
Phenolic Compound, Antioxidant, Antibacterial, and In Silico Studies of Extracts from the Aerial Parts of Lactuca saligna L.

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
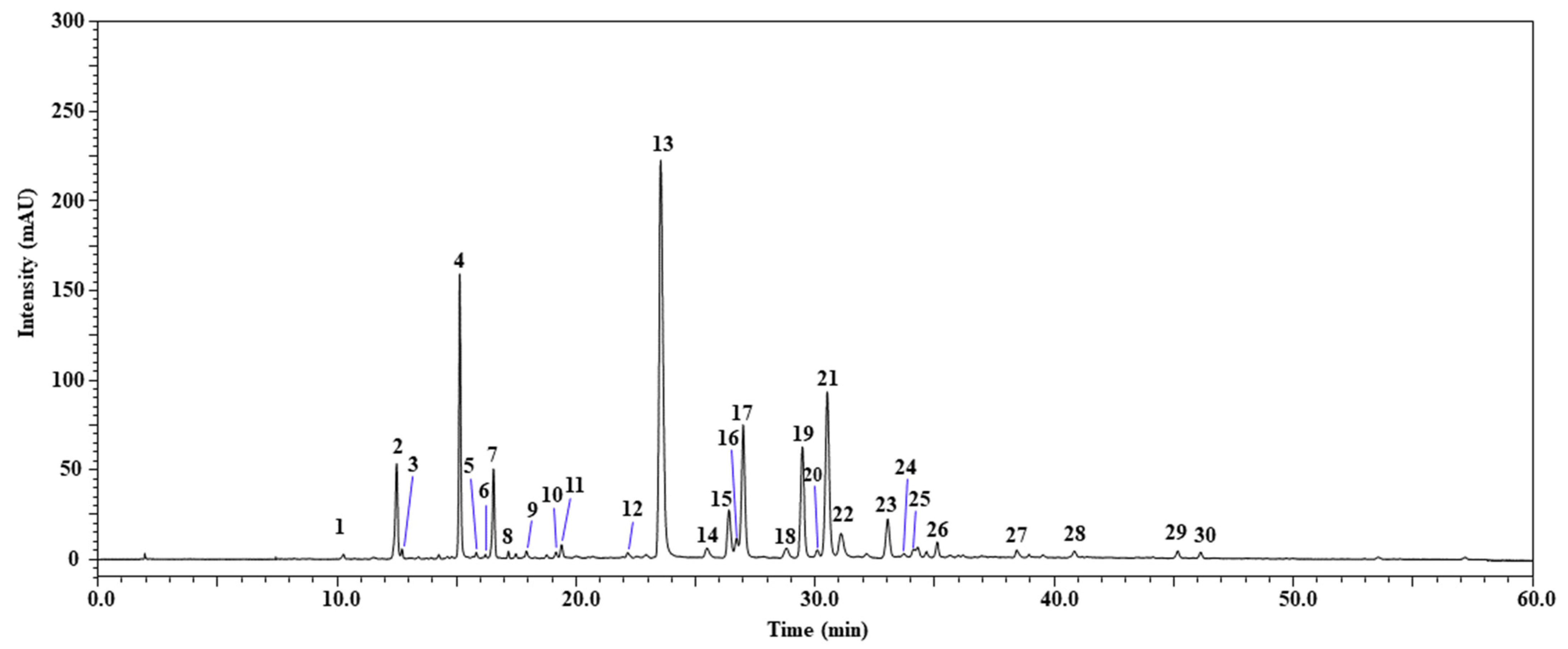
2.1. Identification of Phenolic Compounds by HPLC-PDA/ESI-MS
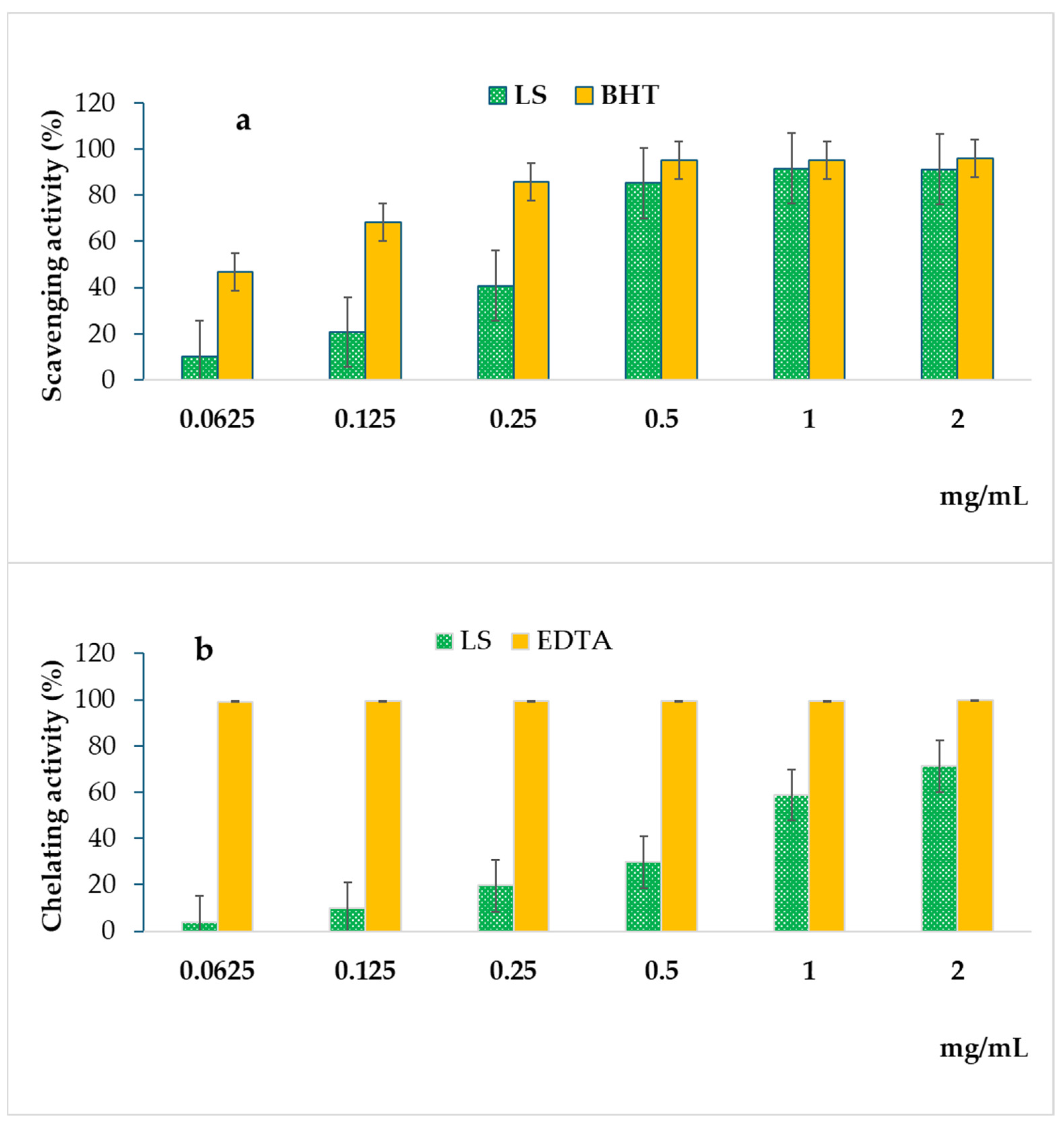
2.2. Antioxidant Activity
2.3. Antibacterial Activity
2.4. In Silico Analysis
2.4.1. Virtual Screening
2.4.2. ADMET Analysis
2.4.3. Molecular Docking
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Materials and Phenolic Compound Extraction
3.3. Polyphenolic Compound Analysis by HPLC-PDA/ESI-MS
3.4. Antioxidant Activity
3.4.1. DPPH Assay
3.4.2. FRAP Assay
3.4.3. Ferrous Ions (Fe2+) Chelating Activity
3.5. Antibacterial Activity
3.5.1. Bacterial Strains and Culture
3.5.2. The Determination of MIC and MBC
3.6. In Silico Analysis
3.6.1. Compounds and Bioinformatics Tools
3.6.2. Protein Preparation and Active Site Prediction
3.6.3. Structure-Based Virtual Screening
3.6.4. ADMET Analysis
3.6.5. Molecular Docking
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sen, T.; Samanta, S.K. Medicinal Plants, Human Health and Biodiversity: A Broad Review. Adv. Biochem. Eng. Biotechnol. 2014, 147, 59–110. [Google Scholar] [CrossRef]
- Jamila, F.; Mostafa, E. Ethnobotanical Survey of Medicinal Plants Used by People in Oriental Morocco to Manage Various Ailments. J. Ethnopharmacol. 2014, 154, 76–87. [Google Scholar] [CrossRef]
- Chroho, M.; Aazza, M.; Bouymajane, A.; El Majdoub, Y.O.; Cacciola, F.; Mondello, L.; Zair, T.; Bouissane, L. HPLC-PDA/ESI-MS Analysis of Phenolic Compounds and Bioactivities of the Ethanolic Extract from Flowers of Moroccan Anacyclus clavatus. Plants 2022, 11, 3423. [Google Scholar] [CrossRef]
- Chlif, N.; Bouymajane, A.; Oulad El Majdoub, Y.; Diouri, M.; Rhazi Filali, F.; Bentayeb, A.; Altemimi, A.B.; Mondello, L.; Cacciola, F. Phenolic Compounds, in Vivo Anti-Inflammatory, Analgesic and Antipyretic Activities of the Aqueous Extracts from Fresh and Dry Aerial Parts of Brocchia cinerea (Vis.). J. Pharm. Biomed. Anal. 2022, 213, 114695. [Google Scholar] [CrossRef] [PubMed]
- Lebeda, A.; Dolezalová, I.; Feráková, V.; Astley, D. Geographical Distribution of Wild Lactuca Species (Asteraceae, Lactuceae). Bot. Rev. 2004, 70, 328–356. [Google Scholar] [CrossRef]
- Lebeda, A.; Křístková, E.; Kitner, M.; Mieslerová, B.; Pink, D.A. Wild Lactuca Saligna: A Rich Source of Variation for Lettuce Breeding. In Enhancing Crop Genepool Use: Capturing Wild Relative and Landrace Diversity for Crop Improveme; CABI: Wallingford, UK, 2016; pp. 32–46. [Google Scholar]
