
A Review on Polyphenols in Salicornia ramosissima with Special Emphasis on Their Beneficial Effects on Brain Ischemia

1
Institute of Biomedicine of Seville (IBiS), Hospital Universitario Virgen del Rocío, CSIC, Universidad de Sevilla, 41013 Seville, Spain
2
Department of Neurology, Hospital Universitario Virgen Macarena, 41009 Seville, Spain
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2023, 15(3), 793; https://doi.org/10.3390/nu15030793
Submission received: 26 December 2022
/
Revised: 29 January 2023
/
Accepted: 1 February 2023
/
Published: 3 February 2023
(This article belongs to the Special Issue Neuroprotection with Bioactive Compounds)
Abstract
:There has been an increasing interest in the consumption of halophytes as a healthy food in the last few years. Salicornia ramosissima is a seasonal Mediterranean halophyte with an interesting profile of bioactive compounds, including more than 60 identified polyphenols with a broad range of biological activities. Accumulating evidence supports the role of dietary polyphenols in the prevention of cardiovascular diseases, such as stroke. Stroke is the second cause of death worldwide and it is estimated that a substantial proportion of stroke incidence and recurrence may be prevented by healthier dietary patterns. Here, we have grouped the phenolic acids and flavonoids identified in S. ramosissima and reviewed their potential protective effect on brain ischemia, which are mostly related to the reduction of oxidative stress and inflammation, the inhibition of cell death pathways and their role in the preservation of the vascular function. Despite the fact that most of these compounds have been reported to be neuroprotective through multiple mechanisms, human studies are still scarce. Given the safe profile of polyphenols identified in S. ramosissima, this halophyte plant could be considered as a source of bioactive compounds for the nutraceutical industry.
1. Introduction
Halophytes comprise more than 350 annual and perennial species capable of tolerating the severe environmental condition of high salinity. In recent years, these plants have aroused interest as a valuable environmental resource for their ability to generate biomass in drought and salinity and a high resistance to conventional plant diseases. Moreover, halophytes contribute to carbon stabilization and play a role in soil phyto-desalinization and phytoremediation [1]. In addition to these benefits, halophytes have been attributed with health-promoting effects, including anti-inflammatory and antioxidant activities [2].
Salicornia ramosissima, also known as glasswort, is an annual halophyte of the S. europaea agg that can be found on the coastline of Europe, including Portugal and Spain, where its aerial parts are frequently consumed as a fresh vegetable [3]. More recently, the use of halophytes has been suggested as an alternative natural solution to reduce the sodium content of food products that are traditionally produced using salt [4]. S. ramosissima has a good nutritional profile, being a source of mineral and fiber [5], but also bioactive compounds. synthesized in response to abiotic stress due to high salinity and UV radiation, such as polyphenols, dietary consumption of which is associated with many health benefits.
Improving nutritional lifestyle is a major strategy in controlling modifiable risk factors related to cardiovascular and cerebrovascular diseases. Stroke, which is the most common neurovascular disease, represents the second cause of death and the third cause of disability worldwide according to the World Health Organization. Hemorrhagic stroke is caused by bleeding blood vessels and represents about 15% of all stroke cases. Ischemic stroke, which is the most common type, is caused by the occlusion of a blood vessel by a thrombus, resulting in the local lack of oxygen and nutrients leading to brain cell death at the infarcted area [6]. Given the high demand for oxygen and glucose of the tissue, the local disruption of blood flow to a brain area leads to cell death by necrosis within minutes (ischemic core). This event is followed by the disruption of normal cell function in the surrounding area (penumbra), triggered by energetic failure, inflammation, acidosis, excitotoxicity, release of Reactive Oxygen Species (ROS) and the impairment of the blood–brain barrier (BBB), among other mechanisms [7].
Reperfusion therapies using thrombolytics or mechanical thrombectomy are the current available treatments for ischemic stroke [8]. Although reperfusion is certainly the therapeutic objective, it can also induce additional injury by triggering inflammation and oxidative stress [9]. Two major categories of experimental models of brain ischemia are used in the search for new therapies for stroke in a variety of animal species including rats and mice, namely global and focal ischemia. The most common method used for focal ischemia is middle cerebral artery occlusion (MCAO), which can be permanent or transient, in order to allow reperfusion. In global ischemia, multiple cervical vessels are temporarily occluded [10].
It has been shown that a substantial proportion of strokes can be attributed to unhealthy lifestyle behaviors [11] and up to 80% of stroke recurrence might be prevented by the application of a multifactorial approach that includes dietary modification [12]. Recently, our group reported that diet supplementation with S. ramosissima ethanolic extract with high content of polyphenols protected both flies and mice from the deleterious effects of ischemia [13]. Here, we aim to review bioactive compounds identified in the edible plant S. ramosissima and discuss their potential as therapeutic nutritional interventions for stroke therapy using the available bibliography on experimental models of brain ischemia.
2. Natural Bioactive Compounds Found in S. ramosissima
Saline stress response in Salicornia species comprises major events in plant tissues including osmotic, anatomical and physiological adaptations, as well as metabolic changes that involve the production of secondary metabolites [14]. On the basis of their structure and chemical nature, there are three main groups of secondary metabolites biosynthesized by plants: (i) terpenes, (ii) phenolics and (iii) sulfur and nitrogen-containing compounds (glucosinolates and alkaloids, respectively) [15].
Phenolics are a broad group of secondary metabolites that range from an aromatic ring (bearing one or more hydroxyl substituents) to more complex examples. Plant phenolics are biosynthesized through the shikimic acid and phenylpropanoid metabolism pathways [16,17]. The biosynthesis of phenolic compounds in plants begins with the conversion of glucose to glucose-6-phosphate to produce either phosphoenolpyruvate (PEP) by glycolysis or erythrose-4-phosphate by the pentose phosphate pathway (PPP). PEP and erythrose-4-phosphate are used together to produce phenylalanine and tyrosine by the shikimic acid pathway, which involves seven sequential enzymatic steps. Phenylalanine and tyrosine are then channeled into the phenylpropanoid pathway to generate several phenolic compounds by means of five rate-limiting enzymes [15,17,18].
Among the compounds of interest in Salicornia species are polyphenols, characterized by the presence of phenol rings in their structure, ranging from simple molecules to complex polymers. Polyphenol subgroups include phenolic acids, flavonoids, lignans and stilbenes [14,17].
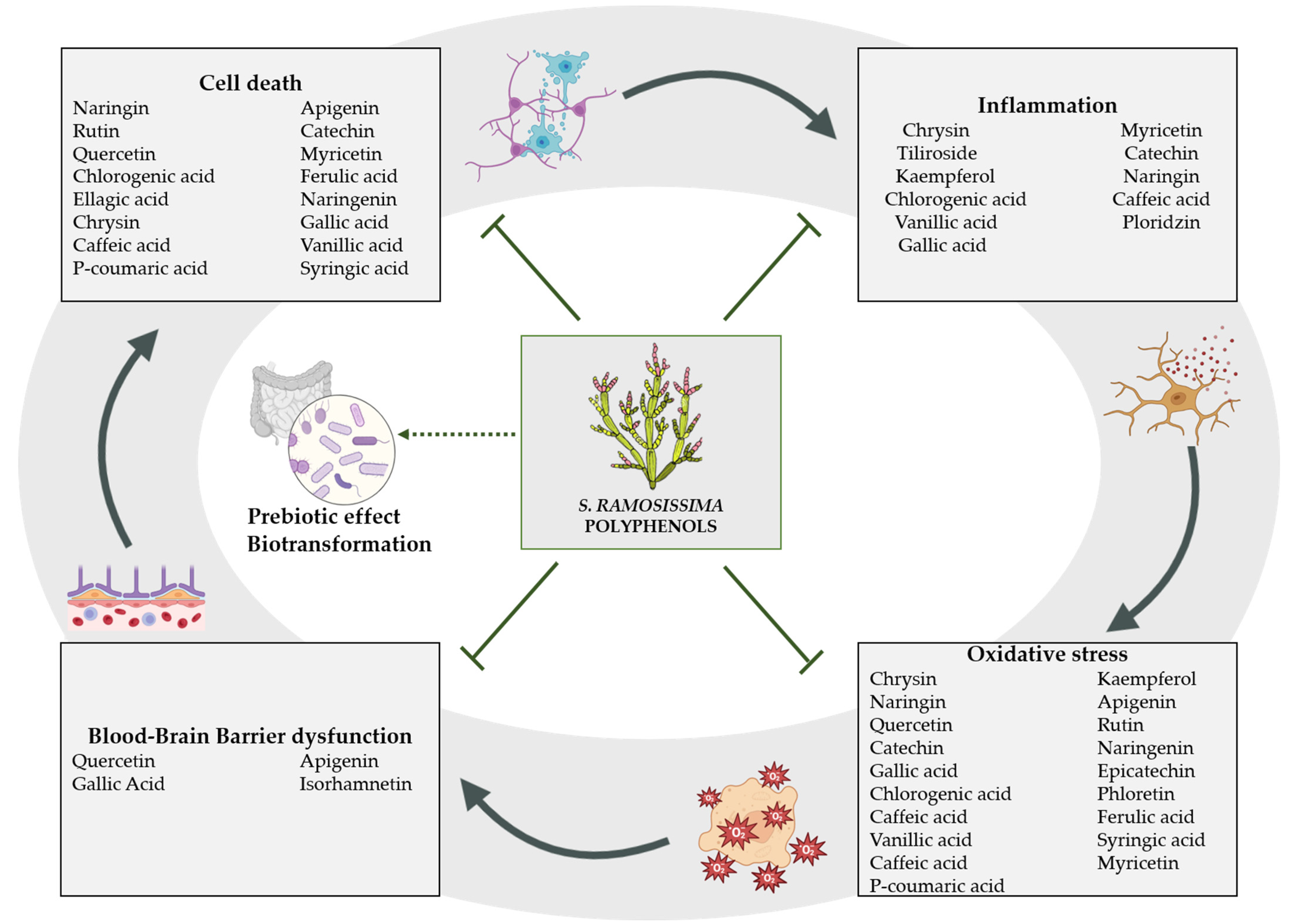
The main polyphenols found in S. ramosissima are phenolic acids and flavonoids, as shown in Table 1. However, it has been reported that the polyphenol content varies in response to salinity levels and other stress factors [14,19]. These compounds have been of interest in recent decades due to their therapeutic effect in different conditions, including a protective role in ischemia, anti-thrombotic effects and other health-promoting neurovascular benefits.
3. Bioactive Compounds with Neuroprotective Effect against Brain Ischemia
3.1. Polyphenols
3.1.1. Phenolic Acids
Phenolic acids are formed by the substitution for hydrogen atoms on benzene rings by a carboxylic acid group and at least one hydroxyl [26]. Phenolic acids are the most commonly studied and a diverse class of plant polyphenols. High concentrations are found in a variety of plant-based foods such as seeds, skins of fruits and leaves of vegetables [27]. Phenolic acids are mainly divided into two sub-groups: hydroxybenzoic (HBA) and hydroxycinnamic acid (HCA), which are based on one-carbon side-chain (C6–C1) and three-carbon side-chain (C6–C3) structure, respectively. HBAs are found abundantly in oilseeds, cereals, coffee, cowpea, black currant, raspberry, squash shells and seeds, and blackberry. HCAs are sourced mainly from coffee, cherries, cereals, peaches, spinach, citrus juices and fruits, plums, tomatoes, potatoes and almonds [28]. Phenolic acids are abundantly consumed in regular human diet, with a daily intake of about 200–400 mg/day depending on the dietary regime [29,30], since these compounds are not evenly distributed in foods [31].
