The Effect of Habitual Fat Intake, IL6 Polymorphism, and Different Diet Strategies on Inflammation in Postmenopausal Women with Central Obesity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
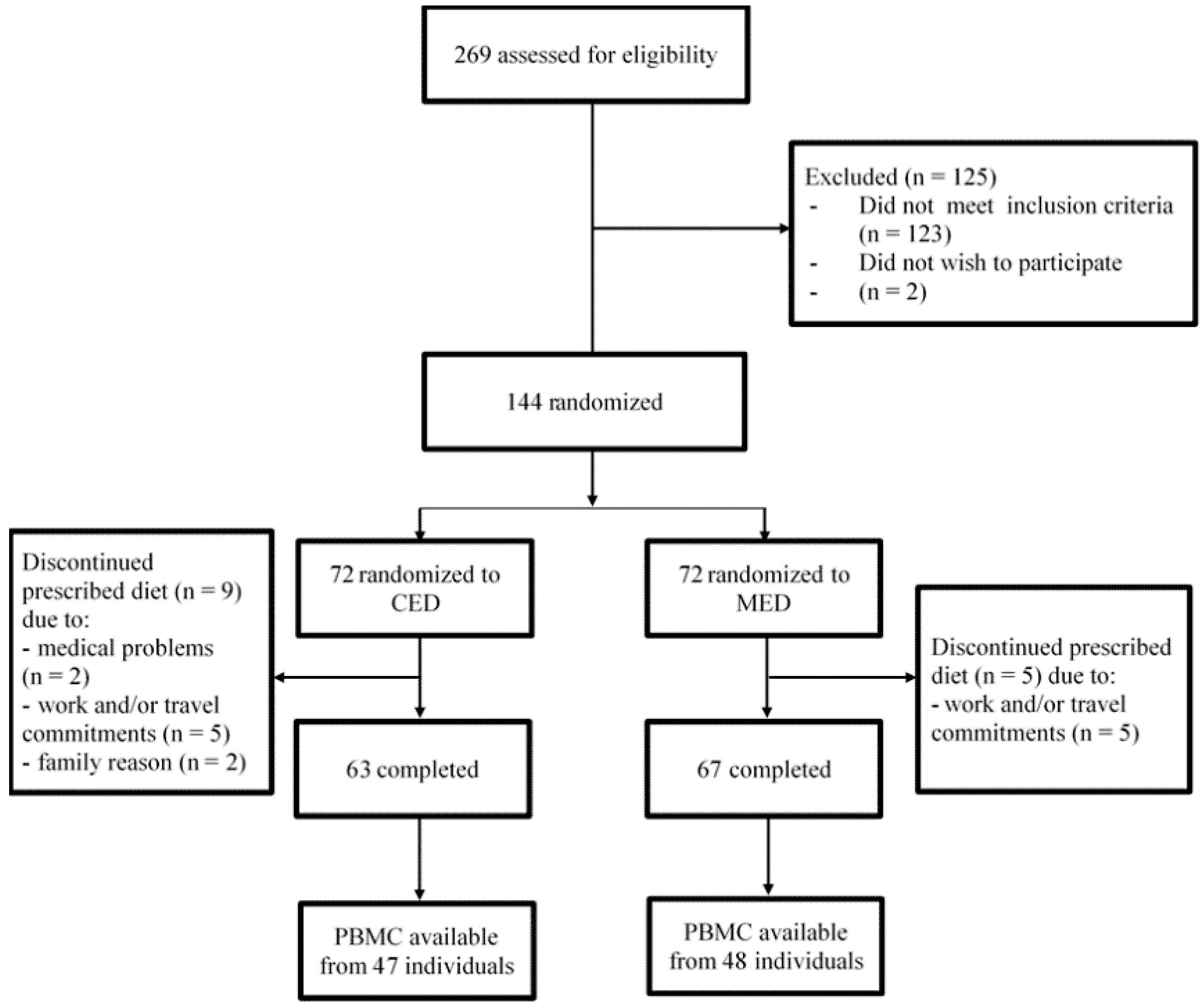
2.1. Study Design and Subjects
2.2. Dietary Assessment
2.3. Anthropometry
2.4. Analysis of Transcription Levels in PBMCs
2.5. Measurement of Blood Biomarkers
2.6. Statistical Analysis
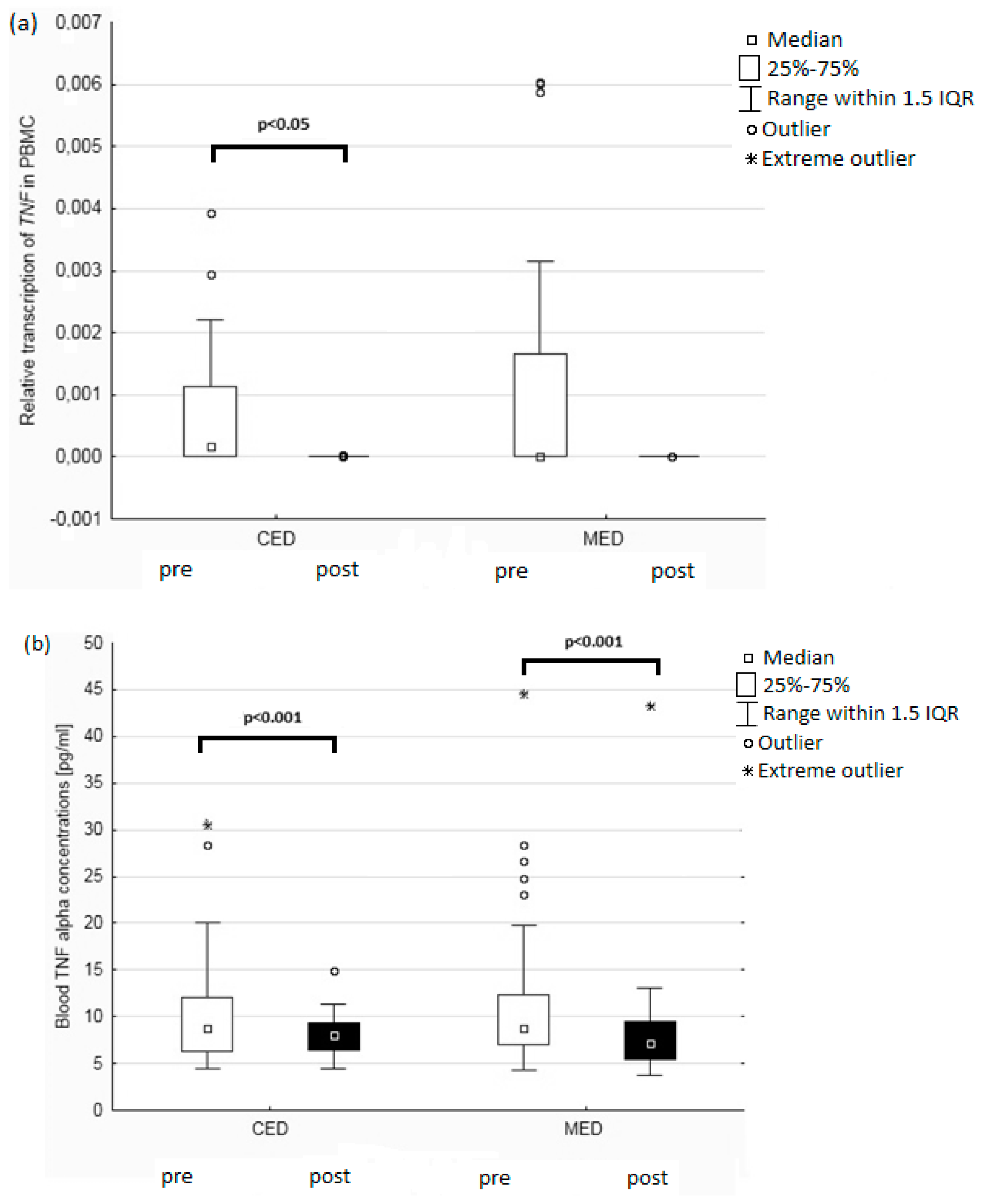
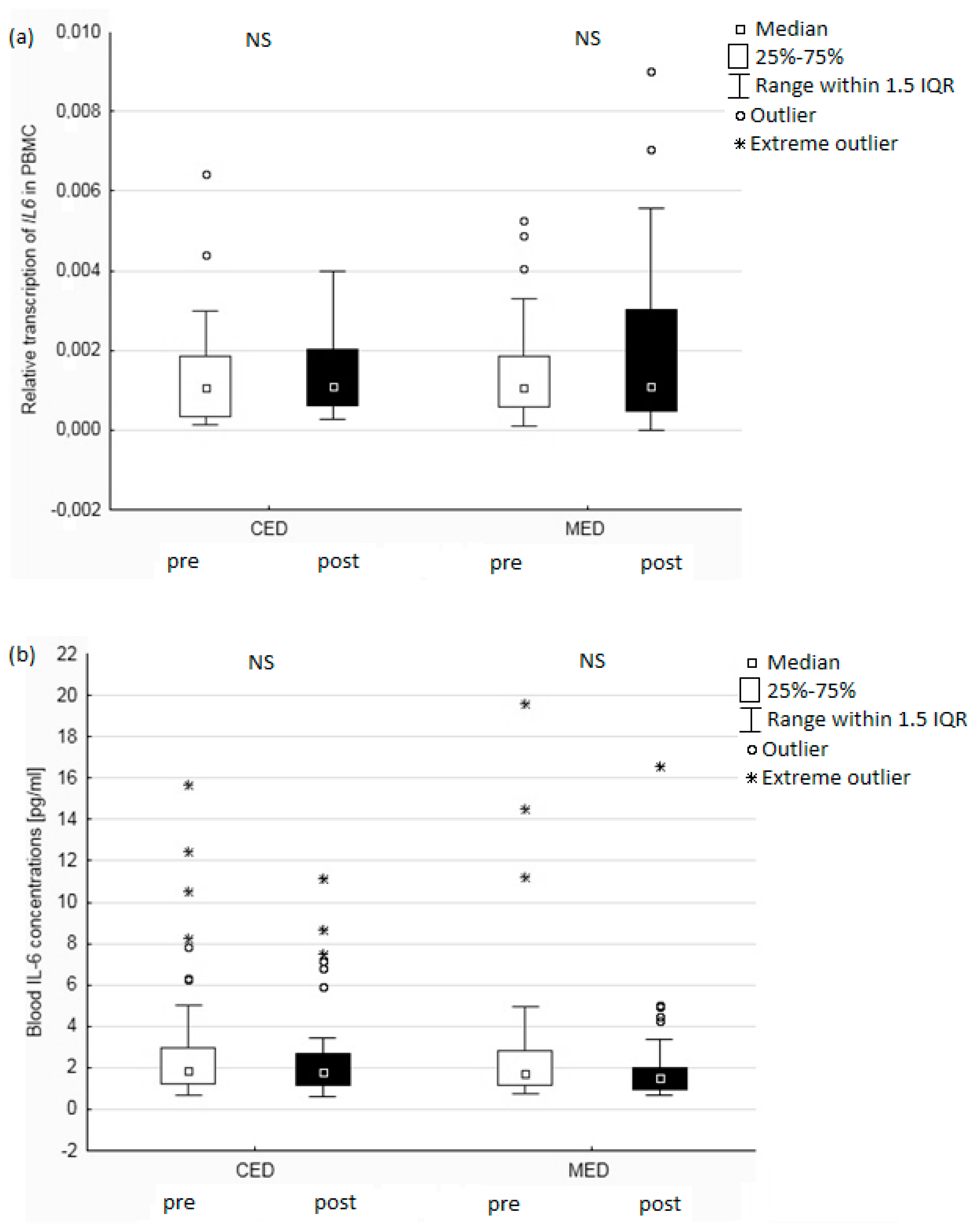
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, D.; Ceriello, A.; Esposito, K. The Effects of Diet on Inflammation. J. Am. Coll. Cardiol. 2006, 48, 677–685. [Google Scholar] [CrossRef] [PubMed]
- McArdle, M.A.; Finucane, O.M.; Connaughton, R.M.; McMorrow, A.M.; Roche, H.M. Mechanisms of Obesity-Induced Inflammation and Insulin Resistance: Insights into the Emerging Role of Nutritional Strategies. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, H.; Simental-Mendía, L.E.; Rodríguez-Ramírez, G.; Reyes-Romero, M.A. Obesity and Inflammation: Epidemiology, Risk Factors, and Markers of Inflammation. Int. J. Endocrinol. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppack, S.W. Pro-Inflammatory Cytokines and Adipose Tissue. Proc. Nutr. Soc. 2001, 60, 349–356. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin. Biochim. Biophys. Acta BBA Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Wallenius, V.; Wallenius, K.; Ahrén, B.; Rudling, M.; Carlsten, H.; Dickson, S.L.; Ohlsson, C.; Jansson, J.-O. Interleukin-6-Deficient Mice Develop Mature-Onset Obesity. Nat. Med. 2002, 8, 75–79. [Google Scholar] [CrossRef]
