
Omega-3 Polyunsaturated Fatty Acid: A Pharmaco-Nutraceutical Approach to Improve the Responsiveness to Ursodeoxycholic Acid

 , and
, and 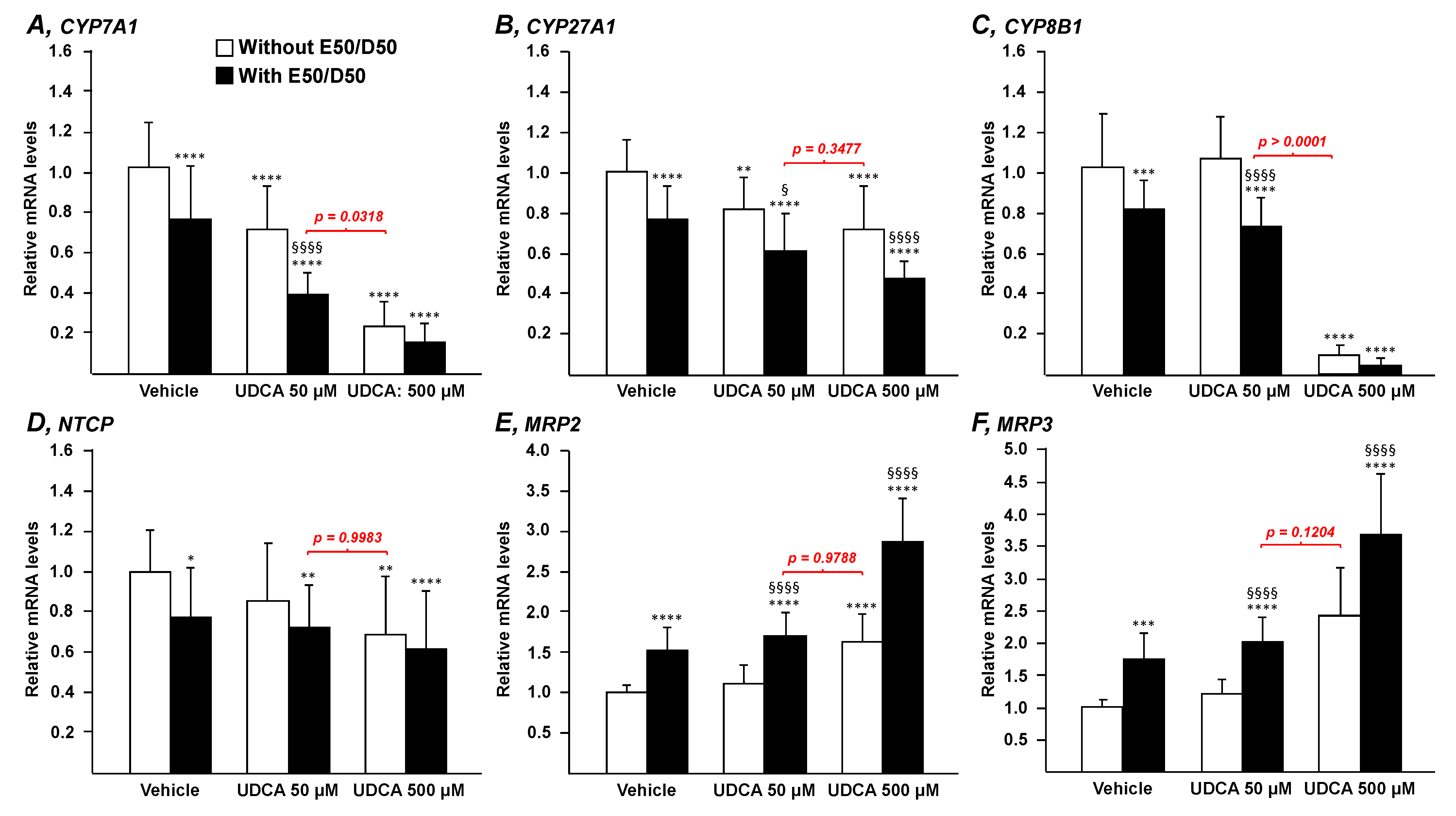{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Isolation, Reverse Transcription and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.3. Caspase-3 Assay
2.4. ELISA
2.5. Statistics
3. Results
3.1. N-3 Polyunsaturated Fatty Acids Improve the Transcriptional Signature of UDCA in HepG2 Cells
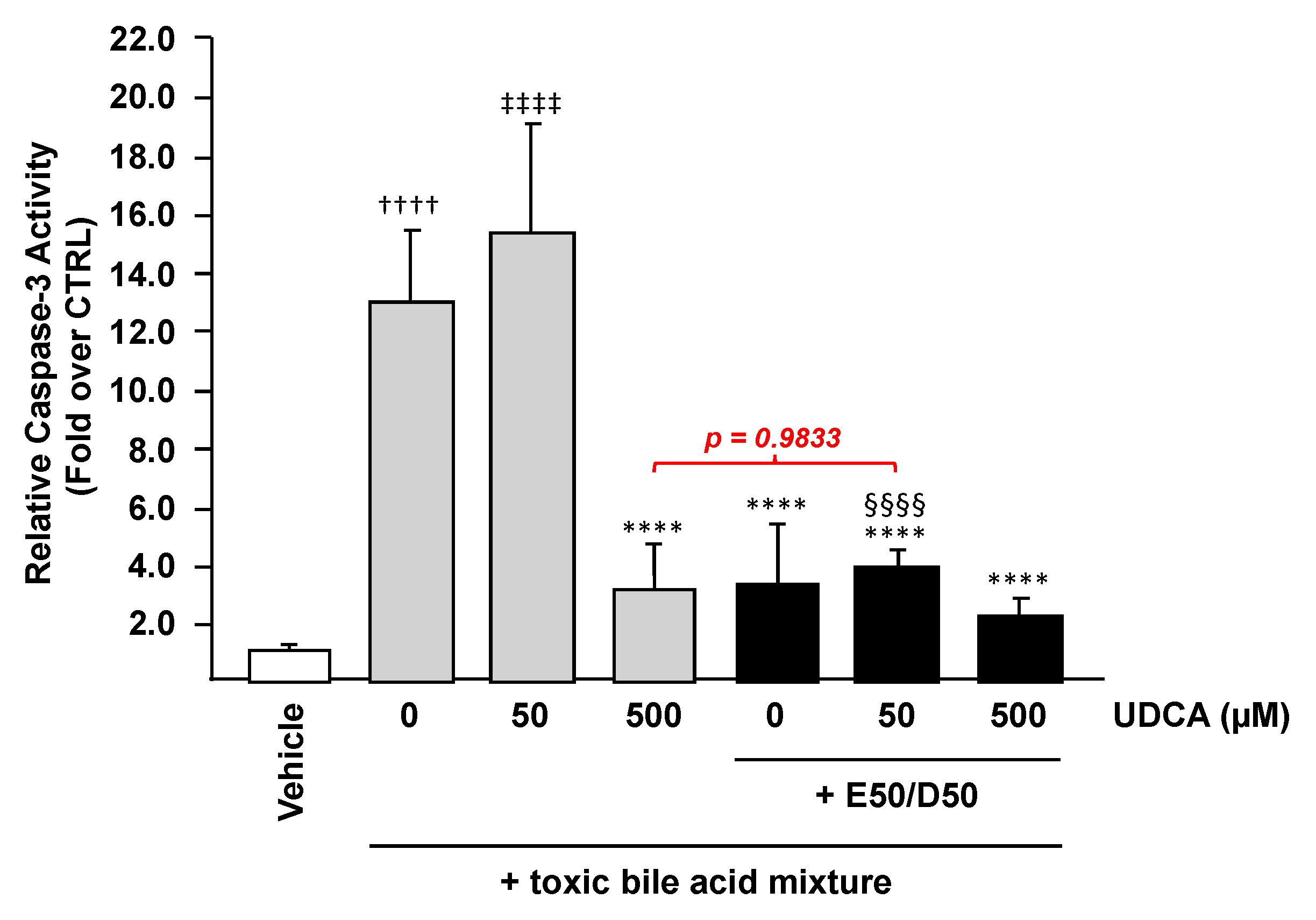
3.2. N-3 Polyunsaturated Fatty Acids Improve the Hepatoprotective Effects of Ursodeoxycholic Acid against BA-Induced Apoptosis in HepG2 Cells
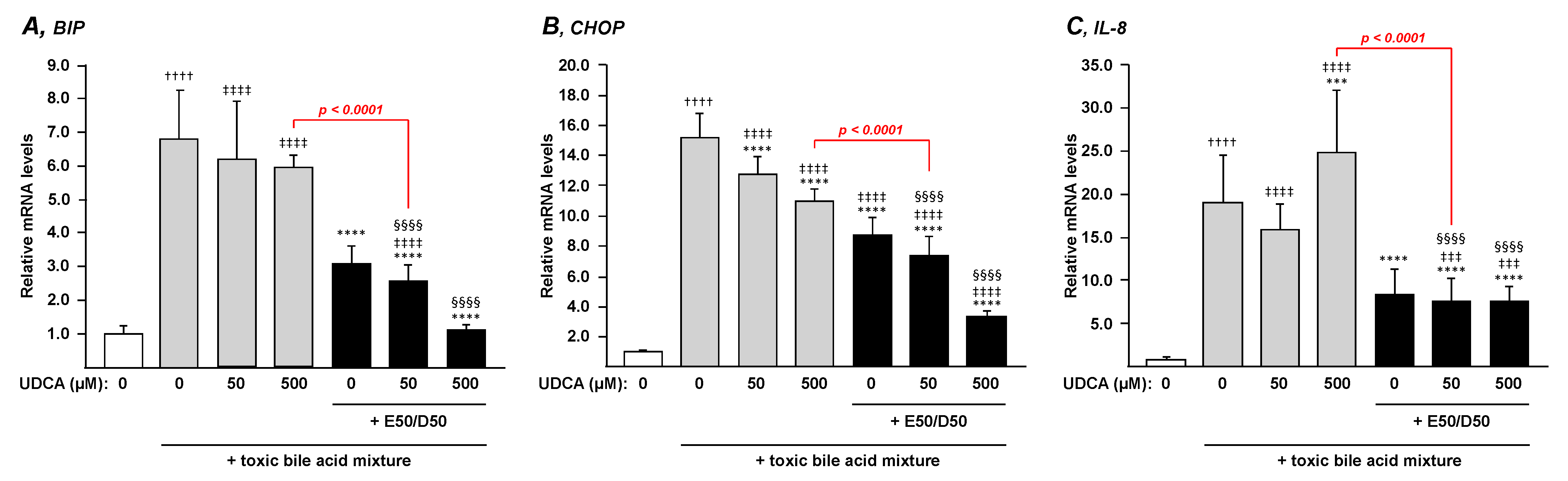
3.3. N-3 Polyunsaturated Fatty Acids Suppress the BA-Induced Endoplasmic Reticulum Stress and Inflammation in HepG2 Cells
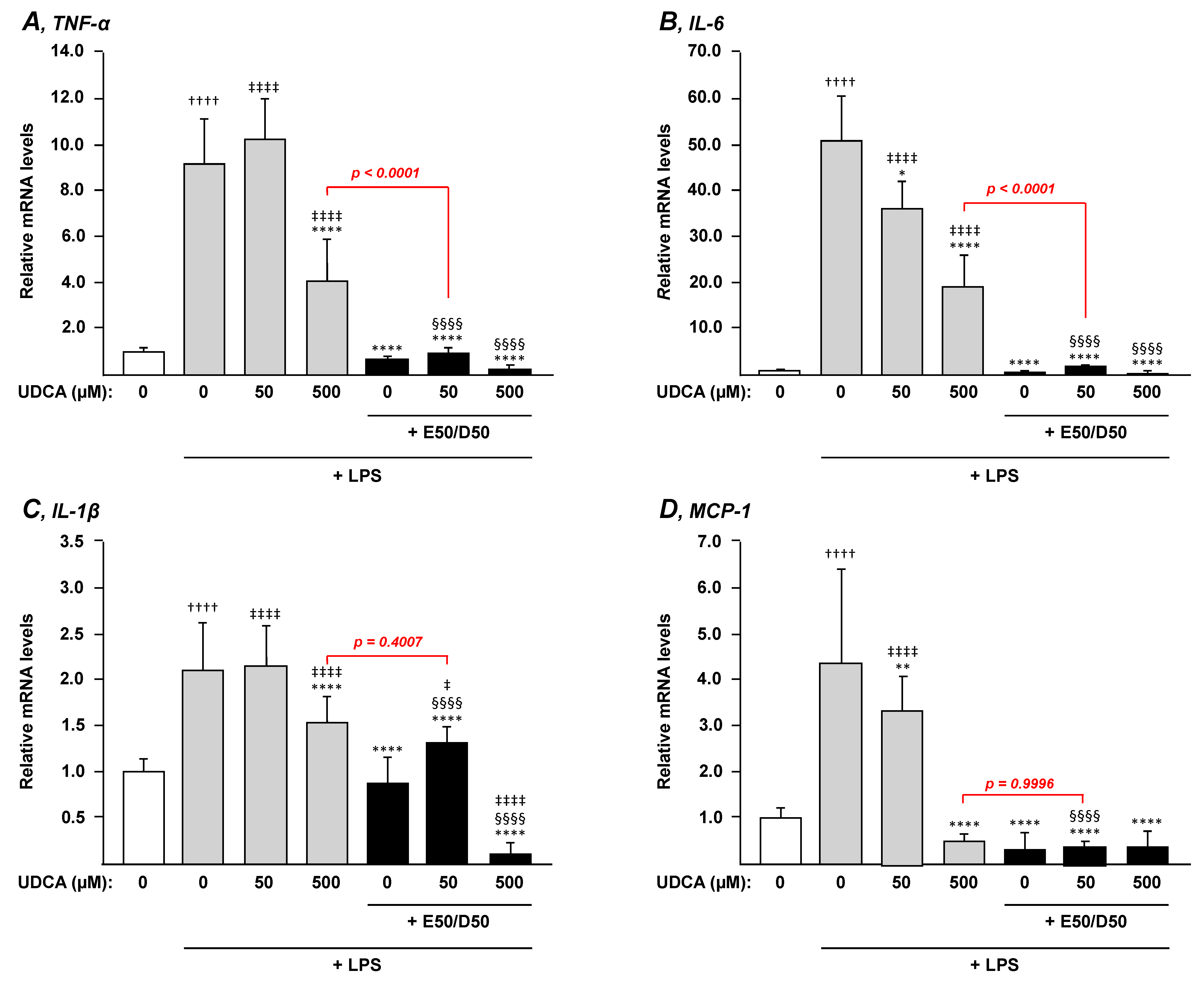
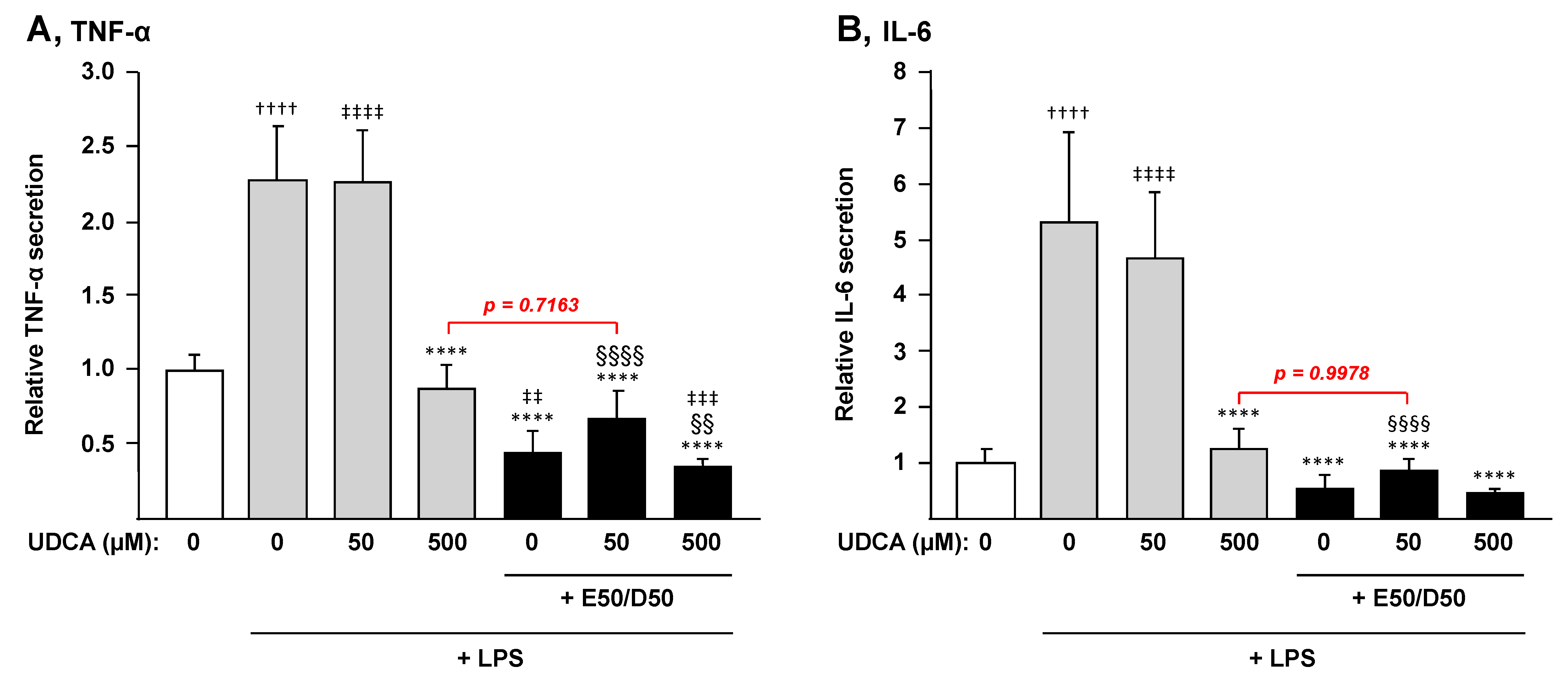
3.4. The Combination n-3 PUFAs Plus UDCA Suppresses the LPS-Dependent Induction of Pro-inflammatory Mediators in THP-1 Macrophages
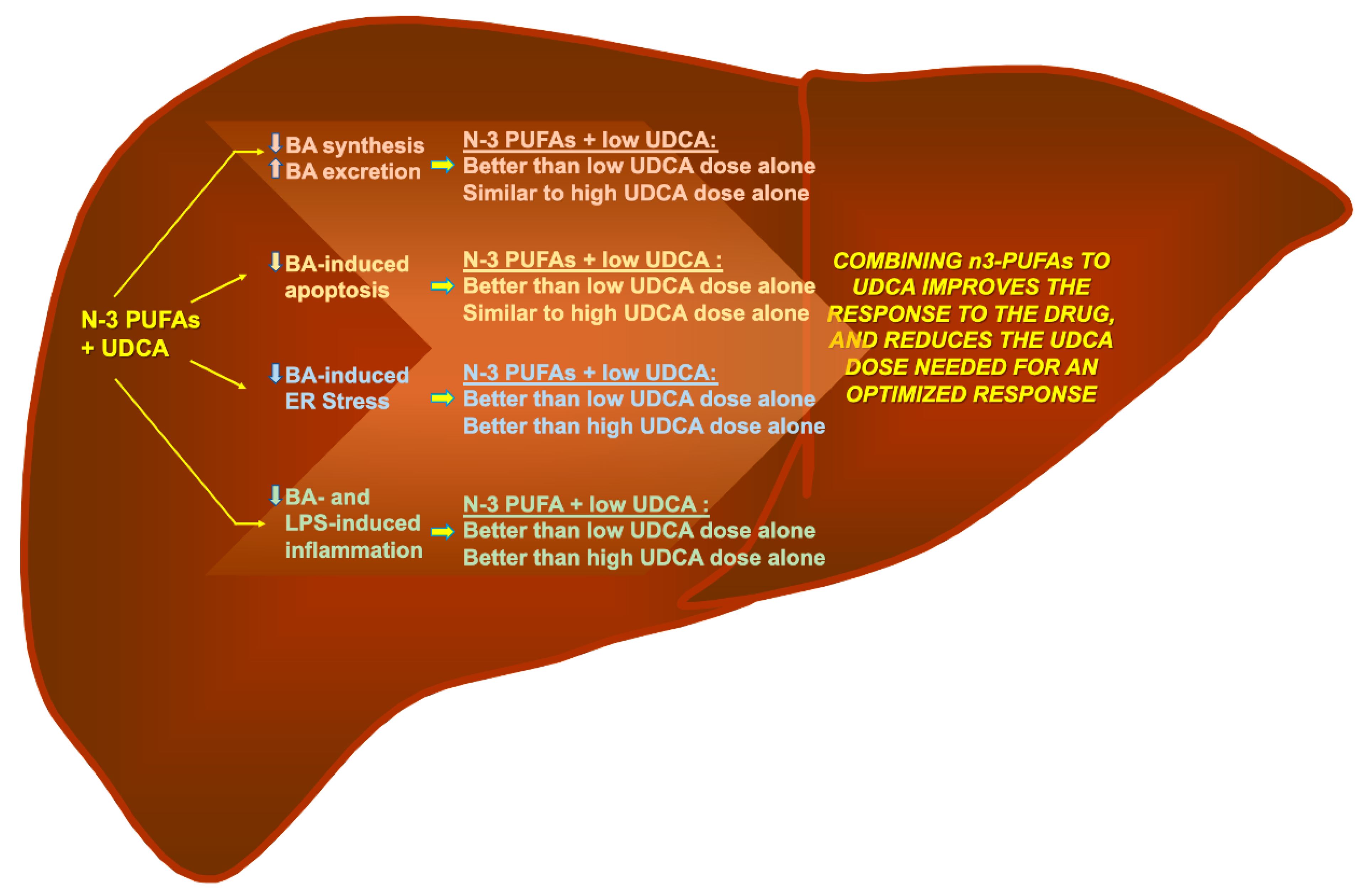
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paumgartner, G.; Beuers, U. Ursodeoxycholic acid in cholestatic liver disease: Mechanisms of action and therapeutic use revisited. Hepatology 2002, 36, 525–531. [Google Scholar] [CrossRef]
- Roma, M.G.; Toledo, F.D.; Boaglio, A.C.; Basiglio, C.L.; Crocenzi, F.A.; Pozzi, E.J. Ursodeoxycholic acid in cholestasis: Linking action mechanisms to therapeutic applications. Clin. Sci. 2011, 121, 523–544. [Google Scholar] [CrossRef] [Green Version]
- Floreani, A.; Mangini, C. Primary biliary cholangitis: Old and novel therapy. Eur. J. Intern. Med. 2018, 47, 1–5. [Google Scholar] [CrossRef]
