
Lupin as a Source of Bioactive Antioxidant Compounds for Food Products

1
Department of Food, Environmental and Nutritional Sciences (DeFENS), University of Milan, Via Celoria 2, 20133 Milan, Italy
2
Council for Agricultural Research and Economics, Research Centre for Animal Production and Aquaculture (CREA-ZA), Via Piacenza 29, 26900 Lodi, Italy
*
Author to whom correspondence should be addressed.
Molecules 2023, 28(22), 7529; https://doi.org/10.3390/molecules28227529
Submission received: 18 October 2023
/
Revised: 6 November 2023
/
Accepted: 8 November 2023
/
Published: 10 November 2023
(This article belongs to the Special Issue Biological Activity of Phenolics and Polyphenols in Nature Products)
Abstract
:Four species of lupin (white lupin, yellow lupin, blue lupin and Andean lupin) are widely cropped thanks to the excellent nutritional composition of their seeds: high protein content (28–48 g/100 g); good lipid content (4.6–13.5 g/100 g, but up to 20.0 g/100 g in Andean lupin), especially unsaturated triacylglycerols; and richness in antioxidant compounds like carotenoids, tocols and phenolics. Particularly relevant is the amount of free phenolics, highly bioaccessible in the small intestine. However, the typical bitter and toxic alkaloids must be eliminated before lupin consumption, hindering its diffusion and affecting its nutritional value. This review summarises the results of recent research in lupin composition for the above-mentioned three classes of antioxidant compounds, both in non-debittered and debittered seeds. Additionally, the influence of technological processes to further increase their nutritional value as well as the effects of food manufacturing on antioxidant content were scrutinised. Lupin has been demonstrated to be an outstanding raw material source, superior to most crops and suitable for manufacturing foods with good antioxidant and nutritional properties. The bioaccessibility of lupin antioxidants after digestion of ready-to-eat products still emerges as a dearth in current research.
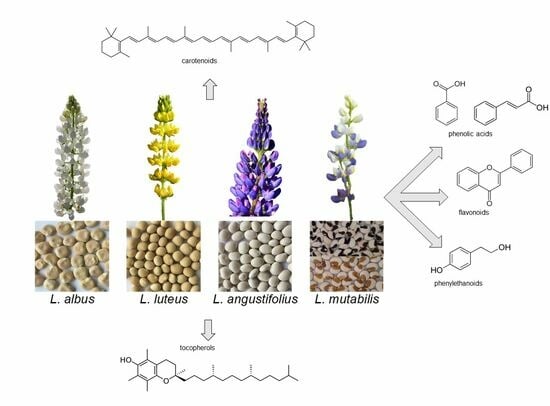
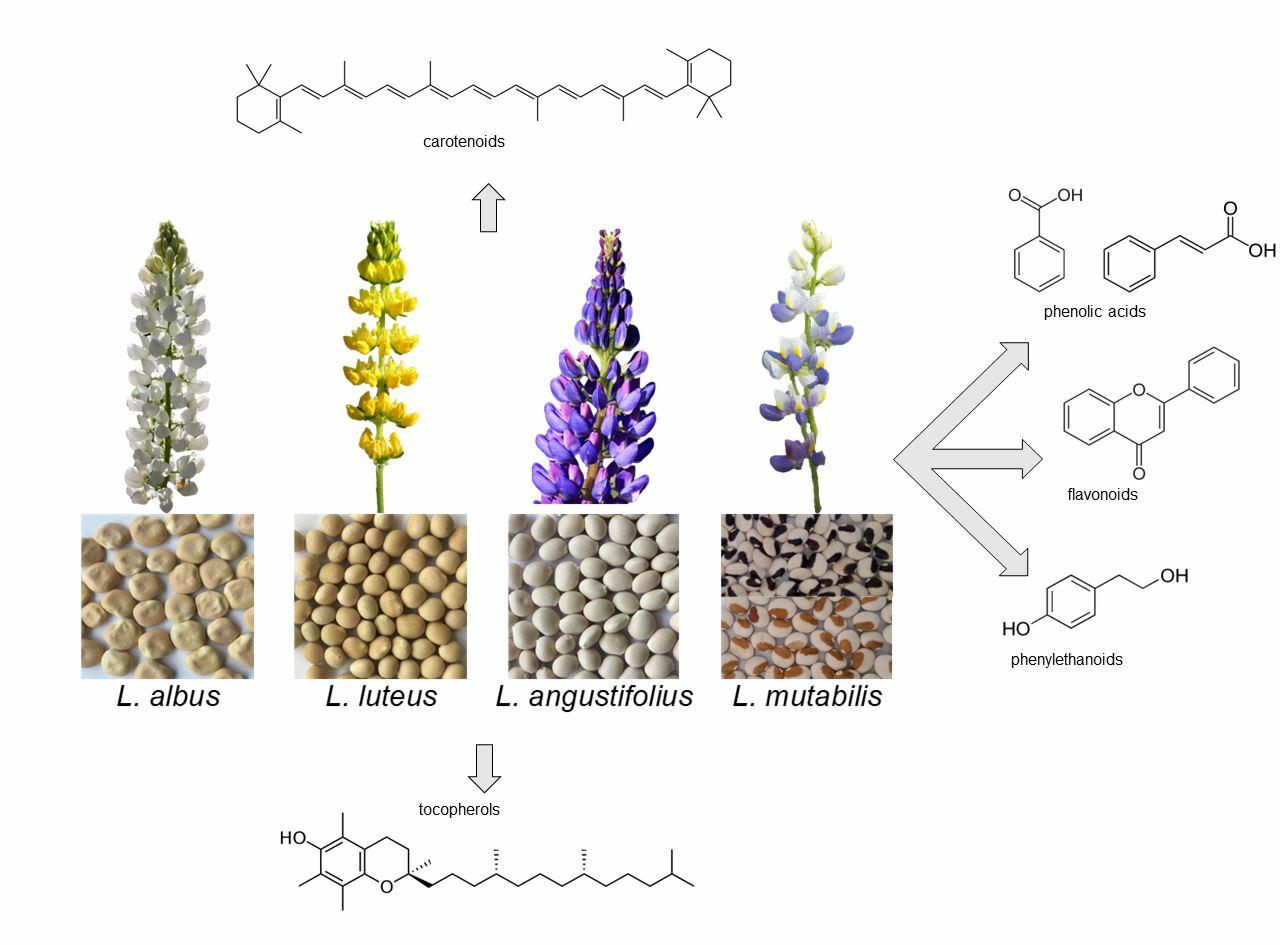
1. Introduction
Lupins are annual plants belonging to the Fabaceae family, genus Lupinus. More than 400 species are known [1], originating from the Mediterranean region, North Africa and North and South America. Four Lupinus species, i.e., white lupin (Lupinus albus L.), yellow lupin (Lupinus luteus L.), blue or narrow leaf lupin (Lupinus angustifolius L.) and Andean lupin (Lupinus mutabilis Sweet.) are widely grown as green manure, forage and food. L. angustifolius, L. albus and L. luteus (all from the Mediterranean region) are the most widespread species worldwide, while L. mutabilis (a.k.a. chocho or tarwi) is cultivated mainly in South America. According to the Food and Agriculture Organization (FAO), in 2021 the lupin cropped area was almost 1 million hectares worldwide, for a total production of 1.385 million metric tons (https://www.fao.org/faostat; last accessed on 23 September 2023); Australia is by far the most major lupin producer, with 865,619 t (about 63% of world production), followed by Poland (221,390 t), the Russian Federation (69,723 t), Morocco (56,856 t), Germany (53,400 t), Chile (37,049 t), Greece (15,830 t), Peru (15,790 t), France (15,130 t) and South Africa (9876 t) (https://www.fao.org/faostat; last accessed on 23 September 2023).
Lupins are considered improver crops because they can grow in a wide range of soil and climate conditions where other crops fail and because (like other legumes) they fix the atmospheric nitrogen in the soil through a symbiotic relationship with Rhizobium bacteria: the fixed nitrogen becomes available also for subsequent crops.
