
Exploring the Exopolysaccharide Production Potential of Bacterial Strains Isolated from Tunisian Blue Crab Portunus segnis Microbiota


 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
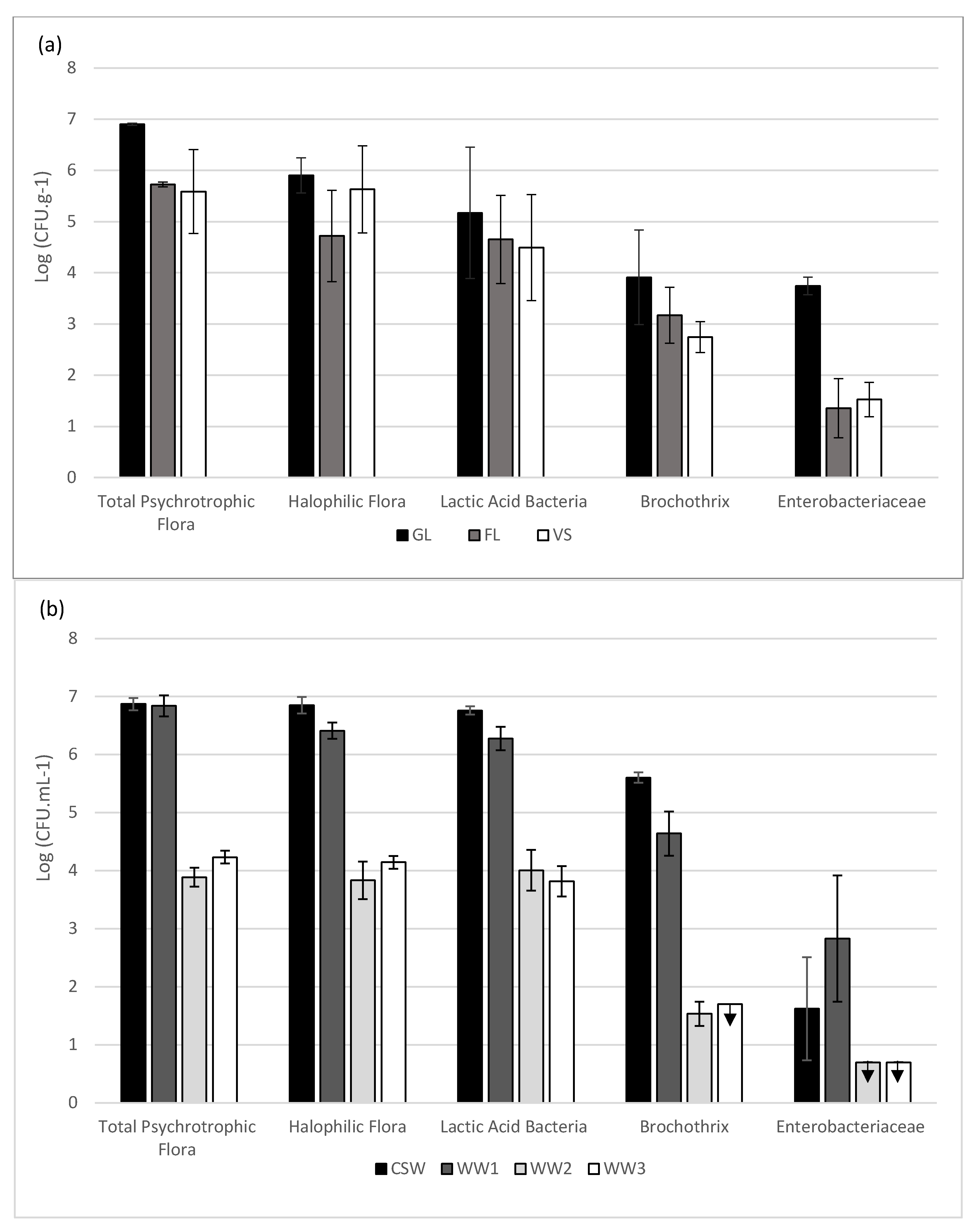
2.1. Enumeration of the Different Bacterial Groups
2.2. Bacterial Identification by 16S rRNA Gene Sequencing
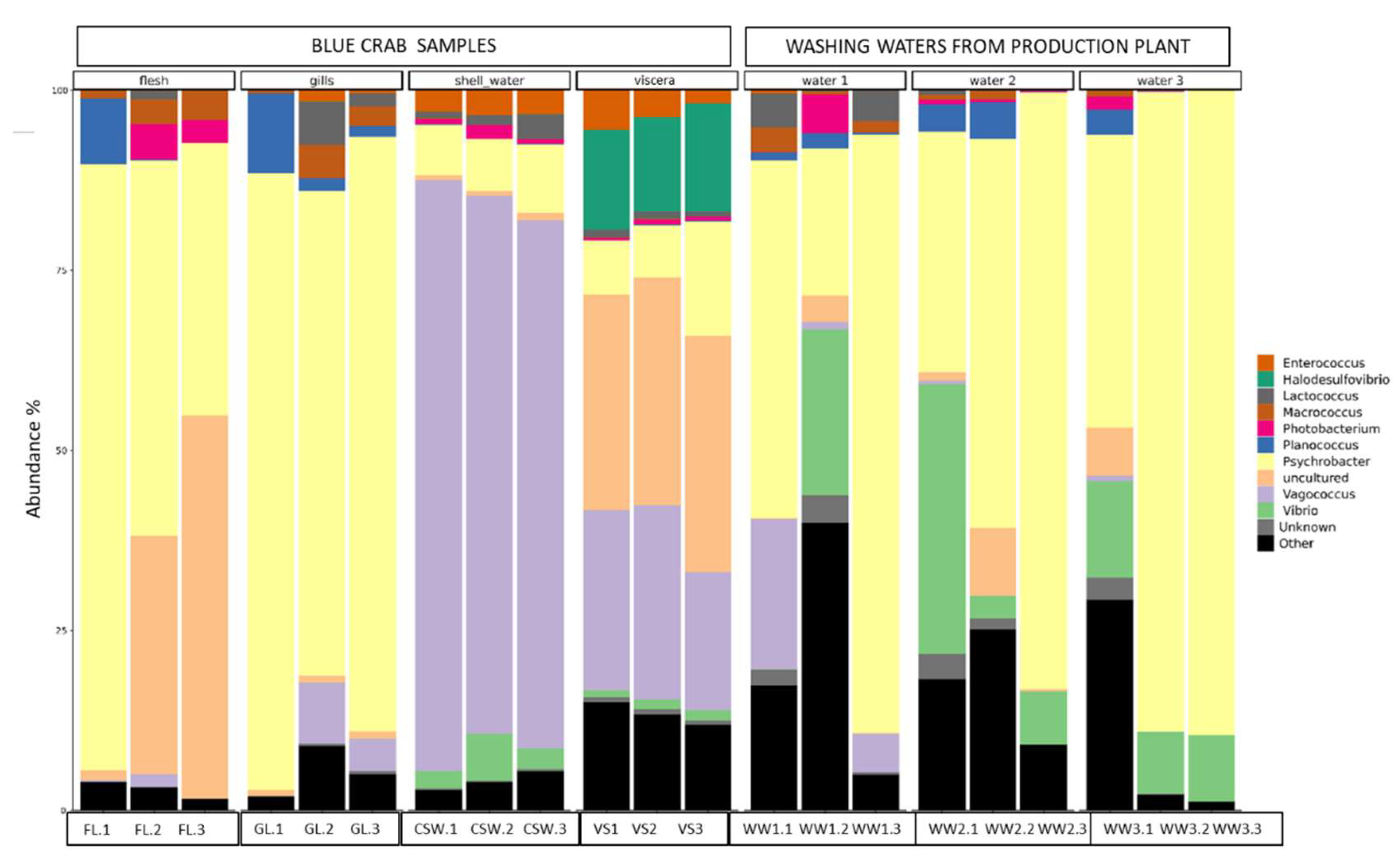
2.3. Bacterial Diversity Using Metabarcoding Analysis
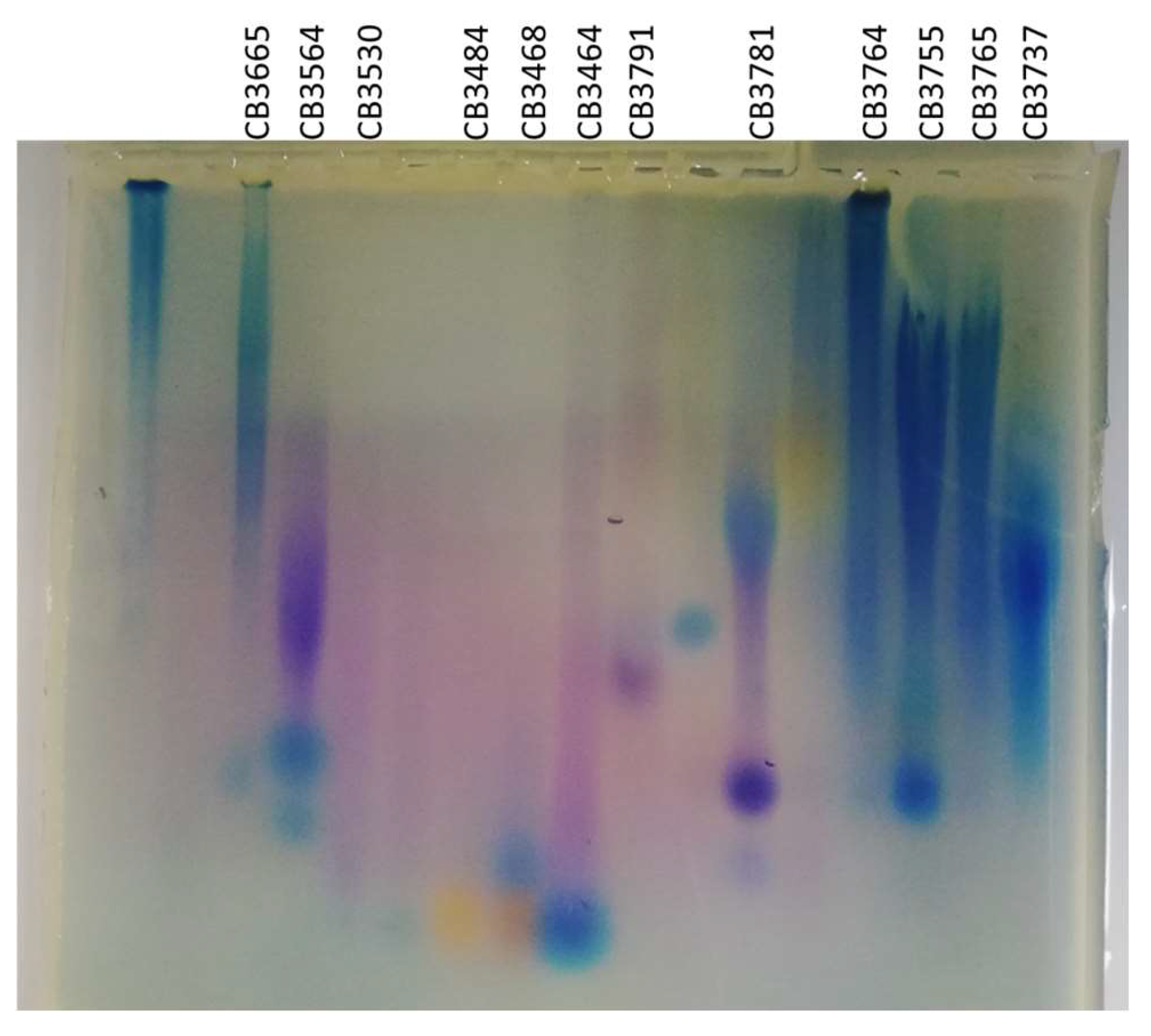
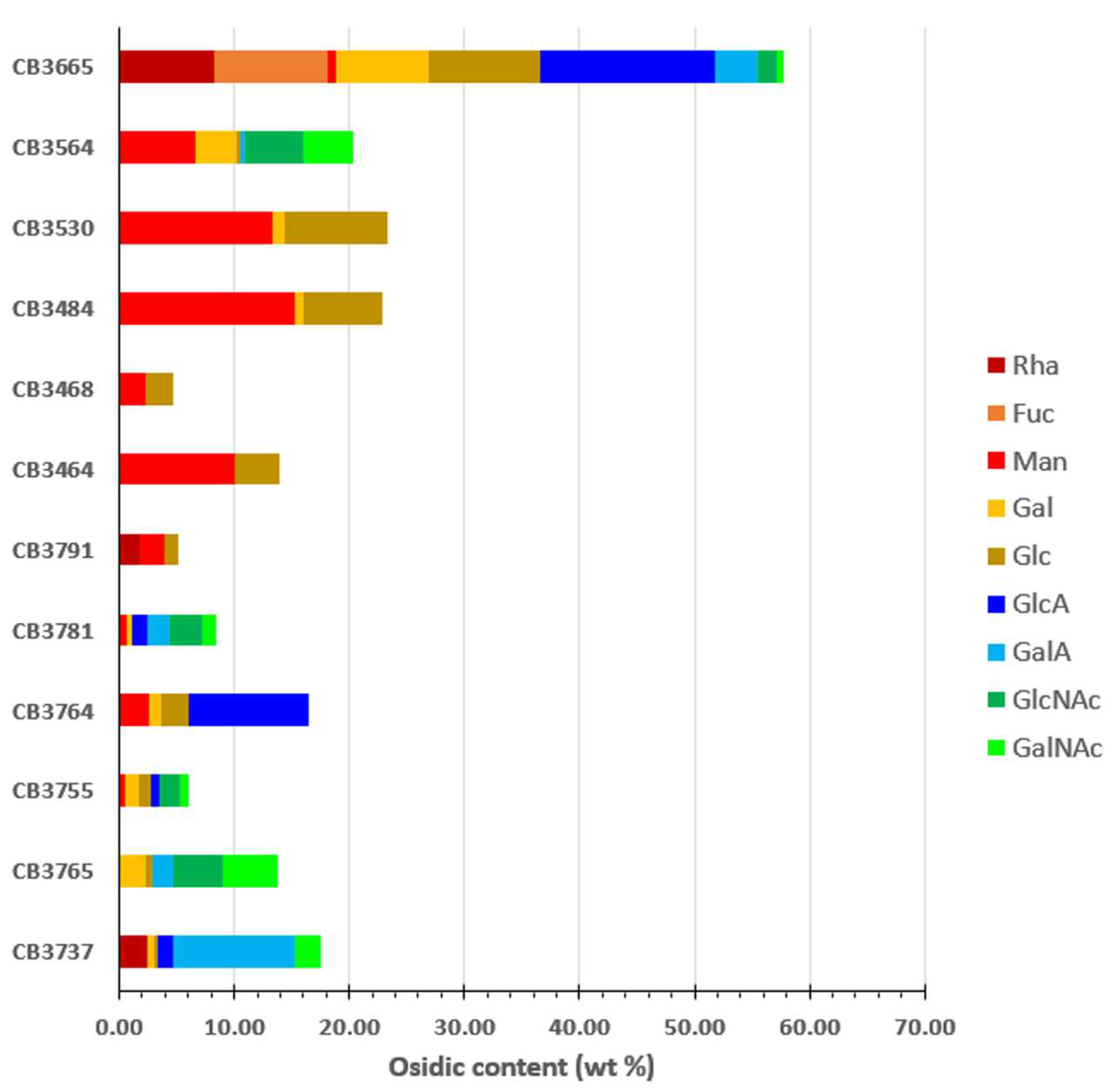
2.4. EPS-Producing Strains
3. Discussion
4. Materials and Methods
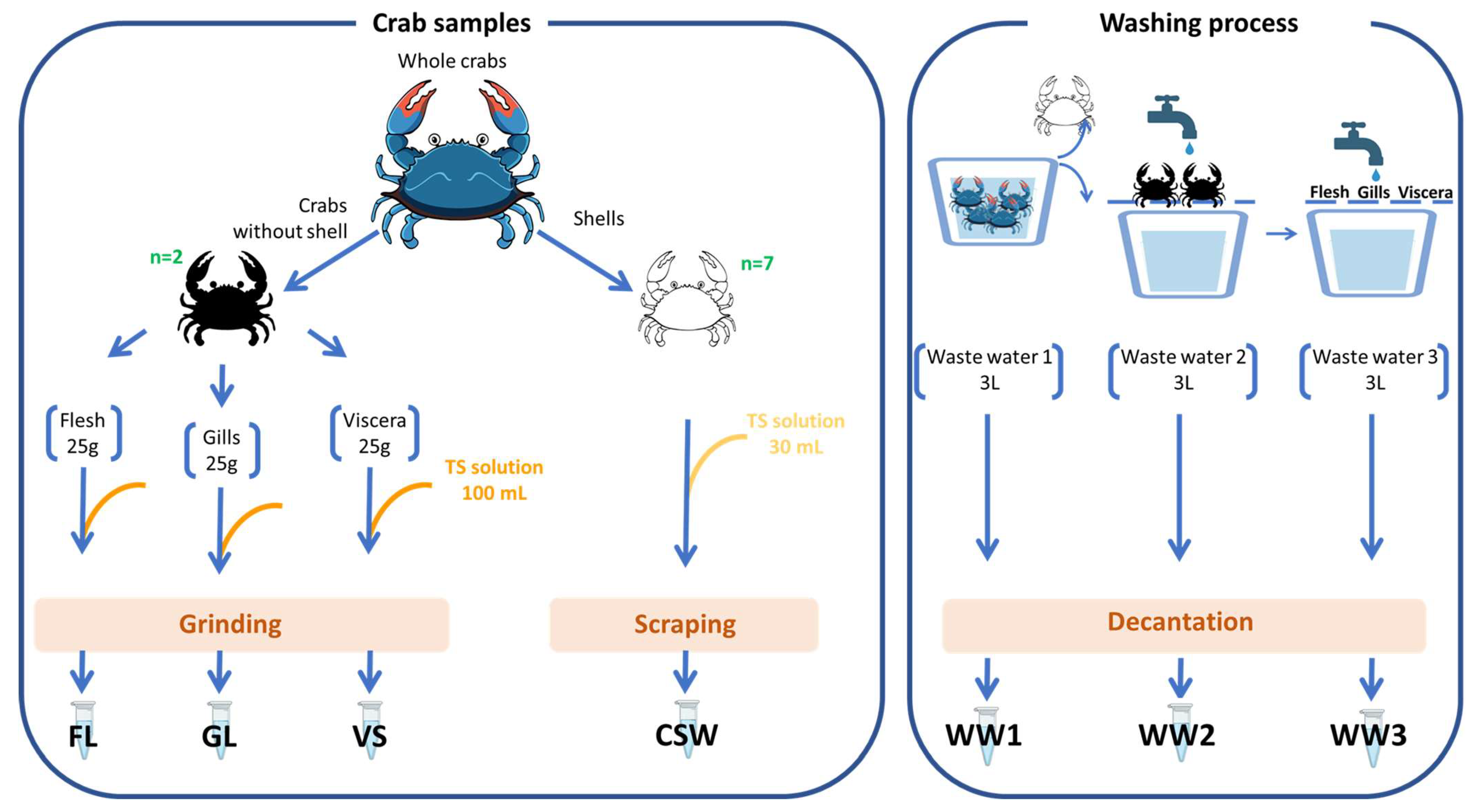
4.1. Blue Crab Sampling and Handling
4.2. Enumeration of Bacterial Groups
4.3. Collection and Identification of Bacterial Isolates
4.3.1. Bacterial Strain Collection
4.3.2. Bacterial Isolate Identification by 16S rRNA Gene Sequencing
4.4. Blue Crab Microbiota 16S rDNA Gene Metabarcoding Analysis
4.4.1. DNA Extraction
4.4.2. Metabarcoding Analysis
4.5. Screening for EPS-Producing Bacteria
4.5.1. EPS Production Screening
4.5.2. Agarose Gel Electrophoresis Analysis
4.6. EPS Characterization
4.6.1. Preparation of the EPS
4.6.2. Osidic Composition
4.6.3. Molecular Weight Determination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Castriota, L.; Falautano, M.; Maggio, T.; Perzia, P. The Blue Swimming Crab Portunus segnis in the Mediterranean Sea: Invasion Paths, Impacts and Management Measures. Biology 2022, 11, 1473. [Google Scholar] [CrossRef]
- Fox, H.M. The Migration of a Red Sea Crab through the Suez Canal. Nature 1924, 113, 714–715. [Google Scholar] [CrossRef]
- Annabi, A.; Bardelli, R.; Vizzini, S.; Mancinelli, G. Baseline Assessment of Heavy Metals Content and Trophic Position of the Invasive Blue Swimming Crab Portunus segnis (Forskål, 1775) in the Gulf of Gabès (Tunisia). Mar. Pollut. Bull. 2018, 136, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.K.; ÇAkir, M.; Ciriaco, S.; Corsini-Foka, M.; Deidun, A.; El Zrelli, R.; et al. New Mediterranean Biodiversity Records (October 2015). Mediterr. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef]
- Rabaoui, L.J.; Arculeo, M.; Mansour, L.; Tlig-Zouari, S.; Fahd, K.; Arabia, S. Occurrence of the Lessepsian Species Portunus segnis (Crustacea: Decapoda) in the Gulf of Gabes (Tunisia): First Record and New Information on Its Biology and Ecology. Cah. Biol. Mar. 2015, 56, 169–175. [Google Scholar]
- Rifi, M.; Khadija, O.; Jamila, B.S.; Zaouali, J. Première Mention Du Crabe Lessepsien Portunus segnis Dans Les Eaux Marines Tunisiennes. In Proceedings of the 4ème Congrès Franco-Maghrébin De Zoologie & 5èmes Journées Franco-Tunisiennes De Zoologie, Korba, Tunisia, 13–17 November 2014. [Google Scholar]
