Obesity and Age-Related Changes in the Brain of the Zucker Lepr fa/fa Rats

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Behavioral Tests, and Tissue Treatment
2.2. Western Blot Analysis
2.3. Immunohistochemistry
2.4. Morphological Analysis
2.5. Statistical Analysis
3. Results
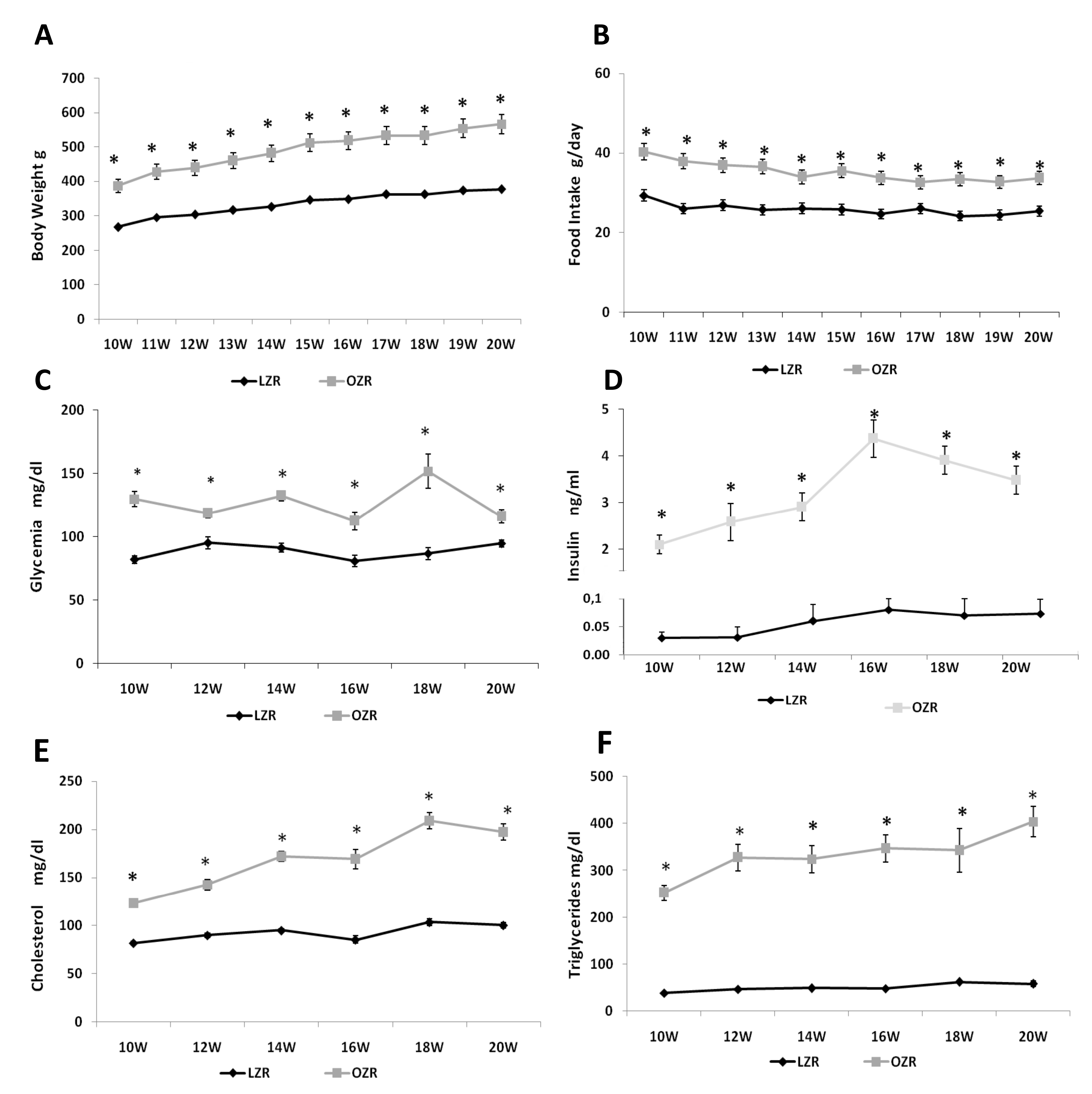
3.1. General and Blood Analysis Indicate a Condition of Dismetabolis Similar to MetS
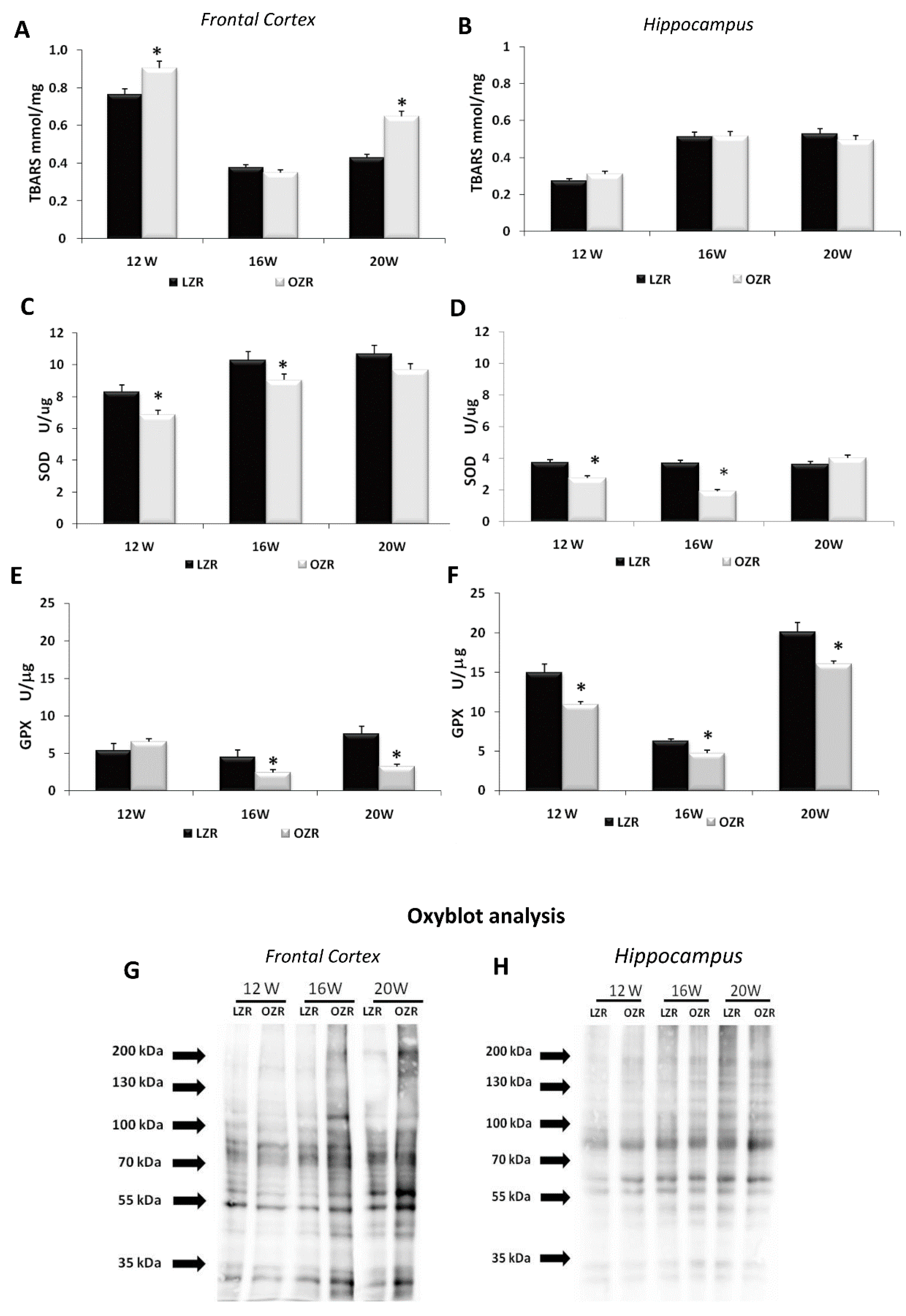
3.2. Oxidative Stress Conditions in Brain Areas
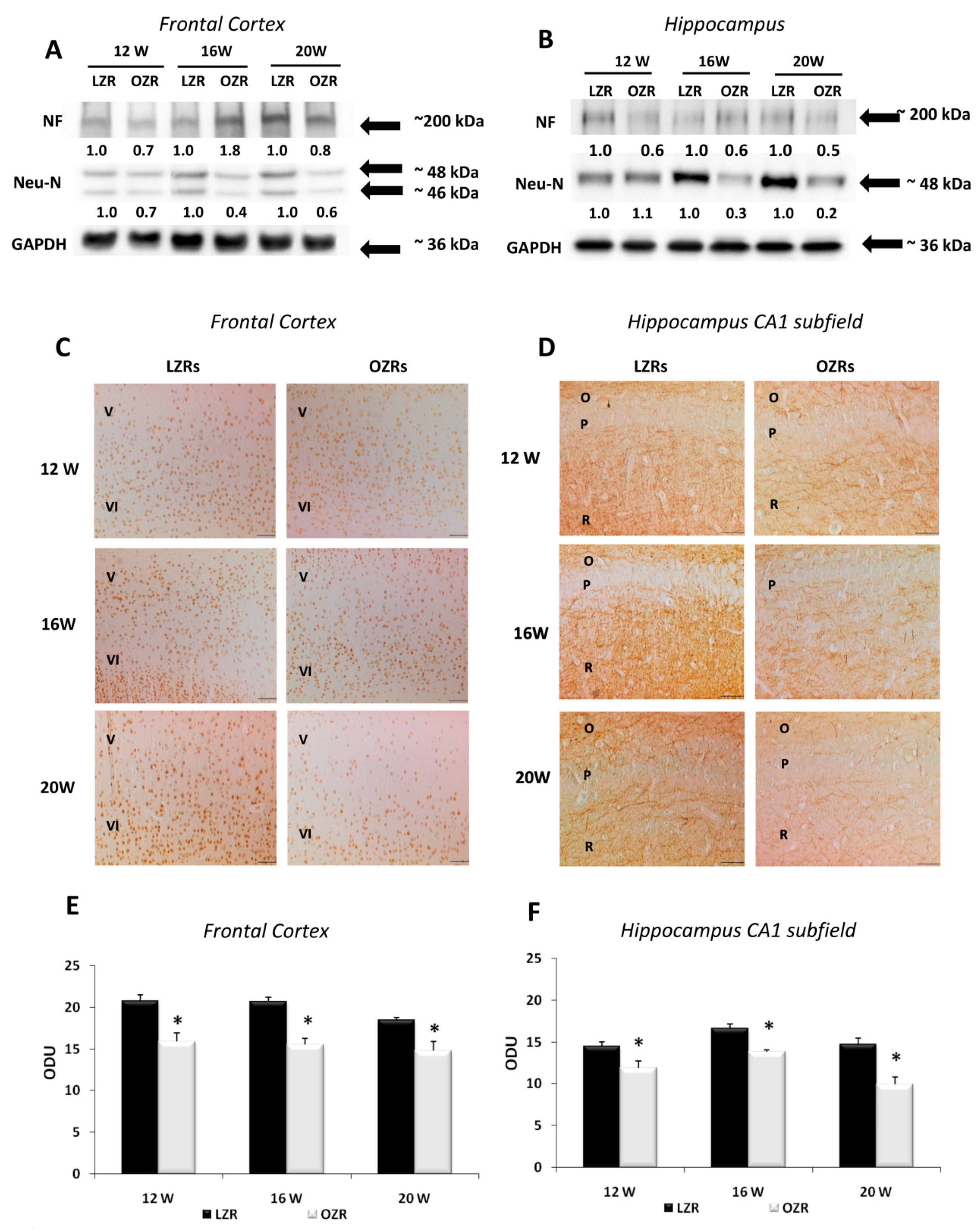
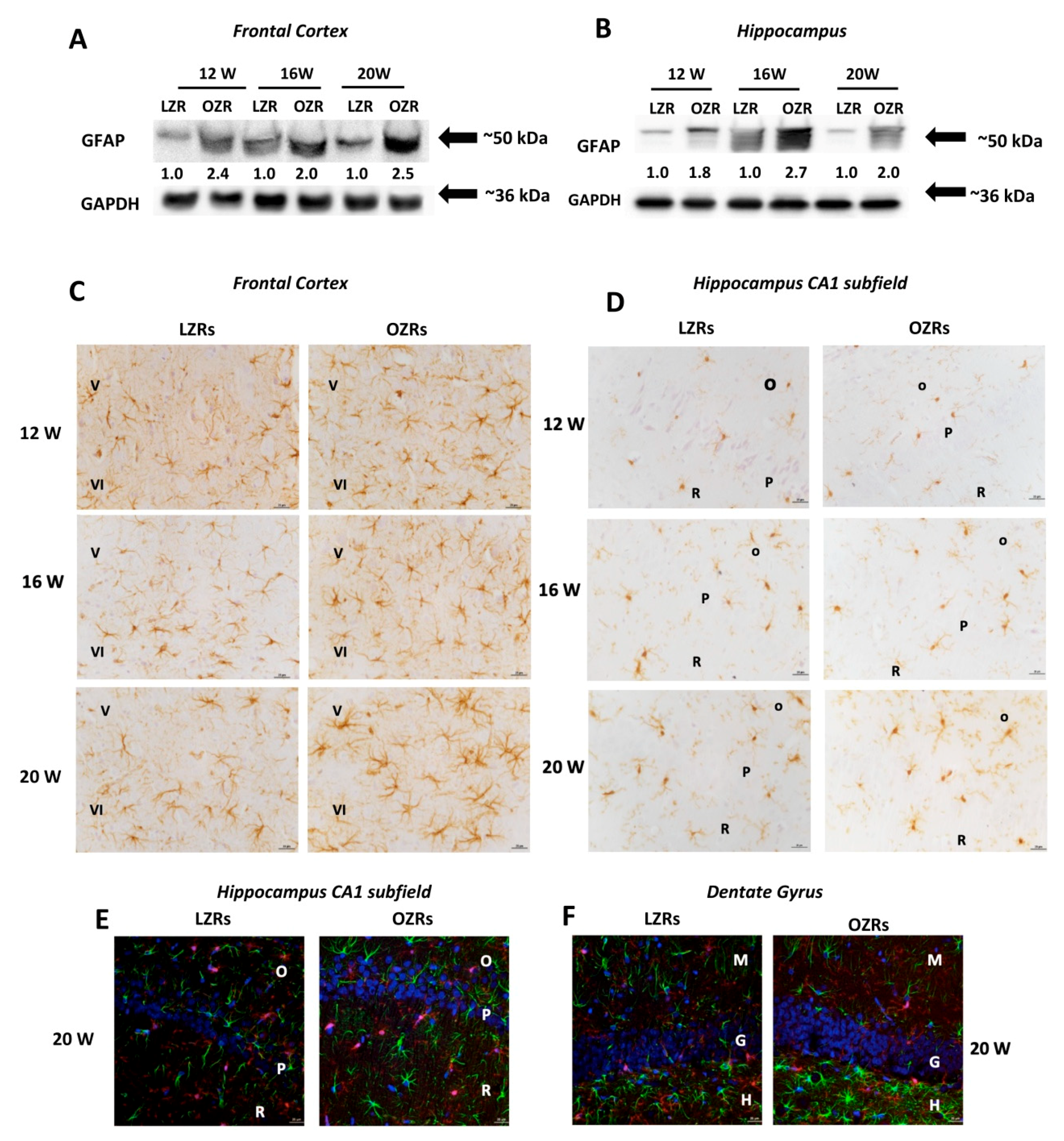
3.3. Neuronal and Glial Markers Analysis Highlighted Neurodegeneration and Gliosis in Obese Rats