Gallic acid (GA, 3, 4, 5-trihydroxybenzoic acid) is among the best studied polyphenols and its antioxidant and neuroprotective properties have been widely investigated in vivo and in vitro [32]. Sun et al. revealed that GA pretreatment protected SH-SY5Y neuroblastoma cells against hypoxia/reoxygenation injury induced by sodium hydrosulfite (Na2S2O4) in a concentration-dependent manner. This in vitro experiment showed that pretreatment with GA reversed ROS-induced lipid peroxidation, decreasing malondialdehyde (MDA) levels, and reduced mitochondrial dysfunction by inhibiting the mitochondrial permeability transition pore (MPTP) opening. The transient MCAO (tMCAO) rat model also shows the neuroprotective effects of GA in vivo. GA intravenous treatment 20 min before the onset of ischemia decreased infarct volume and neurological deficits. GA reduced the release of cytochrome complex (Cyt C, an inducer of caspase-dependent death) to cytoplasm in the infarct area and apoptosis [33]. GA has also been reported to improve the BBB disruption caused by global ischemia/reperfusion (I/R), probably thanks to its antioxidant properties. In this study, it was demonstrated that treatment with GA once daily for 10 days before ischemia ameliorated depression and anxiety-like behaviors. GA also improved hippocampal electrical activity induced by I/R, which has long been hypothesized to play a role in cognition [34]. In addition, anti-inflammatory properties have been attributed to GA. Intraperitoneal GA treatment at different time points after tMCAO induction downregulated the expression of brain ionized calcium binding adaptor molecule-1 (Iba-1), which is a marker for activated microglia, the main immune effector cells in the central nervous system (CNS). Consistently, GA modulated cytokine production towards an M2 anti-inflammatory profile [35]. Interestingly, both studies found that microglial phenotype modulation plays a role in the GA neuroprotective effect on BBB [34,35].
As a powerful antioxidant, caffeic acid also exerts anti-inflammatory effects. Zhou et al. showed that treatment with caffeic acid for 5 days starting immediately before cerebral I/R ameliorated neurological symptoms, neuron loss in the ischemic core and the infarct volume 24 h after reperfusion. Moreover, during the chronic phase after reperfusion, caffeic acid also attenuated delayed injuries, especially the astrocyte proliferation in the boundary zone adjacent to the ischemic core. However, treatment with caffeic acid did not reduce the brain edema in the acute phase [36]. Similarly, Liang et al. revealed in a global cerebral I/R injury model in rats that pretreatment with a single dose of caffeic acid 30 min prior to ischemia can significantly improve learning and memory function as well as preserve neurons in the hippocampus in a dose dependent manner during the chronic phase after I/R. Authors showed that the mechanism of protection of this phenolic acid is related to an increase of superoxide dismutase (SOD) activity, inhibition of Nuclear Factor Kappa ꞵ (NF-κꞵ) activation in the nucleus and reduced MDA content [37]. Interestingly, both studies found that 5-lipoxygenase (5-LOX, a pro-inflammatory mediator) was inhibited by caffeic acid. Because inflammation and oxidative stress are involved in neuronal injury, the inhibition of 5-LOX by caffeic acid may partially explain its therapeutic effect on brain ischemia.
Chlorogenic acid (CGA) is the ester of caffeic acid and quinic acid and has been reported to have antioxidant [38], anti-inflammatory [39] and anti-apoptotic properties [40]. In this sense, intraperitoneal administration of CGA to rats 2 h after MCAO reduced brain infarction and attenuated neurological dysfunction and brain edema. CGA treatment was shown to impede caspase activation, which is a crucial mediator of apoptosis, but also alleviated the increase in poly ADP-ribose polymerase (PARP) activity, a DNA repair enzyme involved in tissue damage during ischemia [41]. In a second study, these authors reported the role of CGA in the disruption of the neuroinflammatory cascade during I/R by reducing the activation of NF-κB, as well as glial cells’ reactivity, two main regulators of the production of proinflammatory cytokines [39]. CGA is quickly absorbed in the rat stomach in its intact form [42] and it has been estimated that, in humans, one third of CGA is absorbed in the small intestine [43]. Accordingly, oral administration of CGA had a dose-dependent neuroprotective effect on global brain I/R in rats, which was abolished in the presence of an inhibitor of the nuclear erythroid 2-related factor 2 (Nrf2) pathway. The oxidative stress reduction improved the brain tissue pathology and decreased cortical cell apoptosis. Moreover, CGA mitigated memory impairment and reversed the inhibition of the neurotrophic factors brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF), alleviating nerve injury [44]. In agreement with this, Zheng et al. recently reported the protective role of CGA pretreatment on rat neonatal stroke by regulating the Nrf2-NF-κB signaling pathway [45].
Ferulic acid (FA, 4-hydroxy-3-methoxycinnamic acid), a derivative of cinnamic acid, is one of the main active ingredients used in traditional Chinese medicine. Pharmacological studies have shown this phenolic acid to have a variety of biological activities, especially on oxidative stress and inflammation [46], endothelial injury [47] and regulation of lipid metabolism [48]. FA was reported to exert neuroprotective effects against experimental stroke by activating p38 mitogen-activated protein kinase (MAPK) signaling, which is involved in cell growth and differentiation, among other cell processes. Intravenous administration of FA using different treatment regimes (pre- and post-ischemia) reduced the area of cerebral infarction and suppressed inflammation and apoptosis after tMCAO in rats [49]. The anti-apoptotic effect of FA in brain ischemia was also confirmed by Ren et al., who reported that FA treatment for 5 consecutive days after ischemia in a global brain I/R model in rats improved spatial learning and memory deficits, increased the generation of the cellular antioxidant defenses (SOD and GSH) and reduced MDA levels. They suggested that FA exerts anti-apoptotic effects by increasing B-cell lymphoma-2 (Bcl-2) levels and inhibiting caspase-3 and Bax-mediated apoptotic signaling in the hippocampus in a concentration-dependent manner [50]. Bcl-2 is a key regulator of the mitochondrial apoptosis pathway localized in the outer membrane of mitochondria and forms a heterodimer with the apoptotic protein Bax. This prevents Bax homodimerization and triggers the activation of terminal caspases, releasing Cyt C and ROS into the cytoplasm and promoting apoptosis [51].
Another cinnamic acid derivative, p-coumaric acid (PC, 4-hydroxycinnamic acid), has also been studied for the treatment of brain ischemia in vivo. Guven et al. tested the effect of intraperitoneal PC 5 min after focal brain ischemia in rats and revealed an improvement in neurological deficit, decreased MDA levels and increased SOD activity, confirming the alleviation of oxidative stress levels by PC treatment after brain I/R injury. This study suggested that PC activates both extracellular signal-regulated kinase (ERK) and Akt signaling pathways, leading to the inhibition of apoptosis [52]. Regarding its oral availability, PC is rapidly absorbed by the monocarboxylic acid transporter present in colon epithelial cells in an intact form [53]. A second study confirmed the neuroprotective effect of PC when administered orally prior to the induction of global brain I/R in mice. PC treatment for two weeks reduced infarction volume and improved brain oxidative status, revealed by a decrease in MDA and increased levels of the antioxidant enzymes SOD and catalase [54].
Vanillic acid (VA, 4-Hydroxy-3-methoxybenzoic acid) is a dihydroxybenzoic acid derivative used for treating various ailments. VA has shown neuroprotective effects in in vivo models of ischemia. VA treatment for 14 days prior to focal brain I/R in rats reduced both neurological deficit and cerebral infarct volume. VA also downregulated the expression of NF-κB-related downstream inflammatory genes and showed excellent free radical scavenging activity [55]. Furthermore, in a similar study using global ischemia, VA decreased cognitive impairments, probably as a consequence of reduced inflammation and apoptotic cell death in the CA1 region of the hippocampus [56].
The neuroprotective effect of syringic acid (SA, 4-hydroxy-3,5-di-methoxybenzoic acid), a polyphenolic derivative of benzoic acid, has been demonstrated using in vitro and in vivo studies. SA pretreatment exerted antioxidant effects in a model of hippocampal neurons subjected to oxygen-glucose deprivation/reoxygenation (OGD/R)-induced cell injury, possibly through the JNK and p38 signaling pathways. Furthermore, SA attenuated the loss of cell viability and apoptosis [57]. Additionally, SA treatment showed similar effects when administered 5 min after permanent MCAO (pMCAO), ameliorating oxidative stress and reducing neuronal degeneration [58].
Other less studied phenolic acids in the stroke field include sinapic acid (SP, sinapine) and ellagic acid (EA). SP is a small naturally occurring hydroxycinnamic acid and its effects have been studied in global brain I/R. Intraperitoneal treatment with SP protected rats against neuronal damage, especially preserving hippocampal CA1 neuronal density, which is critical in learning and memory processes and consistent with an improvement in cognitive decline [59]. On the other hand, EA is a low-molecular-weight polyphenol derived from fruits, vegetables and nuts. EA exhibited neuroprotective effects through the Bcl-2 pathway both in vitro and in a global brain I/R injury in rats. EA treatment prevented primary neuron death and decreased the ratio of Bcl-2/Bax expression, reducing apoptosis. In vivo data showed that EA treatment significantly reduced infarct volume using a prothrombotic stroke model. EA was shown to increase the number of Bcl-2 positive neurons and the Bcl-2/Bax heterodimer ratio in the ischemic semi-dark zone and ameliorated the neurological deficit [60].
Interestingly, we recently found cannabidiolic acid (CBDA) to be present in S. ramosissima samples [13]. CBDA is the carboxylated precursor of CBD, a well-known anti-inflammatory and neuroprotective cannabinoid that was shown to reduce brain damage and neuronal loss in adult and neonatal models of ischemic stroke [61,62]. However, CBDA has been less studied than its decarboxylated form and its effects on ischemia have not yet been studied.
The neuroprotective effect of some derivatives of phenolic acids identified in S. ramosissima have also been described. The administration of 4-hydroxy-3,5-di-tret-butyl cinnamic acid, a cinnamic acid derivative, contributed to the restoration of energy-producing mitochondrial function in the hippocampus as well as ameliorated oxidative stress and apoptosis in rats submitted to pMCAO [63].
Although not all S. ramosissima phenolic acids have been studied in experimental models of cerebral ischemia, numerous studies support the therapeutic activity of this type of compound in different diseases. This is due to the large range of biological effects they display, which include inhibition of intracellular Ca2+ concentration and the consequent mitochondrial damage and free radical production that ultimately leads to the initiation of neuronal death processes, as well as anti-inflammatory properties, through the inhibition of microglial activation and the modulation of pro-inflammatory mediators such as LOX-5. Table 2 summarizes the available in vivo studies analyzing the effect on brain ischemia of phenolic acids present in S. ramosissima.
3.1.2. Flavonoids
Flavonoids, like phenolic acids, constitute one of the largest subgroups of polyphenols. High amounts of these compounds can be found in fruits and vegetables, such as citrus fruit, berries and apples, as well as seeds, legumes and beverages of plant origin [64]. Among flavonoids’ functions in the plant, they play a role in response to environmental factors, such as the coloration of fruits and flowers, and participate in the defense against UV radiation and pathogen infection. These compounds are found in plant vacuoles and their chemical structure consists of a skeleton along with three rings (C6-C3-C6), which are called A, B and C [65].
Regarding their classification, flavonoids are grouped according to their chemical structure, taking into account the degree of unsaturation and oxidation of the rings and the carbon of the C ring on which the B ring is attached. In this way, subgroups include flavanones, present mainly in citrus; flavanols, present in eucalyptus and buckwheat leaves; flavones, found in mint, chamomile and ginkgo biloba; chalcones, which lack the C ring and are found in tomatoes or strawberries; anthocyanins, responsible for the red and blue pigment of plants and therefore found to a greater extent in currants, grapes, strawberries or blueberries; and iso-flavonoids, mainly found in leguminous plants [66].
Many flavonoids described in S. ramosissima have been studied for their antioxidant activity. In addition, some have shown neuroprotective potential. Table 2 summarizes in vivo studies on the effect of flavonoids present in S. ramosissima on brain ischemia. This is the case for chrysin, a flavone which behaves as an antioxidant and anti-inflammatory compound, preventing neurotoxicity in vitro [67]. Moreover, oral administration of chrysin three weeks prior to brain ischemia in rats improved sensorimotor and memory parameters, as well as increased hippocampal cell survival in adult male Wistar rats [68]. The mechanisms by which this flavonoid exerts neuroprotection are varied. First, chrysin has been shown to maintain the integrity of the cell membrane, thereby lessening necrosis [67]. Oral administration prior to tMCAO in mice was also reported to decrease infarct volume and neurological deficit through reducing oxidative stress by dropping the formation of ROS, as well as behaving as a scavenger for O2. Moreover, anti-inflammatory activity has also been described, achieved by two routes; firstly, the inhibition of the expression of proinflammatory genes such as iNOS and COX-2 and, secondly, through the suppression of the NFκB pathway [69].