- Bejaoui, S.; Ghribi, F.; Hatira, S.; Chetoui, I.; Rebah, I.; Cafsi, M.h.E. First Investigation in the Biochemical Analysis of the Invasive Crab Portunus segnis from Tunisian Waters. J. Am. Oil Chem. Soc. 2017, 94, 673–682. [Google Scholar] [CrossRef]
- Affes, S.; Aranaz, I.; Acosta, N.; Heras, Á.; Nasri, M.; Maalej, H. Chitosan Derivatives-Based Films as pH-Sensitive Drug Delivery Systems with Enhanced Antioxidant and Antibacterial Properties. Int. J. Biol. Macromol. 2021, 182, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, M.; Hajji, S.; Affes, S.; Taktak, W.; Maâlej, H.; Nasri, M.; Nasri, R. Development of a Controlled Bioconversion Process for the Recovery of Chitosan from Blue Crab (Portunus segnis) Exoskeleton. Food Hydrocoll. 2018, 77, 534–548. [Google Scholar] [CrossRef]
- Hamdi, M.; Feki, A.; Bardaa, S.; Li, S.; Nagarajan, S.; Mellouli, M.; Boudawara, T.; Sahnoun, Z.; Nasri, M.; Nasri, R. A Novel Blue Crab Chitosan/Protein Composite Hydrogel Enriched with Carotenoids Endowed with Distinguished Wound Healing Capability: In Vitro Characterization and In Vivo Assessment. Mater. Sci. Eng. C 2020, 113, 110978. [Google Scholar] [CrossRef] [PubMed]
- Affes, S.; Aranaz, I.; Hamdi, M.; Acosta, N.; Ghorbel-Bellaaj, O.; Heras, Á.; Nasri, M.; Maalej, H. Preparation of a Crude Chitosanase from Blue Crab Viscera as Well as Its Application in the Production of Biologically Active Chito-Oligosaccharides from Shrimp Shells Chitosan. Int. J. Biol. Macromol. 2019, 139, 558–569. [Google Scholar] [CrossRef]
- Maalej, H.; Maalej, A.; Affes, S.; Hmidet, N.; Nasri, M. A Novel Digestive A-Amylase from Blue Crab (Portunus segnis) Viscera: Purification, Biochemical Characterization and Application for the Improvement of Antioxidant Potential of Oat Flour. Int. J. Mol. Sci. 2021, 22, 1070. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine Microorganisms as an Untapped Source of Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising Bioactive Compounds from the Marine Environment and Their Potential Effects on Various Diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine Bioactive Compounds and Health Promoting Perspectives; Innovation Pathways for Drug Discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar] [CrossRef]
- Concórdio-Reis, P.; Alves, V.D.; Moppert, X.; Guézennec, J.; Freitas, F.; Reis, M.A.M. Characterization and Biotechnological Potential of Extracellular Polysaccharides Synthesized by Alteromonas Strains Isolated from French Polynesia Marine Environments. Mar. Drugs 2021, 19, 522. [Google Scholar] [CrossRef] [PubMed]
- Benhadda, F.; Zykwinska, A.; Colliec-Jouault, S.; Sinquin, C.; Thollas, B.; Courtois, A.; Fuzzati, N.; Toribio, A.; Delbarre-Ladrat, C. Marine Versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications. Mar. Drugs 2023, 21, 582. [Google Scholar] [CrossRef] [PubMed]
- Parkar, D.; Jadhav, R.; Pimpliskar, D.R.M. Marine Bacterial Extracellular Polysaccharides: A Review. J. Coast. Life Med. 2017, 5, 29–35. [Google Scholar] [CrossRef]