- Gochanour, E.; Jayasekera, C.; Kowdley, K. Primary Sclerosing Cholangitis: Epidemiology, Genetics, Diagnosis, and Current Management. Clin. Liver Dis. 2020, 15, 125–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.M.P.; Steer, C.J. The therapeutic effects of ursodeoxycholic acid as an anti-apoptotic agent. Expert Opin. Investig. Drugs 2005, 10, 1243–1253. [Google Scholar] [CrossRef]
- Beuers, U. Drug Insight: Mechanisms and sites of action of ursodeoxycholic acid in cholestasis. Nat. Rev. Gastroenterol. Hepatol. 2006, 3, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Lazaridis, K.N.; Gores, G.J.; Lindor, K.D. Ursodeoxycholic acid ‘mechanisms of action and clinical use in hepatobiliary disorders’. J. Hepatol. 2001, 35, 134–146. [Google Scholar] [CrossRef]
- Jazrawi, R.P.; De Caestecker, J.S.; Goggin, P.M.; Britten, A.J.; Joseph, A.E.; Maxwell, J.D.; Northfield, T.C. Kinetics of hepatic bile acid handling in cholestatic liver disease: Effect of ursodeoxycholic acid. Gastroenterology 1994, 106, 134–142. [Google Scholar] [CrossRef]
- Marschall, H.U.; Wagner, M.; Zollner, G.; Fickert, P.; Diczfalusy, U.; Gumhold, J.; Silbert, D.; Fuchsbichler, A.; Benthin, L.; Grundström, R.; et al. Complementary stimulation of hepatobiliary transport and detoxification systems by rifampicin and ursodeoxycholic acid in humans. Gastroenterology 2005, 129, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.; García, N.; Martí-Climent, J.M.; Peñuelas, I.; Richter, J.A.; Medina, J.F. Assessment of biliary bicarbonate secretion in humans by positron emission tomography. Gastroenterology 1999, 117, 167–172. [Google Scholar] [CrossRef]
- Zollner, G.; Fickert, P.; Silbert, D.; Fuchsbichler, A.; Marschall, H.U.; Zatloukal, K.; Denk, H.; Trauner, M. Adaptive changes in hepatobiliary transporter expression in primary biliary cirrhosis. J. Hepatol. 2003, 38, 717–727. [Google Scholar] [CrossRef]