One of the most commonly studied flavonoids is quercetin, which is involved in several plant physiological processes, such as seed germination, photosynthesis and plant growth. Quercetin and its derivatives can present different biological activities due to modifications at significant positions of the quercetin molecule, such as glycosylation or methylation [70]. Quercetin was shown to protect human brain microvascular endothelial cells from I/R in vitro. At the cellular level, quercetin promoted cell viability and induced cell migration and angiogenesis. Quercetin induced the activation of the protective Keap1/Nrf2 signaling pathway, and consequently reduced ROS formation. Moreover, this flavonoid may have the ability to maintain the integrity of the BBB by sustaining tight-junction protein expression [71]. The protective effect on the neuro-vasculature has also been reported in vivo. In addition to reducing infarct volume and neurological function, intracerebroventricular injection of quercetin 30 min before global ischemia in rats was able to maintain the BBB integrity, including reduced permeability and ultrastructural alterations. Quercetin was found to activate the canonical Wnt/β-catenin signaling, which is involved in cell proliferation [72]. Unfortunately, oral bioavailability of quercetin is poor and can be affected by many factors such as glycosylation, solubility, vitamin C status or food matrix [73].
Within the flavonol subgroup, kaempferol, has been shown to protect against endothelial damage in vitro by lessening oxidative stress, specifically by reducing the levels of ROS and NO [74]. Kaempferol was also shown to protect against neurovascular disease in vivo. Preventive treatment by gavage for one week significantly reduced the volume of the infarct area as well as the neurological deficit score 24 h after tMCAO in rats. The mechanism of protection involved the regulation of various pathological pathways related to oxidative stress and inflammation, including Nrf2, Akt, NF-kβ and Gsk3β [75,76]. Similarly, two glycoside derivatives, namely kaempferol-3-O-rutinoside and kaempferol-3-O- glucoside, were found to be effective in the reduction of brain infarct by reducing brain cell inflammation mediated by the inhibition of NF-kβ and STAT3 [77].
Naringin, which is responsible for the bitterness of grapefruit and citrus skin, has also been reported to be neuroprotective in I/R models. It was shown in PC12 cells that naringin treatment decreased oxidative stress and apoptosis after OGD, partially attributed to increased Bcl-2 and decreased Bax levels. In the same way, naringin modulated several pathways involved in the inflammatory process, such as NF-kβ, Akt or mTOR [78]. Similarly, naringin promoted cell viability and reduced apoptosis after OGD in rat primary neurons, probably related to the activation of the Akt pathway [79]. Furthermore, prophylactic treatment with intraperitoneal naringin for seven days before tMCAO in male rats decreased the infarct volume in the brain, as well as apoptosis level [79]. Interestingly, naringin was found to cross the BBB when administered intravenously. Narigin can also exert neuroprotection when administered just at the beginning of reperfusion after tMCAO in rats and it was found to directly scavenge ONOO− [80].
Naringenin is also a citrus flavonoid that can be found in beverages such as wine and coffee. Several studies have reported positive health effects, including a reduction in the vascular risk [81] or an anti-inflammatory activity [82]. Several in vivo studies have shown naringenin to offer neuroprotection in both ischemic and hemorrhagic stroke models. On the one hand, treating C57BL/6 male mice with naringenin by oral gavage 4 days before the induction of subarachnoid hemorrhage was shown to reduce the associated neurological deficit and cerebral edema. In addition, a reduction in inflammation and apoptosis was observed. Interestingly, the protective effect of naringenin in this model was abolished in the presence of either an AMPK or a Sirtuin-3 inhibitor [83]. On the other hand, oral administration of naringenin was shown to protect against experimental ischemic stroke. Preventive treatment for 21 days in male Wistar rats prior to tMCAO significantly reduced the infarct area and improved neurological deficit by suppressing NF-κB-mediated neuroinflammation [84].
Phloretin is a member of the dihydrochalcone flavonoids and is well known in dermatology for its anti-aging and depigmenting effects [85]. In addition, phloretin can exert a wide range of biological activities. Administration of intravenous phloretin was long ago proposed as a tool to drop the hazardous supply of blood glucose to the post-ischemic brain [86]. More recently, Liu and colleagues showed that intraperitoneal administration of phloretin for 14 days prevented neurological deficits and reduced infarct volume 24 h after tMCAO in rats. The mechanism underlying this protective effect seems to involve the activation of the Nrf2 defense pathway [87]. Accordingly, it has been reported that phloretin induced the activation of Nrf2 pathway mediated by the phosphorylation of AMPK, which consequently reduced the inflammatory phenotype of mice macrophages [88]. As happens with phloretin, phloridzin also plays a role in mediating glucose uptake in the brain [89] and has been shown to protect male mice from experimental stroke [90].
Epicatechin, a major polyphenol component of green tea and cocoa, has been shown to protect vasculature and to have neuroprotective capacity when studied in vivo. Epicatechin is rapidly absorbed in humans and has been reported to cross the BBB [91]. Oral consumption of pure (–) epicatechin enhanced flow-mediated vasodilation in humans [92] and several studies have demonstrated that oral administration of this flavonoid can prevent both ischemic [93,94] and hemorrhagic stroke [95] using rodent models. Consistently, all the studies have attributed this protective effect to the activation of the Nrf2 pathway.
As epicatechin, catechin is one of the major phenolic components of green tea. This flavonoid has been shown to have a great scavenging capacity, one of the mechanisms involved in its protective effect against ischemia [96]. One of the first studies with catechin on ischemia was carried out in 1998 in which Mongolian gerbil rodents were supplemented with catechin in drinking water for two weeks prior and one week after tMCAO. A dose-dependent increase in hippocampal cell viability was observed with the treatment. As expected, a decrease in O2− was reported due to its scavenging activity, as well as a decrease in iNOS [97]. A different study later supported these results. After supplementing male Wistar rats with a green tea catechin extract in drinking water 5 days prior to tMCAO, authors reported a decrease in the infarct area of the brain and an improvement in the neurological deficit. The mechanisms underlying these effects include its scavenging activity as well as in the consequent effect on inflammation and vascular damage [98].
Rutin is a flavonoid glycoside with multiple pharmacological activities and substantial evidence has attributed its cytoprotective effect in ischemic tissues, including the brain. In vitro, treatment of PC12 cells prior to OGD attenuated the deleterious effect of hypoxia through a considerable decrease of ROS and the blockade of the apoptosis pathway by means of reduction of Bax and increase of Bcl-2 [99]. In addition, several studies have reported that oral supplementation with rutin can reduce the area of cerebral infarction and regulate the total amount of oxidative stress parameters, such as carbonylated proteins or antioxidant enzymes [100]. However, rutin has poor solubility, which can affect absorption and limit its oral application [101].
Apigenin, within the flavone subgroup, has beneficial effects on the vasculature which may reduce the risk of neurovascular disease. When given to rats, apigenin has behaved as a potent vasorelaxant [102]. Moreover, apigenin was shown to play a role in vascular inflammation [103] but also in platelet aggregation and secretion [104]. When used as a treatment against hypoxia in cellular models, apigenin exerted protection through the reduction of ROS and apoptosis [105] and induced angiogenesis by increasing the tube formation through a caveolin-1 dependent pathway [106]. Intraperitoneal treatment with apigenin attenuated brain damage after tMCAO in rats [105,107] and has been reported to preserve the proper functioning of the BBB by increasing caveolin-1 and to promote angiogenesis by rising VEGFS levels [107]. In a different study, intraperitoneal treatment with apigenin for 28 days following brain ischemia was shown to preserve nerve cells in rat brain hippocampus through multiple mechanisms involving histone deacetylases [108]. As happens with other flavonoids, apigenin is practically insoluble in highly polar solvents such as water and oral bioavailability is poor, as ingested apigenin is either excreted unabsorbed or rapidly metabolized after absorption [109].
Myricetin is mainly present in the glycoside form (O-glycosides) in food and beverages, such as tea and wine [110]. Among different biological actions, protective potential of myricetin against brain ischemia has been recently reviewed [111]. Using OGD in SH-SY5Y cells, Taheri et al. showed a beneficial effect of myricetin treatment on mitochondrial dysfunction and oxidative stress, two critical mechanisms of ischemic neuronal death [112]. In addition, myricetin was shown to reduce BV2 microglia proinflammatory phenotype [113] and decreased the enhancement of endothelial permeability and inflammation in human brain micro-vessel endothelial cell after OGD by regulating Akt and Nrf2 pathways [114], which could potentially protect the BBB breakdown after stroke. In experimental models of stroke in rats, oral administration of myricetin significantly reduced brain infarct volume and neurological symptomatology. It reduced apoptosis, inflammation and oxidative stress, and these protective effects were also related to Nrf2 and Akt pathways [115].
Taxifolin, also called dihydro-quercetin, is mostly found in milk thistle [116]. It is a flavanol-type flavonoid which has been shown to play a role in inflammation [117], cardiovascular risk [118] and neurological diseases, such as Alzheimer [119], among other health promoting benefits. Turovskaya et al. have shown that taxifolin can protect hippocampal neurons from OGD in vitro. Authors reported that taxifolin was able to reduce oxidative stress by lessening ROS production as well as enhancing cell viability by suppressing the apoptosis pathway through the increase in Bcl-2 [120]. These results are supported by in vivo studies using taxifolin. Intravenous treatment with taxifolin after pMCAO resulted in a dose-dependent decrease in the infarct area related to the reduction of COX-2 and iNOS, as well as a decrease in the adhesion protein ICAM-1. Moreover, a reduction in ROS was observed by the inhibition of NFκB [121].
Isorhamnetin is one of the main compounds of Ginko biloba leaves and Hippophae rhamnoides fruit [122]. Several studies support the use of this flavonoid as a therapeutic agent against cardio-cerebrovascular diseases [123,124]. At the cellular level, isorhamnetin treatment in HT22 mouse hippocampal neurons elicited a reduction of OGD-induced apoptosis, oxidative stress and inflammation. The mechanisms involving these effects are varied, including the increase in Akt, SIRT1, Nrf2, and HO-1 [125]. Intraperitoneal treatment with isorhamnetin attenuated brain damage after tMCAO in mice by reducing cerebral edema through the maintenance of tight-junction proteins at the BBB. In addition, isorhamnetin inhibited apoptosis through the down-regulation of caspase-3 and reduced oxidative stress and inflammation [126].
Tiliroside is a glycosidic flavonoid with several health promoting effects including anti-inflammatory [127] and antioxidant [128] activities. Regarding neuroinflammation, tiliroside was shown to inhibit deleterious NF-κB and p38 signaling pathways in LPS + IFNγ-activated BV2 microglia [129]. However, its role in cerebral ischemia has not yet been studied.

{kind=link}
Table 2.
Summary of in vivo studies on the neuroprotective effect of phenolic acids and flavonoids identified in S. ramosissima against cerebral ischemia.
Table 2.
Summary of in vivo studies on the neuroprotective effect of phenolic acids and flavonoids identified in S. ramosissima against cerebral ischemia.