- Qi, M.; Zheng, C.; Wu, W.; Yu, G.; Wang, P. Exopolysaccharides from Marine Microbes: Source, Structure and Application. Mar. Drugs 2022, 20, 512. [Google Scholar] [CrossRef] [PubMed]
- Joint, I.; Mühling, M.; Querellou, J. Culturing Marine Bacteria—An Essential Prerequisite for Biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L. Mini Review: Molecular Techniques for Identification and Characterization of Marine Biodiversity. Ann. Mar. Biol. Res. 2016, 3, 1015. [Google Scholar]
- Bakermans, C. Adaptations to Marine Versus Terrestrial Low Temperature Environments as Revealed by Comparative Genomic Analyses of the Genus Psychrobacter. FEMS Microbiol. Ecol. 2018, 94, fiy102. [Google Scholar] [CrossRef]
- Yang, X. Moraxellaceae. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 826–833. [Google Scholar] [CrossRef]
- Andrade, J.P.S.; Oliveira, C.P.; Tovar, A.M.F.; Mourão, P.A.S.; Vilanova, E. A Color-Code for Glycosaminoglycans Identification by Means of Polyacrylamide Gel Electrophoresis Stained with the Cationic Carbocyanine Dye Stains-All. Electrophoresis 2018, 39, 666–669. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Sinquin, C.; Marchand, L.; Bonnetot, S.; Zykwinska, A.; Verrez-Bagnis, V.; Colliec-Jouault, S. Influence of the Carbon and Nitrogen Sources on Diabolican Production by the Marine Vibrio diabolicus Strain Cncm I-1629. Polymers 2022, 14, 1994. [Google Scholar] [CrossRef]
- Drouillard, S.; Jeacomine, I.; Buon, L.; Boisset, C.; Courtois, A.; Thollas, B.; Morvan, P.Y.; Vallée, R.; Helbert, W. Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain. Mar. Drugs 2015, 13, 6723–6739. [Google Scholar] [CrossRef]
- Drouillard, S.; Jeacomine, I.; Buon, L.; Boisset, C.; Courtois, A.; Thollas, B.; Morvan, P.Y.; Vallée, R.; Helbert, W. Structure of the Exopolysaccharide Secreted by a Marine Strain Vibrio alginolyticus. Mar. Drugs 2018, 16, 164. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.O.; Dhama, K.; Abdel-Latif, H.M.R. The Functionality of Probiotics in Aquaculture: An Overview. Fish Shellfish Immunol. 2021, 117, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Parlapani, F.F.; Michailidou, S.; Anagnostopoulos, D.A.; Koromilas, S.; Kios, K.; Pasentsis, K.; Psomopoulos, F.; Argiriou, A.; Haroutounian, S.A.; Boziaris, I.S. Bacterial Communities and Potential Spoilage Markers of Whole Blue Crab (Callinectes sapidus) Stored under Commercial Simulated Conditions. Food Microbiol. 2019, 82, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Reed, E.; Ottesen, A. Exploring the Microbiome of Callinectes sapidus (Maryland Blue Crab). Genome Announc. 2018, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, H.; Tang, L.; Mu, C.; Ye, C.; Chen, L.; Wang, C. High-Throughput Sequencing Reveals the Core Gut Microbiota of the Mud Crab (Scylla paramamosain) in Different Coastal Regions of Southern China. BMC Genom. 2019, 20, 829. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.W.; Sizemore, R.K. Incidence of Vibrio Species Associated with Blue Crabs (Callinectes sapidus) Collected from Galveston Bay, Texas. Appl. Environ. Microbiol. 1982, 43, 1092–1097. [Google Scholar] [CrossRef]