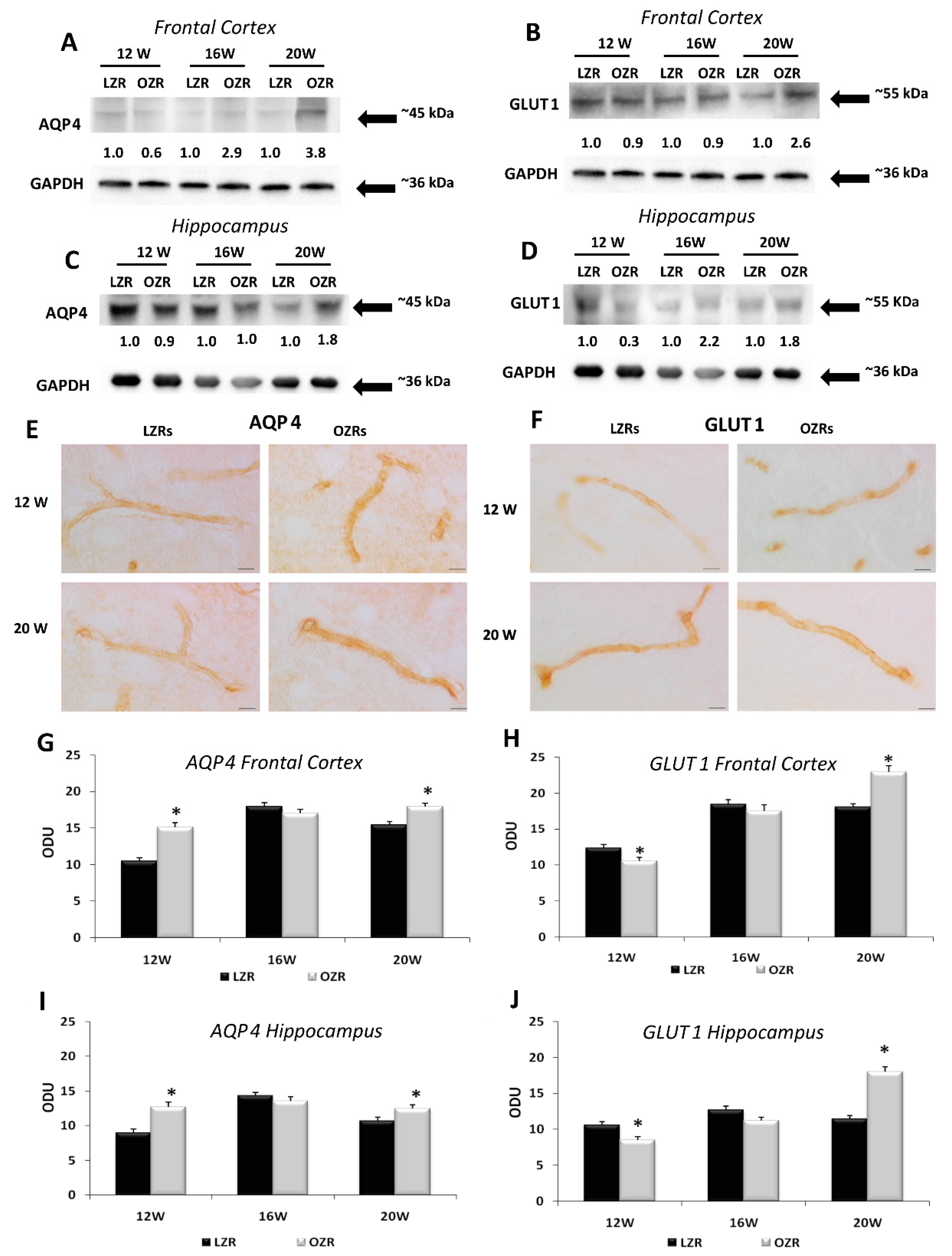
3.4. Analysis of Blood–Brain Barrier Markers Revealed an Impairment of BBB in Obese Rats
3.5. Behavioral Analysis: Anxiety-Like Behavior and Cognitive Impairment in Obese Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AQP4 | aquaporin-4 |
| BBB | blood brain barrier |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GFAP | glial fibrillary acid protein |
| GLUT1 | glucose transporter protein-1 |
| Iba-1 | ionized calcium binding adaptor molecule 1 |
| LZRs | lean Zucker rats |
| MetS | metabolic syndrome |
| NF | neurofilament-200 KDa |
| OZRs | obese Zucker rats |
References
- Arshad, N.; Lin, T.S.; Yahaya, M.F. Metabolic syndrome and its effect on the brain: Possible mechanism. CNS Neurol. Disord. Drug Targets 2018, 17, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Ricci, G.; Pirillo, I.; Tomassoni, D.; Sirignano, A.; Grappasonni, I. Metabolic syndrome, hypertension, and nervous system injury: Epidemiological correlates. Clin. Exp. Hypertens. 2017, 39, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. American Heart Association; Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism.Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss: An update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Ford, E.S.; McGuire, L.C.; Mokdad, A.H. Increasing trends in waist circumference and abdominal obesity among US adults. Obesity 2007, 15, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinkohl, I.; Janke, J.; Hadzidiakos, D.; Slooter, A.; Winterer, G.; Spies, C.; Pischon, T. Associations of the metabolic syndrome and its components with cognitive impairment in older adults. BMC Geriatr. 2019, 19, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization: Obesity and Overweight; WHO: Geneva, Switzerland, 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 27 September 2019).
