Sword Bean (Canavalia gladiata) Pod Exerts Anti-Allergic and Anti-Inflammatory Effects through Modulation of Th1/Th2 Cell Differentiation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of the Extract
2.3. Cell Culture
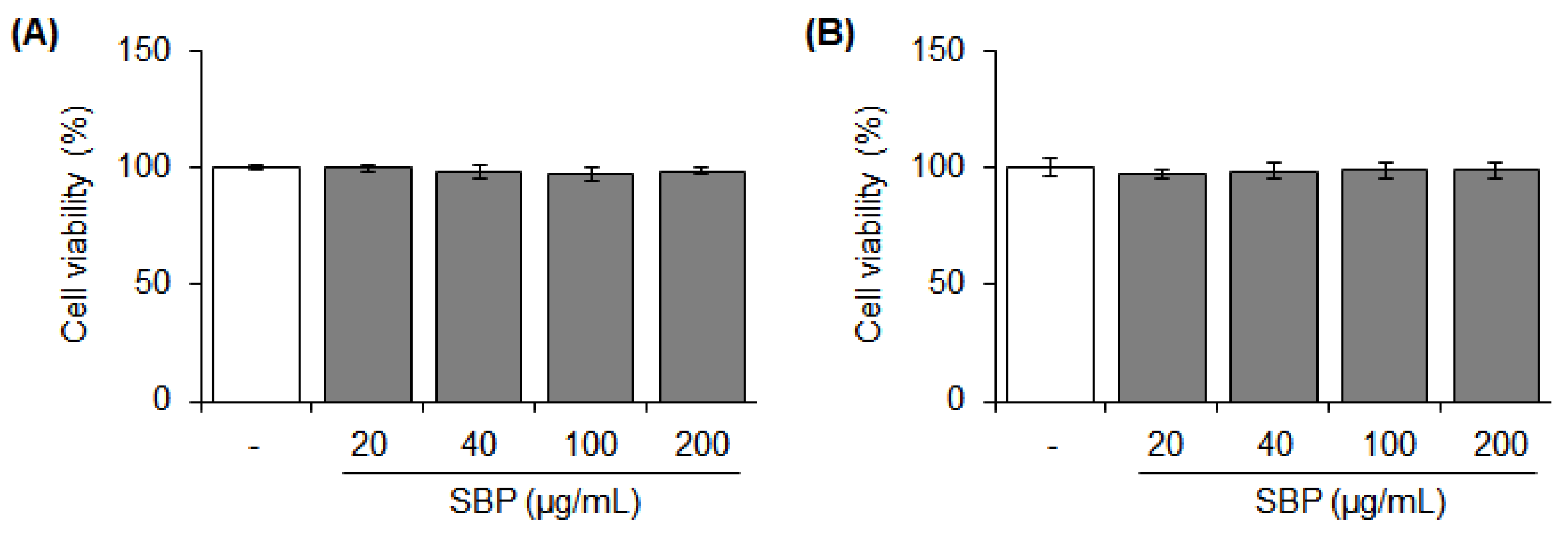
2.4. Cell Viability
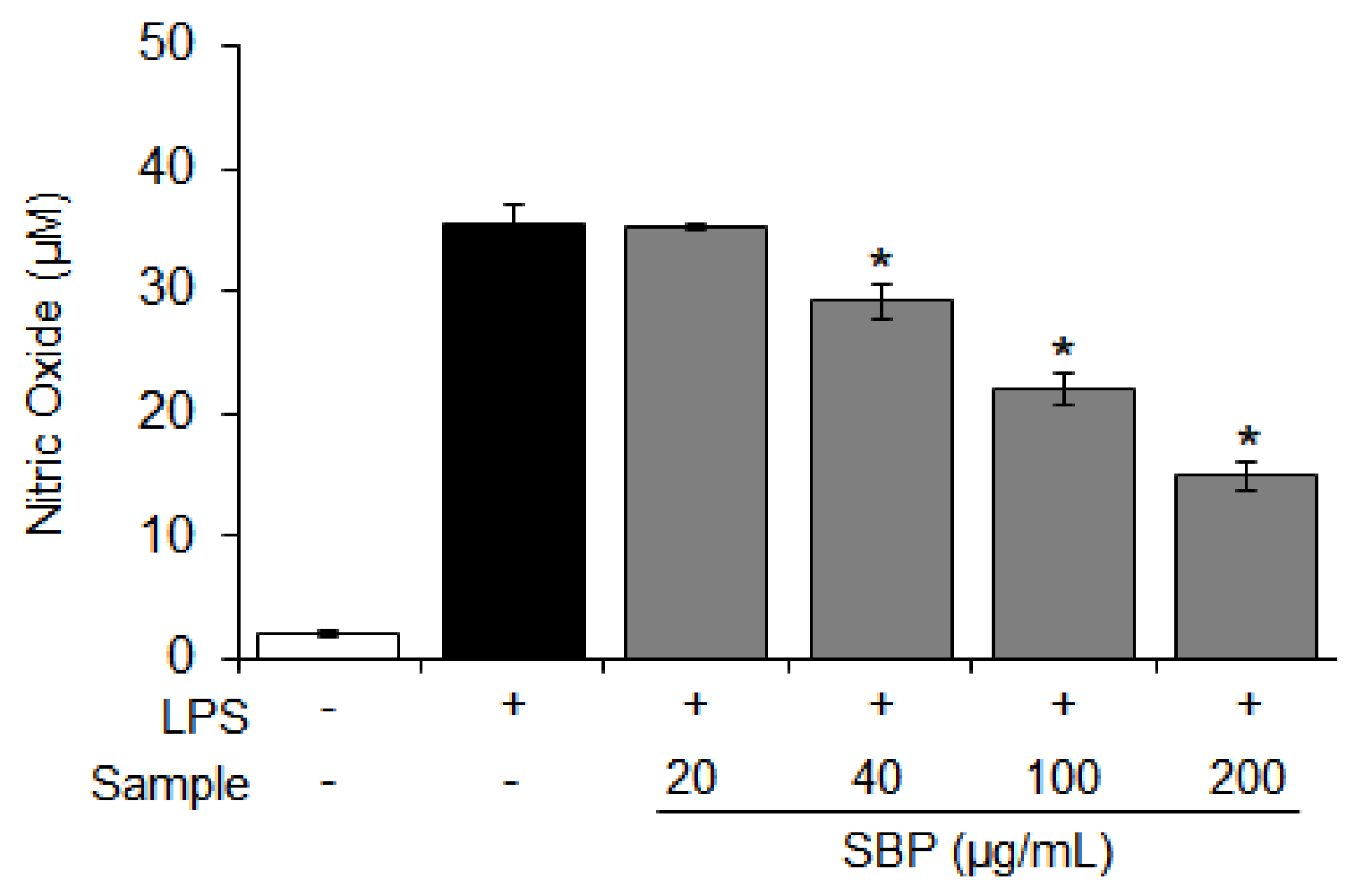
2.5. Nitric Oxide (NO) Production
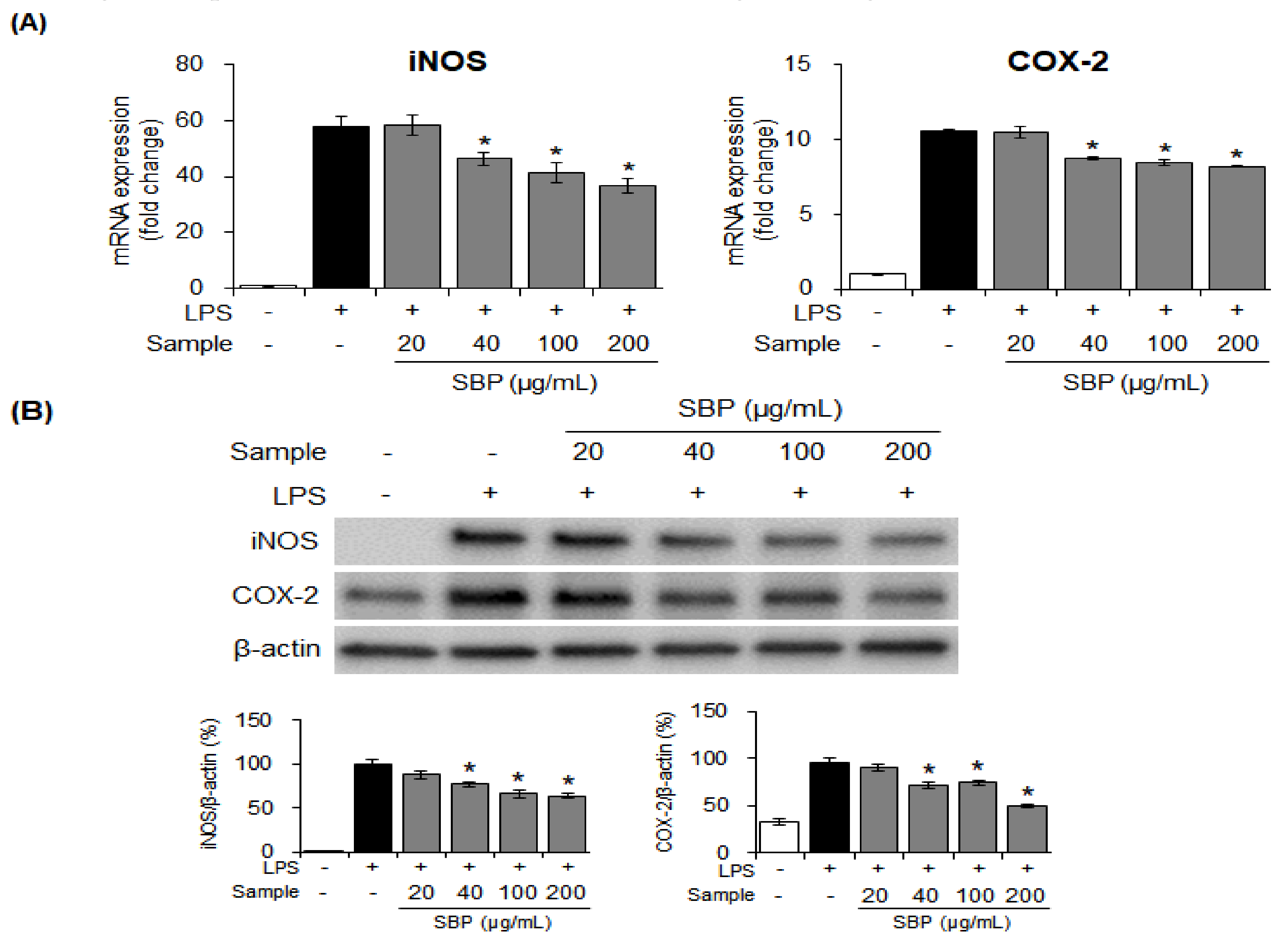
2.6. Quantitative Reverse Transcriptase-PCR (qRT–PCR)
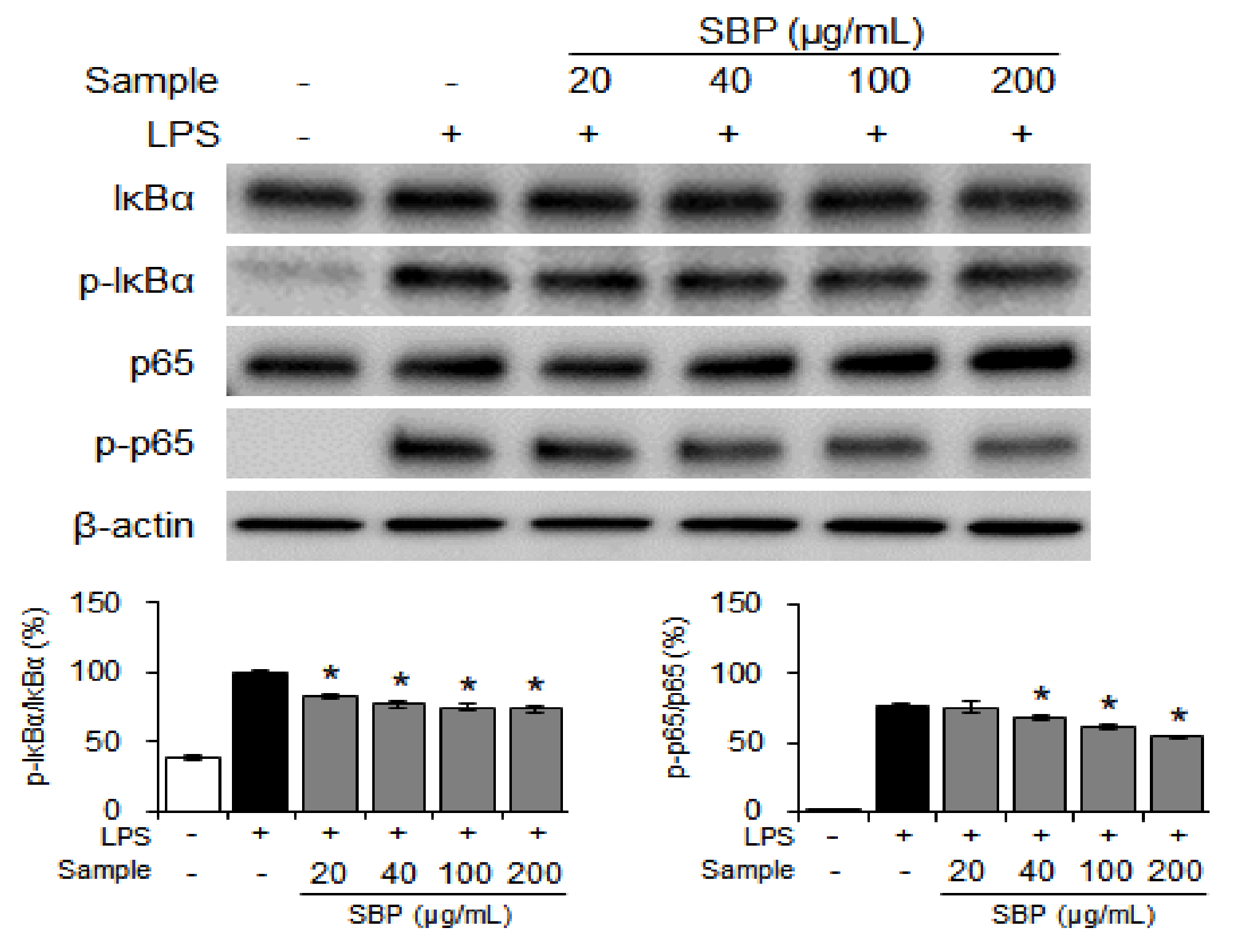
2.7. Western Blot Analysis
2.8. Degranulation in RBL-2H3 Mast Cells
2.9. Experimental Animals
2.10. Sensitization and Challenge
2.11. Measurement of Hepatotoxicity in the Plasma
2.12. Measurement of IgE and Histamine Production
2.13. Measurement of Cytokine Levels in the BALF and Plasma
2.14. Statistical Analysis
3. Results
3.1. Cytotoxicity of the SBP Extract on Raw264.7 and RBL-2H3 Cells
3.2. Effect of the SBP Extract on NO Production in LPS-Induced Raw264.7 Cells
3.3. Effects of the SBP Extract on iNOS and COX-2 mRNA and Protein Expression in LPS-induced Raw264.7 Cells
3.4. Effect of the SBP Extract on NF-κB Signaling in LPS-Induced Raw264.7 Cells
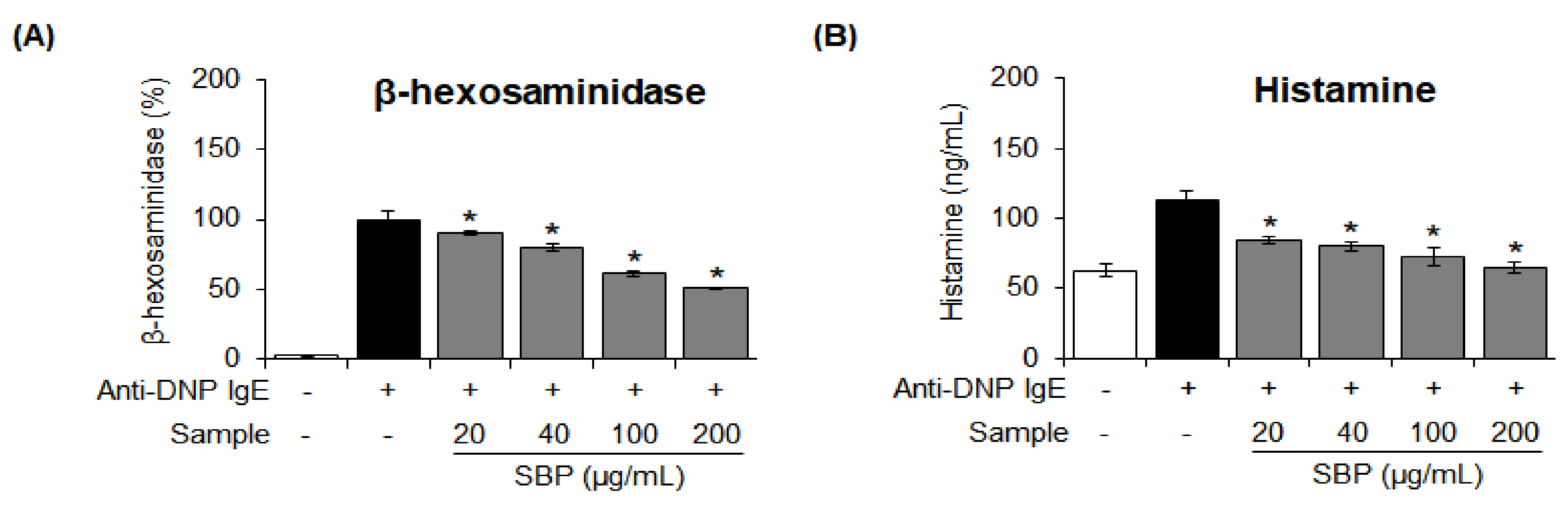
3.5. Effect of SBP Extract on the β-Hexosaminidase Release and Histamine Degranulation Stimulated by Anti-DNP IgE in RBL-2H3 Cells
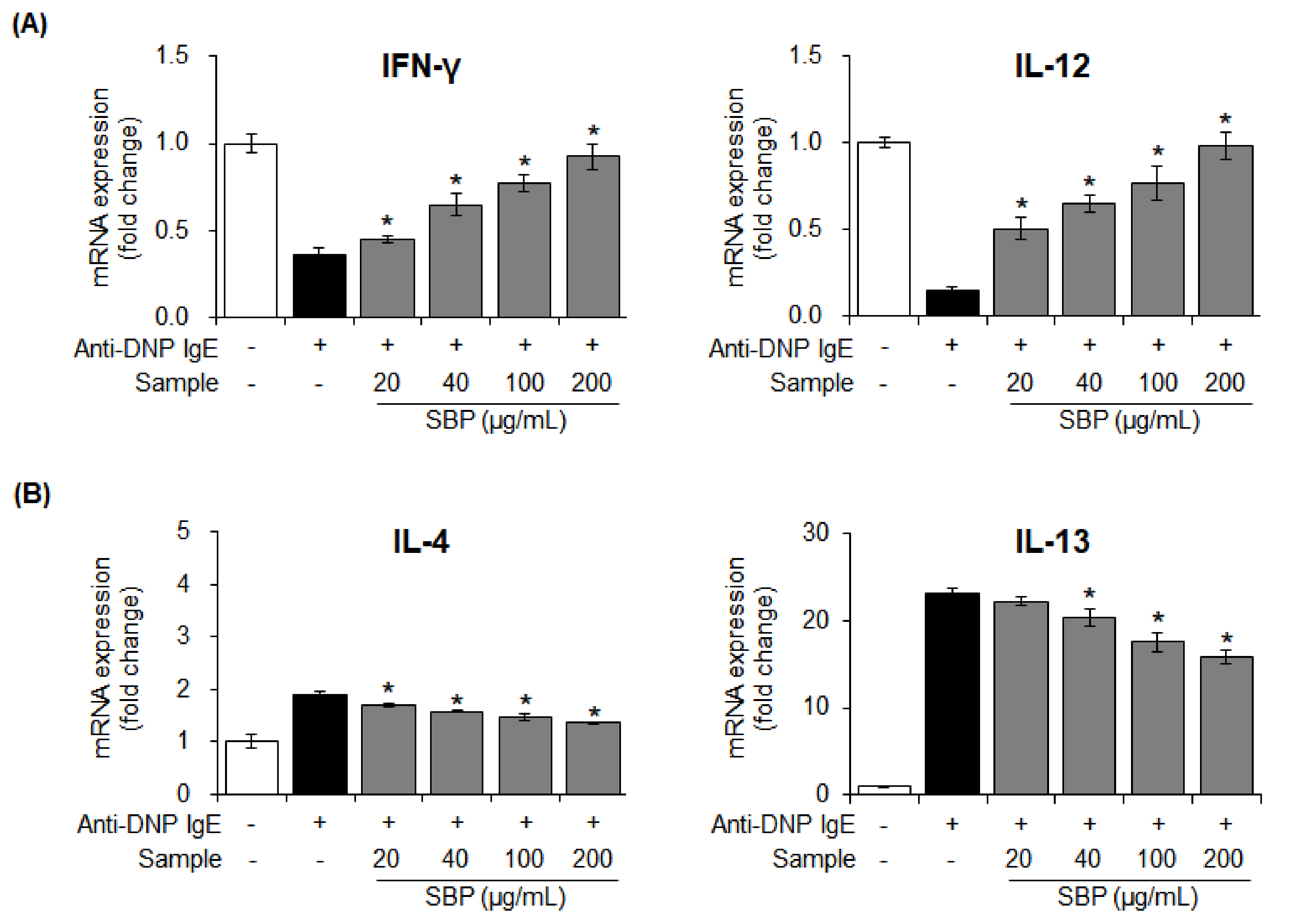
3.6. Effects of SBP Extract on the mRNA Expression of Pro- and Anti-Inflammatory Cytokines Stimulated by Anti-DNP IgE in RBL-2H3 Cells
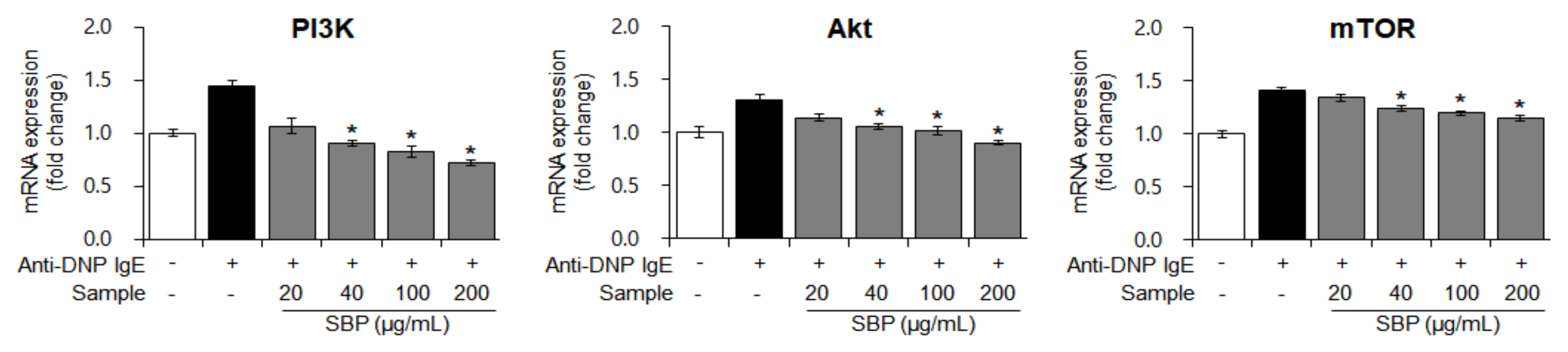
3.7. Effects of SBP Extract on the mRNA Expression of PI3K/mTOR Signaling Factors Stimulated by Anti-DNP IgE in RBL-2H3 Cells
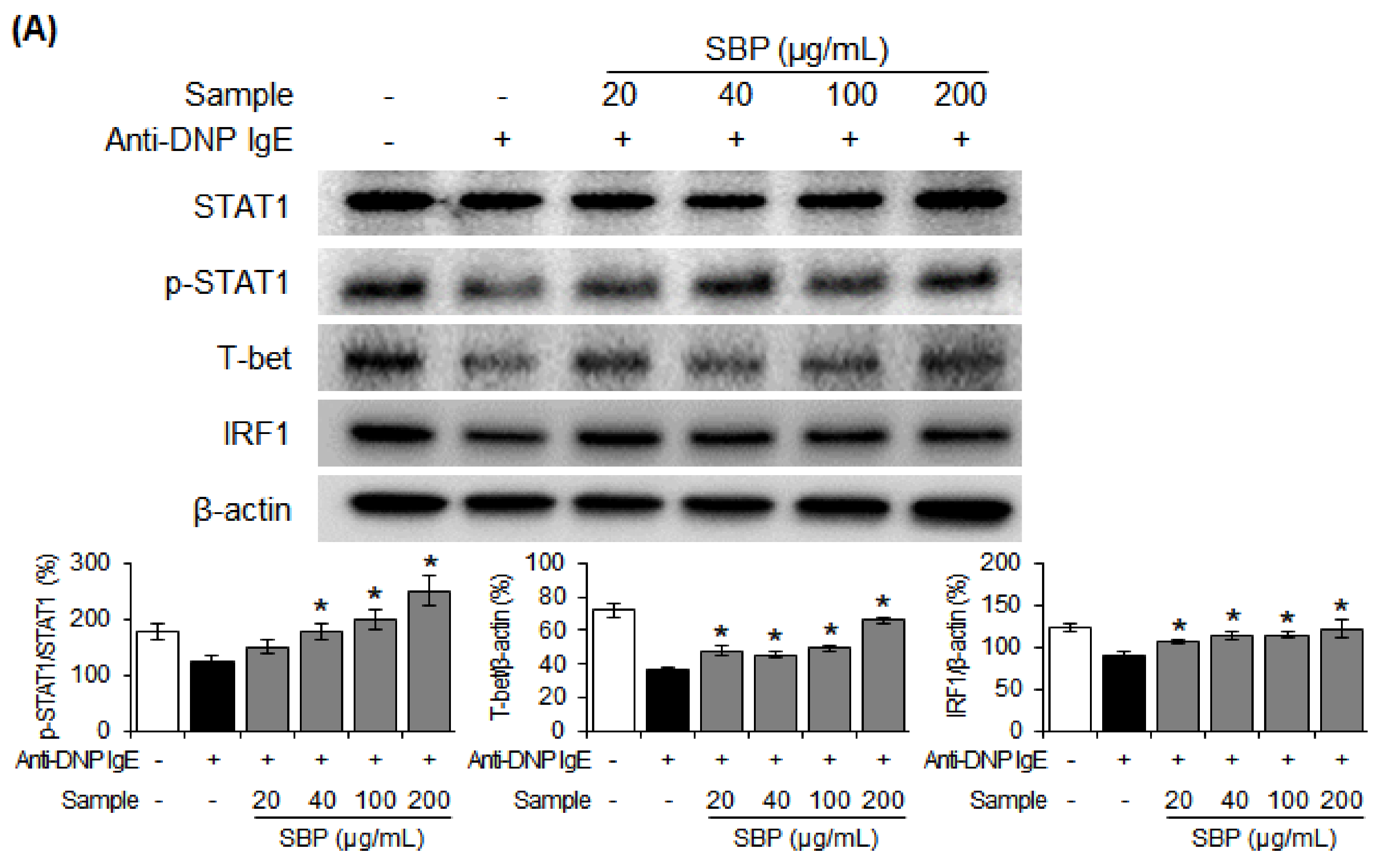
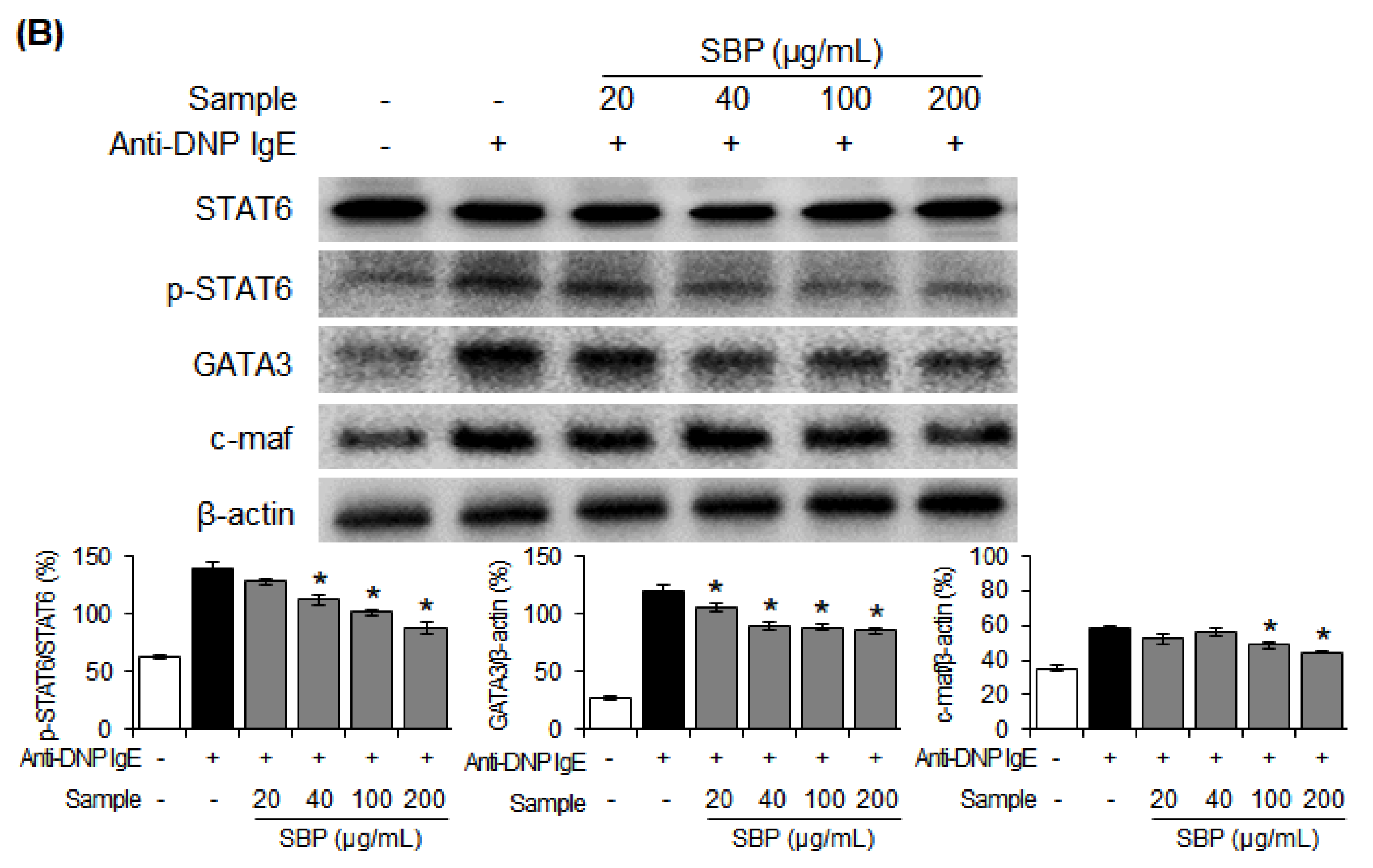
3.8. Effect of SBP Extract on the Protein Expression of Th1/Th2 Differentiation Transcription Factors in RBL-2H3 Cells Stimulated by Anti-DNP IgE
3.9. Effects of the SBP Extract on Body and Tissue Weights of OVA/Alum-Sensitized Mice
3.10. Hepatotoxicity of the SBP Extract in OVA/Alum-Sensitized Mice
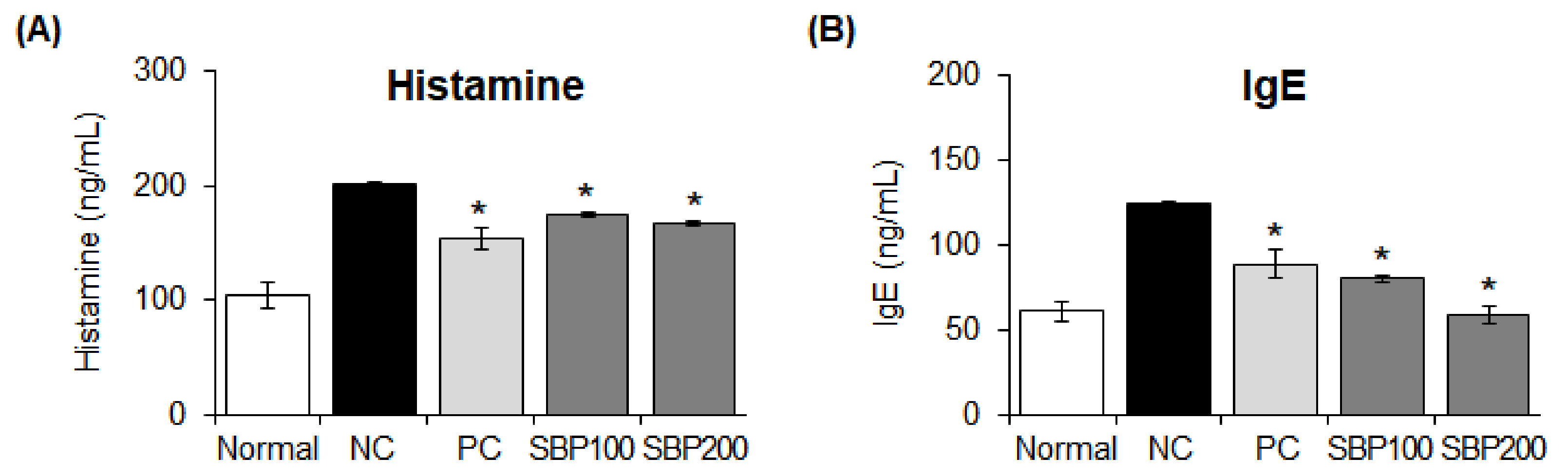