- Tubiash, H.S.; Sizemore, R.K.; Colwell, R.R. Bacterial Flora of the Hemolymph of the Blue Crab, Callinectes sapidus: Most Probable Numbers. Appl. Microbiol. 1975, 29, 388–392. [Google Scholar] [CrossRef]
- Shields, J.D.; Overstreet, R.M. Diseases, Parasites, and Other Symbionts. In The Blue Crab Callinectes sapidus; Kennedy, V., Cronin, L., Eds.; Maryland Sea Grant: College Park, MD, USA, 2003; Volume 1, pp. 299–417. [Google Scholar]
- Kim, M.; Kwon, T.-H.; Jung, S.-M.; Cho, S.-H.; Jin, S.Y.; Park, N.-H.; Kim, C.-G.; Kim, J.-S. Antibiotic Resistance of Bacteria Isolated from the Internal Organs of Edible Snow Crabs. PLoS ONE 2013, 8, e70887. [Google Scholar] [CrossRef]
- McCarthy, Ú.; Stagg, H.; Donald, K.; Garden, A.; Weir, S. Psychrobacter sp. Isolated from the Kidney of Salmonids at a Number of Aquaculture Sites in Scotland. Bull. Eur. Assoc. Fish. Pathol. 2013, 33, 67–72. [Google Scholar]
- Parlapani, F.F.; Ferrocino, I.; Michailidou, S.; Argiriou, A.; Haroutounian, S.A.; Kokokiris, L.; Rantsiou, K.; Boziaris, I.S. Microbiota and Volatilome Profile of Fresh and Chill-Stored Deepwater Rose Shrimp (Parapenaeus longirostris). Food Res. Int. 2020, 132, 109057. [Google Scholar] [CrossRef]
- Ismail, N.; Faridah, M.; Ahmad, A.; Alia’m, A.A.; Khai, O.S.; Mohd Sofa, M.F.A.; Manca, A. Marine Bacteria Associated with Horseshoe Crabs, Tachypleus gigas and Carcinoscorpius rotundicauda. In Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management; Carmichael, R.H., Botton, M.L., Shin, P.K.S., Cheung, S.J., Eds.; Springer International Publishing: Cham, Switerland, 2015; pp. 313–320. [Google Scholar] [CrossRef]
- Li, S.; Sun, L.; Wu, H.; Hu, Z.; Liu, W.; Li, Y.; Wen, X. The Intestinal Microbial Diversity in Mud Crab (Scylla paramamosain) as Determined by Pcr-Dgge and Clone Library Analysis. J. Appl. Microbiol. 2012, 113, 1341–1351. [Google Scholar] [CrossRef]
- Anacleto, P.; Teixeira, B.; Marques, P.; Pedro, S.; Nunes, M.L.; Marques, A. Shelf-Life of Cooked Edible Crab (Cancer pagurus) Stored under Refrigerated Conditions. LWT—Food Sci. Technol. 2011, 44, 1376–1382. [Google Scholar] [CrossRef]
- Lorentzen, G.; Rotabakk, B.T.; Olsen, S.H.; Skuland, A.V.; Siikavuopio, S.I. Shelf Life of Snow Crab Clusters (Chionoecetes opilio) Stored at 0 and 4 °C. Food Control 2016, 59, 454–460. [Google Scholar] [CrossRef]