- Joffe, Y.; van der Merwe, L.; Evans, J.; Collins, M.; Lambert, E.; September, A.; Goedecke, J. Interleukin-6 Gene Polymorphisms, Dietary Fat Intake, Obesity and Serum Lipid Concentrations in Black and White South African Women. Nutrients 2014, 6, 2436–2465. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Chae, J.S.; Paik, J.K.; Seo, H.S.; Jang, Y.; Cavaillon, J.-M.; Lee, J.H. Effects of Aging and Menopause on Serum Interleukin-6 Levels and Peripheral Blood Mononuclear Cell Cytokine Production in Healthy Nonobese Women. AGE 2012, 34, 415–425. [Google Scholar] [CrossRef]
- Malutan, A.M.; Dan, M.; Nicolae, C.; Carmen, M. Proinflammatory and Anti-Inflammatory Cytokine Changes Related to Menopause. Menopause Rev. 2014, 3, 162–168. [Google Scholar] [CrossRef]
- Fishman, D.; Faulds, G.; Jeffery, R.; Mohamed-Ali, V.; Yudkin, J.S.; Humphries, S.; Woo, P. The Effect of Novel Polymorphisms in the Interleukin-6(IL-6) Gene on IL-6 Transcription and Plasma IL-6 Levels, and an Association with Systemic-Onset Juvenile Chronic Arthritis. J. Clin. Investig. 1998, 102, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Kubaszek, A.; Pihlajamaki, J.; Punnonen, K.; Karhapaa, P.; Vauhkonen, I.; Laakso, M. The C-174G Promoter Polymorphism of the IL-6 Gene Affects Energy Expenditure and Insulin Sensitivity. Diabetes 2003, 52, 558–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razquin, C.; Martinez, J.A.; Martinez-Gonzalez, M.A.; Fernández-Crehuet, J.; Santos, J.M.; Marti, A. A Mediterranean Diet Rich in Virgin Olive Oil May Reverse the Effects of the -174G/C IL6 Gene Variant on 3-Year Body Weight Change. Mol. Nutr. Food Res. 2010, 54, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Couture, P.; Desroches, S.; Lamarche, B. Effect of the Mediterranean