3.11. Effects of the SBP Extract on Allergy Mediators in OVA/Alum-Sensitized Mice
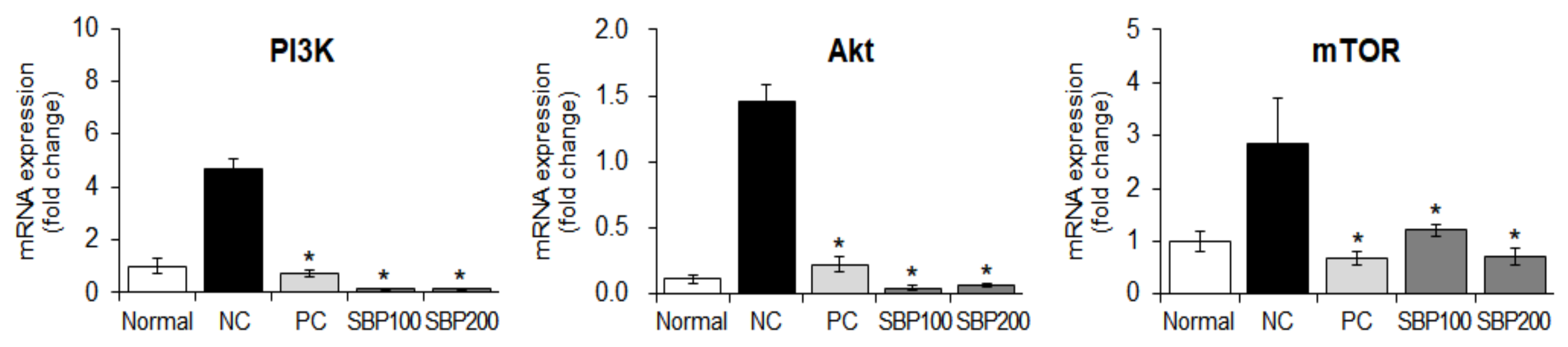
3.12. Effect of the SBP Extract on PI3K/mTOR Signaling in OVA/Alum-Sensitized Mice
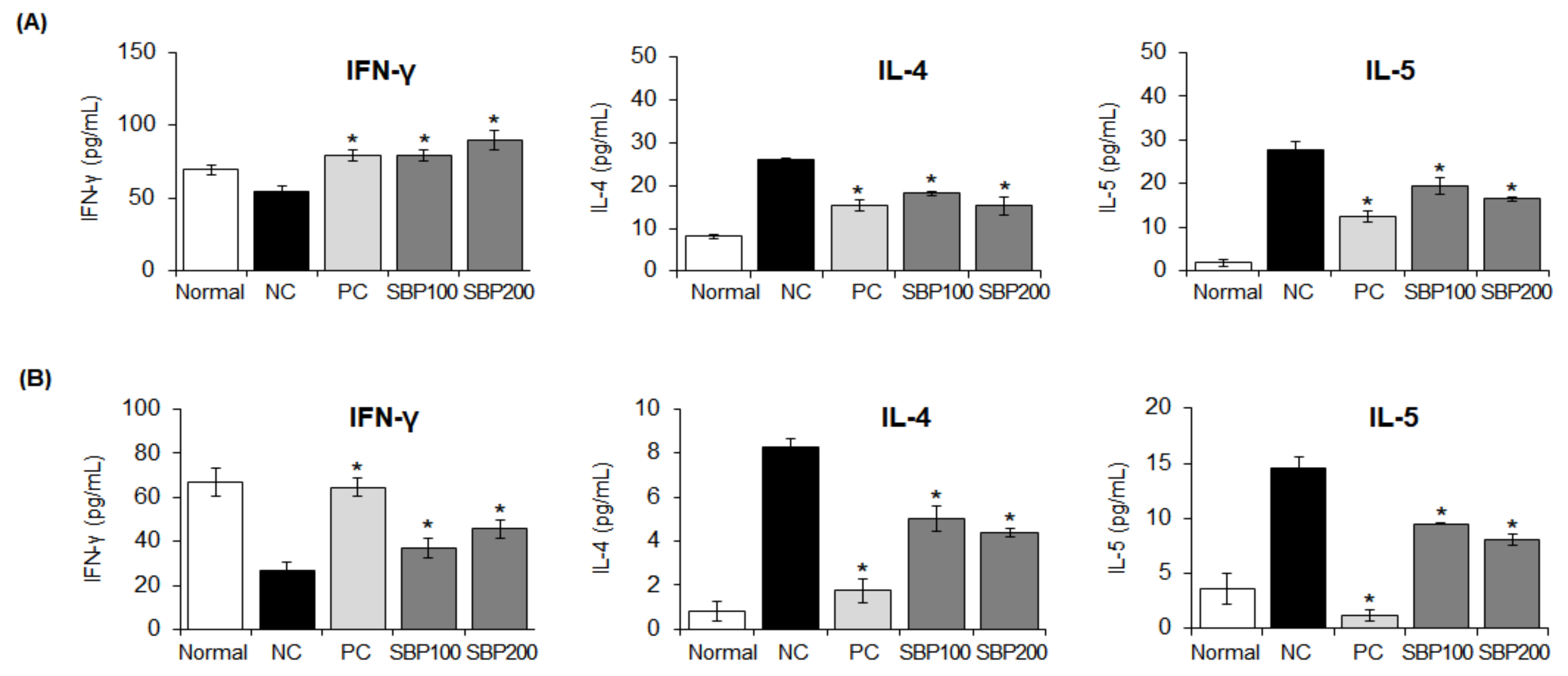
3.13. Effect of the SBP Extract on Cytokine Production in OVA/Alum-Sensitized Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galli, S.J. New Concepts about the Mast Cell. N. Engl. J. Med. 1993, 328, 257–265. [Google Scholar] [CrossRef]
- Kim, S.R.; Heaton, H.; Liu, L.Y.; King, B.A. Rapid Repigmentation of Vitiligo Using Tofacitinib plus Low-Dose, Narrowband uv-b Phototherapy. JAMA Dermatol. 2018, 154, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Youn, D.H.; Kang, J.W.; Ahn, K.S.; Kwak, H.J.; Um, J.Y. Taeumjowi-Tang, a Traditional Korean Sasang Remedy, Improves Obesity-Atopic Dermatitis Comorbidity by Regulating Hypoxia-Inducible Factor 1 Alpha. Front. Pharmacol. 2019, 10, 1458. [Google Scholar] [CrossRef] [Green Version]
- Nauta, A.J.; Engels, F.; Knippels, L.M.; Garssen, J.; Nijkamp, F.P.; Redegeld, F.A. Mechanisms of Allergy and Asthma. Eur. J. Pharmacol. 2008, 585, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Kim, J.Y.; Kang, M.K.; Yang, M.S.; Park, H.W.; Min, K.U.; Cho, S.H.; Kang, H.R. Allergies Are Still on the Rise? A 6-Year Nationwide Population-Based Study in Korea. Allergol. Int. 2016, 65, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainardi, T.; Kapoor, S.; Bielory, L. Complementary and Alternative Medicine: Herbs, Phytochemicals and Vitamins and Their Immunologic Effects. J. Allergy Clin. Immunol. 2009, 123, 283–294. [Google Scholar] [CrossRef]
- Kim, J.M.; Kim, D.J.; Kim, T.H.; Baek, J.M.; Kim, H.S.; Cho, M. Effects of Water Extract of Glycyrrhiza uralensis on β-Hexosaminidase Release and Expression of the Cytokines of RBL-2H3 Mast Cells. Korean J. Med. Crop. Sci. 2010, 18, 231–237. [Google Scholar]