- McDermott, A.; Whyte, P.; Brunton, N.; Lyng, J.; Fagan, J.; Bolton, D.J. The Effect of Organic Acid and Sodium Chloride Dips on the Shelf-Life of Refrigerated Irish Brown Crab (Cancer agurus) Meat. LWT-Food Sci. Technol. 2018, 98, 141–147. [Google Scholar] [CrossRef]
- Talpur, M.A.D.; Memon, D.A.; Khan, M.I.; Ikhwanuddin, M.; Danish-Daniel, M.; Abol-Munafi, A. A Novel of Gut Pathogenic Bacteria of Blue Swimming Crab Portunus pelagicus (Linneaus, 1758) and Pathogenicity of Vibrio harveyi a Transmission Agent in Larval Culture under Hatchery Conditions. Res. J. Appl. Sci. 2011, 6, 116–127. [Google Scholar] [CrossRef]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Theocharis, A.D.; Pavão, M.S.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: Key Players in Cancer Cell Biology and Treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef]
- Köwitsch, A.; Chhalotre, A.; Groth, T. Effect of Thiolated Glycosaminoglycans on the Behavior of Breast Cancer Cells: Toward the Development of in Vitro Models of Cancer. Int. J. Artif. Organs 2017, 40, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.S.; Lee, E.S.; Park, P.W. Diverse Functions of Glycosaminoglycans in Infectious Diseases. Prog. Mol. Biol. Transl. Sci. 2010, 93, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Casale, J.; Crane, J. Biochemistry, Glycosaminoglycans. In Statpearls [Internet]; Statpearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Morla, S. Glycosaminoglycans and Glycosaminoglycan Mimetics in Cancer and Inflammation. Int. J. Mol. Sci. 2019, 20, 63. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, X.; Lyu, Z.; Gu, H.; Li, D.; Chen, H. Glycosaminoglycans (Gags) and Gag Mimetics Regulate the Behavior of Stem Cell Differentiation. Colloids Surf. B Biointerfaces 2017, 150, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Colliec Jouault, S.; Chevolot, L.; Helley, D.; Ratiskol, J.; Bros, A.; Sinquin, C.; Roger, O.; Fischer, A.M. Characterization, Chemical Modifications and in Vitro Anticoagulant Properties of an Exopolysaccharide Produced by Alteromonas infernus. Biochim. Biophys. Acta 2001, 1528, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Zykwinska, A.; Marchand, L.; Bonnetot, S.; Sinquin, C.; Colliec-Jouault, S.; Delbarre-Ladrat, C. Deep-Sea Hydrothermal Vent Bacteria as a Source of Glycosaminoglycan-Mimetic Exopolysaccharides. Molecules 2019, 24, 1703. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Boursicot, V.; Colliec-Jouault, S. Marine-Derived Exopolysaccharides. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 919–939. [Google Scholar] [CrossRef]
- Rougeaux, H.; Kervarec, N.; Pichon, R.; Guezennec, J. Structure of the Exopolysaccharide of Vibrio diabolicus Isolated from a Deep-Sea Hydrothermal Vent. Carbohydr. Res. 1999, 322, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Bramhachari, P.V.; Kishor, P.B.; Ramadevi, R.; Kumar, R.; Rao, B.R.; Dubey, S.K. Isolation and Characterization of Mucous Exopolysaccharide (Eps) Produced by Vibrio furnissii Strain Vb0s3. J. Microbiol. Biotechnol. 2007, 17, 44–51. [Google Scholar]