Diet with and without Weight Loss on Markers of Inflammation in Men with Metabolic Syndrome: Effect of the MedDiet with and Without WL. Obesity 2013, 21, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Bajerska, J.; Chmurzynska, A.; Muzsik, A.; Krzyżanowska, P.; Mądry, E.; Malinowska, A.M.; Walkowiak, J. Weight Loss and Metabolic Health Effects from Energy-Restricted Mediterranean and Central-European Diets in Postmenopausal Women: A Randomized Controlled Trial. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Alberti, K.G.M.; Zimmet, P.; Shaw, J. The Metabolic Syndrome—A New Worldwide Definition. Lancet 2005, 366, 1059–1062. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Telle-Hansen, V.H.; Christensen, J.J.; Ulven, S.M.; Holven, K.B. Does dietary fat affect inflammatory markers in overweight and obese individuals? A review of randomized controlled trials from 2010 to 2016. Genes Nutr. 2017, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Bawadi, H.; Katkhouda, R.; Al-Haifi, A.; Tayyem, R.; Elkhoury, C.F.; Jamal, Z. Energy Balance and Macronutrient Distribution in Relation to C-Reactive Protein and HbA1c Levels among Patients with Type 2 Diabetes. Food Nutr. Res. 2016, 60, 29904. [Google Scholar] [CrossRef]
- Chawla, A. Control of Macrophage Activation and Function by PPARs. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Surmi, B.K.; Hasty, A.H. Macrophage Infiltration into Adipose Tissue: Initiation, Propagation and Remodeling. Future Lipidol. 2008, 3, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, V.E. Weight loss is a critical factor to reduce inflammation. Clin. Nutr. ESPEN. 2018, 28, 21–35. [Google Scholar] [CrossRef]