The fruit of the lupins is a pod that splits open along two seams. The seeds may be eaten as snacks, in salads or side dishes, while their flour is used in the preparation of many foods, like vegetable milk, cheese and meat, fermented products, mayonnaise, chips, noodles, pasta, baked goods, etc. [2,3,4,5,6,7,8]. Furthermore, from the seeds, it is possible to extract an oil rich in monounsaturated and polyunsaturated fatty acids [9,10] as well as protein isolates with outstanding physical, technological and functional properties [11,12,13]. The flour does not contain gluten-forming proteins and is suitable for the manufacturing of gluten-free foods for people with coeliac disease.
The aim of this review is to describe the results of current research on the content of carotenoids, tocols and phenolic compounds both in non-debittered and debittered seeds, as well as to provide some information on their fate during food processing.
2. Chemical Composition of the Seeds
The excellent composition of lupin seeds and flour confers to lupin-based foods many health benefits [14], thanks to anticancer, antimicrobial, antidiabetic, antihypertensive, antioxidant, anti-inflammatory and antimicrobial activities [15,16].
Lupin seeds are characterised by high protein content, which may vary from 28 to 48 g/100 g depending on species, variety, growing conditions and soil types [17,18]; the lowest values are found in L. angustifolius and the highest values in L. mutabilis [19,20,21,22]. The main storage proteins are globulins (80–90%), which are classified into four groups: α-conglutin (11S globulin), β-conglutin (7S globulin), γ-conglutin (7S basic globulin) and δ-conglutin (2S sulphur-rich albumin) [23,24]; prolamins and glutelins are scarce [25]. Lysine, isoleucine, leucine, phenylalanine and tyrosine are in proportions appropriate for adults [26], while methionine and cysteine (sulphur-containing amino acids), valine and tryptophan are the principal limiting compounds [27,28].
Lupin seeds contain high levels of dietary fibre [22,29], both before and after the removal of the hulls (a common step before milling). In fact, the flour on average contains 41.5 g/100 g dietary fibre (about 75% insoluble), which mainly consists of nonstarch polysaccharides (cellulose 79%, hemicellulose 14% and lignin 7%) located in the endosperm cell walls [30].
Lipid content in L. albus, L. luteus, and L. angustifolius seeds ranges from 4.6 to 13.5 g/100 g [18,31,32,33], while the L. mutabilis seeds have a higher concentration: Ruiz-Lopez et al. [18] determined a 14.7 g/100 g lipid content, while Briceño Berru et al. [19] and Carvajal-Larenas et al. [34] recorded even higher values (on average 16.1 and 18.9 g/100 g, respectively), but some genotypes even reached 20 g/100 g [35]. The oil extracted from lupin beans is characterised by scarce saturated fatty acids (10–27%) and abundant unsaturated fatty acids (78–87%), mainly oleic acid (25–63%), linoleic acid (13–57%) and linolenic acid (3–11%) [22,31,33,36,37,38], giving lupin oil outstanding nutritional and technological properties, while nonetheless maintaining remarkably high oxidative stability [9,39].
Lupin seeds contain very little starch (5–12%), while nonstarch polysaccharides are abundant. Cellulose, hemicellulose and pectin are found in the hulls, while the oligosaccharides are predominant in the cotyledons [23]. The total content of seed oligosaccharides (mainly stachyose and raffinose) varies depending on species, cultivar and environment: for example, in L. albus is 53.0 g/kg dry matter (DM) [40] while in L. mutabilis is 41.5 g/kg DM [41].
3. Antinutritional Factors: Quinolizidine Alkaloids
Compared to other legumes, lupin seeds are low in antinutritional factors such as trypsin inhibitors, lectines and saponins [29]. The content of phytic acid, a compound that reduces the bioavailability of some minerals through cation chelation, is very low (0.25–0.62 g/100 g in white lupin seeds) [29,45] and significantly inferior to soybean (1.54–2.27%) [45].
The main antinutritional compounds in lupin seeds are the quinolizidine alkaloids [27,46,47], which have a bitter taste and are often toxic, so before consumption, they must be removed through appropriate processes. The varieties with genetically low alkaloid content [48,49] have low yields, are more susceptible to pests and diseases and over time are inclined to regain their bitterness [49].
In general, debittering is achieved by removing the alkaloids from whole seeds using water as a solvent. Nevertheless, the chemical composition of the seeds is altered because several water-soluble molecules are also carried away: minerals, carbohydrates and oligosaccharides decrease while lipids and proteins increase as a consequence of concentration due to the elimination of the other compounds [20,50,51].
The traditional debittering method starts with preliminary hydration in water, followed by cooking and repeated washings for several days [50,52]. Besides disrupting cell walls and facilitating alkaloid removal, cooking blocks germination, coagulates proteins, inactivates enzymes and sanitises the product [53,54]. To shorten processing time and save water, several improved debittering methods have been proposed. Generally, they include longer boiling times [47], post-cooking washing with warm water [55], different rinsing solutions [47,56,57], germination [56,58], fermentation [59] or a combination of these methods [50,60]. Recently, Estivi et al. [61] reported that the use of sodium chloride or citric acid solutions significantly shortened debittering time, reduced water consumption and decreased alkaloid content to commercial values.
4. Carotenoids
The carotenoids, lipid-soluble antioxidants synthesised by many photosynthetic organisms, are classified as carotenes (tetraterpenoid hydrocarbons) or xanthophylls (with one or more oxygenated functions) [62]. Their single- and double-bond repeats influence the antioxidant properties, while the polar groups shape the interactions with the cellular membranes [63]. In the chloroplasts, the carotenoids act as light collectors and also safeguard against photosensitisation. The animals do not synthesise them, thus they are obtained from foods. The carotenoids are implicated in many human functions. For example, α- and β-carotenes are precursors of vitamin A, which is essential for proper visual functions, cellular reproduction and normal embryo development [64]. Furthermore, the carotenoids shield the cells from free radicals and singlet oxygen, avoiding oxidative damage. Zeaxanthin and lutein protect the macula region of the retina, contributing to the prevention of cataracts. Additional benefits linked to carotenoids are to protect against solar radiation, increase the immune response, hinder some types of cancer and assist in the prevention of cardiovascular and degenerative diseases [62,65].
Carotenoids (mainly lutein, but also α- and β-carotene, β-cryptoxanthin, zeaxanthin and violaxanthin) are scarce in lupins and therefore have not been extensively studied. Fernández-Marín et al. [66] observed 4.1 mg/kg DM in bitter seeds of L. luteus, while El-Difrawi and Hudson [67] reported 4.7 mg/kg in L. albus. In bitter seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis, the carotenoid contents were 1.90–5.26, 0.99, 6.49 and 0.69–2.89 mg/kg DM, respectively [19]. Recently, Siger et al. [68] analysed 55 L. angustifolius breeding lines and observed values ranging from 3.02 to 5.65 mg/100 g; similarly, Siger et al. [31] found 8.9–12.7, 6.1–6.3 and 17.2–62.5 mg/kg in sweet L. albus, L. luteus and L. angustifolius genotypes, while Estivi et al. [69] observed a total carotenoid content ranging from 1.70 to 1.81 mg/kg in L. mutabilis. Interestingly, debittering tends to increase their concentration, due to the loss of several water-soluble molecules [19,44].
5. Tocols
The tocols are another group of lipid-soluble antioxidants produced by photosynthetic organisms. They are separated into tocopherols (saturated phytyl group) and tocotrienols (tri-unsaturated phytyl group) but each class comprises four homologues (α, β, γ and δ), which differ in number and position of methyl groups in the chromane ring. For their capacity to reduce free radicals, in plants, they protect the photosynthetic apparatus from lipid peroxidation [71] and shield the polyunsaturated fatty acids from oxidation [72]. Like the carotenoids, the tocols are not synthesised by animals and must be acquired with food. Thanks to the capacity to quench the free radicals, they protect the polyunsaturated fatty acids in the membrane and limit free-radical damage in the tissues, contributing to the prevention of chronic diseases like cardiovascular, neurological and inflammatory diseases, cancer, etc. [73]. Only α-tocopherol has proven vitamin E activity [74] but the other tocols show comparable or even better antioxidant activity [75,76]; interestingly, the tocotrienols may even be superior to tocopherols in the prevention of cardiovascular disease [77].