- Khan, M.I.R.; Khan, N.A. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Springer: Singapore, 2017; ISBN 9789811052545. [Google Scholar]
- Bouymajane, A.; Rhazi Filali, F.; Oulad El Majdoub, Y.; Ouadik, M.; Abdelilah, R.; Cavò, E.; Miceli, N.; Taviano, M.F.; Mondello, L.; Cacciola, F. Phenolic Compounds, Antioxidant and Antibacterial Activities of Extracts from Aerial Parts of Thymus zygis Subsp. gracilis, Mentha suaveolens and Sideritis incana from Morocco. Chem. Biodivers. 2022, 19, e202101018. [Google Scholar] [CrossRef]
- Souza, M.P.; de Lima, B.R.; Sá, I.S.C.; de Freitas, F.A. Bioactive Compounds Isolated from Amazonian Fruits and Their Possible Applications. Stud. Nat. Prod. Chem. 2023, 79, 205–239. [Google Scholar] [CrossRef]
- Mehta, S.K.; Gowder, S.J.T.; Mehta, S.K.; Gowder, S.J.T. Members of Antioxidant Machinery and Their Functions. In Basic Principles and Clinical Significance of Oxidative Stress; IntechOpen: River, Croatia, 2015; ISBN 978-953-51-2200-5. [Google Scholar]
- Ndhlala, A.R.; Moyo, M.; Van Staden, J. Natural Antioxidants: Fascinating or Mythical Biomolecules? Molecules 2010, 15, 6905. [Google Scholar] [CrossRef]
- Moussa, Z.; Judeh, Z.M.A.; Ahmed, S.A.; Moussa, Z.; Judeh, Z.M.A.; Ahmed, S.A. Nonenzymatic Exogenous and Endogenous Antioxidants. In Free Radical Medicine and Biology; IntechOpen: River, Croatia, 2019; ISBN 978-1-78985-144-1. [Google Scholar]
- Pedro, A.C.; Paniz, O.G.; Fernandes, I.d.A.A.; Bortolini, D.G.; Rubio, F.T.V.; Haminiuk, C.W.I.; Maciel, G.M.; Magalhães, W.L.E. The Importance of Antioxidant Biomaterials in Human Health and Technological Innovation: A Review. Antioxidants 2022, 11, 1644. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Levy, S.B.; Bonnie, M. Antibacterial Resistance Worldwide: Causes, Challenges and Responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrob. Resist. Infect. Contr. 2019, 8, 1–28. [Google Scholar] [CrossRef]
- Anand, U.; Jacobo-Herrera, N.; Altemimi, A.; Lakhssassi, N. A Comprehensive Review on Medicinal Plants as Antimicrobial Therapeutics: Potential Avenues of Biocompatible Drug Discovery. Metabolites 2019, 9, 258. [Google Scholar] [CrossRef]
- Brown, E.D.; Wright, G.D. Antibacterial Drug Discovery in the Resistance Era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef]
- Cheynier, V.; Tomas-Barberan, F.A.; Yoshida, K. Polyphenols: From Plants to a Variety of Food and Nonfood Uses. J. Agric. Food Chem. 2015, 63, 7589–7594. [Google Scholar] [CrossRef] [PubMed]
- Sadiki, F.Z.; Bouymajane, A.; Sbiti, M.; Channaoui, S.; Micalizzi, G.; Cacciola, F.; Dugo, P.; Mondello, L.; El Idrissi, M. Chemical Profile, Antibacterial, Antioxidant and Insecticidal Properties of the Essential Oil from Tetraclinis articulata (Vahl) Masters Cones. J. Essent. Oil Res. 2022, 34, 383–393. [Google Scholar] [CrossRef]