| Polyphenol | Model | Treatment | Observed Effects | Molecular Mechanism | Ref. |
|---|---|---|---|---|---|
| Gallic acid | Male SD rats | 20 min before tMCAO (25, 50 mg/kg; i.v.) | Decreased infarct volume Anti-apoptosis Alleviated mitochondrial dysfunction | ↓ Cyt C ↓ MPTP | [33] |
| Male Wistar rats | Once daily for 10 days before transient 4VO (100 mg/kg; p.o.) | Ameliorated brain oxidative stress Improved the BBB disruption Alleviated anxiety, depression, locomotion behaviors | ↑ SOD ↓ MDA | [34] | |
| Male C57BL/6J mice | 30 min, 1, 12, 24, 48 and 72 h after ischemia in tMCAO (50, 100, 150 mg/kg; i.p.) | Reduced infarct area and edema Improved BBB disruption Anti-inflammatory Improved neurological function Inhibited microglial activation | ↓ IL-1β, TNF-α, IL-6 ↑ IL-10 ↓ MMP-9 ↑ ZO-1, Claudin-5 ↓ Iba-1 | [35] | |
| Caffeic acid | Male SD rats | 30 min before and from 0 h to 5th day after tMCAO (10, 50 mg/kg; i.p.) | Decreased infarct volume and neuron loss Ameliorated neurological dysfunction Attenuated late astrocyte proliferation | ↓ 5-LOX | [36] |
| Male SD rats | 30 min before BCCAO combined with hypotension (10,30, 50 mg/kg; i.p.) | Preserved hippocampal neurons Anti-apoptosis Improved learning and memory function Reduced brain oxidative stress Anti-inflammatory | ↑ SOD ↓ MDA ↓ 5-LOX ↓ NF-κBp65 | [37] | |
| Chlorogenic acid | Male SD rats | For 7 days before BCCAO (20, 100, 500 mg/kg; p.o.) | Reduced infarct volume and hippocampal neuron loss Anti-apoptosis Relieved nerve injury Ameliorated oxidative stress | ↑ BDNF, NGF ↑ SOD, GSH ↓ MDA, ROS ↑ Nrf2/NQO-1/HO-1 | [44] |
| Male SD rats | 2 h after pMCAO (30 mg/kg; i.p.) | Alleviated brain infarction and edema Anti-apoptosis Improved neurobehavioral deficits | ↓ ROS, LPO ↓ Caspase-3, caspase-7 ↓ PARP | [41] | |
| Male SD rats | 2 h after pMCAO (30 mg/kg; i.p.) | Ameliorated oxidative stress Inhibits the activation of astrocytes and microglia Anti-inflammatory | ↓ ROS, LPO ↓ GFAP, Iba-1 ↓ NF-κB ↓ IL-1β, TNF-α | [39] | |
| Ferulic acid | Male SD rats | Pre (2 and 4 h) and post (0,2 and 24 h) tMCAO (100 mg/kg; i.v.) | Alleviated brain infarction Anti-apoptosis Suppressed reactive astrocytosis Improved neurological deficits | ↑ p38 MAPK/p90RSK/CREB/Bcl-2 signaling pathway ↓ GFAP ↓ Mitochondrial Bax ↓ Cyt C, Caspase-3 | [49] |
| Male SD rats | 5 consecutive days after BCCAO (28, 56, 112 mg/kg) | Reduced hippocampal neuron loss Anti-apoptosis Improved memory deficits Anti-oxidative stress | ↑ Bcl-2/Bax ratio ↓ Caspase-3 ↑ SOD, GSH ↓ MDA | [50] | |
| P-coumaric acid | Male SD rats | 5 min after pMCAO (100 mg/kg; i.p.) | Anti-oxidative stress Anti-apoptosis Ameliorated neurological deficits | ↑ Nrf1, SOD ↓ MDA ↓ caspase-3, caspase-9 ↑ ERK, Akt ↓ ASK1 | [52] |
| Male ICR mice | 2 weeks before BCCAO (100 mg/kg; p.o.) | Reduced infarction size Ameliorated brain oxidative stress Anti-apoptosis | ↑ SOD, CAT ↓ MDA ↓ calcium | [54] | |
| Vanillic acid | Male SD rats | Once daily for 14 days before tMCAO (50, 100 mg/kg; p.o.) | Ameliorated cerebral infarct volume Anti-inflammatory Ameliorated oxidative stress Reduce neurological deficits | ↓ NF-κB ↓ IL-1β, IL-6, TNF-α ↓ MDA ↑ CAT, SOD | [55] |
| Male Wistar rats | Once daily for 14 days before BCCAO (100 mg/kg; p.o.) | Reduced hippocampal neuron loss Anti-inflammatory Anti-apoptosis Reversed cognitive deficits | ↑ IL-10, IL-6, TNF-α | [56] | |
| Syringic acid | Male SD rats | 5 min after pMCAO (10 mg/kg; i.p.) | Reduced histopathological changes Anti-oxidative stress Anti-apoptosis | ↑ NRF1, SOD ↓ MDA ↓ Caspase-3, Caspase-9 | [58] |
| Sinapic acid | Male Wistar rats | 0 and 90 min aftertransient 4VO (10 mg/kg; i.p.) | Reduced hippocampal neuronal loss Improved cognitive impairment | [59] | |
| Ellagic acid | Male SD rats | Once daily for 14 days before photothrombotic nerve injury (10, 30 mg/kg; p.o.) | Decreased the volume of infarction Decreased apoptosis Ameliorated neurological deficits | ↑ Bcl-2 | [60] |
| Chrysin | Male Wistar rats | Once daily 3 weeks prior to BCCAO, (10, 30, 100 mg/kg; p.o.) | Anti-apoptosis Attenuated memory impairment and sensorimotor parameters Ameliorated oxidative stress Decreased reactive hyperemia | ↑ GPx ↓ MDA ↓ NO ↓ PGE2 | [68] |
| Male C57/BL6 mice | Once daily for 7 days before tMCAO (75 mg/kg; p.o.) | Reduced infarct volume and neuron loss Anti-inflammatory activity Anti-oxidative effects | ↓ NF-κB, COX-2 ↓ iNOS ↑ SOD ↓ MDA ↓ GFAP, Iba-1 | [69] | |
| Kaempferol | Male SD rats | Once daily for 1 week before tMCAO (1.75, 3.49, 6.99 mM, 1 mL/kg; p.o.) | Decrease infarction volume Improved neurological deficit Anti-inflammatory Anti-oxidative effects | ↑ Nrf2 ↑ Akt ↓ NF-kβ, Gsk3β | [75] |
| Naringin | Male SD rats | Once daily for 7 days before tMCAO (5 mg/kg; i.p.) | Decreased infarction volume Anti-apoptosis | ↓ TNF-α ↓ IL-6 | [79] |
| Male SD rats | Once at reperfusion after tMCAO (80, 120, 160 mg/kg; i.v.) | Decreased infarction volume Reduced neurological damage Anti-apoptosis | ↓ ONOO− | [80] | |
| Phloretin | Male SD rats | Once daily for 14 days prior to tMCAO (20, 40, 80 mg/kg; i.p.) | Reduced infarct volume Anti-oxidative stress Reduced neurological damage | ↑ Nrf2 | [87] |
| Quercetin | Male SD rats | Twice daily for 3 days before BCCAO (25 μmol/kg; i.cv.) | Reduced hippocampal neuron loss Improved neurologic function Reduced brain edema Improved BBB permeability | ↑ Claudin-5, ZO-1 ↓ MMP-9 ↑ Wnt/β-catenin signaling | [72] |
| Epicatechin | Male C57BL/6 mice | 90 min prior to pMCAO (5, 10, 15 mg/kg; p.o.) | Reduced infarct volume and neuron loss Improved motor coordination Anti-oxidative stress | ↑ Nrf2 ↓ Iba-1 | [93] |
| Male C57BL/6 mice | 90 min prior to tMCAO (2.5, 5, 15, 30 mg/kg; p.o.) | Decreased infarction volume Improved neurological score | ↑ Nrf2 | [94] | |
| Apigenin | Male SD rats | Once daily for 7/14 days after tMCAO (25 mg/kg; i.p.) | Reduced infarct volume Anti-apoptosis Improved BBB function Magnification in angiogenesis | ↑ VEGFs ↑ Caveolin-1 | [107] |
| Male SD rats | Once daily for 7 days after tMCAO (25 mg/kg; i.p.) | Decreased infarction volume Improved neurological score | ↓ ROS | [105] | |
| Male SD rats | Once daily for 25 days after tMCAO (20, 40 mg/kg; i.p.) | Decreased infarction volume Improved neuron viability Improve neurological score | ↑ BDNF ↑ Syn-1 | [106] | |
| Myricetin | Male SD rats | Once daily for 7 days prior to pMCAO (1, 5, 25 mg/kg; p.o.) | Decreased infarction volume Anti-inflammatory Anti-apoptosis Decreased oxidative stress | ↓ TNF-α, IL-6, IL-1β ↑ SOD ↓ MDA | [115] |
| Rutin | Male Wistar rats | Pretreatment for 21 days before tMCAO (25 mg/kg; orally) | Decreased oxidative stress Attenuated apoptosis Reduction in infarct size Improved neurobehavioral deficits | ↑ GPx, GR, SOD, CAT, GSH ↓ H2O2, PC ↓ p53 | [100] |
| Catechin | Mongolian gerbils | Once daily for 14 days prior and 7 days post tMCAO (5, 50 mg/kg; solved in drinking water) | Improved hippocampal neuron viability | ↓ iNOS ↓O2− | [97] |
| Male Wistar rats | 5 days prior tMCAO (0.25%, 0.5%; solved in drinking water) | Decreased infarction volume Improve neurological score | ↓ MDA ↓ iNOS ↓ NF-κB | [98] | |
| Naringenin | Male Wistar rats | Once daily for 21 days prior tMCAO (10, 25, 50 mg/kg; p.o.) | Decreased infarction volume Improve neurological score Improved neuron viability | ↑ SOD ↓ iNOS ↓ NF-κB,TNF-α | [84] |
| Phloridzin | Male ddY mice | 0 and 6 h after tMCAO (40, 120, 200 mg/kg; i.p.) (10, 40 µg; i.c.v.) | Decreased infarction volume Improved neurological score Decreased FBG | ↓SGLT | [90] |
| Taxifolin | Male Long-Evans rats | 1 h after pMCAO (0.1, 1 µg/kg; i.v.) | Decreased infarction volume | ↓ iNOS, COX-2 ↓ ICAM-1 ↓ NF-κB | [121] |
| Isorhamnetin | Male ICR mice | 0 h after tMCAO (5 mg/kg; i.p.) | Decreased infarction volume Reduced brain edema Improved BBB function | ↑ Claudin-5, ZO-1, occludin ↓ TNF-α, IL-6, IL-1β ↓ MDA | [126] |
Symbols: (↑) increase; (↓) decrease. 4VO: 4-vessel occlusion; 5-LOX: 5-lipoxygenase; Akt: protein kinase B; ASK1: apoptosis signal-regulating kinase 1; Bax: Bcl-2-associated X protein; BBB: blood–brain barrier; BCCAO: bilateral common carotid arteries occlusion; Bcl-2: B-cell lymphoma 2; BDNF: brain-derived neurotrophic factor; CAT: catalase; COX-2: Cyclooxygenase 2; CREB: cAMP response element-binding; Cyt C: cytochrome complex; ERK: extracellular signal-regulated kinase; FBG: fasting blood glucose; GFAP: glial fibrillary acidic protein; GPx: glutathione peroxidase; GR: glutathione reductase; GSH: glutathione; Gsk3B: glycogen synthase kinase 3 beta; H2O2: hydrogen peroxide; HO-1: hemo oxigenasa-1; i.c.v.: intracerebroventricular injection; i.v.: intravenous injection; i.p.: intraperitoneal injection; Iba-1: brain ionized calcium binding adaptor molecule-1; IL: interleukin; iNOS: inducible nitric oxide synthase; LPO: lipid peroxidation; MAPK: mitogen-activated protein kinase; MDA: malondialdehyde; MMP-9: matrix metallopeptidase 9; MnSOD: manganese superoxide dismutase; MPTP: mitochondrial permeability transition pore; NF-Κꞵ: Nuclear Factor Kappa ꞵ; NGF: nerve growth factor; NQO-1: NAD(P)H quinone dehydrogenase 1; Nrf11: Nuclear respiratory factor 1; Nrf2: nuclear factor erythroid 2-related factor 2; O2−: superoxide anion; ONOO−: peroxy-nitrite; p.o.: per os; p90RSK: P90 ribosomal S6 kinase; PARP: poly ADP-ribose polymerase; PC: protein carbonyl; PGE2: prostaglandin E2; pMCAO: permanent middle cerebral artery occlusion; ROS: reactive oxygen species; SD: Sprague Dawley; SGLT: sodium-glucose transporter; SOD(2): superoxide dismutase (2); syn-1: synapsin 1; tMCAO: transient middle cerebral artery occlusion; TNF-α: tumor necrosis factor-α; VEGFs: vascular endothelial growth factors; ZO-1: zonula occludens-1.
4. Conclusions and Future Perspectives
In the present study, we have reviewed the effects of the different phenolic acids and flavonoids present in S. ramosissima. In vivo studies have reported that most of the identified polyphenols can ameliorate neuron loss in the ischemic core and reduce infarct volume. The associated functional improvements include the attenuation of neurological dysfunction and depression and anxiety-like behaviors, as well as balancing learning and memory processes and sensorimotor parameters. In addition, some of these polyphenolic compounds can mitigate BBB disruption and cerebral edema. Mechanisms underlying these health benefits include polyphenols’ powerful ability to eliminate ROS, but also their action on different signaling pathways involved in oxidative stress, inflammation and apoptosis.
In addition to the aforementioned properties, polyphenols have shown to exert neuroprotective effects indirectly by regulating gut microbiota. The beneficial health effects of polyphenols mainly depend on their bioavailability and absorption rate [130]. A large proportion of phenolic compounds are not directly absorbed and remain in the colon to be metabolized by gut microbiota, where they are converted into polyphenolic metabolites with higher bioavailability by gut microorganisms [131]. Interestingly, when administered in combination with oat β-glucan, polyphenols were shown to regulate the gut microbiota community phenotype and increased probiotics in high-fat-diet fed mice [132]. Accordingly, it was recently reported that polyphenol enriched oat extracts increased the proliferation of beneficial gut microbiota, showing a positive correlation between antioxidant activity and prebiotic effect, which suggest that these polyphenols could be used to regulate the gut microbiota composition [131]. In this sense, certain changes in gut microbiota have been described to increase the risk of a cerebrovascular event, and, inversely, stroke can induce dysbiosis [133]. Therefore, a therapeutic effect of phenolic acids and flavonoids by the modulation of gut microbiota in experimental models is plausible and future studies targeting polyphenols–gut interplay are warranted.