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Balkau, B.; Charles, M.A. Comment on the provisional report from the WHO consultation. European Group for the Study of Insulin Resistance (EGIR). Diabet. Med. 1999, 16, 442–443. [Google Scholar] [CrossRef]
- Emerging Risk Factors Collaboration; Wormser, D.; Kaptoge, S.; Di Angelantonio, E.; Wood, A.M.; Pennells, L.; Thompson, A.; Sarwar, N.; Kizer, J.R.; Lawlor, D.A.; et al. Separate and combined associations of body-mass index and abdominal adiposity with cardiovascular disease: Collaborative analysis of 58 prospective studies. Lancet 2011, 377, 1085–1095. [Google Scholar] [CrossRef] [Green Version]
- Flegal, K.M.; Kit, B.K.; Orpana, H.; Graubard, B.I. Association of all-cause mortality with overweight and obesity using standard body mass index categories: A systematic review and meta-analysis. JAMA 2013, 309, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Sripetchwandee, J.; Chattipakorn, N.; Chattipakorn, S.C. Links between obesity-induced brain insulin resistance, brain mitochondrial dysfunction, and dementia. Front. Endocrinol. 2018, 9, 496. [Google Scholar] [CrossRef]
- Anjum, I.; Fayyaz, M.; Wajid, A.; Sohail, W.; Ali, A. Does obesity increase the risk of dementia: A Literature Review. Cureus 2018, 10, e2660. [Google Scholar] [CrossRef] [Green Version]
- Milionis, H.J.; Florentin, M.; Giannopoulos, S. Metabolic syndrome and Alzheimer’s disease: A link to a vascular hypothesis? CNS Spectr. 2008, 13, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Lo, R.Y.; Cole, D.; Ducharme, S.; Chen, D.G.; Mapstone, M.; Porsteinsson, A. Alzheimer’s Disease Neuroimaging Initiative. Longitudinal effects of metabolic syndrome on Alzheimer and vascular related brain pathology. Dement. Geriatr. Cogn. Dis. Extra 2014, 4, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Etchegoyen, M.; Nobile, M.H.; Baez, F.; Posesorski, B.; González, J.; Lago, N.; Milei, J.; Otero-Losada, M. Metabolic Syndrome and Neuroprotection. Front. Neurosci. 2018, 20, 196. [Google Scholar] [CrossRef]
- Van Dyken, P.; Lacoste, B. Impact of Metabolic Syndrome on Neuroinflammation and the Blood-Brain Barrier. Front. Neurosci. 2018, 12, 930. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef] [PubMed]
- Maes, H.H.; Neale, M.C.; Eaves, L.J. Genetic and environmental factors in relative body weight and human adiposity. Behav. Genet. 1997, 27, 325–351. [Google Scholar] [CrossRef]
- van Praag, H.; Fleshner, M.; Schwartz, M.W.; Mattson, M.P. Exercise, energy intake, glucose homeostasis, and the brain. J. Neurosci. 2014, 34, 15139–15149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Freire, D.; Knable, L.; Zhao, W.; Gong, B.; Mazzola, P.; Ho, L.; Levine, S.; Pasinetti, G.M. Childhood and adolescent obesity and long-term cognitive consequences during aging. J. Comp. Neurol. 2015, 523, 757–768. [Google Scholar] [CrossRef]
- Aleixandre de Artiñano, A.; Miguel Castro, M. Experimental rat models to study the metabolic syndrome. Br. J. Nutr. 2009, 102, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Yorek, M.A. Alternatives to the Streptozotocin-Diabetic Rodent. Int. Rev. Neurobiol. 2016, 127, 89–112. [Google Scholar] [CrossRef] [Green Version]
- Olsson, I.A.S.; Westlund, K. More than numbers matter: The effect of social factors on behaviour and welfare of laboratory rodents and non-human primates. Appl. Anim. Behav. Sci. 2007, 103, 229–254. [Google Scholar] [CrossRef]
- Gonder, J.C.; Laber, K. Renewed Look at Laboratory Rodent Housing and Management. ILAR J. 2007, 48, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loo, P.L.P.; Mol, J.A.; Koolhaas, J.M.; van Zutphen, L.F.M.; Baumans, V. Modulation of aggression in male mice: Influence of cage cleaning regime and scent-marking. Physiol. Behav. 2001, 5, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Krohn, T.C.; Sørensen, D.B.; Ottesen, J.L.; Hansen, A.K. The effects of individual housing on mice and rats: A review. Anim. Welf. 2006, 15, 343–352. [Google Scholar]
- Georgsson, L.; Barrett, J.; Gietzen, D. The effects of group-housing and relative weight on feeding behaviour in rats. Scand. J. Lab. Animsci. 2001, 28, 201–209. [Google Scholar]
- Tomassoni, D.; Nwankwo, I.E.; Gabrielli, M.G.; Bhatt, S.; Muhammad, A.B.; Lokhandwala, M.F.; Tayebati, S.K.; Amenta, F. Astrogliosis in the brain of obese Zucker rat: A model of metabolic syndrome. Neurosci. Lett. 2013, 543, 136–141. [Google Scholar] [CrossRef]
- Brooks, S.D.; DeVallance, E.; d’Audiffret, A.C.; Frisbee, S.J.; Tabone, L.E.; Shrader, C.D.; Frisbee, J.C.; Chantler, P.D. Metabolic syndrome impairs reactivity and wall mechanics of cerebral resistance arteries in obese Zucker rats. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1846–H1859. [Google Scholar] [CrossRef] [Green Version]