- Ji, M.; Gong, X.; Li, X.; Wang, C.; Li, M. Advanced Research on the Antioxidant Activity and Mechanism of Polyphenols from Hippophae Species—A Review. Molecules 2020, 25, 917. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.-A. New Insights into the Mechanisms of Polyphenols beyond Antioxidant Properties; Lessons from the Green Tea Polyphenol, Epigallocatechin 3-Gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Shaker, B.; Ahmad, S.; Lee, J.; Jung, C.; Na, D. In Silico Methods and Tools for Drug Discovery. Comput. Biol. Med. 2021, 137, 104851. [Google Scholar] [CrossRef] [PubMed]
- Ilgün, S.; Akkol, E.K.; Ilhan, M.; Polat, D.Ç.; Kılıç, A.B.; Coşkun, M.; Sobarzo-Sánchez, E. Sedative Effects of Latexes Obtained from Some Lactuca L. Species Growing in Turkey. Molecules 2020, 25, 1587. [Google Scholar] [CrossRef] [PubMed]
- El-Fattah, H.A.; Khalil, A.T.; Mansour, E.S.; Waight, E.S. Triterpenes and Triterpene Esters from Lactuca saligna. Int. J. Pharmacogn. 1992, 30, 21–26. [Google Scholar] [CrossRef]
- Viacava, G.E.; Roura, S.I.; Berrueta, L.A.; Iriondo, C.; Gallo, B.; Alonso-Salces, R.M. Characterization of Phenolic Compounds in Green and Red Oak-Leaf Lettuce Cultivars by UHPLC-DAD-ESI-QToF/MS Using MSE Scan Mode. J. Mass. Spectrom. 2017, 52, 873–902. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Agustí, A.; Gratacós-Cubarsí, M.; Sárraga, C.; García-Regueiro, J.A.; Castellari, M. Analysis of Eleven Phenolic Compounds Including Novel P-Coumaroyl Derivatives in Lettuce (Lactuca sativa L.) by Ultra-High-Performance Liquid Chromatography with Photodiode Array and Mass Spectrometry Detection. Phytochem. Anal. 2011, 22, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Abu-Reidah, I.M.; Contreras, M.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Reversed-Phase Ultra-High-Performance Liquid Chromatography Coupled to Electrospray Ionization-Quadrupole-Time-of-Flight Mass Spectrometry as a Powerful Tool for Metabolic Profiling of Vegetables: Lactuca Sativa as an Example of Its Application. J. Chromatogr. A 2013, 1313, 212–227. [Google Scholar] [CrossRef]
- Pepe, G.; Sommella, E.; Manfra, M.; De Nisco, M.; Tenore, G.C.; Scopa, A.; Sofo, A.; Marzocco, S.; Adesso, S.; Novellino, T.; et al. Evaluation of Anti-Inflammatory Activity and Fast UHPLC–DAD–IT-TOF Profiling of Polyphenolic Compounds Extracted from Green Lettuce (Lactuca sativa L.; Var. Maravilla de Verano). Food Chem. 2015, 167, 153–161. [Google Scholar] [CrossRef]