- Calmus, Y.; Gane, P.; Rouger, P.; Poupon, R. Hepatic expression of class I and class II major histocompatibility complex molecules in primary biliary cirrhosis: Effect of ursodeoxycholic acid. Hepatology 1990, 11, 12–15. [Google Scholar] [CrossRef]
- Koga, H.; Sakisaka, S.; Ohishi, M.; Sata, M.; Tanikawa, K. Nuclear DNA fragmentation and expression of Bcl-2 in primary biliary cirrhosis. Hepatology 1997, 25, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.G.; Wolfhagen, F.H.; Verma, A.; Van Buuren, H.R.; Jazrawi, R.P.; Levy, J.H.; Northfield, T.C.; Schalm, S.W. Soluble intercellular adhesion molecule-1 in primary biliary cirrhosis: Effect of ursodeoxycholic acid and immunosuppressive therapy. Eur. J. Gastroenterol. Hepatol. 1997, 9, 155–161. [Google Scholar] [CrossRef]
- Neuman, M.; Angulo, P.; Malkiewicz, I.; Jorgensen, R.; Shear, N.; Dickson, E.R.; Haber, J.; Katz, G.; Lindor, K. Tumor necrosis factor-alpha and transforming growth factor-beta reflect severity of liver damage in primary biliary cirrhosis. J. Gastroenterol. Hepatol. 2002, 17, 196–202. [Google Scholar] [CrossRef]
- Neuman, M.G.; Cameron, R.G.; Haber, J.A.; Katz, G.G.; Blendis, L.M. An electron microscopic and morphometric study of ursodeoxycholic effect in primary biliary cirrhosis. Liver Int. 2002, 22, 235–244. [Google Scholar] [CrossRef]
- Terasaki, S.; Nakanuma, Y.; Ogino, H.; Unoura, M.; Kobayashi, K. Hepatocellular and biliary expression of HLA antigens in primary biliary cirrhosis before and after ursodeoxycholic acid therapy. Am. J. Gastroenterol. 1991, 86, 1194–1199. [Google Scholar] [PubMed]
- Yokomori, H.; Oda, M.; Wakayashi, G.; Kitajima, M.; Ishii, H. Ursodeoxycholic Acid Therapy Attenuated Expression of Adhesion Molecule in Primary Biliary Cirrhosis. Intern. Med. 2003, 42, 1259–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gochanour, E.M.; Kowdley, K.V. Investigational drugs in early phase development for primary biliary cholangitis. Expert. Opin. Investig. Drugs 2021, 30, 131–141. [Google Scholar] [CrossRef]