As reported in Table 2, the most abundant tocol in lupins is γ-tocopherol (γ-T), representing > 97% of total tocols [19,31,32,78], followed by β-tocopherol (β-T) and δ-tocopherol (δ-T) (approximately 1% each) and by α-tocopherol (α-T; 0.3%). Briceño Berru et al. [19] reported that γ-tocopherol was plentiful in L. luteus and L. mutabilis and was less abundant in L. angustifolius and L. albus. Nevertheless, Siger et al. [31] and Martinez-Villaluenga et al. [78] observed higher γ-tocopherol concentrations in white lupin (126–130 and 201–516 mg/kg, respectively) than in yellow lupin (86–87 and 96–112 mg/kg). The total tocol content (Table 2) of L. albus can vary between 63 and 154 mg/kg [19,31,32,79] with a reported maximum of 581 mg/kg [78]; that of L. luteus may range from 86 to 187 mg/kg [19,31,78]; that of L. angustifolius can span between 65 and 123 mg/kg [19,31,32,68]; and that of L. mutabilis can vary from 169 to 250 mg/kg [19,41,44,69].
6. Free Phenolics
Another class of excellent oxygen radical scavengers includes the phenolic compounds, built by one or more aromatic rings with one or more hydroxyl groups. They are secondary metabolites produced by plants to counter environmental stresses that induce oxidative damage: the accrued synthesis of phenolic metabolites [80] stabilises the free radicals and defends membranes, lipids, proteins and DNA from oxidative harm [81]. Consequently, they safeguard the health of cell walls, providing anti-inflammatory, cardio-protective, antimicrobial, antithrombotic and vasodilatory effects [82]. The phenolics are found in three forms: soluble free, soluble conjugated, i.e., o-glycosylated to sugar moieties or other low molecular weight components, and insoluble bound, i.e., linked to cell wall constituents such as polysaccharides, protein, lignin, cutin or suberin [83]. In lupin, the most abundant fraction is by far the soluble free (>90%) [44], followed by the soluble conjugated and the insoluble bound (Table 3, Table 4, Table 5, Table 6 and Table 7).
Their bioavailability is related to their structural diversity because while the simplest free phenolic acids, like caffeic acid, can easily cross the intestinal barrier, more complex molecules are hardly absorbed [84]; nevertheless, after a bacteria-mediated partial degradation in the colon, they exhibit antioxidant capacity and may still perform a critical role in reducing the risk of bowel cancer [85].

{kind=link}
Table 3.
Free phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. albus with different solvents (* H2O; ** 1%NaCl; *** 1% citric acid).
Table 3.
Free phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. albus with different solvents (* H2O; ** 1%NaCl; *** 1% citric acid).
| Reference | [86] | [87] | [88] | [89] | [21] | [90] | [91] | [92] | [70] | [70] | [70] | [93] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bit | bit | bit | bit | bit | bit | bit | bit | deb * | deb ** | deb *** | deb * | |
| n = 11, 3 y | n = 1 | n = 2 | n = 11 | n = 2 | n = 4 | n = 4 | n = 2 | n = 2 | n = 2 | n = 2 | n = 3 | |
| Total flavonoids | 460 | 0.28–0.54 | 382–570 | 130–465 | 379–401 | 51.4–53.7 | 134–168 | 215–275 | ||||
| Apigenin | nd–9.49 | 1.10–2.66 | ||||||||||
| Apigenin der † | 0.23–0.37 | 382–570 | 1130–1310 | 130–465 | 379–401 | 30.5–49.8 | 101.9–128.7 | 164–208 | 6.79–17.31 | |||
| Apiin | 0.04–0.05 | |||||||||||
| Diosmin der | nd–1.31 | nd–2.76 | 3.94–4.81 | |||||||||
| Genistein | nd–1.78 | 1.43–2.00 | nd–13.26 | |||||||||
| Genistein der | nd–20.88 | nd–66.12 | nd–103.21 | nd–4.95 | ||||||||
| Hesperidin | nd–0.13 | |||||||||||
| Kaempferol | nd–2.75 | |||||||||||
| Myricetin | 4.2–25.9 | 11.2–21.2 | ||||||||||
| Naringenin der | nd–2.58 | nd–2.27 | 1.73–4.11 | |||||||||
| Quercetin | nd–2.61 | 0.64–1.44 | ||||||||||
| Total phenolic acids | 150 | 3.50–4.32 | 1440–3240 | 40.0–46.2 | 53.1–120.6 | 82.6–107.5 | 32.1–85.9 | |||||
| Caffeic | 151–766 | 443–766 | 0.09–0.58 | |||||||||
| p-coumaric | nd–6.44 | 2.47–6.55 | 0.11–0.18 | |||||||||
| p-coumaric der | 1440–3240 | |||||||||||
| p-hydroxybenzoic | 2.26–28.49 | 0.65–0.67 | 22.8–27.8 | |||||||||
| p-hydroxybenzoic der | 5.39–17.8 | |||||||||||
| p-aminobenzoic | 0.71–0.77 | |||||||||||
| Ferulic | 1.39–19.34 | 1.35–1.57 | 5.18–11.8 | |||||||||
| Protocatechuic | nd–7.5 | 13.0–14.7 | ||||||||||
| Sinapic acid der. | 53.1–120.6 | 82.6–107.5 | 32.1–85.9 | |||||||||
| Vanillic der | 0.61–0.94 | |||||||||||
| Other phenolics | 530 | 4.1–4.2 | ||||||||||
| Total free phenols | 1140 | 7.88–8.96 | 382–570 | 130–465 | 419–447 | 109–172 | 216–276 | 247–361 | 6.79–31.27 |
nd: not detected, i.e., below the detection limit; † der, derivative.
Therefore, almost all the data available in the literature refer to the soluble free fraction (Table 3, Table 4, Table 5 and Table 6), more abundant and with better bioaccessibility [94]. In L. albus seed samples, varying quantities of the flavonoids apigenin, apigenin derivative (der), catechin der, genistein, genistein der, kaempferol, myricetin and quercetin were detected, as well as the phenolic acids caffeic, cinnamic der, p-coumaric, p-coumaric der, p-hydroxybenzoic, p-hydroxybenzoic der, 2,4-hydroxybenzoic der, chlorogenic, vanillic, ferulic and protocatechuic [21,86,87,89,90,92,93,95]. In L. luteus, the flavonoids detected were apigenin der, diosmin der, genistein, genistein der and naringenin der [21,92], along with the phenolic acids caffeic, p-coumaric, p-hydroxybenzoic, vanillic, ferulic and protocatechuic [92,93,95]. In L. angustifolius instead the flavonoids apigenin, apigenin der, genistein and genistein der were scored as well as the phenolic acids caffeic, cinnamic der, p-coumaric, p-hydroxybenzoic, p-hydroxybenzoic der, chlorogenic, vanillic, ferulic, sinapic and protocatechuic [21,92,93,95,96,97]. Finally, in L. mutabilis the flavonoids apigenin der, catechin der, diosmin der, genistein and genistein der were recorded as well as naringenin der along with the phenylethanoids tyrosol and tyrosol der and the phenolic acids cinnamic der, p-hydroxybenzoic der, 2,4-hydroxybenzoic der and vanillic der [44,93]. The most abundant phenolics were the flavonoids (85–100% of total phenolics), followed by the phenylethanoids (not detected–13%) and the phenolic acids (not detected–2%). Genistein der (464.69 mg/kg DM) and genistein (135.78 mg/kg DM) represented 67% and 20% of total flavonoids, respectively, corresponding to 64% and 19% of total phenolics [93].
Table 4.