- Bramhachari, P.V.; Dubey, S.K. Isolation and Characterization of Exopolysaccharide Produced by Vibrio harveyi Strain Vb23. Lett. Appl. Microbiol. 2006, 43, 571–577. [Google Scholar] [CrossRef]
- Chalkiadakis, E.; Dufourcq, R.; Schmitt, S.; Brandily, C.; Kervarec, N.; Coatanea, D.; Amir, H.; Loubersac, L.; Chanteau, S.; Guezennec, J.; et al. Partial Characterization of an Exopolysaccharide Secreted by a Marine Bacterium, Vibrio neocaledonicus sp. Nov., from New Caledonia. J. Appl. Microbiol. 2013, 114, 1702–1712. [Google Scholar] [CrossRef]
- Rodrigues, C.; Bhosle, N.B. Exopolysaccharide Production by Vibrio fischeri, a Fouling Marine Bacterium. Biofouling 1991, 4, 301–308. [Google Scholar] [CrossRef]
- Enos-Berlage, J.L.; McCarter, L.L. Relation of Capsular Polysaccharide Production and Colonial Cell Organization to Colony Morphology in Vibrio parahaemolyticus. J. Bacteriol. 2000, 182, 5513–5520. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Li, J.; Han, F.; Duan, G.; Lu, X.; Gu, Y.; Yu, W. Antibiofilm Activity of an Exopolysaccharide from Marine Bacterium Vibrio sp. Qy101. PLoS ONE 2011, 6, e18514. [Google Scholar] [CrossRef] [PubMed]
- Delbarre-Ladrat, C.; Salas, M.L.; Sinquin, C.; Zykwinska, A.; Colliec-Jouault, S. Bioprospecting for Exopolysaccharides from Deep-Sea Hydrothermal Vent Bacteria: Relationship between Bacterial Diversity and Chemical Diversity. Microorganisms 2017, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Guo, W.; Wang, L.; Jing, C. Extracellular Polymeric Substances from Shewanella oneidensis Mr-1 Biofilms Mediate the Transformation of Ferrihydrite. Sci. Total Environ. 2021, 784, 147245. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, P.; Tang, X.; Xu, C. Characterisation and Bioactivities of an Exopolysaccharide from an Antarctic Bacterium Shewanella frigidimarina W32–2. Aquaculture 2021, 530, 735760. [Google Scholar] [CrossRef]
- Feng, L.; Qian, T.; Yang, G.; Mu, J. Characteristics of Exopolysaccharides Produced by Isolates from Natural Bioflocculant of Ruditapes philippinarum Conglutination Mud. Front. Microbiol. 2022, 13, 1068922. [Google Scholar] [CrossRef] [PubMed]
- Paulo, A.M.S.; Amorim, C.L.; Castro, P.M.L. Effect of Salt on Eps Production by Halotolerant Bacteria for Aerobic Granular Sludge Treatment. In Proceedings of the 12th Symposium of the European Society of Biochemical Engineering Sciences (Esbes), Lisbon, Portugal, 9–12 September 2018. [Google Scholar]
- Yu, L.; Sun, G.; Wei, J.; Wang, Y.; Du, C.; Li, J. Activation of Macrophages by an Exopolysaccharide Isolated from Antarctic Psychrobacter sp. B-3. Chin. J. Oceanol. Limnol. 2016, 34, 1064–1071. [Google Scholar] [CrossRef]