- Jeong, S.W.; Kim, G.S.; Lee, W.S.; Kim, Y.H.; Kang, N.J.; Jin, J.S.; Lee, G.M.; Kim, S.T.; Abd El-Aty, A.M.; Shim, J.H.; et al. The Effects of Different Night-Time Temperatures and Cultivation Durations on the Polyphenolic Contents of Lettuce: Application of Principal Component Analysis. J. Adv. Res. 2015, 6, 493–499. [Google Scholar] [CrossRef]
- Qin, X.X.; Zhang, M.Y.; Han, Y.Y.; Hao, J.H.; Liu, C.J.; Fan, S.X. Beneficial Phytochemicals with Anti-Tumor Potential Revealed through Metabolic Profiling of New Red Pigmented Lettuces (Lactuca sativa L.). Int. J. Mol. Sci. 2018, 19, 1165. [Google Scholar] [CrossRef]
- Kalisz, S.; Oszmiański, J.; Kolniak-Ostek, J.; Grobelna, A.; Kieliszek, M.; Cendrowski, A. Effect of a Variety of Polyphenols Compounds and Antioxidant Properties of Rhubarb (Rheum rhabarbarum). LWT 2020, 118, 108775. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, W.; Li, S.; Zhou, W.; Zhang, H.; Wang, J.; Liu, X.; Zhang, J.; Liang, L.; Xu, X. Antioxidant Capacity and Interaction of Endogenous Phenolic Compounds from Tea Seed Oil. Food Chem. 2022, 376, 131940. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Wang, Z.T. Antioxidative Activity of Natural Products from Plants. Life Sci. 2000, 66, 709–723. [Google Scholar] [CrossRef]
- Makori, S.I.; Mu, T.H.; Sun, H.N. Physicochemical Properties, Antioxidant Activities, and Binding Behavior of 3,5-Di-O-Caffeoylquinic Acid with Beta-Lactoglobulin Colloidal Particles. Food Chem. 2021, 347, 129084. [Google Scholar] [CrossRef]
- Chen, X.Q.; Yang, J.H.; Cho, S.S.; Kim, J.H.; Xu, J.Q.; Seo, K.; Ki, S.H. 5-Caffeoylquinic Acid Ameliorates Oxidative Stress-Mediated Cell Death via Nrf2 Activation in Hepatocytes. Pharm. Biol. 2020, 58, 999–1005. [Google Scholar] [CrossRef]
- Mishra, B.; Priyadarsini, K.I.; Kumar, M.S.; Unnikrishnan, M.K.; Mohan, H. Effect of O-Glycosilation on the Antioxidant Activity and Free Radical Reactions of a Plant Flavonoid, Chrysoeriol. Bioorg Med. Chem. 2003, 11, 2677–2685. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and Anti-Inflammatory Activities of Quercetin and Its Derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Ganai, A.A.; Farooqi, H. Bioactivity of Genistein: A Review of In Vitro and In Vivo Studies. Biomed. Pharmacother. 2015, 76, 30–38. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Zhang, N.; Zhou, Q.; Fan, D.; Wang, M. Lipophilized Apigenin Derivatives Produced during the Frying Process as Novel Antioxidants. Food Chem. 2022, 379, 132178. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial Plant Compounds, Extracts and Essential Oils: An Updated Review on Their Effects and Putative Mechanisms of Action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef] [PubMed]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyzowska, A. Plant Extracts Rich in Polyphenols: Antibacterial Agents and Natural Preservatives for Meat and Meat Products. Crit. Rev. Food Sci. Nutr. 2021, 61, 149–178. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial Activity of Flavonoids and Their Structure–Activity Relationship: An Update Review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinic Acid: A Spectroscopic Study and Biological Screening for Antimicrobial Activity. LWT-Food Sci. Technol. 2016, 65, 471–479. [Google Scholar] [CrossRef]
- Alizadeh, S.R.; Ebrahimzadeh, M.A. Quercetin Derivatives: Drug Design, Development, and Biological Activities, a Review. Eur. J. Med. Chem. 2022, 229, 114068. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, H.; Yuan, M.; Zhou, J.; Tu, Q.; Liu, J.J.; Wang, J. Synthesis and Biological Evaluation of Apigenin Derivatives as Antibacterial and Antiproliferative Agents. Molecules 2013, 18, 11496–11511. [Google Scholar] [CrossRef]
- Lv, P.C.; Li, H.Q.; Xue, J.Y.; Shi, L.; Zhu, H.L. Synthesis and Biological Evaluation of Novel Luteolin Derivatives as Antibacterial Agents. Eur. J. Med. Chem. 2009, 44, 908–914. [Google Scholar] [CrossRef]
- Khan, F.; Bamunuarachchi, N.I.; Tabassum, N.; Kim, Y.M. Caffeic Acid and Its Derivatives: Antimicrobial Drugs toward Microbial Pathogens. J. Agric. Food Chem. 2021, 69, 2979–3004. [Google Scholar] [CrossRef] [PubMed]
- Norinder, U.; Bergström, C.A.S. Prediction of ADMET Properties. ChemMedChem 2006, 1, 920–937. [Google Scholar] [CrossRef]
- En-Nahli, F.; Hajji, H.; Ouabane, M.; Aziz Ajana, M.; Sekatte, C.; Lakhlifi, T.; Bouachrine, M. ADMET Profiling and Molecular Docking of Pyrazole and Pyrazolines Derivatives as Antimicrobial Agents. Arab. J. Chem. 2023, 16, 105262. [Google Scholar] [CrossRef]
- Ed-Dra, A.; Filai, F.R.; Bou-Idra, M.; Zekkori, B.; Bouymajane, A.; Moukrad, N.; Benhallam, F.; Bentayeb, A. Application of Mentha Suaveolens Essential Oil as an Antimicrobial Agent in Fresh Turkey Sausages. J. Appl. Biol. Biotechnol. 2018, 6, 7–12. [Google Scholar] [CrossRef]
- Bouymajane, A.; Filali, F.R.; Ed-Dra, A.; Aazza, M.; Nalbone, L.; Giarratana, F.; Alibrando, F.; Miceli, N.; Mondello, L.; Cacciola, F. Chemical Profile, Antibacterial, Antioxidant, and Anisakicidal Activities of Thymus zygis subsp. gracilis Essential Oil and Its Effect against Listeria monocytogenes. Int. J. Food Microbiol. 2022, 383, 109960. [Google Scholar] [CrossRef]