- Nicoletti, A.; Maurice, J.B.; Thorburn, D. Guideline review: British Society of Gastroenterology/UK-PSC guidelines for the diagnosis and management of primary sclerosing cholangitis. Frontline Gastroenterol. 2021, 12, 62–66. [Google Scholar] [CrossRef]
- Olsson, R.; Boberg, K.M.; De Muckadell, O.S.; Lindgren, S.; Hultcrantz, R.; Folvik, G.; Bell, H.; Gangsøy–Kristiansen, M.; Matre, J.; Rydning, A.; et al. High-dose ursodeoxycholic acid in primary sclerosing cholangitis: A 5-year multicenter, randomized, controlled study. Gastroenterology 2005, 129, 1464–1472. [Google Scholar] [CrossRef] [Green Version]
- Levy, C. Novel Therapies for Cholestatic Liver Disease. Gastroenterol. Hepatol. 2019, 15, 493–496. [Google Scholar]
- Fetterman, J.W.; Zdanowicz, M.M. Therapeutic potential of n-3 polyunsaturated fatty acids in disease. Am. J. Health-Syst. Pharm. 2009, 66, 1169–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Fernández-Galilea, M.; Martínez-Fernández, L.; González-Muniesa, P.; Pérez-Chávez, A.; Martínez, J.A.; Moreno-Aliaga, M.J. Oxidative Stress and Non-Alcoholic Fatty Liver Disease: Effects of Omega-3 Fatty Acid Supplementation. Nutrients 2019, 11, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieślak, A.; Trottier, J.; Verreault, M.; Milkiewicz, P.; Vohl, M.-C.C.; Barbier, O. N-3 Polyunsaturated Fatty Acids Stimulate Bile Acid Detoxification in Human Cell Models. Can. J. Gastroenterol. Hepatol. 2018, 2018, 6031074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohma, R.; Takahashi, M.; Takada, H.; Takada, H.; Kuwayama, H. Protective effect of n-3 polyunsaturated fatty acid on primary culture of rat hepatocytes. J. Gastroenterol. Hepatol. 2007, 22, 1965–1970. [Google Scholar] [CrossRef]