- Smidowicz, A.; Regula, J. Effect of Nutritional Status and Dietary Patterns on Human Serum C-Reactive Protein and Interleukin-6 Concentrations. Adv. Nutr. 2015, 6, 738–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imayama, I.; Ulrich, C.M.; Alfano, C.M.; Wang, C.; Xiao, L.; Wener, M.H.; Campbell, K.L.; Duggan, C.; Foster-Schubert, K.E.; Kong, A.; et al. Effects of a Caloric Restriction Weight Loss Diet and Exercise on Inflammatory Biomarkers in Overweight/Obese Postmenopausal Women: A Randomized Controlled Trial. Cancer Res. 2012, 72, 2314–2326. [Google Scholar] [CrossRef] [PubMed]
- Kolehmainen, M.; Ulven, S.M.; Paananen, J.; de Mello, V.; Schwab, U.; Carlberg, C.; Myhrstad, M.; Pihlajamäki, J.; Dungner, E.; Sjölin, E.; et al. Healthy Nordic Diet Downregulates the Expression of Genes Involved in Inflammation in Subcutaneous Adipose Tissue in Individuals with Features of the Metabolic Syndrome. Am. J. Clin. Nutr. 2015, 101, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, S.J.; Feskens, E.J.; Bos, M.B.; Hoelen, D.W.; Heijligenberg, R.; Bromhaar, M.G.; de Groot, L.C.; de Vries, J.H.; Müller, M.; Afman, L.A. A Saturated Fatty Acid–Rich Diet Induces an Obesity-Linked Proinflammatory Gene Expression Profile in Adipose Tissue of Subjects at Risk of Metabolic Syndrome. Am. J. Clin. Nutr. 2009, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, C.; Alvehus, M.; Burén, J.; Ryberg, M.; Larsson, C.; Lindahl, B.; Mellberg, C.; Söderström, I.; Chorell, E.; Olsson, T. Attenuated Low-Grade Inflammation Following Long-Term Dietary Intervention in Postmenopausal Women with Obesity: Diet Effects on Low-Grade Inflammation in Obesity. Obesity 2017, 25, 892–900. [Google Scholar] [CrossRef]