- Kimber, M.S.; Martin, F.; Lu, Y.; Houston, S.; Vedadi, M.; Dharamsi, A.; Fiebig, K.M.; Schmid, M.; Rock, C.O. The Structure of (3R)-Hydroxyacyl-Acyl Carrier Protein Dehydratase (FabZ) from Pseudomonas aeruginosa. J. Biol. Chem. 2004, 279, 52593–52602. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; Ferrari, S.; Luciani, R.; Tassone, G.; Costi, M.P.; Mangani, S. Structural Comparison of Enterococcus faecalis and Human Thymidylate Synthase Complexes with the Substrate DUMP and Its Analogue FdUMP Provides Hints about Enzyme Conformational Variabilities. Molecules 2019, 24, 1257. [Google Scholar] [CrossRef]
- Price, A.C.; Choi, K.H.; Heath, R.J.; Li, Z.; White, S.W.; Rock, C.O. Inhibition of β-Ketoacyl-Acyl Carrier Protein Synthases by Thiolactomycin and Cerulenin: Structure and Mechanism. J. Biol. Chem. 2001, 276, 6551–6559. [Google Scholar] [CrossRef] [PubMed]
- Moser, J.; Gerstel, B.; Meyer, J.E.W.; Chakraborty, T.; Wehland, J.; Heinz, D.W. Crystal Structure of the Phosphatidylinositol-Specific Phospholipase C from the Human Pathogen Listeria monocytogenes. J. Mol. Biol. 1997, 273, 269–282. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nakashima, R.; Sakurai, K.; Baucheron, S.; Giraud, E.; Doublet, B.; Cloeckaert, A.; Nishino, K. Crystal Structure of the Multidrug Resistance Regulator RamR Complexed with Bile Acids. Sci. Rep. 2019, 9, 177. [Google Scholar] [CrossRef]
- Gruszczyk, J.; Fleurie, A.; Olivares-Illana, V.; Béchet, E.; Zanella-Cleon, I.; Moréra, S.; Meyer, P.; Pompidor, G.; Kahn, R.; Grangeasse, C.; et al. Structure Analysis of the Staphylococcus aureus UDP-N-Acetyl-Mannosamine Dehydrogenase Cap5O Involved in Capsular Polysaccharide Biosynthesis. J. Biol. Chem. 2011, 286, 17112–17121. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N° | Compounds | tR (min) | UVmax (nm) | [M − H]− m/z | Phenolic Content * | References |
|---|---|---|---|---|---|---|
| 1 | di-Hydroxybenzoic acid-hexoside | 10.30 | 314 | 315 | - | [26] |
| 2 | Caffeoyltartaric acid | 12.51 | 327 | 311 | 76.39 | [27] |
| 3 | Caffeic acid-hexoside | 12.73 | 289 | 341 | 9.96 | [26] |
| 4 | 5-Caffeoylquinic acid | 15.15 | 298, 325 | 353 | 171.74 | [27] |
| 5 | Quercetin hexose-glucuronide | 15.83 | 342 | 639 | 2.05 | [26] |
| 6 | Quercetin-O-di-hexoside | 16.22 | 339 | 625, 301 | 1.38 | [26] |
| 7 | Caffeic acid | 16.57 | 294, 323 | 179 | 66.05 | [28] |
| 8 | Apigenin glucoside | 17.18 | 355 | 431 | 1.03 | [28] |
| 9 | 3-p-Coumaroylquinic acid | 17.93 | 311 | 337 | 11.70 | [26] |
| 10 | Caffeoylmalic acid | 19.18 | 326 | 295 | 9.40 | [27] |
| 11 | Caffeoylferuloylquinic acid | 19.41 | 325 | 367 | 16.27 | [29] |
| 12 | Quercetin-3-O-glucuronide | 22.19 | 279, 341 | 477, 301 | 3.33 | [26] |
| 13 | Dicaffeoyltartaric acid | 23.55 | 299, 328 | 473, 311, 179 | 472.77 | [27] |
| 14 | Dicaffeoyltartaric acid isomer | 25.48 | 299, 328 | 473, 311, 179 | 16.79 | [27] |
| 15 | Quercetin hexose | 26.40 | 254, 350 | 463, 303+ | 33.96 | [28] |