Free phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. luteus.
| Reference | [87] | [88] | [21] | [89] | [90] | [91] |
|---|---|---|---|---|---|---|
| bit | bit | bit | bit | bit | deb | |
| n = 1 | n = 2 | n = 2 | n = 4 | n = 2 | n = 1 | |
| Total flavonoids | 650 | 6.12–15.75 | 1380–1438 | 706–1144 | 1246–1508 | 399 |
| Apigenin der † | nd–7.35 | 138–142 | 706–1144 | 1246–1508 | 27.82 | |
| Apiin | 0.08–1.30 | |||||
| Diosmin der | 98.43 | |||||
| Genistein | 40.39 | |||||
| Genistein der | 230.00 | |||||
| Hesperidin | 0.15–0.53 | |||||
| Kaempferol | nd–0.17 | |||||
| Luteolin | nd–1.14 | |||||
| Luteolin der | nd–5.48 | |||||
| Morin dihydrate | nd–4.67 | |||||
| Naringenin der | 2.03 | |||||
| Rutin | nd–0.40 | |||||
| Rutin der | nd–0.60 | |||||
| Total phenylethanoids | 49.42 | |||||
| Tyrosol der | 49.42 | |||||
| Total phenolic acids | 60 | 6.29–6.39 | 10.0–27.6 | 41.5–82.1 | ||
| Anthranilic | nd–0.08 | |||||
| Caffeic | nd–0.10 | 1.02–1.22 | ||||
| p-coumaric | 0.56–4.47 | 0.03–0.68 | ||||
| p-hydroxybenzoic | 0.54–4.55 | 1.06–2.24 | ||||
| Ferulic | 0.26–0.39 | |||||
| Protocatechuic | 10.0–27.6 | 35.9–73.6 | ||||
| Gallic | 0.18–0.33 | |||||
| Vanillic | 0.31–0.68 | |||||
| Other phenolics | 890 | 3.62–3.70 | ||||
| Total free phenols | 1600 | 16.03–25.84 | 1380–1438 | 719–1165 | 1288–1590 | 448.09 |
nd: not detected, i.e., below the detection limit; † der, derivative.
Table 5.
Free phenolics (mg/kg DM) of bitter (bi), sweet (sw) and debittered (deb) seeds of L. angustifolius.
Table 5.
Free phenolics (mg/kg DM) of bitter (bi), sweet (sw) and debittered (deb) seeds of L. angustifolius.
| Reference | [68] | [88] | [21] | [91] | [92] | [96] | [93] |
|---|---|---|---|---|---|---|---|
| bit/sw | bit | bit | bit | bit | bit | deb | |
| n = 55 | n = 3 | n = 2 | n = 2 | n = 2 | n = 1 | n = 1 | |
| Total flavonoids | 1007–1643 | 1.84–2.91 | 784–1027 | 666–692 | 687–731 | 20.1 | 14.82 |
| Apigenin | 35.2–49.1 | ||||||
| Apigenin der † | 0.17–0.25 | 735–992 | 666–692 | 687–731 | 14.82 | ||
| Apiin | 0.12–0.19 | ||||||
| Diosmin der | |||||||
| Genistein | 20.10 | ||||||
| Genistein der | |||||||
| Hesperidin | 0.19–0.29 | ||||||
| Luteolin | 1.23–2.31 | ||||||
| Naringenin der | 0.03–0.05 | ||||||
| Rutin der | nd–0.09 | ||||||
| Total phenolic acids | 50.4–199.4 | 1.87–2.71 | 58.0–58.1 | 4.36 | |||
| Anthranilic | 0.031–0.035 | ||||||
| Caffeic | nd–0.11 | 0.62–0.63 | |||||
| Cinnamic der | 0.30 | ||||||
| p-coumaric | 0.46–0.62 | 0.34–0.42 | |||||
| p-hydroxybenzoic | 42.7–43.7 | 4.60 | |||||
| p-hydroxybenzoic der | |||||||
| Ferulic | 1.10–1.47 | ||||||
| Protocatechuic | 12.5–13.8 | ||||||
| Vanillic der | 0.31–0.40 | ||||||
| Other phenolics | 3.20–4.45 | ||||||
| Total free phenols | 7.68–9.44 | 784–1027 | 666–692 | 745–789 | 14.82 |
nd: not detected, i.e., below the detection limit; † der, derivative.
The estimates of total free phenols may vary widely (see Table 3, Table 4, Table 5 and Table 6) due to the different genotypes tested, the diverse edaphic and climatic growing conditions, the varying testing methodologies, etc. Siger et al. [92] compared three different lupin species and noticed an increasing free phenolic content from L. albus (433 mg/kg DM) to L. angustifolius (767.0 mg/kg DM) to L. luteus (1439 mg/kg DM). A similar trend was reported by Czubinski et al. [21]: L. albus (476 mg/kg DM), L. angustifolius (906 mg/kg DM) and L. luteus (1409 mg/kg DM); by Magalhães et al. [90]: L. albus (223 mg/kg DM), L. angustifolius (679 mg/kg DM) and L. luteus (960 mg/kg DM); and by Ferchichi et al. [88]: L. albus (8.42 mg/kg DM), L. angustifolius (8.37 mg/kg DM) and L. luteus (20.93 mg/kg DM) Likewise, Estivi et al. [93] observed growing concentrations of total free phenolics in L. albus (9.23 mg/kg DM), L. angustifolius (14.82 mg/kg DM), L. luteus (448 mg/kg DM) and L. mutabilis (731 mg/kg DM). The diversity within and among species is probably due to genetic selection and adaptation to cropping areas with different soil, climate and pathogens [88].
The literature available on the phenolic composition and antioxidant capacity of lupins deals mainly with raw seeds (either bitter or sweet). The debittering process, characterised by a boiling step followed by repeated washings, leads to a drastic loss of water-soluble compounds [51,60], including, unfortunately, phenolics [44,57]. After debittering L. mutabilis seeds, Villacrés et al. [57] recorded a reduction in total phenolic content of up to 96–97%. Similarly, in the same species, Brandolini et al. [44] reported a 76% phenolic decrease, mainly due to phenylethanoid (−95%) and flavonoid (−71%) loss, while phenolic acids (−57%) were less affected. Estivi et al. [70] described that after debittering with 1.0% sodium chloride or 1.0% citric acid solution, the soluble-free fraction was still the most abundant and represented 51.8–67.0% of the total, while with water-debittering it could be as low as 35.1% due to longer treatment and greater wash-out.
Table 6.
Free phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. mutabilis.
| Reference | [44] | [44] | [93] |
|---|---|---|---|
| bit | deb | deb | |
| n = 3 | n = 3 | n = 33 | |
| Total flavonoids | 2739–2871 | 771–917 | 295–1340 |
| Apigenin der † | 154–200 | 5.27–36.94 | 2.70–27.29 |
| Catechin der | 926–986 | nd–36.49 | |
| Diosmin der | 159–328 | 33–95 | 7.23–168.19 |
| Genistein | 11.88–12.78 | 99–125 | 38.03–352.12 |
| Genistein der | 1280–1375 | 550–723 | 219.57–1062.84 |
| Naringenin der | 108–138 | 5.94–7.42 | nd–11.58 |
| Total phenylethanoids | 494–1148 | 7.70–28.39 | |
| Tyrosol | 398–1044 | 2.17–19.29 | nd–49.21 |
| Tyrosol der | 96–138 | 5.09–9.09 | nd–60.27 |
| Total phenolic acids | 64–71 | 24–37 | |
| Cinnamic der | 6.39–30.39 | 5.75–14.36 | nd–11.92 |
| p-hydroxybenzoic der | 1.99–12.38 | 0.87–8.02 | |
| 2,4-hydroxybenzoic der | 4.04–4.53 | 7.73–14.45 | |
| Vanillic der | 24–45 | 0.54–9.11 | |
| Total free phenols | 3298–4025 | 803–956 | 341–1393 |
nd: not detected, i.e., below the detection limit; † der, derivative.
Table 7.
Conjugated and bound phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
Table 7.