- Kim, S.B.; Kang, B.H.; Kwon, H.S.; Kang, J.H. Anti Inflammatory and Antiallergic Activity of Fermented Turmeric by Lactobacillus johnsonii IDCC 9203. Korean J. Microbiol. Biotechnol. 2011, 39, 266–273. [Google Scholar]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From Growth Signal Integration to Cancer, Diabetes and Ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, X.; Xu, Z.; Hao, L.; Zhang, Y.; Liu, Z. PI3K-AKT-mTOR Signaling Pathway: The Intersection of Allergic Asthma and Cataract. Pharmazie 2019, 74, 598–600. [Google Scholar] [CrossRef]
- Youyou, S.; Chong, L.; Lin, P.; Li, H.; Zhu, L.; Wu, Q.; Li, C. MicroRNA-133a Alleviates Airway Remodeling in Asthma through PI3K/AKT/mTOR Signaling Pathway by Targeting IGF1R. J. Cell. Physiol. 2019, 234, 4068–4080. [Google Scholar] [CrossRef]
- Cheng, H.; Shcherba, M.; Pendurti, G.; Liang, Y.; Piperdi, B.; Perez-Soler, R. Targeting the PI3K/AKT/mTOR Pathway: Potential for Lung Cancer Treatment. Lung Cancer Manag. 2014, 3, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Athari, S.S.; Mehrabi Nasab, E.M.; Zhao, L. PI3K/AKT/mTOR and TLR4/MyD88/NF-κB Signaling Inhibitors Attenuate Pathological Mechanisms of Allergic Asthma. Inflammation 2021, 44, 1895–1907. [Google Scholar] [CrossRef]
- Kapoor, Y.; Kumar, K. Structural and Clinical Impact of Anti-Allergy Agents: An Overview. Bioorg Chem. 2020, 94, 103351. [Google Scholar] [CrossRef]
- Poluzzi, E.; Raschi, E.; Godman, B.; Koci, A.; Moretti, U.; Kalaba, M.; Wettermark, B.; Sturkenboom, M.; Ponti, F.D. Pro-arrhythmic potential of oral antihistamines (H1): Combining adverse event reports with drug utilization data across Europe. PLoS ONE 2015, 10, e0119551. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.L.; Anderson, M.L.; Dublin, S.; Hanlon, J.T.; Hubbard, R.; Walker, R.; Yu, O.; Crane, P.K.; Larson, E.B. Cumulative use of strong anticholinergics and incident dementia: A prospective cohort study. JAMA Intern Med. 2015, 175, 401–407. [Google Scholar] [CrossRef]
- Anti-Inflammatory Effect of Epigallocatechin Gallate in a Mouse Model of Ovalbumin-Induced Allergic Rhinitis. Int. Immunopharmacol. 2017, 49, 102–108. [CrossRef]