- Micioni Di Bonaventura, M.V.; Martinelli, I.; Moruzzi, M.; Micioni Di Bonaventura, E.; Giusepponi, M.E.; Polidori, C.; Lupidi, G.; Tayebati, S.K.; Amenta, F.; Cifani, C.; et al. Brain alterations in high fat diet induced obesity: Effects of tart cherry seeds and juice. Nutrients 2020, 12, 623. [Google Scholar] [CrossRef] [Green Version]
- Bailey, K.R.; Crawley, J.N. Anxiety-Related Behaviors in Mice. In Methods of Behavior Analysis in Neuroscience; Buccafusco, J.J., Ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Micioni Di Bonaventura, M.V.; Lutz, T.A.; Romano, A.; Pucci, M.; Geary, N.; Asarian, L.; Cifani, C. Estrogenic suppression of binge-like eating elicited by cyclic food restriction and frustrative-nonreward stress in femalerats. Int. J. Eat. Disord. 2017, 50, 624–635. [Google Scholar] [CrossRef]
- Micioni Di Bonaventura, M.V.; Ubaldi, M.; Liberati, S.; Ciccocioppo, R.; Massi, M.; Cifani, C. Caloric restriction increases the sensitivity to the hyperphagic effect of nociceptin/orphanin FQ limiting its ability to reduce binge eating in female rats. Psychopharmacology 2013, 228, 53–63. [Google Scholar] [CrossRef]
- Tayebati, S.K.; Martinelli, I.; Moruzzi, M.; Amenta, F.; Tomassoni, D. Choline and Choline alphoscerate Do Not Modulate Inflammatory Processes in the Rat Brain. Nutrients 2017, 9, 1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbatini, M.; Tomassoni, D.; Amenta, F. Hypertensive brain damage: Comparative evaluation of protective effect of treatment with dihydropyridine derivatives in spontaneously hypertensive rats. Mech. Ageing Dev. 2001, 122, 2085–2105. [Google Scholar] [CrossRef]
- Yagi, K. Simple assay for the level of total lipid peroxides in serum or plasma. Methods Mol. Biol. 1998, 108, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.; Browne, R. The analysis of free radicals, lipid peroxides, antioxidant enzymes and compounds related to oxidative stress as applied to the clinical chemistry laboratory. Adv. Exp. Med. Biol. 1994, 366, 43–58. [Google Scholar] [CrossRef]
- Martinelli, I.; Tomassoni, D.; Moruzzi, M.; Traini, E.; Amenta, F.; Tayebati, S.K. Obesity and metabolic syndrome affect the cholinergic transmission and cognitive functions. CNS Neurol. Disord. Drug Targets 2017, 16, 664–676. [Google Scholar] [CrossRef]
- Dommermuth, R.; Ewing, K. Metabolic syndrome: Systems thinking in heart disease. Prim. Care 2018, 45, 109–129. [Google Scholar] [CrossRef]
- Michikawa, M. Cholesterol paradox: Is high total or low HDL cholesterol level a risk for Alzheimer’s disease? J. Neurosci. Res. 2003, 72, 141–146. [Google Scholar] [CrossRef]
- Sickmann, H.M.; Waagepetersen, H.S.; Schousboe, A.; Benie, A.J.; Bouman, S.D. Obesity and type 2 diabetes in rats are associated with altered brain glycogen and amino-acid homeostasis. J. Cereb. Blood Flow Metab. 2010, 30, 1527–1537. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, I.; Tomassoni, D.; Moruzzi, M.; Roy, P.; Cifani, C.; Amenta, F.; Tayebati, S.K. Cardiovascular changes related to metabolic syndrome: Evidence in obese zucker Rats. Int. J. Mol. Sci. 2020, 21, 2035. [Google Scholar] [CrossRef] [Green Version]
- Yuan, A.; Nixon, R.A. Specialized roles of neurofilament proteins in synapses: Relevance to neuropsychiatric disorders. Brain Res. Bull. 2016, 126, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Cleveland, D.W. Neuronal intermediate filaments. Annu. Rev. Neurosci. 1996, 19, 187–217. [Google Scholar] [CrossRef] [PubMed]
- McMurray, C.T. Neurodegeneration: Diseases of the cytoskeleton? Cell Death Differ. 2000, 7, 861–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camargo-De-Morais, M.; De Freitas, M.; De Mattos, A.G.; Schröder, N.; Zilles, A.C.; Lisboa, C.S.; Arteni, N.; Barlem, A.; Schierholt, R.; Zwetsch, G.; et al. Effects of brain ischemia on intermediate filaments of rat hippocampus. Neurochem. Res. 1996, 21, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayebati, S.K.; Tomassoni, D.; Amenta, F. Neuroinflammatory Markers in Spontaneously Hypertensive Rat Brain: An Immunohistochemical Study. CNS Neurol. Disord. Drug Targets 2016, 15, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Tomassoni, D.; Bellagamba, G.; Postacchini, D.; Venarucci, D.; Amenta, F. Cerebrovascular and brain microanatomy in spontaneously hypertensive rats with streptozotocin-induced diabetes. Clin. Exp. Hypertens. 2004, 26, 305–321. [Google Scholar] [CrossRef]
- Malone, J.I.; Hanna, S.K.; Saporta, S. Hyperglycemic brain injury in the rat. Brain Res. 2006, 6, 9–15. [Google Scholar] [CrossRef]