It is well known that consumption of certain foods, such as cereals, fruit or vegetables, can reduce many risk factors related to neurovascular disease [134]. The wide range of health benefits of polyphenols has drawn the attention of the food industry and many phenolic acids and flavonoids found in S. ramosissima are also available commercially as dietary supplements with different health claims.
Phenolic acids are easily absorbed by the gastrointestinal tract [135,136] and some of the aforementioned compounds are marketed as food products. As an example, cinnamic acid is marketed in liquid formulation as oral drops intended to support bladder dysfunction, cystitis or asthma, among other conditions, and ferulic acid tablets are recommended as antioxidants. In addition, sitostanol is used as an ingredient in food products designed to lower cholesterol levels and both the Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA) allows manufacturers of products that contain sito-stanol to claim this health benefit.
Most compounds within the flavonoid group are marketed as herbal extracts or natural isolated compounds from different plants. Phloretin is marketed as a nutricosmetic, a dietary supplement capsule containing green apple extract to support skin health. Several flavanols supplements from green tea extract (catechin, epicatechin, epigallocatechin) are intended for boost sport performance. In addition, Taxifolin tablets are commercialized as a help to improve venous circulation. Narigining and apigenin are suggested, among other uses, for improving cognitive function. Chrysin has been shown to be an inhibitor of aromatase enzyme activity [137] and is used as a phytoestrogen to regulate the conversion of testosterone into estrogen. Naturally extracted luteolin, quercetin and rutin are used to fight oxidative stress and inflammation and restore immune health. However, it is well known that the low bioavailability of dietary flavonoids is a major limitation for their use [138].
In addition, some polyphenolic-rich mixtures are also available as botanical extracts, such as tea, coffee or berry extracts.
Despite clinical trials showing that polyphenol supplements are safe to consume and well tolerated [139], to our knowledge there is only one human study assessing a S. ramosissima extract. Authors reported an inhibitory effect of Salicornia on hyperkinesis, an analgesic effect and a role on skin barrier architecture [139,140,141]. However, this study investigated S. ramosissima as a skin cream application and further clinical studies are needed on the oral safety and tolerability of S. ramosissima polyphenol-rich extracts in humans. In this sense, preclinical studies support both the safety [142] and the use of S. ramosissima polyphenolic profile for the treatment of ischemia [13,143]. Moreover, Salicornia extracts from other Salicornia species have been better studied in neuroprotection, such as S. europaea [144] or S. herbacea [145]. Altogether, since neurodegenerative processes develop not only in the acute stage of ischemia, but also progress throughout the survival period after ischemia [146], S. ramosissima polyphenolic profile may be a promising complementary agent in the future against the development of post-ischemic brain neurodegeneration, as depicted in Figure 1.
Author Contributions
Conceptualization, C.d.R. and J.M.; writing—original draft preparation, A.M.N., M.R.-B. and C.d.R.; writing—review and editing, C.d.R. and J.M.; supervision, C.d.R. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
The authors received financial support from “CSF-Proyectos estratégicos de I+D+i. Proyectos cofinanciados en un 80% por fondos del Programa Operativo FEDER de Andalucía 2014–2020”, grant number PE-0527-2019. This research was partially funded by “Consejería de Transformación Económica, Industria, Conocimiento y Universidades (CTEICU) y 80% cofinanciados por la UE, PO FEDER Andalucía 2014–2020”, grant number [PY20_01351]. C.R. received financial support from the Sara Borrell program funded by ISCIII, grant number CD21/00148. M.R. received financial support from the PFIS program funded by ISCIII grant number FI22/00202.
Data Availability Statement
No applicable.
Acknowledgments
We thank Yasin Hanaee for his help with language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lopes, M.; Sanches-Silva, A.; Castilho, M.; Cavaleiro, C.; Ramos, F. Halophytes as source of bioactive phenolic compounds and their potential applications. Crit. Rev. Food Sci. Nutr. 2021, 2, 1–24. [Google Scholar] [CrossRef]
- Giordano, R.; Saii, Z.; Fredsgaard, M.; Hulkko, L.S.S.; Poulsen, T.B.G.; Thomsen, M.E.; Henneberg, N.; Zucolotto, S.M.; Arendt-Nielsen, L.; Papenbrock, J.; et al. Pharmacological Insights into Halophyte Bioactive Extract Action on Anti-Inflammatory, Pain Relief and Antibiotics-Type Mechanisms. Molecules 2021, 26, 3140. [Google Scholar] [CrossRef]
- Antunes, M.D.; Gago, C.; Guerreiro, A.; Sousa, A.R.; Julião, M.; Miguel, M.G.; Faleiro, M.L.; Panagopoulos, T. Nutritional Characterization and Storage Ability of Salicornia ramosissima and Sarcocornia perennis for Fresh Vegetable Salads. Horticulturae 2021, 7, 6. [Google Scholar] [CrossRef]
- Lopes, M.; Cavaleiro, C.; Ramos, F. Sodium Reduction in Bread: A Role for Glasswort (Salicornia ramosissima J. Woods). Compr. Rev. Food Sci. Food Saf. 2017, 16, 1056–1071. [Google Scholar] [CrossRef]
- Choi, S.C.; Kim, B.J.; Rhee, P.L.; Chang, D.K.; Son, H.J.; Kim, J.J.; Rhee, J.C.; Kim, S.I.; Han, Y.S.; Sim, K.H.; et al. Probiotic Fermented Milk Containing Dietary Fiber Has Additive Effects in IBS with Constipation Compared to Plain Probiotic Fermented Milk. Gut Liver 2011, 5, 22–28. [Google Scholar] [CrossRef]
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic stroke. Nat. Rev. Dis. Prim. 2019, 5, 70. [Google Scholar] [CrossRef]
- Kuriakose, D.; Xiao, Z. Pathophysiology and Treatment of Stroke: Present Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef]
- Berge, E.; Whiteley, W.; Audebert, H.; De Marchis, G.M.; Fonseca, A.C.; Padiglioni, C.; de la Ossa, N.P.; Strbian, D.; Tsivgoulis, G.; Turc, G. European Stroke Organisation (ESO) guidelines on intravenous thrombolysis for acute ischaemic stroke. Eur. Stroke J. 2021, 6, I-LXII. [Google Scholar] [CrossRef]
- Soares, R.O.S.; Losada, D.M.; Jordani, M.C.; Evora, P.; Castro, E.S.O. Ischemia/Reperfusion Injury Revisited: An Overview of the Latest Pharmacological Strategies. Int. J. Mol. Sci. 2019, 20, 5034. [Google Scholar] [CrossRef]
- Woodruff, T.M.; Thundyil, J.; Tang, S.C.; Sobey, C.G.; Taylor, S.M.; Arumugam, T.V. athophysiology, treatment, and animal and cellular models of human ischemic stroke. Mol. Neurodegener. 2011, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Hankey, G.J. Secondary stroke prevention. Lancet Neurol. 2014, 13, 178–194. [Google Scholar] [CrossRef]
- Hackam, D.G.; Spence, J.D. Combining multiple approaches for the secondary prevention of vascular events after stroke: A quantitative modeling study. Stroke 2007, 38, 1881–1885. [Google Scholar] [CrossRef]
- Garcia-Rodriguez, P.; Ma, F.; Rio, C.D.; Romero-Bernal, M.; Najar, A.M.; Cadiz-Gurrea, M.L.; Leyva-Jimenez, F.J.; Ramiro, L.; Menendez-Valladares, P.; Perez-Sanchez, S.; et al. Diet Supplementation with Polyphenol-Rich Salicornia ramosissima Extracts Protects against Tissue Damage in Experimental Models of Cerebral Ischemia. Nutrients 2022, 14, 5077. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gavilan, I.; Ramirez, E.; de la Fuente, V. Bioactive Compounds in Salicornia patula Duval-Jouve: A Mediterranean Edible Euhalophyte. Foods 2021, 10, 410. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019; pp. 1–15. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Falleh, H.; Trabelsi, N.; Boulaaba, M.; Smaoui, A.; Abdelly, C. Influence of biological, environmental and technical factors on phenolic content and antioxidant activities of Tunisian halophytes. Comptes Rendus Biol. 2008, 331, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Irfan, M.I.-D.M.; Raghib, F.; Ahmad, B. Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2019; p. 9. [Google Scholar] [CrossRef]
- Lima, A.R.; Castaneda-Loaiza, V.; Salazar, M.; Nunes, C.; Quintas, C.; Gama, F.; Pestana, M.; Correia, P.J.; Santos, T.; Varela, J.; et al. Influence of cultivation salinity in the nutritional composition, antioxidant capacity and microbial quality of Salicornia ramosissima commercially produced in soilless systems. Food Chem. 2020, 333, 127525. [Google Scholar] [CrossRef]
- Silva, A.M.; Lago, J.P.; Pinto, D.; Moreira, M.M.; Grosso, C.; Cruz Fernandes, V.; Delerue-Matos, C.; Rodrigues, F. Salicornia ramosissima Bioactive Composition and Safety: Eco-Friendly Extractions Approach (Microwave-Assisted Extraction vs. Conventional Maceration). Appl. Sci. 2021, 11, 4744. [Google Scholar] [CrossRef]
- Oliveira-Alves, S.C.; Andrade, F.; Prazeres, I.; Silva, A.B.; Capelo, J.; Duarte, B.; Cacador, I.; Coelho, J.; Serra, A.T.; Bronze, M.R. Impact of Drying Processes on the Nutritional Composition, Volatile Profile, Phytochemical Content and Bioactivity of Salicornia ramosissima J. Woods. Antioxidants 2021, 10, 1312. [Google Scholar] [CrossRef]
- Surget, G.; Stiger-Pouvreau, V.; Le Lann, K.; Kervarec, N.; Couteau, C.; Coiffard, L.J.; Gaillard, F.; Cahier, K.; Guerard, F.; Poupart, N. Structural elucidation, in vitro antioxidant and photoprotective capacities of a purified polyphenolic-enriched fraction from a saltmarsh plant. J. Photochem. Photobiol. B Biol. 2015, 143, 52–60. [Google Scholar] [CrossRef]
- Guerreiro, A.; Rassal, C.; Afonso, C.M.; Galego, L.; Serra, M.; Rodrigues, M.A. Healthy, Tasty and Sustainable Mediterranean Food. UMAMI Taste and Polyphenols of Twiggy Glasswort (Salicornia ramosissima). In International Congress on Engineering and Sustainability in the XXI Century; Springer: Berlin/Heidelberg, Germany, 2017; pp. 191–198. [Google Scholar] [CrossRef]
- Ferreira, D.; Isca, V.M.; Leal, P.; Seca, A.M.; Silva, H.; de Lourdes Pereira, M.; Silva, A.M.; Pinto, D.C. Salicornia ramosissima: Secondary metabolites and protective effect against acute testicular toxicity. Arab. J. Chem. 2018, 11, 70–80. [Google Scholar] [CrossRef]
