 (2.1–4.5) | 3.3 (2.5–4.7) | 4.36 (2.6–5.9) |
| FEMg %, median (IQR) | 13.2 (3.1–30.9) | 16.9 (3.5–44.6) | 28.6 (21.9–53.9) |
| Calcium mg/24 h, mean (SD) | 2.62 ± 2.07 | 3.16 ± 1.75 | 5.23 ± 2.32 |
| FECa %, median (IQR) | 2 (0.35–10.2) | 3.27 (0.29–13.3) | 8.5 (6.1–24) |
| Protein mg/24 h, median (IQR) | 12.1 (10.3–16) | 11.4 (9.3–16.1) | 12.9 (9.7–18.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, X.; Friesen, X.; Mathias Brandenburg, V.; Liehn, E.A.; Steppan, S.; Kiessling, F.; Kramann, R.; Floege, J.; Krüger, T.; Kaesler, N. Magnesium Improves Cardiac Function in Experimental Uremia by Altering Cardiac Elastin Protein Content. Nutrients 2023, 15, 1303. https://doi.org/10.3390/nu15061303
Barros X, Friesen X, Mathias Brandenburg V, Liehn EA, Steppan S, Kiessling F, Kramann R, Floege J, Krüger T, Kaesler N. Magnesium Improves Cardiac Function in Experimental Uremia by Altering Cardiac Elastin Protein Content. Nutrients. 2023; 15(6):1303. https://doi.org/10.3390/nu15061303
Chicago/Turabian StyleBarros, Xoana, Xenia Friesen, Vincent Mathias Brandenburg, Elisa Anamaria Liehn, Sonja Steppan, Fabian Kiessling, Rafael Kramann, Jürgen Floege, Thilo Krüger, and Nadine Kaesler. 2023. "Magnesium Improves Cardiac Function in Experimental Uremia by Altering Cardiac Elastin Protein Content" Nutrients 15, no. 6: 1303. https://doi.org/10.3390/nu15061303