- Kalayci, R.; Kaya, M.; Uzun, H.; Bilgic, B.; Ahishali, B.; Arican, N.; Elmas, I.; Küçük, M. Influence of hypercholesterolemia and hypertension on the integrity of the blood-brain barrier in rats. Int. J. Neurosci. 2009, 119, 1881–1904. [Google Scholar] [CrossRef]
- Erdös, B.; Snipes, J.A.; Miller, A.W.; Busija, D.W. Cerebrovascular dysfunction in Zucker obese rats is mediated by oxidative stress and protein kinase C. Diabetes 2004, 53, 1352–1359. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C.; Neher, J.J. Microglial phagocytosis of live neurons. Nat. Rev. Neurosci. 2014, 15, 209–216. [Google Scholar] [CrossRef]
- Jolivel, V.; Bicker, F.; Binamé, F.; Ploen, R.; Keller, S.; Gollan, R.; Jurek, B.; Birkenstock, J.; Poisa-Beiro, L.; Bruttger, J.; et al. Perivascular microglia promote blood vessel disintegration in the ischemic penumbra. Acta Neuropathol. 2015, 129, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Garden, G.A.; Möller, T. Microglia biology in health and disease. J. Neuroimmune Pharm. 2006, 1, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.K.; Choi, J.H.; Nam, S.M.; Park, O.K.; Yoo, D.Y.; Kim, W.; Yi, S.S.; Won, M.H.; Seong, J.K.; Yoon, Y.S. Activation of microglia and induction of pro-inflammatory cytokines in the hippocampus of type 2 diabetic rats. Neurol. Res. 2014, 36, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Tomassoni, D.; Bramanti, V.; Amenta, F. Expression of aquaporins 1 and 4 in the brain of spontaneously hypertensive rats. Brain Res. 2010, 1325, 155–163. [Google Scholar] [CrossRef]
- Zhou, J.; Kong, H.; Hua, X.; Xiao, M.; Ding, J.; Hu, G. Altered blood–brain barrier integrity in adult aquaporin-4 knockout mice. Neuroreport 2008, 19, 1–5. [Google Scholar] [CrossRef]
- Hou, W.K.; Xian, Y.X.; Zhang, L.; Lai, H.; Hou, X.G.; Xu, Y.X.; Yu, T.; Xu, F.Y.; Song, J.; Fu, C.L.; et al. Influence of blood glucose on the expression of glucose trans-porter proteins 1 and 3 in the brain of diabetic rats. Chin. Med. J. 2007, 120, 1704–1709. [Google Scholar] [CrossRef]
- Erickson, M.A.; Dohi, K.; Banks, W.A. Neuroinflammation: A common pathway in CNS diseases as mediated at the blood-brain barrier. Neuroimmunomodulation 2012, 19, 121–130. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Sapolsky, R.M. Stress and cognitive function. Curr. Opin. Neurobiol. 1995, 5, 205–216. [Google Scholar] [CrossRef]
- Gispen, W.H.; Biessels, G.J. Cognition and synaptic plasticity in diabetes mellitus. Trends Neurosci. 2000, 23, 542–549. [Google Scholar] [CrossRef]
- Winocur, G.; Greenwood, C.E.; Piroli, G.G.; Grillo, C.A.; Reznikov, L.R.; Reagan, L.P.; McEwen, B.S. Memory impairment in obese Zucker rats: An investigation of cognitive function in an animal model of insulin resistance and obesity. Behav. Neurosci. 2005, 119, 1389–1395. [Google Scholar] [CrossRef] [Green Version]
- Messier, C.; Awad, N.; Gagnon, M. The relationships between atherosclerosis, heart disease, type 2 diabetes and dementia. Neurol. Res. 2004, 26, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Dinel, A.L.; André, C.; Aubert, A.; Ferreira, G.; Layé, S.; Castanon, N. Cognitive and emotional alterations are related to hippocampal inflammation in a mouse model of metabolic syndrome. PLoS ONE 2011, 6, e24325. [Google Scholar] [CrossRef] [PubMed]
- Rebolledo-Solleiro, D.; Roldán-Roldán, G.; Díaz, D.; Velasco, M.; Larqué, C.; Rico-Rosillo, G.; Vega-Robledo, G.B.; Zambrano, E.; Hiriart, M.; Pérez de la Mora, M. Increased anxiety-like behavior is associated with the metabolic syndrome in non-stressed rats. PLoS ONE 2017, 12, e0176554. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.G.; Shaw, W.N.; Neel, M.-A.; Little, L.A.; Eichberg, J. Zucker diabetic fatty rat as a model for non-insulin-dependent diabetes mellitus. ILAR J. 1990, 32, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, K.; Ramarao, P. Animal models in type 2 diabetes research: An overview. Indian J. Med. Res. 2007, 125, 451–472. [Google Scholar]
- Wohlfart, P.; Lin, J.; Dietrich, N.; Kannt, A.; Elvert, R.; Herling, A.W.; Hammes, H.P. Expression patterning reveals retinal inflammation as a minor factor in experimental retinopathy of ZDF rats. Acta Diabetol. 2014, 51, 553–558. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary and Secondary Antibodies | Company Cat. No | Dilution Western Blot | Dilution IHC |
|---|---|---|---|
| Anti Neuron-Specific nuclear protein (NEUronal Nuclei Neu-N) | Monoclonal antibody Merk-Millipore Cat. MAB377 | 1:1000 | 1:500 |
| Anti-Neurofilament 200kDa, clone RT-97 | Monoclonal antibody Merk-Millipore Cat. MAB5262 | 1:1000 | 1:500 |
| Anti-Glial Fibrillary Acidic Protein (GFAP) | Monoclonal antibody Merk-Millipore USA Cat. MAB3402 | 1:1000 | 1:500 |
| Anti-Glucose transporter 1 GLUT-1 (c-Terminus) clone 5B12.3 | Monoclonal antibody Merk-Millipore USA Cat. MABS132 | 1:500 | 1:500 |