- van Spreekens, K.J.A. The Suitability of a Modification of Long and Hammer’s Medium for the Enumeration of More Fastidious Bacteria from Fresh Fishery Products. Arch. Fur Leb. 1974, 25, 213–219. [Google Scholar]
- Jérôme, M.; Passerini, D.; Chevalier, F.; Marchand, L.; Leroi, F.; Macé, S. Development of a Rapid Qpcr Method to Quantify Lactic Acid Bacteria in Cold-Smoked Salmon. Int. J. Food Microbiol. 2022, 363, 109504. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating Deep Phylogenetic Relationships among Cyanobacteria and Plastids by Small Subunit Rrna Sequence Analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Kisand, V.; Cuadros, R.; Wikner, J. Phylogeny of Culturable Estuarine Bacteria Catabolizing Riverine Organic Matter in the Northern Baltic Sea. Appl. Environ. Microbiol. 2002, 68, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using Qiime 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. Dada2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Olesen, S.W.; Duvallet, C.; Alm, E.J. Dbotu3: A New Implementation of Distribution-Based Otu Calling. PLoS ONE 2017, 12, e0176335. [Google Scholar] [CrossRef]
- McKnight, D.T.; Huerlimann, R.; Bower, D.S.; Schwarzkopf, L.; Alford, R.A.; Zenger, K.R. Microdecon: A Highly Accurate Read-Subtraction Tool for the Post-Sequencing Removal of Contamination in Metabarcoding Studies. Environ. DNA 2019, 1, 14–25. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The Silva Ribosomal Rna Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Sabrina, M.; Christine, D.L. Blue Crab Portunis Segnis Microbiome—16S rDNA metabarcoding—Projet VALOPOLYMER, Ifremer, Scientific Information Systems for the sea. 2022. [Google Scholar] [CrossRef]
- Kamerling, J.P.; Gerwig, G.J.; Vliegenthart, J.F.; Clamp, J.R. Characterization by Gas-Liquid Chromatography-Mass Spectrometry and Proton-Magnetic-Resonance Spectroscopy of Pertrimethylsilyl Methyl Glycosides Obtained in the Methanolysis of Glycoproteins and Glycopeptides. Biochem. J. 1975, 151, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Montreuil, J.; Bouquelet, S.; Debray, H.; Fournet, B.; Spik, G.; Strecker, G. Glycoproteins. In Carbohydrate Analysis: A Practical Approach, 2nd ed.; Chaplin, M.F., Kennedy, J.K., Eds.; Irl Press: Oxford, UK, 1986; pp. 143–204. [Google Scholar]
 ) represents the results under the limit of plate counting quantification for 3 replicates. Results are expressed as mean and standard deviation of 3 biological replicates.
) represents the results under the limit of plate counting quantification for 3 replicates. Results are expressed as mean and standard deviation of 3 biological replicates.
) represents the results under the limit of plate counting quantification for 3 replicates. Results are expressed as mean and standard deviation of 3 biological replicates.