Conjugated and bound phenolics (mg/kg DM) of bitter (bit) and debittered (deb) seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
| Conjugated | Bound | |||||||
|---|---|---|---|---|---|---|---|---|
| Reference | [70] | [70] | [70] | [44] | [44] | [70] | [70] | [70] |
| deb * | deb ** | deb *** | bit | deb * | deb * | deb ** | deb *** | |
| L. albus | L. albus | L. albus | L. mutabilis | L. mutabilis | L. albus | L. albus | L. albus | |
| n = 2 | n = 2 | n = 2 | n = 3 | n = 3 | n = 2 | n = 2 | n = 2 | |
| Total flavonoids | 78.99–82.08 | 70.26–85.28 | 118.72–118.94 | 82–109 | 106–122 | 82.69–115.34 | 101.19–124.4 | 59.23–110.17 |
| Apigenin der † | nd–39.49 | nd–0.52 | nd | 10.20–13.13 | 12.26–14.68 | 0.15–1.77 | 0.42–0.84 | 1.35–6.01 |
| Catechin der | nd–0.41 | nd–0.88 | 3.54–4.06 | 19–31 | 14–17 | 23.94–26.24 | 27.37–28.25 | 8.74–11.7 |
| Genistein | nd–2.68 | nd–3.15 | 1.55–4.17 | 9.83–14.22 | nd–2.13 | nd–1.75 | nd–1.77 | |
| Genistein der | 33.63–73.04 | 63.22–82.8 | 105.62–111.76 | 40–59 | 60–76 | 40.41–78.23 | 53.71–83.44 | 29.23–83.91 |
| Naringenin der | 5.46–6.36 | 1.97–3.01 | 2.08–4.87 | 7.82–9.58 | 5.75–6.95 | 10.89–14.27 | 11.42–18.39 | 11.44–15.25 |
| Total phenolic acids | 108–145 | 54–72 | ||||||
| Cinnamic | 2.57–3.43 | 6.53–7.99 | ||||||
| p-coumaric | 0.08–0.16 | 0.01–0.23 | ||||||
| Ferulic | 1.66–2.16 | 1.56–2.31 | ||||||
| m-hydroxybenzoic der | 3.08–9.64 | 3.64–9.42 | ||||||
| m-hydroxybenzoic | 3.71–5.24 | 6.10–11.05 | ||||||
| p-hydroxybenzoic der | 55–70 | 17–23 | ||||||
| p-hydroxybenzoic | 4.81–6.87 | 2.40–3.44 | ||||||
| Salicylic der | 0.66–0.71 | 0.42–0.48 | ||||||
| Syringic der | 19–31 | 5.72–7.28 | ||||||
| Vanillic | 8.62–20.12 | 7.82–9.58 | ||||||
| Total phenols | 190–254 | 161–187 | 83 | 101 | 59 |
nd: not detected, i.e., below the detection limit; † der, derivative; * debittered with H2O; ** debittered with 1%NaCl; *** debittered with 1% citric acid.
7. Conjugated and Bound Phenolics
As mentioned above, the available information on the conjugated and bound phenolics of lupins is scant. Estivi et al. [70], testing the effects of different debittering methods on phenolic compounds, observed that the soluble-conjugated and the insoluble-bound fractions represented 17.1–27.0% and 11.0–37.9% of the total, respectively. All the conjugated compounds detected belonged to the flavonoids (Table 7) and were apigenin der, catechin der, genistein, genistein der and naringenin der. Water debittering led to a lower conjugated phenolics content (79 mg/kg DM) than 1% saline solution (85 mg/kg) and, especially, 1% citric acid solution (119 mg/kg). Apigenin der and naringenin der were better preserved by the water method, and genistein der by the other two methods.
Brandolini et al. [44], studying the effect of processing in bitter seeds of L. mutabilis, found the flavonoids apigenin der, catechin der, genistein, genistein der (the most abundant flavonoid) and naringenin der in the bound fraction, as well as the phenolic acids cinnamic, p-coumaric, ferulic, m-hydroxybenzoic der, p-hydroxybenzoic der (the most abundant phenolic acid), salicylic der, syringic der and vanillic (Table 7). Debittering did not significantly alter the flavonoids (from 82–109 to 106–122 mg/kg DM), marginally decreasing the content of catechin der and naringenin der and slightly increasing those of apigenin, genistein and genistein der. On the other hand, the phenolic acids were halved (from 108–145 to 54–72 mg/kg DM), due to a strong reduction in syringic der and p-hydroxybenzoic der, p-hydroxybenzoic and vanillic acid, not balanced by a slight increase in m-hydroxybenzoic and cinnamic acids. Estivi et al. [70] found the same flavonoids (Table 7) in their debittered samples of L. albus and observed that the 1% NaCl solution better preserved them, followed by water and 1% citric acid solution (101, 83 and 59 mg/kg DM, respectively).
8. Influence of Technological Processes
Germination is an easy and useful process to increase the nutritional value of legumes and cereals; additionally, it causes a reduction in the content of alkaloids, phytynians, and oligosaccharides [98].
Rumiyati et al. [99] studied the effect of germination of Australian sweet lupin (ASL) seeds for nine days and noticed that the total phenolic contents and the antioxidant activity of germinated flour were significantly increased (700 and 1400%, respectively) following germination. Additionally, the concentration of phytosterols and the antioxidant activity of oil extracts from germinated ASL flour also increased significantly (300 and 800%, respectively). Similarly, Dueñas et al. [96] found that nine-day germination modified the quantitative and qualitative phenolic composition of lupin (Lupinus angustifolius) seeds with a significant increase in flavonoids (63% after nine days), total phenolic compounds (84%) and antioxidant activity. After germinating L. albus and L. angustifolius seeds for two weeks, Andor et al. [100] observed a significant increase in cinnamic acid derivatives (in particular, caffeic acid: 160% and 298%, respectively) and genistein (120% and 157%). These results are in close agreement with those of Fernández-Orozco et al. [101], who found an increase of 53% of total phenols in lupin sprouts after nine days, and Frías et al. [102], who observed an enhancement in the antioxidant activity of white lupin (Lupinus albus) after a nine-day germination and an increase in vitamins E and C content.
Estivi et al. [69] germinated white lupin seeds for up to six days and observed that total carotenoid (mainly lutein) content increased up to 12-fold in comparison to ungerminated seeds. On the other hand, the total tocol content was almost unchanged; however, while before germination γ-tocopherol represented 95–99%, during germination, the α-tocopherol increased from not-quantifiable up to 74.8 mg/kg (37%), while the γ-tocopherol decreased progressively. Therefore, this shift in composition improves vitamin E activity.
The solid-state fermentation of L. angustifolius flour with different bacteria (Bifidobacterium animalis subsp. lactis DSM10140, B. longum subsp. longum DSM20097, B. breve DSM20213, L. plantarum DSM2648, L. plantarum KX881779 and L. reuteri KX88177), performed up to 72 h, always led to a significant increase in both total phenolic content (TPC) and antioxidant capacity [103,104]. Likewise, Łopusiewicz et al. [105] observed an increase in TPC (almost two-fold) and antioxidant capacity in lupin beverages fermented for 21 days with commercial kefir grains. Khan et al. [106] explored a combination of germination and fermentation of L. angustifolius seeds with a tempeh starter fungus (Rhizopus sp.) and found that a 12-h germination increased the TPC from 4.8 to 10.9 mg gallic acid equivalent/100 g DM, while fermentation of the 12-h germinated seeds further improved the TPC to 47.7 mg gallic acid equivalent/100 g DM; the antioxidant capacity improved accordingly.
Brandolini et al. [44] examined the effects of different processing techniques (extrusion and spray-drying) on the carotenoids, tocols and phenolic compounds (mainly free phenols) of debittered seeds of three Andean lupins. They observed that the extrusion did not modify the content of tocopherols, slightly increased that of phenolics and marginally reduced (14.5%) that of carotenoids, while spray-drying diminished tocopherols, carotenoids and phenolics (30.0, 35.4 and 48.4%), mainly because of processing conditions; the coating agent dilution effect was minimal.