| 16 | di-4-Hydroxyphenylacetyl-hexose | 26.71 | 347 | 447 | - | [28] |
| 17 | Luteolin 7-glucoronide | 27.00 | 252, 347 | 461 | 224.30 | [28] |
| 18 | p-Coumaroylcaffeoyltartaric acid | 28.78 | 320 | 457 | 17.88 | [27,28] |
| 19 | Quercetin 6-acetyl-3-O-glucoside | 29.47 | 255, 354 | 505, 301 | 82.36 | [30] |
| 20 | Quercetin malonylglucoside | 30.09 | 363 | 549, 505, 303+ | 8.56 | [27] |
| 21 | 3,5-di-O-Caffeoylquinic acid | 30.51 | 297, 326 | 515, 353, 179 | 196.79 | [26] |
| 22 | Quercetin 3-O-rhamnoside | 31.07 | 331 | 447, 301 | 17.96 | [28] |
| 23 | Apigenin 7-O-glucuronide | 33.03 | 267, 334 | 445, 269 | 8.89 | [28] |
| 24 | Luteolin 7-glucoronide | 33.70 | 341 | 461 | 7.92 | [28] |
| 25 | 3,5-di-O-Caffeoylquinic acid isomer | 34.11 | 326 | 515 | 13.43 | [26] |
| 26 | Apigenin 7-O-glucoside | 35.10 | 267, 282, 346 | 431 | 3.59 | [31] |
| 27 | Unknown | 38.42 | 331 | 473, 269 | - | - |
| 28 | Luteolin | 40.85 | 350 | 285 | 13.20 | [28] |
| 29 | Genistein | 45.16 | 331 | 269 | 9.63 | [31] |
| 30 | Chrysoeriol | 46.12 | 345 | 299 | 1.52 | [31] |
| Samples | DPPH Assay IC50 (mg/mL) | FRAP Assay ASE/mL | FIC Assay IC50 (mg/mL) |
|---|---|---|---|
| L. saligna extract | 0.297 ± 0.012 a | 13.952 ± 0.248 a | 1.422 ± 0.003 a |
| BHT | 0.065 ± 0.008 b | 1.131 ± 0.037 b | ND |
| EDTA | ND | ND | 0.007 ± 0.000 b |
| Bacteria | Gram Type | MIC | MBC | MBC/MIC | Effect |
|---|---|---|---|---|---|
| Escherichia coli | − | 10.41 ± 0.23 | 83.33 ± 0.12 | 8 | Bacteriostatic |
| Pseudomonas aeruginosa | − | 5.20 ± 0.19 | 83.33 ± 0.20 | 16 | Bacteriostatic |
| Salmonella typhimurium | − | 10.41 ± 0.14 | 166.66 ± 0.12 | 16 | Bacteriostatic |
| Listeria monocytogenes | + | 5.20 ± 0.13 | 10.83 ± 0.12 | 2 | Bactericidal |
| Enterococcus faecalis | + | 5.20 ± 0.22 | 10.83 ± 0.15 | 2 | Bactericidal |
| Staphylococcus aureus | + | 1.30 ± 0.31 | 5.20 ± 0.16 | 4 | Bactericidal |
| Total Energy kcal/mol | Binding Affinity kcal/mol | Hydrogen Bonds | |
|---|---|---|---|
| Pseudomonas aeroginosa | |||
| Apigenin 7-O-glucuronide | −98.32 | −7.6 | Trp(60), Tyr(93), Cys(79) |
| Quercetin-3-O-glucuronide | −102.63 | −6.8 | Tyr(75), Thy(115), Ser(129), Tyr(47) |
| 3-p-Coumaroylquinic acid | −88.32 | −6.5 | Tyr(93), Ser(129) |
| Escherichia coli | |||
| Apigenin 7-O-glucuronide | −100.1 | −8.7 | Asn(396) |
| Quercetin-3-O-glucuronide | −92.16 | −7.9 | Leu(9), Asp(25), Asn(17), Glu(80) |
| 3-p-Coumaroylquinic acid | −71.21 | −7.1 | Gly(391), Thr(302), Gly(205), Met(204), Val(270) |
| Salmonella Typhimurium | |||
| Apigenin 7-O-glucuronide | −88.25 | −8.4 | Thr(85), Cys(67), Asp(152) |
| Quercetin-3-O-glucuronide | −93.48 | −8.7 | Ser(137), Thr(85), Asp(124) |