- Isca, V.M.; Seca, A.M.; Pinto, D.C.; Silva, H.; Silva, A.M. Lipophilic profile of the edible halophyte Salicornia ramosissima. Food Chem. 2014, 165, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shahzad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Nat. Prod. Commun. 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Radtke, J.; Linseisen, J.; Wolfram, G. Phenolic acid intake of adults in a Bavarian subgroup of the national food consumption survey. Z. Ernahr. 1998, 37, 190–197. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Medina-Remon, A.; Perez-Jimenez, J.; Martinez-Gonzalez, M.A.; Covas, M.I.; Corella, D.; Salas-Salvado, J.; Gomez-Gracia, E.; Lapetra, J.; Aros, F.; et al. Dietary intake and major food sources of polyphenols in a Spanish population at high cardiovascular risk: The PREDIMED study. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 953–959. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A Role of Gallic Acid in Oxidative Damage Diseases: A Comprehensive Review. Nat. Prod. Commun. 2019, 14, 1934578X19874174. [Google Scholar] [CrossRef]
- Sun, J.; Li, Y.Z.; Ding, Y.H.; Wang, J.; Geng, J.; Yang, H.; Ren, J.; Tang, J.Y.; Gao, J. Neuroprotective effects of gallic acid against hypoxia/reoxygenation-induced mitochondrial dysfunctions in vitro and cerebral ischemia/reperfusion injury in vivo. Brain Res. 2014, 1589, 126–139. [Google Scholar] [CrossRef]
- Jahangiri, H.M.; Sarkaki, A.; Farbood, Y.; Dianat, M.; Goudarzi, G. Gallic acid affects blood-brain barrier permeability, behaviors, hippocampus local EEG, and brain oxidative stress in ischemic rats exposed to dusty particulate matter. Environ. Sci. Pollut. Res. 2020, 27, 5281–5292. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Wang, L.; Mao, Y. Gallic acid attenuates cerebral ischemia/re-perfusion-induced blood-brain barrier injury by modifying polarization of microglia. J. Immunotoxicol. 2022, 19, 17–26. [Google Scholar] [CrossRef]
- Zhou, Y.; Fang, S.H.; Ye, Y.L.; Chu, L.S.; Zhang, W.P.; Wang, M.L.; Wei, E.Q. Caffeic acid ameliorates early and delayed brain injuries after focal cerebral ischemia in rats. Acta Pharmacol. Sin. 2006, 27, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Shi, B.; Luo, W.; Yang, J. The protective effect of caffeic acid on global cerebral ischemia-reperfusion injury in rats. Behav. Brain Funct. 2015, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm 2011, 403, 136–138. [Google Scholar] [CrossRef]
- Shah, M.A.; Kang, J.B.; Park, D.J.; Kim, M.O.; Koh, P.O. Chlorogenic acid alleviates cerebral ischemia-induced neuroinflammation via attenuating nuclear factor kappa B activation. Neurosci. Lett. 2022, 773, 136495. [Google Scholar] [CrossRef]
- Huang, S.M.; Chuang, H.C.; Wu, C.H.; Yen, G.C. Cytoprotective effects of phenolic acids on methylglyoxal-induced apoptosis in Neuro-2A cells. Mol. Nutr. Food Res. 2008, 52, 940–949. [Google Scholar] [CrossRef]
- Shah, M.A.; Kang, J.B.; Park, D.J.; Kim, M.O.; Koh, P.O. Chlorogenic acid alleviates neurobehavioral disorders and brain damage in focal ischemia animal models. Neurosci. Lett. 2021, 760, 136085. [Google Scholar] [CrossRef]
- Lafay, S.; Gil-Izquierdo, A.; Manach, C.; Morand, C.; Besson, C.; Scalbert, A. Chlorogenic acid is absorbed in its intact form in the stomach of rats. J. Nutr. 2006, 136, 1192–1197. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar] [CrossRef]
- Liu, D.; Wang, H.; Zhang, Y.; Zhang, Z. Protective Effects of Chlorogenic Acid on Cerebral Ischemia/Reperfusion Injury Rats by Regulating Oxidative Stress-Related Nrf2 Pathway. Drug Des. Dev. Ther. 2020, 14, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, L.; Chen, B.; Fang, Y.; Lin, W.; Zhang, T.; Feng, X.; Tao, X.; Wu, Y.; Fu, X.; et al. Chlorogenic acid exerts neuroprotective effect against hypoxia-ischemia brain injury in neonatal rats by activating Sirt1 to regulate the Nrf2-NF-kappaB signaling pathway. Cell Commun. Signal. 2022, 20, 84. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.N.; Wu, W.J.; Sun, C.Z.; Liu, H.F.; Chen, W.B.; Zhan, Q.P.; Lei, Z.G.; Xin, X.; Ma, J.J.; Yao, K.; et al. Antioxidant and Anti-inflammatory Capacity of Ferulic Acid Released from Wheat Bran by Solid-state Fermentation of Aspergillus niger. Biomed. Environ. Sci. 2019, 32, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tian, L.; Liu, Y.; Liu, J.; Huang, J. Ferulic Acid Protects Endothelial Cells from Hypoxia-Induced Injury by Regulating MicroRNA-92a. Appl. Bionics Biomech. 2022, 2022, 6148361. [Google Scholar] [CrossRef]
- Yogeeta, S.K.; Hanumantra, R.B.R.; Gnanapragasam, A.; Subramanian, S.; Rajakannu, S.; Devaki, T. Attenuation of Abnormalities in the Lipid Metabolism during Experimental Myocardial Infarction Induced by Isoproterenol in Rats: Beneficial Effect of Ferulic Acid and Ascorbic Acid. Basic Clin. Pharmacol. Toxicol. 2006, 98, 467–472. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Tang, N.Y.; Kao, S.T.; Hsieh, C.L. Ferulic Acid Administered at Various Time Points Protects against Cerebral Infarction by Activating p38 MAPK/p90RSK/CREB/Bcl-2 Anti-Apoptotic Signaling in the Subacute Phase of Cerebral Ischemia-Reperfusion Injury in Rats. PLoS ONE 2016, 11, e0155748. [Google Scholar] [CrossRef]
- Ren, Z.; Zhang, R.; Li, Y.; Li, Y.; Yang, Z.; Yang, H. Ferulic acid exerts neuroprotective effects against cerebral ischemia/reperfusion-induced injury via antioxidant and anti-apoptotic mechanisms in vitro and in vivo. Int. J. Mol. Med. 2017, 40, 1444–1456. [Google Scholar] [CrossRef] [Green Version]
- Anilkumar, U.; Prehn, J.H. Anti-apoptotic BCL-2 family proteins in acute neural injury. Front. Cell Neurosci. 2014, 8, 281. [Google Scholar] [CrossRef]
- Guven, M.; Aras, A.B.; Akman, T.; Sen, H.M.; Ozkan, A.; Salis, O.; Sehitoglu, I.; Kalkan, Y.; Silan, C.; Deniz, M.; et al. Neuroprotective effect of p-coumaric acid in rat model of embolic cerebral ischemia. Iran. J. Basic Med. Sci. 2015, 18, 356–363. [Google Scholar]
- Konishi, Y.; Hitomi, Y.; Yoshioka, E. Intestinal absorption of p-coumaric and gallic acids in rats after oral administration. J. Agric. Food Chem. 2004, 52, 2527–2532. [Google Scholar] [CrossRef]
- Sakamula, R.; Thong-Asa, W. Neuroprotective effect of p-coumaric acid in mice with cerebral ischemia reperfusion injuries. Metab. Brain Dis. 2018, 33, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, Y.; Zhang, S.Y. Vanillic Acid Improve Neural Function after Focal Cerebral Ischemia-reperfusion Rats. Int. J. Pharmacol. 2018, 14, 488–494. [Google Scholar] [CrossRef]
- Khoshnam, S.E.; Sarkaki, A.; Rashno, M.; Farbood, Y. Memory deficits and hippocampal inflammation in cerebral hypoperfusion and reperfusion in male rats: Neuroprotective role of vanillic acid. Life Sci. 2018, 211, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, L.; Sun, S.; Yi, Z.; Jiang, X.; Jia, D. Neuroprotective effects of syringic acid against OGD/R-induced injury in cultured hippocampal neuronal cells. Int. J. Mol. Med. 2016, 38, 567–573. [Google Scholar] [CrossRef]
- Guven, M.; Aras, A.B.; Topaloglu, N.; Ozkan, A.; Sen, H.M.; Kalkan, Y.; Okuyucu, A.; Akbal, A.; Gokmen, F.; Cosar, M. The protective effect of syringic acid on ischemia injury in rat brain. Turk. J. Med. Sci. 2015, 45, 233–240. [Google Scholar] [CrossRef]
- Kim, Y.O.; Lee, S.W.; Oh, M.S.; Lee, H.J. Effects of sinapic Acid of 4 vessel occlusion model-induced ischemia and cognitive impairments in the rat. Clin. Psychopharmacol. Neurosci. 2011, 9, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.S.; Deng, R.; Li, S.; Li, X.; Li, K.; Kebaituli, G.; Li, X.; Liu, R. Ellagic acid protects against neuron damage in ischemic stroke through regulating the ratio of Bcl-2/Bax expression. Appl. Physiol. Nutr. Metab. 2017, 42, 855–860. [Google Scholar] [CrossRef]
- Mori, M.A.; Meyer, E.; Soares, L.M.; Milani, H.; Guimaraes, F.S.; de Oliveira, R.M.W. Cannabidiol reduces neuroinflammation and promotes neuroplasticity and functional recovery after brain ischemia. Prog. NeuroPsychopharmacol. Biol. Psychiatry 2017, 75, 94–105. [Google Scholar] [CrossRef]
- Hayakawa, K.; Mishima, K.; Fujiwara, M. Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke. Pharmacology 2010, 3, 2197–2212. [Google Scholar] [CrossRef]
- Pozdnyakov, D.I. 4-Hydroxy-3,5-di-tret-butyl cinnamic acid restores the activity of the hippocampal mitochondria in rats under permanent focal cerebral ischemia. Iran. J. Basic Med. Sci. 2021, 24, 1590–1601. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Campos, H.M.; da Costa, M.; da Silva Moreira, L.K.; da Silva Neri, H.F.; Branco da Silva, C.R.; Pruccoli, L.; Dos Santos, F.C.A.; Costa, E.A.; Tarozzi, A.; Ghedini, P.C. Protective effects of chrysin against the neurotoxicity induced by aluminium: In vitro and in vivo studies. Toxicology 2022, 465, 153033. [Google Scholar] [CrossRef]
- Shooshtari, M.K.; Sarkaki, A.; Mansouri, S.M.T.; Badavi, M.; Khorsandi, L.; Dehcheshmeh, M.G.; Farbood, Y. Protective effects of Chrysin against memory impairment, cerebral hyperemia and oxidative stress after cerebral hypoperfusion and reperfusion in rats. Metab. Brain Dis. 2020, 35, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Chen, L.; Xiao, J.; Wang, C.; Jiang, W.; Zhang, R.; Hao, J. Chrysin protects against focal cerebral ischemia/reperfusion injury in mice through attenuation of oxidative stress and inflammation. Int. J. Mol. Sci. 2014, 15, 20913–20926. [Google Scholar] [CrossRef] [PubMed]
- Michala, A.S.; Pritsa, A. Quercetin: A Molecule of Great Biochemical and Clinical Value and Its Beneficial Effect on Diabetes and Cancer. Diseases 2022, 10, 37. [Google Scholar] [CrossRef]
- Li, M.T.; Ke, J.; Guo, S.F.; Wu, Y.; Bian, Y.F.; Shan, L.L.; Liu, Q.Y.; Huo, Y.J.; Guo, C.; Liu, M.Y.; et al. The Protective Effect of Quercetin on Endothelial Cells Injured by Hypoxia and Reoxygenation. Front. Pharmacol. 2021, 12, 732874. [Google Scholar] [CrossRef]
- Jin, Z.; Ke, J.; Guo, P.; Wang, Y.; Wu, H. Quercetin improves blood-brain barrier dysfunction in rats with cerebral ischemia reperfusion via Wnt signaling pathway. Am J. Transl Res. 2019, 11, 4683–4695. [Google Scholar]
- Kaşıkcı, M.B.; Bağdatlıoğlu, N. Bioavailability of Quercetin. Curr. Res. Nutr. Food Sci. J. 2016, 4, 146–151. [Google Scholar] [CrossRef]
- Wang, J.; Li, T.; Feng, J.; Li, L.; Wang, R.; Cheng, H.; Yuan, Y. Kaempferol protects against gamma radiation-induced mortality and damage via inhibiting oxidative stress and modulating apoptotic molecules in vivo and vitro. Environ. Toxicol. Pharmacol. 2018, 60, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mao, J.; Wang, R.; Li, S.; Wu, B.; Yuan, Y. Kaempferol Protects Against Cerebral Ischemia Reperfusion Injury Through Intervening Oxidative and Inflammatory Stress Induced Apoptosis. Front. Pharmacol. 2020, 11, 424. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Del-Rio, L.; Nag, A.; Gutierrez Casado, E.; Ariza, J.; Awad, A.M.; Joseph, A.I.; Kwon, O.; Verdin, E.; de Cabo, R.; Schneider, C.; et al. Kaempferol increases levels of coenzyme Q in kidney cells and serves as a biosynthetic ring precursor. Free. Radic. Biol. Med. 2017, 110, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, C.; Wang, L.F.; Kuang, X.; Liu, K.; Zhang, H.; Du, J.R. Neuroprotective effect of kaempferol glycosiDes. against brain injury and neuroinflammation by inhibiting the activation of NF-kappaB and STAT3 in transient focal stroke. PLoS ONE 2013, 8, e55839. [Google Scholar] [CrossRef]