As mentioned in the introduction, lupin flour can be used to improve the nutritional profile of food products. However, the effect of thermal treatment on phenolics and other antioxidant compounds during food manufacturing has been scarcely studied. Rumiyani et al. [107] observed that muffins baked at 190 °C for 25 min after the addition of 4% or 6% lupin flour had greater total phenolic content and antioxidant activity than their batters and attributed this result to both the release of phenolic compounds from the cellular structures and the formation of phenolic products from thermal degradation. On the other hand, Villarino et al. [108] found that carotenoid recovery in ASL-wheat bread was significantly inferior to the raw composite flours, in the order of 15% (zeaxanthin), 24% (lutein), 48% (β-carotene) and 71% (α-carotene), corresponding to a 62% loss in total carotenoids. During dough mixing in the presence of water and oxygen, lipoxygenases (LOX) oxidise the polyunsaturated fatty acids, causing the degradation of the antioxidant carotenoids. ASL has a high LOX capacity, and this may be the reason behind the low recovery rates of these carotenoids.
9. Conclusions
Lupins are a very promising alternative to animals as a protein source and have an outstanding content of antioxidant compounds. All four cropped species are good sources of proteins, dietary fibre and lipids, rich in unsaturated fatty acids, as well as of micro- and macro-elements, vitamins and antioxidants. Minor differences in antioxidant composition are reported among species. Carotenoids are more abundant in L. angustifolius and L. albus, tocopherols in L. mutabilis and L. albus and phenolics, mainly free flavonoids, in L. mutabilis; conjugated and bound phenolics are scarce and therefore are hardly studied.
The content of all these antioxidants is significantly increased by germination and fermentation. Technological treatments (e.g., debittering, extrusion, spray-drying and baking) may induce a partial loss of these compounds, which, however, are still present in relevant quantities in the end products.
Approaches to increase the availability of lupin antioxidants may include choosing the most promising accessions, adopting optimised debittering methods (not needed for the low-alkaloid varieties) that limit leaching of valuable compounds while also saving time and water consumption, germinating the seeds, selectively fermenting the flours and optimising the processing conditions. Major shortcomings in current knowledge are the limited information about the effect of processing on lupin antioxidants and the complete absence of data on their bioaccessibility, to be studied by appropriate digestion protocols on lupin-enriched ready-to-eat foods.
Author Contributions
Conceptualisation and writing—original draft preparation, A.B. and A.H. visualisation, A.G. and L.E.; investigation, A.B., L.E. and A.G.; supervision, A.B. and A.H.; writing—review and editing, A.B., L.E., A.G. and A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reinhard, H.; Rupp, H.; Sager, F.; Streule, M.; Zoller, O. Quinolizidine Alkaloids and Phomopsins in Lupin Seeds and Lupin Containing Food. J. Chromatogr. A 2006, 1112, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Albuja-Vaca, D.; Yépez, C.; Vernaza, M.G.; Navarrete, D. Gluten-Free Pasta: Development of a New Formulation Based on Rice and Lupine Bean Flour (Lupinus mutabilis) Using a Mixture-Process Design. Food Sci. Technol. 2019, 40, 408–414. [Google Scholar] [CrossRef]
- Al-Saedi, N.; Agarwal, M.; Ma, W.; Islam, S.; Ren, Y. Proteomic Characterisation of Lupin (Lupinus angustifolius) Milk as Influenced by Extraction Techniques, Seed Coat and Cultivars. Molecules 2020, 25, 1782. [Google Scholar] [CrossRef]
- Güémes-Vera, N.; Peña-Bautista, R.J.; Jiménez-Martínez, C.; Dávila-Ortiz, G.; Calderón-Domínguez, G. Effective Detoxification and Decoloration of Lupinus mutabilis Seed Derivatives, and Effect of These Derivatives on Bread Quality and Acceptance. J. Sci. Food Agr. 2008, 88, 1135–1143. [Google Scholar] [CrossRef]
- Jayasena, V.; Leung, P.P.Y.; Nasar-Abbas, S.M. Effect of Lupin Flour Substitution on the Quality and Sensory Acceptability of Instant Noodles. J. Food Qual. 2010, 33, 709–727. [Google Scholar] [CrossRef]
- Jayasena, V.; Khu, W.S.; Nasar-Abbas, S.M. The Development and Sensory Acceptability of Lupin-Based Tofu. J. Food Qual. 2010, 33, 85–97. [Google Scholar] [CrossRef]
- Özcan, M.M.; İpek, D.; Ghafoor, K.; Al Juhaimi, F.; Uslu, N.; Babiker, E.E.; Mohamed Ahmed, I.A.; Alsawmahi, O.N. Physico-Chemical and Sensory Properties of Chips Produced Using Different Lupin (Lupinus albus L.) Flour Formulations and Cooking Methods. Int. J. Food Sci. Technol. 2021, 56, 2780–2788. [Google Scholar] [CrossRef]
- Yaver, E.; Bilgiçli, N. Effect of Ultrasonicated Lupin Flour and Resistant Starch (Type 4) on the Physical and Chemical Properties of Pasta. Food Chem. 2021, 357, 129758. [Google Scholar] [CrossRef]
- Rodríguez, G.; Aguirre, E.; Córdova-Chang, A.; Muñoz-Saenz, J.C.; Baquerizo, M.; Brandolini, A.; Villanueva, E.; Hidalgo, A. Modification of the Nutritional Quality and Oxidative Stability of Lupin (Lupinus mutabilis Sweet) and Sacha Inchi (Plukenetia Volubilis L.) Oil Blends. Molecules 2022, 27, 7315. [Google Scholar] [CrossRef]
- Pascual-Chagman, G.; Santa-Cruz-Olivos, J.; Hidalgo, A.; Benavente, F.; Pérez-Camino, M.C.; Sotelo-Mendez, A.; Paucar-Menacho, L.M.; Encina-Zelada, C.R.; Pascual-Chagman, G.; Santa-Cruz-Olivos, J.; et al. Aceite de Lupinus mutabilis Obtenido Por Prensa Expeller: Análisis de Rendimiento, Caracterización Fisicoquímica, Capacidad Antioxidante, Ácidos Grados y Estabilidad Oxidativa. Sci. Agropecu. 2021, 12, 219–227. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Ereifej, K.; Gammoh, S.; Kubow, S.; Tawalbeh, D. Preparation of Mayonnaise from Extracted Plant Protein Isolates of Chickpea, Broad Bean and Lupin Flour: Chemical, Physiochemical, Nutritional and Therapeutic Properties. J. Food Sci. Technol. 2017, 54, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Kasapis, S.; Farahnaky, A. Lupin Protein: Isolation and Techno-Functional Properties, a Review. Food Hydrocoll. 2021, 112, 106318. [Google Scholar] [CrossRef]
- Paraskevopoulou, A.; Provatidou, E.; Tsotsiou, D.; Kiosseoglou, V. Dough Rheology and Baking Performance of Wheat Flour–Lupin Protein Isolate Blends. Food Res. Int. 2010, 43, 1009–1016. [Google Scholar] [CrossRef]
- Arnoldi, A.; Boschin, G.; Zanoni, C.; Lammi, C. The Health Benefits of Sweet Lupin Seed Flours and Isolated Proteins. J. Funct. Foods 2015, 18, 550–563. [Google Scholar] [CrossRef]