- Takemoto, N.; Kamogawa, Y.; Jun Lee, H.; Kurata, H.; Arai, K.I.; O’Garra, A.; Arai, N.; Miyatake, S. Cutting Edge: Chromatin Remodeling at the IL-4/IL-13 Intergenic Regulatory Region for Th2-Specific Cytokine Gene Cluster. J. Immunol. 2000, 165, 6687–6691. [Google Scholar] [CrossRef] [Green Version]
- Mullen, A.C.; High, F.A.; Hutchins, A.S.; Lee, H.W.; Villarino, A.V.; Livingston, D.M.; Kung, A.L.; Cereb, N.; Yao, T.P.; Yang, S.Y.; et al. Role of T-Bet in Commitment of TH1 Cells before IL-12-Dependent Selection. Science 2001, 292, 1907–1910. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Cote-Sierra, J.; Guo, L.; Paul, W.E. GATA-3 Promotes Th2 Responses through Three Different Mechanisms: Induction of Th2 Cytokine Production, Selective Growth of Th2 Cells and Inhibition of Th1 Cell-Specific Factors. Cell Res. 2006, 16, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.B. Sword Bean (Canavalia ensiformis (L.) DC.) Genetic Resources Regenerated for Potential Medical, Nutraceutical and Agricultural Traits. Genet. Resour. Crop Evol. 2007, 54, 586–592. [Google Scholar]
- Joo, S.J.; Choi, K.J.; Kim, K.S.; Lee, J.W.; Park, S.J. Characteristics of Yogurt Prepared with “Jinpum” Bean and Sword Bean (Canavalin gladiata). Korean J. Postharvest Sci. Technol. 2001, 8, 308–312. [Google Scholar]
- Kim, O.K.; Nam, D.E.; You, Y.H.; Jun, W.J.; Lee, J.M. Protective Effect of Canavalia gladiata on Gastric Inflammation Induced by Alcohol Treatment in Rats. J. Korean Soc. Food Sci. Nutr. 2013, 42, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.P.; Yang, Y.S.; Kim, J.H.; Lee, H.H.; Kim, E.S.; Moon, Y.W.; Kim, J.Y.; Chung, J.K. Chemical Properties and DPPH Radical Scavenging Ability of Sword Bean (Canavalia gladiata) Extract. Korean J. Food Sci. Technol. 2012, 44, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.C.; Park, J.U.; Moon, J.H. Anti-Inflammatory Effects of a Mixture of Coffee and Sword Bean Extracts. Korean J. Food Sci. Technol. 2020, 52, 237–243. [Google Scholar]