- Cao, W.; Feng, S.J.; Kan, M.C. Naringin Targets NFKB1 to Alleviate Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury in PC12 Cells Via Modulating HIF-1alpha/AKT/mTOR-Signaling Pathway. J. Mol. Neurosci. 2021, 71, 101–111. [Google Scholar] [CrossRef]
- Yang, J.; Yuan, L.; Wen, Y.; Zhou, H.; Jiang, W.; Xu, D.; Wang, M. Protective Effects of Naringin in Cerebral Infarction and Its Molecular Mechanism. Med. Sci. Monit. 2020, 26, e918772. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Chen, X.; Lu, S.; Li, W.; Yang, D.; Su, W.; Wang, X.; Shen, J. Naringin Attenuates Cerebral Ischemia-Reperfusion Injury Through Inhibiting Peroxynitrite-Mediated Mitophagy Activation. Mol. Neurobiol. 2018, 55, 9029–9042. [Google Scholar] [CrossRef]
- Heidary Moghaddam, R.; Samimi, Z.; Moradi, S.Z.; Little, P.J.; Xu, S.; Farzaei, M.H. Naringenin and naringin in cardiovascular disease prevention: A preclinical review. Eur. J. Pharmacol. 2020, 887, 173535. [Google Scholar] [CrossRef]
- Zeng, W.; Jin, L.; Zhang, F.; Zhang, C.; Liang, W. Naringenin as a potential immunomodulator in therapeutics. Pharmacol. Res. 2018, 135, 122–126. [Google Scholar] [CrossRef]
- Zeng, H.; Shao, B.; Zhuang, J.; Peng, Y.; Chen, H.; Yu, Q.; Xu, C.; Fu, X.; Zhou, H.; Cao, Y.; et al. Naringenin reduces early brain injury in subarachnoid hemorrhage (SAH) mice: The role of the AMPK/SIRT3 signaling pathway. J. Funct. Foods 2020, 72, 104043. [Google Scholar] [CrossRef]
- Raza, S.S.; Khan, M.M.; Ahmad, A.; Ashafaq, M.; Islam, F.; Wagner, A.P.; Safhi, M.M.; Islam, F. Neuroprotective effect of naringenin is mediated through suppression of NF-kappaB signaling pathway in experimental stroke. Neuroscience 2013, 230, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Anunciato Casarini, T.P.; Frank, L.A.; Pohlmann, A.R.; Guterres, S.S. Dermatological applications of the flavonoid phloretin. Eur. J. Pharmacol. 2020, 889, 173593. [Google Scholar] [CrossRef]
- Oldendorf, W.H.; Crane, P.D.; Lawner, P.M.; Braun, L.D. Rapid, transient drop in brain glucose after intravenous phloretin or 3-0-methyl-D-glucose. Stroke 1983, 14, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Liang, J. Activation of the Nrf2 defense pathway contributes to neuroprotective effects of phloretin on oxidative stress injury after cerebral ischemia/reperfusion in rats. J. Neurol. Sci. 2015, 351, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Dierckx, T.; Haidar, M.; Grajchen, E.; Wouters, E.; Vanherle, S.; Loix, M.; Boeykens, A.; Bylemans, D.; Hardonniere, K.; Kerdine-Romer, S.; et al. Phloretin suppresses neuroinflammation by autophagy-mediated Nrf2 activation in macrophages. J. Neuroinflamm. 2021, 18, 148. [Google Scholar] [CrossRef]
- Betz, A.L.; Drewes, L.R.; Gilboe, D.D. Inhibition of glucose transport into brain by phlorizin, phloretin and glucose analogues. Biochim. Biophys. Acta (BBA)-Biomembr. 1975, 406, 505–515. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Harada, S.; Tokuyama, S. Post-ischemic hyperglycemia exacerbates the development of cerebral ischemic neuronal damage through the cerebral sodium-glucose transporter. Brain Res. 2012, 1489, 113–120. [Google Scholar] [CrossRef]
- Nehlig, A. The neuroprotective effects of cocoa flavanol and its influence on cognitive performance. Br. J. Clin. Pharmacol. 2013, 75, 716–727. [Google Scholar] [CrossRef]
- Schroeter, H.; Heiss, C.; Balzer, J.; Kleinbongard, P.; Keen, C.L.; Hollenberg, N.K.; Sies, H.; Kwik-Uribe, C.; Schmitz, H.H.; Kelm, M. (-)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 1024–1029. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, C.C.; Agrawal, M.; Singh, N.; Moore, J.R.; Biswal, S.; Dore, S. Oral administration of the flavanol (-)-epicatechin bolsters endogenous protection against focal ischemia through the Nrf2 cytoprotective pathway. Eur. J. Neurosci. 2013, 38, 3659–3668. [Google Scholar] [CrossRef]
- Shah, Z.A.; Li, R.C.; Ahmad, A.S.; Kensler, T.W.; Yamamoto, M.; Biswal, S.; Dore, S. The flavanol (-)-epicatechin prevents stroke damage through the Nrf2/HO1 pathway. J. Cereb. Blood Flow Metab. 2010, 30, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.F.; Cho, S.; Wang, J. (-)-Epicatechin protects hemorrhagic brain via synergistic Nrf2 pathways. Ann. Clin. Transl. Neurol. 2014, 1, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef]
- Inanami, O.; Watanabe, Y.; Syuto, B.; Nakano, M.; Tsuji, M.; Kuwabara, M. Oral administration of (-)catechin protects against ischemia-reperfusion-induced neuronal death in the gerbil. Free. Radic. Res. 1998, 29, 359–365. [Google Scholar] [CrossRef]
- Suzuki, M.; Tabuchi, M.; Ikeda, M.; Umegaki, K.; Tomita, T. Protective effects of green tea catechins on cerebral ischemic damage. Med. Sci. Monit. 2004, 10, BR166–BR174. [Google Scholar] [PubMed]
- Nassiri-Asl, M.; Ghorbani, A.; Salehisar, S.; Asadpour, E.; Sadeghnia, H.R. Effect of rutin on oxidative DNA damage in PC12 neurons cultured in nutrients deprivation condition. Iran. J. Basic Med. Sci. 2020, 23, 390–395. [Google Scholar] [CrossRef]
- Khan, M.M.; Ahmad, A.; Ishrat, T.; Khuwaja, G.; Srivastawa, P.; Khan, M.B.; Raza, S.S.; Javed, H.; Vaibhav, K.; Khan, A.; et al. Rutin protects the neural damage induced by transient focal ischemia in rats. Brain Res. 2009, 1292, 123–135. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, X.; Zhang, Q.; Wang, L.; Li, Y.; Li, Y. Characterization and Evaluation of the Solubility and Oral Bioavailability of Rutin-Ethanolate Solvate. AAPS PharmSciTech 2020, 21, 241. [Google Scholar] [CrossRef]
- Woodman, O.L.; Chan, E. Vascular and anti-oxidant actions of flavonols and flavones. Clin. Exp. Pharmacol. Physiol. 2004, 31, 786–790. [Google Scholar] [CrossRef]
- Olszanecki, R.; Gebska, A.; Kozlovski, V.I.; Gryglewski, R.J. Flavonoids and nitric oxide synthase. J. Physiol. Pharmacol. 2002, 53, 571–584. [Google Scholar]
- Guerrero, J.A.; Lozano, M.L.; Castillo, J.; Benavente-Garcia, O.; Vicente, V.; Rivera, J. Flavonoids inhibit platelet function through binding to the thromboxane A2 receptor. J. Thromb Haemost 2005, 3, 369–376. [Google Scholar] [CrossRef]
- Ling, C.; Lei, C.; Zou, M.; Cai, X.; Xiang, Y.; Xie, Y.; Li, X.; Huang, D.; Wang, Y. Neuroprotective effect of apigenin against cerebral ischemia/reperfusion injury. J. Int. Med. Res. 2020, 48, 300060520945859. [Google Scholar] [CrossRef] [PubMed]
- Tu, F.; Pang, Q.; Chen, X.; Huang, T.; Liu, M.; Zhai, Q. Angiogenic effects of apigenin on endothelial cells after hypoxia-reoxygenation via the caveolin-1 pathway. Int. J. Mol. Med. 2017, 40, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.; Zhao, Y.; Chen, X.; Zhao, K.; Zhai, Q.; Tu, F. Apigenin Protects the Brain against Ischemia/Reperfusion Injury via Caveolin-1/VEGF In Vitro and In Vivo. Oxidative Med. Cell. Longev. 2018, 2018, 7017204. [Google Scholar] [CrossRef]
- Tu, F.; Pang, Q.; Huang, T.; Zhao, Y.; Liu, M.; Chen, X. Apigenin Ameliorates Post-Stroke Cognitive Deficits in Rats Through Histone Acetylation-Mediated Neurochemical Alterations. Med. Sci. Monit. 2017, 23, 4004–4013. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed. Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin bioactive effects: Moving from preclinical evidence to potential clinical applications. BMC Complement Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef]
- Pluta, R.; Januszewski, S.; Czuczwar, S.J. Myricetin as a Promising Molecule for the Treatment of Post-Ischemic Brain Neurodegeneration. Nutrients 2021, 13, 241. [Google Scholar] [CrossRef]
- Wu, S.; Yue, Y.; Peng, A.; Zhang, L.; Xiang, J.; Cao, X.; Ding, H.; Yin, S. Myricetin ameliorates brain injury and neurological deficits via Nrf2 activation after experimental stroke in middle-aged rats. Food Funct. 2016, 7, 2624–2634. [Google Scholar] [CrossRef]
- Boriero, D.; Carcereri de Prati, A.; Antonini, L.; Ragno, R.; Sohji, K.; Mariotto, S.; Butturini, E. The anti-STAT1 polyphenol myricetin inhibits M1 microglia activation and counteracts neuronal death. FEBS J. 2021, 288, 2347–2359. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, X.; Guo, S.; Shi, L.; He, Q.; Zhang, P.; Yu, S.; Zhao, R. Myricetin ameliorated ischemia/reperfusion-induced brain endothelial permeability by improvement of eNOS uncoupling and activation eNOS/NO. J. Pharmacol. Sci. 2019, 140, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, P.; Fu, T.; Huang, X.; Song, J.; Chen, M.; Tian, X.; Yin, H.; Han, J. Myricetin against ischemic cerebral injury in rat middle cerebral artery occlusion model. Mol. Med. Rep. 2018, 17, 3274–3280. [Google Scholar] [CrossRef] [PubMed]
- Sunil, C.; Xu, B. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Wang, Y.J.; Yang, G.T.; Gao, Q.L.; Tang, M.X. Taxifolin Inhibits Receptor Activator of NF-kappaB Ligand-Induced Osteoclastogenesis of Human Bone Marrow-Derived Macrophages in vitro and Prevents Lipopolysaccharide-Induced Bone Loss in vivo. Pharmacology 2019, 103, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Yang, Y.; Yang, L.; Jiang, H.; Yu, X.; Wang, Y. Cardioprotective effects of dihydroquercetin against ischemia reperfusion injury by inhibiting oxidative stress and endoplasmic reticulum stress-induced apoptosis via the PI3K/Akt pathway. Food Funct 2019, 10, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Yamamoto, Y.; Maki, T.; Hattori, Y.; Ito, H.; Mizuno, K.; Harada-Shiba, M.; Kalaria, R.N.; Fukushima, M.; Takahashi, R.; et al. Taxifolin inhibits amyloid-beta oligomer formation and fully restores vascular integrity and memory in cerebral amyloid angiopathy. Acta NeuroPathol. Commun. 2017, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Turovskaya, M.V.; Gaidin, S.G.; Mal’tseva, V.N.; Zinchenko, V.P.; Turovsky, E.A. Taxifolin protects neurons against ischemic injury in vitro via the activation of antioxidant systems and signal transduction pathways of GABAergic neurons. Mol. Cell Neurosci. 2019, 96, 10–24. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wang, W.Y.; Chang, C.C.; Liou, K.T.; Sung, Y.J.; Liao, J.F.; Chen, C.F.; Chang, S.; Hou, Y.C.; Chou, Y.C.; et al. Taxifolin ameliorates cerebral ischemia-reperfusion injury in rats through its anti-oxidative effect and modulation of NF-kappa B activation. J. BioMed. Sci. 2006, 13, 127–141. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.Y.; Zhang, Z.L.; Rahman, K.; Wang, S.J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. BioMed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef]