- Okagu, I.U.; Ndefo, J.C.; Aham, E.C.; Obeme-Nmom, J.I.; Agboinghale, P.E.; Aguchem, R.N.; Nechi, R.N.; Lammi, C. Lupin-Derived Bioactive Peptides: Intestinal Transport, Bioavailability and Health Benefits. Nutrients 2021, 13, 3266. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, A.R.; El-Nashar, H.A.S.; Younis, T.; Mangat, M.A.; Shahzadi, M.; Ul Haq, A.S.; El-Shazly, M. Genus Lupinus (Fabaceae): A Review of Ethnobotanical, Phytochemical and Biological Studies. J. Pharm. Pharmacol. 2022, 74, 1700–1717. [Google Scholar] [CrossRef] [PubMed]
- Kohajdová, Z.; Karovičová, J.; Schmidt, Š. Lupin Composition and Possible Use in Bakery—A Review. Czech J. Food Sci. 2011, 29, 203–211. [Google Scholar] [CrossRef]
- Ruiz-López, M.A.; Barrientos-Ramírez, L.; García-López, P.M.; Valdés-Miramontes, E.H.; Zamora-Natera, J.F.; Rodríguez-Macias, R.; Salcedo-Pérez, E.; Bañuelos-Pineda, J.; Vargas-Radillo, J.J. Nutritional and Bioactive Compounds in Mexican Lupin Beans Species: A Mini-Review. Nutrients 2019, 11, 1785. [Google Scholar] [CrossRef]
- Briceño Berru, L.; Glorio-Paulet, P.; Basso, C.; Scarafoni, A.; Camarena, F.; Hidalgo, A.; Brandolini, A. Chemical Composition, Tocopherol and Carotenoid Content of Seeds from Different Andean Lupin (Lupinus mutabilis) Ecotypes. Plant Foods Hum. Nutr. 2021, 76, 98–104. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Linnemann, A.R.; Nout, M.J.R.; Koziol, M.; van Boekel, M.A.J.S. Lupinus mutabilis: Composition, Uses, Toxicology, and Debittering. Crit. Rev. Food Sci. Nutr. 2016, 56, 1454–1487. [Google Scholar] [CrossRef]
- Czubinski, J.; Wroblewska, K.; Czyzniejewski, M.; Górnaś, P.; Kachlicki, P.; Siger, A. Bioaccessibility of Defatted Lupin Seed Phenolic Compounds in a Standardized Static In Vitro Digestion System. Food Res. Int. 2019, 116, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Musco, N.; Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Infascelli, F.; Grazioli, R.; Lo Presti, V.; Gresta, F.; Chiofalo, B. Comparison of Nutritional and Antinutritional Traits among Different Species (Lupinus albus L., Lupinus luteus L., Lupinus angustifolius L.) and Varieties of Lupin Seeds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1227–1241. [Google Scholar] [CrossRef]
- Prusinski, J. White Lupin (Lupinus albus L.)—Nutritional and Health Values in Human Nutrition—A Review. Czech J. Food Sci. 2017, 35, 95–105. [Google Scholar] [CrossRef]
- Foley, R.C.; Gao, L.L.; Spriggs, A.; Soo, L.Y.C.; Goggin, D.E.; Smith, P.M.C.; Atkins, C.A.; Singh, K.B. Identification and Characterisation of Seed Storage Protein Transcripts from Lupinus angustifolius. BMC Plant Biol. 2011, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Gulewicz, P.; Martínez-Villaluenga, C.; Frias, J.; Ciesiołka, D.; Gulewicz, K.; Vidal-Valverde, C. Effect of Germination on the Protein Fraction Composition of Different Lupin Seeds. Food Chem. 2008, 107, 830–844. [Google Scholar] [CrossRef]
- FAO/WHO/ONU. Energy and Protein Requirements. Report of a Joint FAO/WHO/UNU Expert Consultation; WHO: Geneva, Switzerland, 1985. [Google Scholar]
- Sujak, A.; Kotlarz, A.; Strobel, W. Compositional and Nutritional Evaluation of Several Lupin Seeds. Food Chem. 2006, 98, 711–719. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Bez, J.; Petersen, I.L.; Joehnke, M.S.; Detzel, A.; Busch, M.; Krueger, M.; Ispiryan, L.; O’Mahony, J.A.; Arendt, E.K.; et al. Techno-Functional, Nutritional and Environmental Performance of Protein Isolates from Blue Lupin and White Lupin. Foods 2020, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Sironi, E.; Vidal-Valverde, C.; Duranti, M. Effects of Oligosaccharide Removing Procedure on the Protein Profiles of Lupin Seeds. Eur. Food Res. Technol. 2006, 223, 691–696. [Google Scholar] [CrossRef]
- Ciesiołka, D.; Gulewicz, P.; Martinez-Villaluenga, C.; Pilarski, R.; Bednarczyk, M.; Gulewicz, K. Products and Biopreparations from Alkaloid-Rich Lupin in Animal Nutrition and Ecological Agriculture. Folia Biol. 2005, 53, 59–66. [Google Scholar] [CrossRef]
- Siger, A.; Grygier, A.; Czubinski, J. Comprehensive Characteristic of Lipid Fraction as a Distinguishing Factor of Three Lupin Seed Species. J. Food Compos. Anal. 2023, 115, 104945. [Google Scholar] [CrossRef]
- Boschin, G.; Arnoldi, A. Legumes Are Valuable Sources of Tocopherols. Food Chem. 2011, 127, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Erbaş, M.; Certel, M.; Uslu, M.K. Some Chemical Properties of White Lupin Seeds (Lupinus albus L.). Food Chem. 2005, 89, 341–345. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E. Nutritional, Rheological and Sensory Evaluation of Lupinus mutabilis Food Products—A Review. Czech J. Food Sci. 2019, 37, 301–311. [Google Scholar] [CrossRef]
- Carvalho, I.S.; Chaves, M.; Pinto Ricardo, C. Influence of Water Stress on the Chemical Composition of Seeds of Two Lupins (Lupinus albus and Lupinus mutabilis). J. Agron. Crop Sci. 2005, 191, 95–98. [Google Scholar] [CrossRef]
- Al-Amrousi, E.F.; Badr, A.N.; Abdel-Razek, A.G.; Gromadzka, K.; Drzewiecka, K.; Hassanein, M.M.M. A Comprehensive Study of Lupin Seed Oils and the Roasting Effect on Their Chemical and Biological Activity. Plants 2022, 11, 2301. [Google Scholar] [CrossRef] [PubMed]
- Sbihi, H.M.; Nehdi, I.A.; Tan, C.P.; Al-Resayes, S.I. Bitter and Sweet Lupin (Lupinus albus L.) Seeds and Seed Oils: A Comparison Study of Their Compositions and Physicochemical Properties. Ind. Crops Prod. 2013, 49, 573–579. [Google Scholar] [CrossRef]
- Yorgancilar, M.; Bilgiçli, N. Chemical and Nutritional Changes in Bitter and Sweet Lupin Seeds (Lupinus albus L.) during Bulgur Production. J. Food Sci. Technol. 2014, 51, 1384–1389. [Google Scholar] [CrossRef] [PubMed]
- Czubiński, J.; Siger, A. Physicochemical Characteristics and Technological Properties of Lupinus mutabilis Oil. Eur. J. Lipid Sci. Technol. 2023, 125, 2200200. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Rayas-Duarte, P. Composition of Lupinus albus. Cereal Chem. 1995, 72, 643–647. [Google Scholar]
- Czubinski, J.; Grygier, A.; Siger, A. Lupinus mutabilis Seed Composition and Its Comparison with Other Lupin Species. J. Food Compos. Anal. 2021, 99, 103875. [Google Scholar] [CrossRef]
- Zelalem, K.; Chandravanshi, B. Levels of Essential and Non-Essential Elements in Raw and Processed Lupinus albus L. (White Lupin, Gibto) Cultivated in Ethiopia. Afr. J. Food Agric. Nutr. Dev. 2014, 14, 9215–9235. [Google Scholar] [CrossRef]
- Trugo, L.C.; von Baer, D.; von Baer, E. Lupin. In Encyclopedia of Food Sciences and Nutrition; Benjamin, C., Ed.; Academic Press: Oxford, UK, 2003; pp. 3623–3629. [Google Scholar]
- Brandolini, A.; Glorio-Paulet, P.; Estivi, L.; Locatelli, N.; Cordova-Ramos, J.S.; Hidalgo, A. Tocopherols, Carotenoids and Phenolics Changes during Andean Lupin (Lupinus mutabilis Sweet) Seeds Processing. J. Food Compos. Anal. 2022, 106, 104335. [Google Scholar] [CrossRef]