- Mališová, L.; Rossmeislová, L.; Kováčová, Z.; Kračmerová, J.; Tencerová, M.; Langin, D.; Šiklová-Vítková, M.; Štich, V. Expression of Inflammation-Related Genes in Gluteal and Abdominal Subcutaneous Adipose Tissue during Weight-Reducing Dietary Intervention in Obese Women. Physiol. Res. 2014, 63, 73–82. [Google Scholar]
- Henningsson, S.; Håkansson, A.; Westberg, L.; Baghaei, F.; Rosmond, R.; Holm, G.; Ekman, A.; Nissbrandt, H.; Eriksson, E. Interleukin-6 Gene Polymorphism −174G/C Influences Plasma Lipid Levels in Women*. Obesity 2006, 14, 1868–1873. [Google Scholar] [CrossRef]
- Klipstein-Grobusch, K.; Möhlig, M.; Spranger, J.; Hoffmann, K.; Rodrigues, F.U.S.; Sharma, A.M.; Klaus, S.; Pfeiffer, A.F.H.; Boeing, H. Interleukin-6 g.−174G>C Promoter Polymorphism Is Associated with Obesity in the EPIC-Potsdam Study*. Obesity 2006, 14, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Han, S.; Cao, X.; Zhu, C.; Wang, X.; Guo, X. Genetic Polymorphisms in Adipokine Genes and the Risk of Obesity: A Systematic Review and Meta-Analysis. Obesity 2012, 20, 396–406. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, R.W.; Kay, T.; Lyle, E.A.; Traxler, S.A.; Deveney, C.W.; Jobe, B.A.; Roberts, C.T., Jr.; Marks, D.; Rosenbaum, J.T. Alterations in peripheral blood lymphocyte cytokine expression in obesity. Clin. Exp. Immunol. 2006, 146, 39–46. [Google Scholar] [CrossRef] [PubMed]
- De Mello, V.D.; Kolehmainen, M; Schwab, U; Mager, U; Laaksonen, D.E.; Pulkkinen, L.; Niskanen, L.; Gylling, H.; Atalay, M.; Rauramaa, R.; et al. Effect of weight loss on cytokine messenger RNA expression in peripheral blood mononuclearcells of obese subjects with the metabolic syndrome. Metabolism 2008, 57, 192–199. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of Diet | CED | MED |