- Kim, S.H.; Kim, K.H.; Chi, G.Y.; Cho, I.S.; Kim, H.Y.; Lee, Y.C. Enhancing Effect of Canavalia gladiata DC Semen on the Hematopoietic Expansion and Function of Stem Cells. Korea J. Herbol. 2012, 27, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Reddy, Y.N. Protective Effect of Canavalia gladiata (Sword Bean) Fruit Extracts and Its Flavanoidal Contents, against Azathioprine-Induced Toxicity in Hepatocytes of Albino Rats. Toxicol. Environ. Chem. 2014, 96, 474–481. [Google Scholar] [CrossRef]
- Hwang, K.A.; Heo, W.; Hwang, H.J.; Han, B.K.; Song, M.C.; Kim, Y.J. Anti-Inflammatory Effect of Immature Sword Bean Pod (Canavalia gladiata) in Lipopolysaccharide-Induced RAW264.7 Cells. J. Med. Food 2020, 23, 1183–1191. [Google Scholar] [CrossRef]
- Hwang, H.J.; Hwang, Y.J.; Kim, Y.J.; Kim, M.N.; Hwang, K.A. Immature Sword Bean Pods (Canavalia gladiata) Inhibit Adipogenesis in C3H10T1/2 Cells and Mice with High-Fat Diet–Induced Obesity. J. Chin. Med. Assoc. 2022, 85, 67–76. [Google Scholar] [CrossRef]
- Huang, F.; Yamaki, K.; Tong, X.; Fu, L.; Zhang, R.; Cai, Y.; Yanagisawa, R.; Inoue, K.I.; Takano, H.; Yoshino, S. Inhibition of the antigen-induced activation of RBL-2H3 cells by sinomenine. Int. Immunopharmacol. 2008, 8, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and Atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A Key Role in Inflammatory Diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in Inflammatory Cells: Chemoprevention Using Food Phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Garcia Leme, J.; Hamamura, L.; Leite, M.P.; Rocha e Silva, M. Pharmacological Analysis of the Acute Inflammatory Process Induced in the Rat’s Paw by Local Injection of Carrageenin and by Heating. Br. J. Pharmacol. 1973, 48, 88–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theoharides, T.C.; Alysandratos, K.D.; Angelidou, A.; Delivanis, D.A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast Cells and Inflammation. Biochim. Biophys. Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef] [Green Version]
- White, M.V. The Role of Histamine in Allergic Diseases. J. Allergy Clin. Immunol. 1990, 86, 599–605. [Google Scholar] [CrossRef]
- Romagnani, P.; Annunziato, F.; Piccinni, M.P.; Maggi, E.; Romagnani, S. Th1/Th2 cells, their associated molecules and role in pathophysiology. Eur. Cytokine Netw. 2000, 11, 510–511. [Google Scholar]