- Tillman, E.M.; Helms, R.A.; Black, D.D. Eicosapentaenoic acid and docosahexaenoic acid synergistically attenuate bile acid-induced hepatocellular apoptosis. J. Parenter. Enter. Nutr. 2012, 36, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Olsson, R.; Boberg, K.M.; De Muckadell, O.S.; Lindgren, S.; Hultcrantz, R.; Folvik, G.; Bell, H.; Gangsøy–Kristiansen, M.; Matre, J.; Rydning, A.; et al. The safety and efficacy of oral docosahexaenoic acid supplementation for the treatment of primary sclerosing cholangitis—A pilot study. Aliment. Pharmacol. Ther. 2012, 35, 255–265. [Google Scholar]
- Poulsen, R.C.; Moughan, P.J.; Kruger, M.C. Docosahexaenoic acid and 17 beta-estradiol co-treatment is more effective than 17 beta-estradiol alone in maintaining bone post-ovariectomy. Exp. Biol. Med. 2008, 233, 592–602. [Google Scholar] [CrossRef]
- Zhu, S.; Lin, G.; Song, C.; Wu, Y.; Feng, N.; Chen, W.; He, Z.; Chen, Y.Q. RA and omega-3 PUFA co-treatment activates autophagy in cancer cells. Oncotarget 2017, 8, 109135. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.-J.J.; Briz, O. Bile-acid-induced cell injury and protection. World J. Gastroenterol. 2009, 15, 1677–1689. [Google Scholar] [CrossRef]
- Bochkis, I.M.; Rubins, N.E.; White, P.; Furth, E.E.; Friedman, J.R.; Kaestner, K.H. Hepatocyte-specific ablation of Foxa2 alters bile acid homeostasis and results in endoplasmic reticulum stress. Nat. Med. 2008, 14, 828–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mencin, A.; Seki, E.; Osawa, Y.; Kodama, Y.; De Minicis, S.; Knowles, M.; Brenner, D.A. Alpha-1 antitrypsin Z protein (PiZ) increases hepatic fibrosis in a murine model of cholestasis. Hepatology 2007, 46, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Yoshimura-Miyakoshi, M.; Sato, Y.; Nakanuma, Y. A possible involvement of endoplasmic reticulum stress in biliary epithelial autophagy and senescence in primary biliary cirrhosis. J. Gastroenterol. 2015, 50, 984–995. [Google Scholar] [CrossRef]