| Ionized calcium binding adaptor molecule 1 (Iba-1) | Thermofisher USA Cat. PA5-27436 | ---- | 1:200 |
| Anti Aquaporine-4 (AQP4) | Monoclonal antibody Merk-Millipore USA AB3594 | 1:1000 | 1:500 |
| Anti-Mouse Biotinylated | Polyclonal antibody Bethyl Lab. Cat. A90-116B | ------ | 1:200 |
| Anti-Mouse HRP Conjugated | Polyclonal antibody Bethyl Lab. Cat. A90-116P | 1:5000 | ------ |
| Anti-Rabbit Biotinylated | Polyclonal antibody Bethyl Lab. A120-101B | ------ | 1:200 |
| Anti-Rabbit HRP Conjugated | Polyclonal antibody Bethyl Lab. Cat. A120-101P | 1:5000 | ------ |
| Anti-Mouse | Alexa Fluor 488® | ------ | 1:100 |
| Anti-Rabbit | Alexa Fluor 594® | ------ | 1:100 |
| 12 Weeks | 16 Weeks | 20 Weeks | ||||
|---|---|---|---|---|---|---|
| LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) | |
| Frontal Cortex | ||||||
| Fifth layer N/50,000 μm2 | 21.6 ± 1.9 | 19.3 ± 1.7 | 19.5 ± 0.8 | 15.3 ± 2.2 | 20.0 ± 1.7 | 13.8 ± 0.7 * |
| Sixth layer N/50,000 μm2 | 40.3 ± 2.9 | 31.8 ± 2.8 | 39.7 ± 2.1 | 26.4 ± 3.9 * | 42.8 ± 3.0 | 24.9 ± 2.0 * |
| Hippocampus | ||||||
| CA1 subfield N/20,000 μm2 | 65.0 ± 3.1 | 71.1 ± 3.2 | 71.3 ± 2.5 | 74.5 ± 4.8 | 76.3 ± 2.7 | 65.1 ± 3.2 * |
| CA3 subfield N/20,000 μm2 | 23.1 ± 1.2 | 24.1 ± 2.8 | 27.3 ± 2.1 | 29.4 ± 2.7 | 25.2 ± 1.2 | 24.6 ± 1.9 |
| Dentate gyrus N/20,000 μm2 | 95.8 ± 2.3 | 96.1 ± 6.2 | 93.2 ± 4.2 | 95.7 ± 4.1 | 102.1 ± 2.6 | 87.5 ± 4.5 * |
| 12 Weeks | 16 Weeks | 20 Weeks | ||||
|---|---|---|---|---|---|---|
| LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) | |
| Frontal Cortex | ||||||
| Fifth layer Astrocytes immunoreaction area | 83.6 ± 5.5 | 114.8 ± 7.5 * | 80.2 ± 5.6 | 100.1 ± 4.1 * | 78.0 ± 4.2 | 113.3 ± 4.4 * |
| Fifth layer Soma area of Iba-1 immunoreactive Cells | 40.3 ± 2.1 | 42.4 ± 1.8 | 39.7 ± 2.8 | 41.3 ± 1.2 | 42.5 ± 1.4 | 47.3 ± 1.7 |
| Hippocampus | ||||||
| CA1 Astrocytes immunoreaction area | 83.9 ± 7.2 | 105.6 ± 4.5 * | 78.7 ± 3.6 | 109.3 ± 4.8 * | 89.6 ± 8.3 | 114.1 ± 6.1 * |
| CA1 Soma area of Iba-1 immunoreactive cells | 42.0 ± 3.9 | 41.6 ± 3.1 | 45.9 ± 4.9 | 55.1 ± 3.0 | 45.8 ± 0.8 | 60.1 ± 1.2 * |
| CA3 Astrocytes immunoreaction area | 83.1 ± 8.7 | 101.2 ± 5.2 * | 93.5 ± 7.0 | 115.3 ± 6.7 * | 101.1 ± 2.7 | 118.7 ± 2.7 * |
| CA3 Soma area of Iba-1 immunoreactive Cells | 40.7 ± 4.8 | 40.8 ± 3.1 | 42.5 ± 2.5 | 49.1 ± 5.6 | 46.0 ± 1.1 | 51.5 ± 2.2 * |
| Dentate gyrus Mean immunoreaction area | 132.7 ± 10.8 | 130.2 ± 7.1 | 129.1 ± 8.0 | 130.4 ± 6.2 | 135.2 ± 5.5 | 130.4 ± 7.2 |
| Dentate gyrus Soma area of Iba-1 immunoreactive Cells | 40.6 ± 4.9 | 39.8 ± 2.2 | 42.2 ± 3.2 | 50.2 ± 4.9 | 44.5 ± 2.1 | 53.4 ± 3.2 * |
| Perivascular astrocytes | ||||||
| Total Brain Mean immunoreaction area | 150.4 ± 9.3 | 164.1 ± 9.1 | 151.4 ± 5.6 | 140.7 ± 8.2 | 131.4 ± 6.4 | 128.7 ± 5.5 |
| Open Field | ||||||
|---|---|---|---|---|---|---|
| 12 Weeks | 16 Weeks | 20 Weeks | ||||
| Parameters | LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) | LZRs (n = 6) | OZRs (n = 6) |
| Tot. Dist. Trav. (cm) | 2741.6 ± 221.9 | 1307.4 ± 95.2 * | 2160.1 ± 101.3 | 1100.1 ± 87.3 * | 1910.9 ± 151.5 | 1009.2 ± 66.4 * |
| Tot. Amb. Cnts | 1870.6 ± 156.2 | 764.3 ± 62.8 * | 1956.0 ± 113.4 | 876.7 ± 90.5 * | 1667.8± 142.5 | 789.5 ± 62.4 * |
| Tot. Vert. Cnts | 94.4 ± 6.9 | 39.3 ± 3.7 * | 78.3 ± 2.6 | 37.7 ± 5.2 * | 83.5 ± 4.6 | 37.5 ± 4.5 * |
| Central Zone. Dist. Trav. | 229.3 ± 7.6 | 103.3 ± 26.7 * | 264.4 ± 22.6 | 91.9 ± 21.9 * | 224.1 ± 36.1 | 56.1 ± 7.8 * |
| Central zone Entries | 159.5 ± 9.7 | 60.2 ± 13.1 * | 103.2 ± 9.8 | 34.5 ± 9.5 * | 89.0 ± 12.1 | 21.5 ± 3.5 * |
| % Dist. Trav. | 8.6 ± 0.6 | 8.4 ± 2.3 | 12.1 ± 0.5 | 9.2 ± 2.9 | 11.8 ± 2.0 | 5.7 ± 0.9 * |
| Passive Avoidance | ||||||
| Latency time 72 h after electric shock (s) | 176.3 ± 33.2 | 174.8 ± 30.7 | 171.3 ± 32.8 | 169.8 ± 30.8 | 177.3 ± 46.2 | 47.7 ± 16.2 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomassoni, D.; Martinelli, I.; Moruzzi, M.; Micioni Di Bonaventura, M.V.; Cifani, C.; Amenta, F.; Tayebati, S.K. Obesity and Age-Related Changes in the Brain of the Zucker Lepr fa/fa Rats. Nutrients 2020, 12, 1356. https://doi.org/10.3390/nu12051356
Tomassoni D, Martinelli I, Moruzzi M, Micioni Di Bonaventura MV, Cifani C, Amenta F, Tayebati SK. Obesity and Age-Related Changes in the Brain of the Zucker Lepr fa/fa Rats. Nutrients. 2020; 12(5):1356. https://doi.org/10.3390/nu12051356
Chicago/Turabian StyleTomassoni, Daniele, Ilenia Martinelli, Michele Moruzzi, Maria Vittoria Micioni Di Bonaventura, Carlo Cifani, Francesco Amenta, and Seyed Khosrow Tayebati. 2020. "Obesity and Age-Related Changes in the Brain of the Zucker Lepr fa/fa Rats" Nutrients 12, no. 5: 1356. https://doi.org/10.3390/nu12051356