- Thomson, A.W.; Turnquist, H.R.; Raimondi, G. Immunoregulatory functions of mTOR inhibition. Nat. Rev. Immunol. 2009, 9, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Noval Rivas, M.N.; Chatila, T.A. Regulatory T Cells in Allergic Diseases. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef] [Green Version]
- Shacter, E.; Weitzman, S.A. Chronic Inflammation and Cancer. Oncology 2002, 16, 217–226. [Google Scholar]
- Tuppo, E.E.; Arias, H.R. The Role of Inflammation in Alzheimer’s Disease. Int. J. Biochem. Cell Biol. 2005, 37, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic Inflammation: Importance of NOD2 and NALP3 in Interleukin-1β Generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Han, B.; Liu, S.; Wang, H.; Zhuang, W.; Huang, Y.; Zhang, R. Derp1-modified dendritic cells attenuate allergic inflammation by regulating the development of T helper type1(Th1)/Th2 cells and regulatory T cells in a murine model of allergic rhinitis. Mol. Immunol. 2017, 90, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, J.A. Cytokines and allergic diseases: Clinical aspects. Allergy Asthma Proc. 1998, 19, 337–341. [Google Scholar] [CrossRef]
- Quraishi, S.A.; Davies, M.J.; Craig, T.J. Inflammatory Responses in Allergic Rhinitis: Traditional Approaches and Novel Treatment Strategies. J. Am. Osteopath. Assoc. 2004, 104, S7–S15. [Google Scholar]
- Zhang, Y.; Jing, Y.; Qiao, J.; Luan, B.; Wang, X.; Wang, L.; Song, Z. Activation of the MTOR Signaling Pathway Is Required for Asthma Onset. Sci. Rep. 2017, 7, 4532. [Google Scholar] [CrossRef] [Green Version]
- Tas, S.W.; Maracle, C.X.; Balogh, E.; Szekanecz, Z. Targeting of Proangiogenic Signalling Pathways in Chronic Inflammation. Nat. Rev. Rheumatol. 2016, 12, 111–122. [Google Scholar] [CrossRef]
- Helal, M.G.; Megahed, N.A.; Abd Elhameed, A.G.A. Saxagliptin Mitigates Airway Inflammation in a Mouse Model of Acute Asthma via Modulation of NF-kB and TLR4. Life Sci. 2019, 239, 117017. [Google Scholar] [CrossRef]