- Tamaki, N.; Hatano, E.; Taura, K.; Tada, M.; Kodama, Y.; Nitta, T.; Iwaisako, K.; Seo, S.; Nakajima, A.; Ikai, I.; et al. CHOP deficiency attenuates cholestasis-induced liver fibrosis by reduction of hepatocyte injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindor, K.D.; Kowdley, K.V.; Luketic, V.A.; Harrison, M.E.; McCashland, T.; Befeler, A.S.; Harnois, D.; Jorgensen, R.; Petz, J.; Keach, J.; et al. High-dose ursodeoxycholic acid for the treatment of primary sclerosing cholangitis. Hepatology 2009, 50, 808–814. [Google Scholar] [CrossRef]
- Chascsa, D.M.H.; Lindor, K.D. Emerging therapies for PBC. J. Gastroenterol. 2020, 55, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Rautiainen, H.; Kärkkäinen, P.; Karvonen, A.L.; Nurmi, H.; Pikkarainen, P.; Nuutinen, H.; Färkkilä, M. Budesonide combined with UDCA to improve liver histology in primary biliary cirrhosis: A three-year randomized trial. Hepatology 2005, 41, 747–752. [Google Scholar] [CrossRef]
- Iravani, S.; Dooghaie-Moghadam, A.; Razavi-Khorasani, N.; Moazzami, B.; Beirami, A.D.; Mansour-Ghanaei, A.; Majidzadeh, A.K.; Mehrvar, A.; Khoshdel, A.; Toosi, M.N.; et al. An update on treatment options for primary sclerosing cholangitis. Gastroenterol. Hepatol. Bed Bench. 2020, 13, 115–124. [Google Scholar]
- Barbier, O.; Torra, I.P.; Duguay, Y.; Blanquart, C.; Fruchart, J.C.; Glineur, C.; Staels, B. Pleiotropic actions of peroxisome proliferator-activated receptors in lipid metabolism and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Ghonem, N.S.; Auclair, A.M.; Hemme, C.L.; Gallucci, G.M.; De la Rosa Rodriguez, R.; Boyer, J.L.; Assis, D.N. Fenofibrate