- Zhang, N.; Pei, F.; Wei, H.; Zhang, T.; Yang, C.; Ma, G.; Yang, C. Isorhamnetin protects rat ventricular myocytes from ischemia and reperfusion injury. Exp. Toxicol. Pathol. 2011, 63, 33–38. [Google Scholar] [CrossRef]
- Iida, A.; Usui, T.; Zar Kalai, F.; Han, J.; Isoda, H.; Nagumo, Y. Protective effects of Nitraria retusa extract and its constituent isorhamnetin against amyloid beta-induced cytotoxicity and amyloid beta aggregation. Biosci. Biotechnol. Biochem. 2015, 79, 1548–1551. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fan, L.; Wang, Y.; Ding, J.; Wang, R. Isorhamnetin Alleviates High Glucose-Aggravated Inflammatory Response and Apoptosis in Oxygen-Glucose Deprivation and Reoxygenation-Induced HT22 Hippocampal Neurons Through Akt/SIRT1/Nrf2/HO-1 Signaling Pathway. Inflammation 2021, 44, 1993–2005. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Song, J.Q.; Pan, S.Y.; Wang, K. Treatment with Isorhamnetin Protects the Brain Against Ischemic Injury in Mice. Neurochem Res. 2016, 41, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Lv, Q.; Zhong, C.; Cui, Y.; He, L.; Zhang, C.; Yu, J. Tiliroside Ameliorates Ulcerative Colitis by Restoring the M1/M2 Macrophage Balance via the HIF-1alpha/glycolysis Pathway. Front. Immunol. 2021, 12, 649463. [Google Scholar] [CrossRef]
- Han, R.; Yang, H.; Lu, L.; Lin, L. Tiliroside as a CAXII inhibitor suppresses liver cancer development and modulates E2Fs/Caspase-3 axis. Sci. Rep. 2021, 11, 8626. [Google Scholar] [CrossRef]
- Velagapudi, R.; Aderogba, M.; Olajide, O.A. Tiliroside, a dietary glycosidic flavonoid, inhibits TRAF-6/NF-kappaB/p38-mediated neuroinflammation in activated BV2 microglia. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 3311–3319. [Google Scholar] [CrossRef]
- Di Meo, F.; Valentino, A.; Petillo, O.; Peluso, G.; Filosa, S.; Crispi, S. Bioactive Polyphenols and Neuromodulation: Molecular Mechanisms in Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 8626. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Ren, X.; Zhang, X.; Wu, Z.; Liu, L. The positive correlation of antioxidant activity and prebiotic effect about oat phenolic compounds. Food Chem. 2023, 402, 134231. [Google Scholar] [CrossRef]
- Li, Y.; Qin, C.; Dong, L.; Zhang, X.; Wu, Z.; Liu, L.; Yang, J.; Liu, L. Whole grain benefit: Synergistic effect of oat phenolic compounds and beta-glucan on hyperlipidemia via gut microbiota in high-fat-diet mice. Food Funct 2022, 13, 12686–12696. [Google Scholar] [CrossRef]
- Peh, A.; O’Donnell, J.A.; Broughton, B.R.S.; Marques, F.Z. Gut Microbiota and Their Metabolites in Stroke: A Double-Edged Sword. Stroke 2022, 53, 1788–1801. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: A Comprehensive Review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef] [PubMed]
- Renouf, M.; Marmet, C.; Giuffrida, F.; Lepage, M.; Barron, D.; Beaumont, M.; Williamson, G.; Dionisi, F. Dose-response plasma appearance of coffee chlorogenic and phenolic acids in adults. Mol. Nutr. Food Res. 2014, 58, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Morand, C.; Manach, C.; Remesy, C. Absorption and metabolism of polyphenols in the gut and impact on health. BioMed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.C.; Zhou, C.; Sherman, M.; Laughton, C.A.; Chen, S. Molecular basis of the inhibition of human aromatase (estrogen synthetase) by flavone and isoflavone phytoestrogens: A site-directed mutagenesis study. Environ. Health Perspect. 1998, 106, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, S.H.; Rupasinghe, H.P. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef]
- Han, M.K.; Barreto, T.A.; Martinez, F.J.; Comstock, A.T.; Sajjan, U.S. Randomised clinical trial to determine the safety of quercetin supplementation in patients with chronic obstructive pulmonary disease. BMJ Open Respir Res. 2020, 7, e000392. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, T.; Tanaka, Y.; Kamimaki, I.; Nagao, T.; Tokimitsu, I. Catechin safely improved higher levels of fatness, blood pressure, and cholesterol in children. Obesity 2008, 16, 1338–1348. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Lilitchan, S.; Tuntipopipat, S.; Tirawanchai, N.; Komindr, S. Ferulic Acid Supplementation Improves Lipid Profiles, Oxidative Stress, and Inflammatory Status in Hyperlipidemic Subjects: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2018, 10, 713. [Google Scholar] [CrossRef]
- Lee, H.-S. Acute Oral Toxicity of Salicornia herbacea L. Extract in Mice. Biomed. Sci. Lett. 2016, 22, 46–52. [Google Scholar] [CrossRef]
- Ferreira, D.; Pinto, D.; Silva, H.; Girol, A.P.; de Lourdes Pereira, M. Salicornia ramosissima J. Woods seeds affected the normal regenerative function on carbon tetrachloride-induced liver and kidney injury. BioMed. Pharmacother. 2018, 107, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Karthivashan, G.; Park, S.Y.; Kweon, M.H.; Kim, J.; Haque, M.E.; Cho, D.Y.; Kim, I.S.; Cho, E.A.; Ganesan, P.; Choi, D.K. Ameliorative potential of desalted Salicornia europaea L. extract in multifaceted Alzheimer’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 7174. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Karthivashan, G.; Kweon, M.H.; Kim, D.H.; Choi, D.K. The Ameliorative Effects of the Ethyl Acetate Extract of Salicornia europaea L. and Its Bioactive Candidate, Irilin B, on LPS-Induced Microglial Inflammation and MPTP-Intoxicated PD-Like Mouse Model. Oxidative Med. Cell. Longev. 2019, 2019, 6764756. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Ulamek, M.; Jablonski, M. Alzheimer’s mechanisms in ischemic brain degeneration. Anat. Rec. 2009, 292, 1863–1881. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Ischemic stroke pathological mechanisms modulated by phenolic acids and flavonoids identified in S. ramosissima. As free radical scavengers, polyphenols counteract ROS generation and the inflammation triggered by oxidative stress. Some phenolic acids and flavonoids can directly act on endothelial cells and reduce BBB dysfunction or modulate mechanisms leading to cell death that prevent neuronal loss. Additionally, phenolic acids and flavonoids may have indirect beneficial effects by favoring the increase of beneficial bacteria, which in turn modulate their metabolisms and bioavailability.
Figure 1.
Ischemic stroke pathological mechanisms modulated by phenolic acids and flavonoids identified in S. ramosissima. As free radical scavengers, polyphenols counteract ROS generation and the inflammation triggered by oxidative stress. Some phenolic acids and flavonoids can directly act on endothelial cells and reduce BBB dysfunction or modulate mechanisms leading to cell death that prevent neuronal loss. Additionally, phenolic acids and flavonoids may have indirect beneficial effects by favoring the increase of beneficial bacteria, which in turn modulate their metabolisms and bioavailability.

Table 1.
Polyphenolic compounds identified in S. ramosissima.
| Polyphenol | Subclass | Compound | Ref. |
|---|---|---|---|
| Flavonoid | Dihydrochalcone | Phloretin | [20] |
| Phloridzin | [20] | ||
| Flavanol | Catechin | [20] | |
| Epicatechin | [20] | ||
| (Epi)gallocatechin | [13] | ||
| Dihydroquercetin (Taxifolin) | [21] | ||
| Flavanone | Naringin | [20] | |
| Naringenin | [20] | ||
| Flavone | Apigenin | [20] | |
| Apigenin-6-arabinosyl-8-glucoside (isoschaftoside) | [21] | ||
| Chrysin | [20] | ||
| Luteolin glucosyllactate | [13] | ||
| Flavonol | Isorhamnetin | [22] | |
| Isorhamnetin 3-glucoside | [22] | ||
| Isorhamnetin-7-O-(6-O-malonyl)-glucoside | [23] | ||
| Isorhamnetin glucopyranoside | [13] | ||
| Kaempferol | [20] | ||
| kaempferol derivative | [21] | ||
| kaempferol-3-O-glucoside | [20] | ||
| kaempferol-3-O-rutinoside | [20] | ||
| Myricetin | [20] | ||
| Quercetin | [20] | ||
| Quercetin-3-O-galactoside | [20] | ||
| Quercetin glucoside | [13] | ||
| Quercetin 3-glucoside (Isoquercitrin) | [21,22,23] | ||
| Quercetin-malonyglucoside | [13,21] | ||
| Quercetin-methyl-ether derivative (isomer 1 and 2) | [21] | ||
| Quercetin-rhamnosyl-hexoside | [13,21] | ||
| Rutin (quercetin 3 -O rhamnosyl glucoside, quercetin rutinoside, vitamin p) | [20] | ||
| Phenolic acids | Hydroxybenzoic acids | Cannabidiolic acid | [13] |
| Salicylic acid derivative | [21] | ||
| Sitostanol | [24] | ||
| Syringic acid | [20] | ||
| Tiliroside | [20] | ||
| Vanillic acid | [20] | ||
| Ellagic acid | [20] | ||
| Gallic acid | [20] | ||
| Gallocatechin | [24] | ||
| Protocatechuic acid | [20] | ||
| Protocatechuic-arabinoside acid | [21] | ||
| Hydroxycinnamic acids | Cinnamic acid | [25] | |
| P-coumaric acid (4-hydroxycinnamic acid) | [13,20,21,23] | ||
| Sinapic acid (3,5-Dimethoxy-4-hydroxycinnamic acid) | [20] | ||
| Ethyl (E)-2-hydroxycinnamate | [24] | ||
| P-coumaric acid benzyl ester derivative | [21] | ||
| Quinic acid | [13,21,23] | ||
| P-coumaroylquinic acid (isomer 1 and 2) | [21] | ||
| Caffeic acid | [20,22] | ||
| Hydrocaffeic acid | [22] | ||
| Caffeic acid-glucuronide-glucoside (isomer 1) | [21] | ||
| Caffeoylquinic acid | [22] | ||
| Chlorogenic acid (3-O-caffeoylquinic acid) | [20,21,23] | ||
| Neochlorogenic acid (5-O-caffeoylquinic acid) | [13,21] | ||
| Dicaffeoylquinic acid (isomer 1, 2, 3 and 4) | [13,22] | ||
| 3,4-Di-O-caffeoylquinic acid | [20,22] | ||
| 3,5-Di-O-caffeoylquinic acid | [20] | ||
| 3,5-Dicaffeoylquinic acid | [21] | ||
| 4,5-Dicaffeoylquinic acid | [21] | ||
| Hydrocaffeoylquinic acid | [13,21,22] | ||
| Dihydrocaffeoyl quinic acid | [22] | ||
| Caffeoyl-hydrocaffeoyl quinic acid | [21,22] | ||
| Tungtungmadic acid (3-Caffeoyl-4-dihydrocaffeoyl quinic acid) (isomer 1 and 2) | [13] | ||
| Ferulic acid | [13,21,23,25] | ||
| Ferulic-glucoside acid | [21] | ||
| Trans-ferulic acid | [20] | ||
| Coumarin | Scopoletin | [13,24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nájar, A.M.; Romero-Bernal, M.; del Río, C.; Montaner, J. A Review on Polyphenols in Salicornia ramosissima with Special Emphasis on Their Beneficial Effects on Brain Ischemia. Nutrients 2023, 15, 793. https://doi.org/10.3390/nu15030793
AMA Style
Nájar AM, Romero-Bernal M, del Río C, Montaner J. A Review on Polyphenols in Salicornia ramosissima with Special Emphasis on Their Beneficial Effects on Brain Ischemia. Nutrients. 2023; 15(3):793. https://doi.org/10.3390/nu15030793
Chicago/Turabian StyleNájar, Ana M., Marina Romero-Bernal, Carmen del Río, and Joan Montaner. 2023. "A Review on Polyphenols in Salicornia ramosissima with Special Emphasis on Their Beneficial Effects on Brain Ischemia" Nutrients 15, no. 3: 793. https://doi.org/10.3390/nu15030793
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.