- Nakano, T.; Ikeda, M.; Wakugawa, T.; Kashiwada, Y.; Kaminuma, O.; Kitamura, N.; Yabumoto, M.; Fujino, H.; Kitamura, Y.; Fukui, H.; et al. Identification of Pyrogallol from Awa-Tea as an Anti-allergic Compound That Suppresses Nasal Symptoms and IL-9 Gene Expression. J. Med. Investig. 2020, 67, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.J.; Koh, D.J.; Kim, S.H.; Park, S.J.; Ryu, J.H.; Kim, D.G.; Lee, J.Y.; Lee, K.T. Anti-Inflammatory Effects of Sinapic Acid through the Suppression of Inducible Nitric Oxide Synthase, Cyclooxygase-2, and Proinflammatory Cytokines Expressions via Nuclear Factor-kappaB Inactivation. J. Agric. Food Chem. 2008, 56, 10265–10272. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5′→3′) |
|---|---|---|
| mGADPH | Forward | GTT GTC TCC TGC GAC TTC A |
| Reverse | GGT GGT CCA GGG TTT CTT A | |
| miNOS | Forward | GGC AGC CTG TGA GAC CTT TG |
| Reverse | GCA TTG GAA GTG AAG CGT TT | |
| mCOX-2 | Forward | TTG CTG TAC AAG CAG TGG CAA AGG |
| Reverse | TGG GAG GCA CTT GCA TTG CAT TGA | |
| mPI3K | Forward | GAA GTT GCT CTA CCC AGT GTC C |
| Reverse | GAT AGC CGT TCT TTT CAT TTG G | |
| mAkt | Forward | ACT CAT TCC AGA CCC ACG AC |
| Reverse | AGC CCG AAG TCC GTT ATC TT | |
| mmTOR | Forward | TGT GAA CGG AAC ATA CGA CC |
| Reverse | TTG CTT GCC CAT CAG AGT CAG | |
| rGADPH | Forward | CCA CAG TCC ATG CCA TCA C |
| Reverse | TCC ACC ACC CTG TTG CTG TA- | |
| rIFN-γ | Forward | AAT GGC AAC ATC AGG TCG GCC ATC ACT |
| Reverse | GCT GTG TGT GTC ACA GAA GTC TCG AAC TC | |
| rIL-12 | Forward | GGA GAG ACT ATC AAG ATA GT |
| Reverse | ATG GTC AGT AGA CTT TTA CA | |
| rIL-4 | Forward | CGA TGA TGC ACT TGC AGA AA |
| Reverse | TGG AAA TTG GGG TAG GAA GG- | |
| rIL-13 | Forward | AGC ACA GAA AGC ATG ATC CG |
| Reverse | GTT TGC TAC GAC GTG CGC TA | |
| rPI3K | Forward | AAC ACA GAA GAC CAA TAC TC |
| Reverse | TTC GCC ATC TAC CAC TAC | |
| rAkt | Forward | GTG GCA AGA TGT GTA TGA G |
| Reverse | CTG GCT GAG TAG GAG AAC | |
| rmTOR | Forward | GGT GGA CGA GCT CTT TGT CA |
| Reverse | AGG AGC CCT AAC ACT CGG AT |
| Group (n = 8) | Treatment (mg/kg) | Condition |
|---|---|---|
| Normal | PBS | None |
| NC | Maltodextrin (200 mg/kg) | OVA/Alum |