Improves Liver Function and Reduces the Toxicity of the Bile Acid Pool in Patients with Primary Biliary Cholangitis and Primary Sclerosing Cholangitis Who Are Partial Responders to Ursodiol. Clin. Pharmacol. Ther. 2020, 108, 1213–1223. [Google Scholar] [CrossRef]
- Corpechot, C.; Chazouillères, O.; Rousseau, A.; Le Gruyer, A.; Habersetzer, F.; Mathurin, P.; Goria, O.; Potier, P.; Minello, A.; Silvain, C.; et al. A Placebo-Controlled Trial of Bezafibrate in Primary Biliary Cholangitis. N. Engl. J. Med. 2018, 378, 2171–2181. [Google Scholar] [CrossRef]
- Cheung, A.C.; Lapointe-Shaw, L.; Kowgier, M.; Meza-Cardona, J.; Hirschfield, G.M.; Janssen, H.L.A.; Feld, J.J. Combined ursodeoxycholic acid (UDCA) and fenofibrate in primary biliary cholangitis patients with incomplete UDCA response may improve outcomes. Aliment. Pharmacol. Ther. 2016, 43, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Caron-Dorval, D.; Verreault, M.; Couture, P.; Deshaies, Y.; Barbier, O.; Vohl, M.C. PPARalpha L162V polymorphism alters the potential of n-3 fatty acids to increase lipoprotein lipase activity. Mol. Nutr. Food Res. 2010, 54, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Trottier, J.; Milkiewicz, P.; Kaeding, J.; Verreault, M.; Barbier, O. Coordinate regulation of hepatic bile acid oxidation and conjugation by nuclear receptors. Mol. Pharm. 2006, 3, 212–222. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Guenard, F.; Barbier, O.; Vohl, M.C. Effect of different concentrations of omega-3 fatty acids on stimulated THP-1 macrophages. Genes Nutr. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.I.; Fang, S.; Kim, K.P.; Ko, Y.; Kim, H.; Oh, J.; Hong, G.Y.; Lee, S.Y.; Kim, J.M.; Noh, I.; et al. Combination treatment with n-3 polyunsaturated fatty acids and ursodeoxycholic acid dissolves cholesterol gallstones in mice. Sci. Rep. 2019, 9, 12740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Lee, K.S.; Lee, D.K.; Lee, S.Y.; Chang, H.Y.; Choi, J.; Lee, J.I. Omega-3 polyunsaturated fatty acid and ursodeoxycholic acid have an additive effect in attenuating diet-induced nonalcoholic steatohepatitis in mice. Exp. Mol. Med. 2014, 46, e127. [Google Scholar] [CrossRef] [Green Version]
- Uppala, J.; Gani, A.R.; Ramaiah, K.V.A. Chemical chaperone, TUDCA unlike PBA, mitigates protein aggregation efficiently and resists ER and non-ER stress induced HepG2 cell death. Sci. Rep. 2017, 7, 3831. [Google Scholar] [CrossRef]
- Xie, Q.; Khaoustov, V.I.; Chung, C.C.; Sohn, J.; Krishnan, B.; Lewis, D.E.; Yoffe, B. Effect of tauroursodeoxycholic acid on endoplasmic reticulum stress–induced caspase-12 activation. Hepatology 2002, 36, 592–601. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thérien, A.; Cieślak, A.; Verreault, M.; Perreault, M.; Trottier, J.; Gobeil, S.; Vohl, M.-C.; Barbier, O. Omega-3 Polyunsaturated Fatty Acid: A Pharmaco-Nutraceutical Approach to Improve the Responsiveness to Ursodeoxycholic Acid. Nutrients 2021, 13, 2617. https://doi.org/10.3390/nu13082617
Thérien A, Cieślak A, Verreault M, Perreault M, Trottier J, Gobeil S, Vohl M-C, Barbier O. Omega-3 Polyunsaturated Fatty Acid: A Pharmaco-Nutraceutical Approach to Improve the Responsiveness to Ursodeoxycholic Acid. Nutrients. 2021; 13(8):2617. https://doi.org/10.3390/nu13082617
Chicago/Turabian StyleThérien, Ariane, Anna Cieślak, Mélanie Verreault, Martin Perreault, Jocelyn Trottier, Stéphane Gobeil, Marie-Claude Vohl, and Olivier Barbier. 2021. "Omega-3 Polyunsaturated Fatty Acid: A Pharmaco-Nutraceutical Approach to Improve the Responsiveness to Ursodeoxycholic Acid" Nutrients 13, no. 8: 2617. https://doi.org/10.3390/nu13082617