|---|---|---|
| Carbohydrates % E | 55 | 45 |
| Protein % E | 18 | 18 |
| Fat % E | 27 | 37 |
| SFA % E | 8 | 8 |
| MUFA % E | 10 | 20 |
| PUFA % E | 9 | 9 |
| Gene | Names of Oligonucleotides * | Primers and Probes |
|---|---|---|
| Gene expression | ||
| IL6 | #40 | 5′ gatgagtacaaaagtcctgatcca 5′ ctgcagccactggttctgt |
| TNF | #29 | 5′ cagcctcttctccttcctgat 5′ gccagagggctgattagaga |
| GAPDH | #60 | 5′ agccacatcgctcagacac 5′ gcccaatacgaccaaatcc |
| ACTB | #64 | 5′ ccaaccgcgagaagatga 5′ ccagaggcgtacagggatag |
| SNP analysis | ||
| IL6 rs1800795 | IL-F | 5′ ttactctttgtcaagacatgcca |
| IL-R | 5′ atgagcctcagacatctccag | |
| IL Anchor | 5′ ctaagctgcacttttccccctagt | |
| IL Sensor | 5′ gtgtcttgcgatgctaaagga | |
| Intake | Median Intake | TNFα [pg/mL] | p-Value | IL6 [pg/mL] | p-Value | |
|---|---|---|---|---|---|---|
| Energy from fat [%kcal] | 36% | low | 9.73 ± 6.03 | 0.184 | 2.24 ± 2.83 | 0.040 |
| high | 11.53 ± 7.11 | 3.64 ± 3.70 | ||||
| Energy from proteins [%kcal] | 17% | low | 8.71 ± 2.52 | 0.005 | 2.39 ± 2.37 | 0.126 |
| high | 12.46 ± 8.59 | 3.44 ± 4.03 | ||||
| Energy from carbohydrates [%kcal] | 46% | low | 11.39 ± 7.12 | 0.254 | 3.40 ± 3.50 | 0.167 |
| high | 9.83 ± 6.02 | 2.45 ± 3.14 | ||||
| Total fat [g/day] | 75 | low | 9.87 ± 6.54 | 0.286 | 2.92 ± 3.73 | 0.996 |
| high | 11.32 ± 6.65 | 2.92 ± 2.94 | ||||
| SFA [g/day] | 27 | low | 10.63 ± 7.57 | 0.967 | 3.14 ± 3.69 | 0.529 |
| high | 10.58 ± 5.56 | 2.70 ± 2.98 | ||||
| MUFA [g/day] | 29 | low | 10.02 ± 6.48 | 0.396 | 2.90 ± 3.74 | 0.949 |
| high | 11.18 ± 6.73 | 2.94 ± 2.94 | ||||
| PUFA [g/day] | 11 | low | 10.59 ± 6.71 | 0.976 | 2.85 ± 3.69 | 0.834 |