) represents the results under the limit of plate counting quantification for 3 replicates. Results are expressed as mean and standard deviation of 3 biological replicates.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Type of Samples | Total Number | ||||||
|---|---|---|---|---|---|---|---|---|
| GL | FL | VS | CSW | WW1 | WW2 | WW3 | ||
| Aeromonas hydrophila/salmonicida | 1 | 1 | ||||||
| Aeromonas salmonicida | 1 | 1 | ||||||
| Aeromonas sp. | 1 | 1 | 2 | |||||
| Aeromonas veronii | 2 | 2 | ||||||
| Brochothrix thermosphacta | 1 | 1 | ||||||
| Buttiauxella brennerae | 1 | 1 | ||||||
| Buttiauxella sp. | 1 | 1 | ||||||
| Carnobacterium divergens | 1 | 1 | ||||||
| Carnobacterium maltaromaticum | 1 | 1 | ||||||
| Enterobacter sp. | 1 | 1 | ||||||
| Enterococcus faecalis | 4 | 1 | 5 | |||||
| Enterococcus sp. | 1 | 1 | 2 | |||||
| Escherichia coli | 1 | 1 | ||||||
| Jeotgalibaca dankookensis | 1 | 1 | 2 | 4 | ||||
| Jeotgalicoccus nanhaiensis | 1 | 1 | ||||||
| Jeotgalicoccus sp. | 2 | 2 | 4 | |||||
| Lactococcus garviae | 1 | 2 | 2 | 4 | 7 | 4 | 4 | 24 |
| Leclercia adecarboxylata | 1 | 1 | ||||||
| Lelliottia sp. | 1 | 1 | ||||||
| Macrococcus caseolyticus | 4 | 1 | 5 | |||||
| Macrococcus sp. | 2 | 1 | 3 | |||||
| Morganella morganii | 1 | 1 | 1 | 3 | ||||
| Planococcus citreus | 1 | 1 | 2 | |||||
| Planococcus rifietoensis | 1 | 1 | 2 | |||||
| Planococcus sp. | 1 | 1 | ||||||
| Proteus hauseri | 1 | 1 | 2 | |||||
| Proteus mirabilis | 1 | 1 | ||||||
| Proteus vulgaris | 1 | 1 | 2 | |||||
| Pseudomonas shahriarae | 1 | 1 | ||||||
| Psychrobacter alimentarius | 1 | 1 | ||||||
| Psychrobacter arenosus | 2 | 2 | ||||||
| Psychrobacter celer | 2 | 3 | 1 | 2 | 3 | 6 | 4 | 21 |
| Psychrobacter cibarius | 1 | 1 | ||||||
| Psychrobacter cryohalolentis | 1 | 1 | ||||||
| Psychrobacter faecalis | 2 | 2 | 4 | |||||
| Psychrobacter faecalis/pulmonis | 1 | 1 | 2 | |||||
| Psychrobacter fozii | 3 | 3 | ||||||
| Psychrobacter glacincola | 1 | 2 | 1 | 1 | 1 | 6 | ||
| Psychrobacter halodurans | 2 | 2 | ||||||
| Psychrobacter marincola | 1 | 1 | 2 | 1 | 2 | 2 | 9 | |
| Psychrobacter marincola/submarinus | 1 | 1 | 1 | 2 | 1 | 6 | ||
| Psychrobacter maritimus | 1 | 2 | 1 | 1 | 1 | 1 | 7 | |
| Psychrobacter pulmonis | 1 | 1 | 1 | 3 | ||||
| Psychrobacter sp. | 3 | 3 | 5 | 1 | 6 | 5 | 23 | |
| Psychrobacter submarinus | 1 | 1 | 1 | 1 | 4 | |||
| Raoultella terrigena | 1 | 1 | ||||||
| Salinicoccus jeotgali/salsiraiae | 2 | 2 | ||||||
| Salinicoccus salsiraiae | 1 | 1 | ||||||
| Salinicoccus sp. | 1 | 2 | 3 | |||||
| Serratia grimesii | 1 | 1 | 2 | |||||
| Serratia liquefaciens | 3 | 2 | 1 | 1 | 7 | |||
| Serratia sp. | 1 | 1 | 2 | |||||
| Shewanella algae | 2 | 2 | ||||||
| Shewanella indica | 1 | 1 | 2 | |||||
| Shewanella sp. | 1 | 1 | 2 | |||||
| Shigella sonnei | 1 | 1 | ||||||
| Staphylococcus pasteuri | 1 | 1 | ||||||
| Staphylococcus sp. | 1 | 1 | 2 | |||||
| Streptococcus parauberis | 1 | 1 | 2 | |||||
| Vagococcus fluvialis | 1 | 2 | 1 | 1 | 5 | |||
| Vagococcus sp. | 1 | 2 | 3 | 1 | 1 | 8 | ||
| Vibrio alginolyticus | 4 | 1 | 2 | 2 | 2 | 7 | 4 | 22 |
| Vibrio diabolicus/neonatus | 1 | 1 | ||||||
| Vibrio furnissii | 1 | 3 | 3 | 1 | 8 | |||
| Vibrio parahaemolyticus | 1 | 1 | ||||||
| Total number of isolates per sample | 33 | 30 | 33 | 31 | 35 | 40 | 45 | 247 |
| Code Number | Bacterial Isolate | Origin | Identification | EPS Production (mg L−1) | Mw (g mol−1) |
|---|---|---|---|---|---|
| 1 | CB3665 | Gills | Buttiauxella sp. | 44.3 | 15 × 106 |
| 2 | CB3564 | Gills | Psychrobacter sp. | 33.6 | 7.4 × 106 |
| 3 | CB3530 | WW2 | Psychrobacter marincola/submarinus | 10.5 | 13 × 106 |
| 4 | CB3484 | Viscera | Psychrobacter pulmonis | 20 | 9.7 × 106 |
| 5 | CB3468 | Gills | Psychrobacter sp. | 17.6 | 32 × 106 |
| 6 | CB3464 | Gills | Psychrobacter halodurans | 22.1 | 11 × 106 |
| 7 | CB3791 | WW3 | Shewanella sp. | 22.8 | 62 × 106 |
| 8 | CB3781 | WW2 | Vibrio alginolyticus | 55.5 | 17 × 106 |
| 9 | CB3764 | CSW | Shewanella algae | 89.6 | 16 × 106 |
| 10 | CB3755 | Viscera | Vibrio alginolyticus | 85.3 | 14 × 106 |
| 11 | CB3765 | CSW | Vibrio alginolyticus | 127.8 | 81 × 106 |
| 12 | CB3737 | Gills | Vibrio furnissii | 77 | 36 × 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migaou, M.; Macé, S.; Maalej, H.; Marchand, L.; Bonnetot, S.; Noël, C.; Sinquin, C.; Jérôme, M.; Zykwinska, A.; Colliec-Jouault, S.; et al. Exploring the Exopolysaccharide Production Potential of Bacterial Strains Isolated from Tunisian Blue Crab Portunus segnis Microbiota. Molecules 2024, 29, 774. https://doi.org/10.3390/molecules29040774
Migaou M, Macé S, Maalej H, Marchand L, Bonnetot S, Noël C, Sinquin C, Jérôme M, Zykwinska A, Colliec-Jouault S, et al. Exploring the Exopolysaccharide Production Potential of Bacterial Strains Isolated from Tunisian Blue Crab Portunus segnis Microbiota. Molecules. 2024; 29(4):774. https://doi.org/10.3390/molecules29040774
Chicago/Turabian StyleMigaou, Mariem, Sabrina Macé, Hana Maalej, Laetitia Marchand, Sandrine Bonnetot, Cyril Noël, Corinne Sinquin, Marc Jérôme, Agata Zykwinska, Sylvia Colliec-Jouault, and et al. 2024. "Exploring the Exopolysaccharide Production Potential of Bacterial Strains Isolated from Tunisian Blue Crab Portunus segnis Microbiota" Molecules 29, no. 4: 774. https://doi.org/10.3390/molecules29040774