| 3-p-Coumaroylquinic acid | −78.18 | −7.7 | Asp(30), Thr(35), Ser(103) |
| Staphylococcus aureus | |||
| Apigenin 7-O-glucuronide | −102.36 | −8.8 | Thr(82), Cys(258), Asn(84), Asp(30) |
| Quercetin-3-O-glucuronide | −93.16 | −9.1 | Asn(84), Thr(119), Val(80), Ala(79), Glu(151) |
| 3-p-Coumaroylquinic acid | −80.63 | −7.6 | Ser(137), Thr(85), Tyr(59) |
| Enterococcus faecalis | |||
| Apigenin 7-O-glucuronide | −86.25 | −7.8 | Glu(59), Pro(195), Ser(218) |
| Quercetin-3-O-glucuronide | −85.47 | −7.7 | Ile(80), His(198), Asn(228) |
| 3-p-Coumaroylquinic acid | −69.22 | −6.7 | Asp(30), Thr(35), Ser(103) |
| Listeria monocytogenes | |||
| Quercetin-3-O-glucuronide | −89.03 | −7.4 | Asp(229), Thr(34) |
| Apigenin 7-O-glucuronide | −98.62 | −7.8 | Asn(207), Leu(62), Glu(128) |
| 3-p-Coumaroylquinic acid | −66.61 | −6.8 | Asp(204), Asn(207) |
| Compounds | Apigenin 7-O-glucuronide | Quercetin-3-O-glucuronide | Chrysoeriol | 3-p-Coumaroylquinic Acid |
|---|---|---|---|---|
| Absorption and Distribution | ||||
| Blood–Brain Barrier | −1.305 | −1.322 | −0.943 | −1.16 |
| Human Gut Absorption | 67 | 70 | 29 | 82 |
| Substrat glycoprotéine P | - | - | - | - |
| Inhibitor of Glycoprotein P | - | - | - | - |
| Metabolism | ||||
| CYP450 2D6 Substrate | No | No | No | No |
| CYP450 3A4 Substrate | Yes | Yes | Yes | Yes |
| CYP450 2D6 Inhibitor | No | No | No | No |
| CYP3A4 Inhibitors | No | No | No | No |
| Excretion and Toxicity | ||||
| Hepatotoxicity | No | No | No | No |
| Carcinogens | No | No | No | No |
| AMES Mutagenicity | No | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouymajane, A.; Filali, F.R.; Moujane, S.; Majdoub, Y.O.E.; Otzen, P.; Channaoui, S.; Ed-Dra, A.; Bouddine, T.; Sellam, K.; Boughrous, A.A.; et al. Phenolic Compound, Antioxidant, Antibacterial, and In Silico Studies of Extracts from the Aerial Parts of Lactuca saligna L. Molecules 2024, 29, 596. https://doi.org/10.3390/molecules29030596
Bouymajane A, Filali FR, Moujane S, Majdoub YOE, Otzen P, Channaoui S, Ed-Dra A, Bouddine T, Sellam K, Boughrous AA, et al. Phenolic Compound, Antioxidant, Antibacterial, and In Silico Studies of Extracts from the Aerial Parts of Lactuca saligna L. Molecules. 2024; 29(3):596. https://doi.org/10.3390/molecules29030596
Chicago/Turabian StyleBouymajane, Aziz, Fouzia Rhazi Filali, Soumia Moujane, Yassine Oulad El Majdoub, Philipp Otzen, Souhail Channaoui, Abdelaziz Ed-Dra, Toufik Bouddine, Khalid Sellam, Ali Ait Boughrous, and et al. 2024. "Phenolic Compound, Antioxidant, Antibacterial, and In Silico Studies of Extracts from the Aerial Parts of Lactuca saligna L." Molecules 29, no. 3: 596. https://doi.org/10.3390/molecules29030596