| high | 10.63 ± 6.54 | 3.00 ± 3.09 | ||||
| Parameter | rs1800795 Genotype Groups | p-Value | |
|---|---|---|---|
| GG | GC and CC | ||
| Body weight [kg] | 82.6 ± 10.6 | 83.6 ± 12.0 | 0.395 |
| Waist circumference [m] | 1.03 ± 0.07 | 1.03 ± 0.08 | 0.888 |
| % fat mass | 48.6 ± 4.8 | 48.3 ± 4.8 | 0.525 |
| BMI | 32.8 ± 3.8 | 32.9 ± 4.5 | 0.974 |
| WHR | 0.92 ± 0.05 | 0.92 ± 0.05 | 0.792 |
| Cholesterol [mg/dL] | 207 ± 39 | 234 ± 44 | <0.01 |
| HDL-C [mg/dL] | 55 ± 9 | 55 ± 12 | 0.921 |
| LDL-C [mg/dL] | 123 ± 36 | 147 ± 39 | <0.01 |
| TG [mg/dL] | 144 ± 74 | 162 ± 74 | 0.295 |
| Glucose [mg/dL] | 97 ± 14 | 96 ± 12 | 0.615 |
| TNFα [pg/mL] | 8.21 ± 3.60 | 11.27 ± 7.30 | 0.117 |
| IL6 [pg/mL] | 2.20 ± 1.56 | 3.03 ± 3.49 | 0.216 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmurzynska, A.; Muzsik, A.; Krzyżanowska-Jankowska, P.; Walkowiak, J.; Bajerska, J. The Effect of Habitual Fat Intake, IL6 Polymorphism, and Different Diet Strategies on Inflammation in Postmenopausal Women with Central Obesity. Nutrients 2019, 11, 1557. https://doi.org/10.3390/nu11071557
Chmurzynska A, Muzsik A, Krzyżanowska-Jankowska P, Walkowiak J, Bajerska J. The Effect of Habitual Fat Intake, IL6 Polymorphism, and Different Diet Strategies on Inflammation in Postmenopausal Women with Central Obesity. Nutrients. 2019; 11(7):1557. https://doi.org/10.3390/nu11071557
Chicago/Turabian StyleChmurzynska, Agata, Agata Muzsik, Patrycja Krzyżanowska-Jankowska, Jarosław Walkowiak, and Joanna Bajerska. 2019. "The Effect of Habitual Fat Intake, IL6 Polymorphism, and Different Diet Strategies on Inflammation in Postmenopausal Women with Central Obesity" Nutrients 11, no. 7: 1557. https://doi.org/10.3390/nu11071557