| PC | Dexamethasone (0.5 mg/kg) | |
| SBP100 | SBP low (100 mg/kg) | |
| SBP200 | SBP high (200 mg/kg) |
| Group | Body (g) | Liver (g) | Spleen (g) | Kidney (g) | Heart (g) | Lung (g) |
|---|---|---|---|---|---|---|
| Normal | 26.71 ± 0.47 | 1.509 ± 0.025 * | 0.101 ± 0.003 * | 0.452 ± 0.024 | 0.146 ± 0.004 * | 0.307 ± 0.010 * |
| NC | 25.72 ± 0.37 | 1.269 ± 0.037 | 0.112 ± 0.003 | 0.396 ± 0.019 | 0.131 ± 0.003 | 0.356 ± 0.015 |
| PC | 24.32 ± 0.24 | 1.063 ± 0.049 * | 0.045 ± 0.003 * | 0.335 ± 0.012 | 0.118 ± 0.004 * | 0.225 ± 0.025 * |
| SBP100 | 25.26 ± 0.32 | 1.138 ± 0.054 | 0.107 ± 0.005 | 0.326 ± 0.005 * | 0.119 ± 0.004 | 0.338 ± 0.010 |
| SBP200 | 25.19 ± 0.22 | 1.192 ± 0.015 | 0.113 ± 0.004 | 0.340 ± 0.010 * | 0.119 ± 0.001 * | 0.330 ± 0.010 |
| Group | AST (mU/mL) | ALT (mU/mL) | GSH (μM) | GPx (mU/mL) |
|---|---|---|---|---|
| Normal | 109.5 ± 5.142 | 61.52 ± 11.95 | 10.25 ± 1.750 * | 672.3 ± 44.96 * |
| NC | 118.4 ± 5.860 | 73.56 ± 8.811 | 4.451 ± 1.477 | 450.6 ± 104.2 |
| PC | 110.6 ± 8.071 | 61.94 ± 7.618 | 12.17 ± 0.941 * | 651.1 ± 31.52 * |
| SBP100 | 109.7 ± 5.329 | 64.72 ± 8.760 | 9.414 ± 1.149 * | 478.1 ± 47.16 |
| SBP200 | 108.0 ± 7.734 | 63.48 ± 6.197 | 10.98 ± 1.070 * | 548.6 ± 52.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, K.-A.; Hwang, Y.J.; Hwang, H.-J.; Lee, S.H.; Kim, Y.J. Sword Bean (Canavalia gladiata) Pod Exerts Anti-Allergic and Anti-Inflammatory Effects through Modulation of Th1/Th2 Cell Differentiation. Nutrients 2022, 14, 2853. https://doi.org/10.3390/nu14142853
Hwang K-A, Hwang YJ, Hwang H-J, Lee SH, Kim YJ. Sword Bean (Canavalia gladiata) Pod Exerts Anti-Allergic and Anti-Inflammatory Effects through Modulation of Th1/Th2 Cell Differentiation. Nutrients. 2022; 14(14):2853. https://doi.org/10.3390/nu14142853
Chicago/Turabian StyleHwang, Kyung-A, Yu Jin Hwang, Hye-Jeong Hwang, Sang Hoon Lee, and Young Jun Kim. 2022. "Sword Bean (Canavalia gladiata) Pod Exerts Anti-Allergic and Anti-Inflammatory Effects through Modulation of Th1/Th2 Cell Differentiation" Nutrients 14, no. 14: 2853. https://doi.org/10.3390/nu14142853