- Saastamoinen, M.; Eurola, M.; Hietaniemi, V. The Chemical Quality of Some Legumes, Peas, Fava Beans, Blue and White Lupins and Soybeans Cultivated in Finland. J. Agric. Sci. Technol. 2013, B3, 92–100. [Google Scholar]
- El-Adawy, T.A.; Rahma, E.H.; El-Bedawey, A.A.; Gafar, A.F. Nutritional Potential and Functional Properties of Sweet and Bitter Lupin Seed Protein Isolates. Food Chem. 2001, 74, 455–462. [Google Scholar] [CrossRef]
- Jiménez-Martínez, C.; Hernández-Sánchez, H.; Alvárez-Manilla, G.; Robledo-Quintos, N.; Martínez-Herrera, J.; Dávila-Ortiz, G. Effect of Aqueous and Alkaline Thermal Treatments on Chemical Composition and Oligosaccharide, Alkaloid and Tannin Contents of Lupinus Campestris Seeds. J. Sci. Food Agr. 2001, 81, 421–428. [Google Scholar] [CrossRef]
- Kroc, M.; Czepiel, K.; Wilczura, P.; Mokrzycka, M.; Święcicki, W. Development and Validation of a Gene-Targeted DCAPS Marker for Marker-Assisted Selection of Low-Alkaloid Content in Seeds of Narrow-Leafed Lupin (Lupinus angustifolius L.). Genes 2019, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Uauy, R.; Gattas, V.; Yañez, E. Sweet Lupins in Human Nutrition. In World Review of Nutrition and Dietetics; Simopoulos, A.P., Ed.; Karger Publishers: Basel, Switzerland, 1995; Volume 77, pp. 75–88. [Google Scholar]
- Erbas, M. The Effects of Different Debittering Methods on the Production of Lupin Bean Snack from Bitter Lupinus albus L. Seeds. J. Food Qual. 2010, 33, 742–757. [Google Scholar] [CrossRef]
- Córdova-Ramos, J.S.; Glorio-Paulet, P.; Camarena, F.; Brandolini, A.; Hidalgo, A. Andean Lupin (Lupinus mutabilis Sweet): Processing Effects on Chemical Composition, Heat Damage, and In Vitro Protein Digestibility. Cereal Chem. 2020, 97, 827–835. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Van Boekel, M.J.A.S.; Koziol, M.; Nout, M.J.R.; Linnemann, A.R. Effect of Processing on the Diffusion of Alkaloids and Quality of Lupinus mutabilis Sweet. J. Food Process Preserv. 2014, 38, 1461–1471. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Nout, M.J.R.; van Boekel, M.A.J.S.; Koziol, M.; Linnemann, A.R. Modelling of the Aqueous Debittering Process of Lupinus mutabilis Sweet. LWT—Food Sci. Technol. 2013, 53, 507–516. [Google Scholar] [CrossRef]
- Gross, U.; Galindo, R.G.; Schoeneberger, H. The Development and Acceptability of Lupine (Lupinus mutabilis) Products. Plant Foods Hum. Nutr. 1983, 32, 155–164. [Google Scholar] [CrossRef]
- Fontanari, G.G.; Batistuti, J.P.; Da Cruz, R.J.; Saldiva, P.H.N.; Arêas, J.A.G. Cholesterol-Lowering Effect of Whole Lupin (Lupinus albus) Seed and Its Protein Isolate. Food Chem. 2012, 132, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.A.; Mohamed, E.A.; Yagoub, A.E.A.; Mohamed, A.R.; Babiker, E.E. Effect of Processing Methods on Alkaloids, Phytate, Phenolics, Antioxidants Activity and Minerals of Newly Developed Lupin (Lupinus albus L.) Cultivar. J. Food Process Preserv. 2017, 41, e12960. [Google Scholar] [CrossRef]
- Villacrés, E.; Quelal, M.B.; Fernández, E.; Garcìa, G.; Cueva, G.; Rosell, C.M. Impact of Debittering and Fermentation Processes on the Antinutritional and Antioxidant Compounds in Lupinus mutabilis Sweet. LWT—Food Sci. Technol. 2020, 131, 109745. [Google Scholar] [CrossRef]
- De Cortes Sánchez, M.; Altares, P.; Pedrosa, M.M.; Burbano, C.; Cuadrado, C.; Goyoaga, C.; Muzquiz, M.; Jiménez-Martínez, C.; Dávila-Ortiz, G. Alkaloid Variation during Germination in Different Lupin Species. Food Chem. 2005, 90, 347–355. [Google Scholar] [CrossRef]
- Jiménez-Martínez, C.; Hernández-Sánchez, H.; Dávila-Ortiz, G. Diminution of Quinolizidine Alkaloids, Oligosaccharides and Phenolic Compounds from Two Species of Lupinus and Soybean Seeds by the Effect of Rhizopus oligosporus. J. Sci. Food Agr. 2007, 87, 1315–1322. [Google Scholar] [CrossRef]
- Jiménez Martínez, C.; Mora Escobedo, R.; Cardador Martínez, A.; Muzquiz, M.; Mercedes, M.P.M.; Gloria, D.O. Effect of Aqueous, Acid, and Alkaline Thermal Treatments on Antinutritional Factors Content and Protein Quality in Lupinus Campestris Seed Flour. J. Agr. Food Chem. 2010, 58, 1741–1745. [Google Scholar] [CrossRef]
- Estivi, L.; Buratti, S.; Fusi, D.; Benedetti, S.; Rodríguez, G.; Brandolini, A.; Hidalgo, A. Alkaloid Content and Taste Profile Assessed by Electronic Tongue of Lupinus albus Seeds Debittered by Different Methods. J. Food Compos. Anal. 2022, 114, 104810. [Google Scholar] [CrossRef]
- Van Den Berg, H.; Faulks, R.; Granado, H.F.; Hirschberg, J.; Olmedilla, B.; Sandmann, G.; Southon, S.; Stahl, W. The Potential for the Improvement of Carotenoid Levels in Foods and the Likely Systemic Effects. J. Sci. Food Agr. 2000, 80, 880–912. [Google Scholar] [CrossRef]
- Britton, G. Structure and Properties of Carotenoids in Relation to Function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef]
- Zile, M.H. Vitamin A and Embryonic Development: An Overview. J. Nutr. 1998, 128, 455S–458S. [Google Scholar] [CrossRef]
- Krinsky, N.I. The Biological Properties of Carotenoids. Pure Appl. Chem. 1994, 66, 1003–1010. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Milla, R.; Martín-Robles, N.; Arc, E.; Kranner, I.; Becerril, J.M.; García-Plazaola, J.I. Side-Effects of Domestication: Cultivated Legume Seeds Contain Similar Tocopherols and Fatty Acids but Less Carotenoids than Their Wild Counterparts. BMC Plant Biol. 2014, 14, 1599. [Google Scholar] [CrossRef] [PubMed]
- El-Difrawi, E.A.; Hudson, B.J.F. Identification and Estimation of Carotenoids in the Seeds of Four Lupinus Species. J. Sci. Food Agr. 1979, 30, 1168–1170. [Google Scholar] [CrossRef]
- Siger, A.; Michalak, M.; Bąkowska, E.; Dwiecki, K.; Nogala-Kałucka, M.; Grześ, B.; Piasecka-Kwiatkowska, D. The Effect of the Genotype-Environment Interaction on the Concentration of Carotenoids, Tocochromanol, and Phenolic Compounds in Seeds of Lupinus angustifolius Breeding Lines. J. Food Compos. Anal. 2023, 123, 105511. [Google Scholar] [CrossRef]
- Estivi, L.; Pascual Chagman, G.J.; Santa Cruz Olivos, J.E.; Savasi, P.; Brandolini, A.; Hidalgo, A. Changes in Colour, Tocopherols and Carotenoids during the Germination of Lupin Seeds. J. Food Compos. Anal. 2023, 124, 105682. [Google Scholar] [CrossRef]
- Estivi, L.; Fusi, D.; Brandolini, A.; Hidalgo, A. Effect of Debittering with Different Solvents and Ultrasound on Carotenoids, Tocopherols, and Phenolics of Lupinus albus Seeds. Antioxidants 2022, 11, 2481. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, R.; Matsushita, S. Products Formed by Photosensitized Oxidation of Tocopherols. Agric. Biol. Chem. 1979, 43, 2151–2156. [Google Scholar] [CrossRef]
- Goffman, F.D.; Böhme, T. Relationship between Fatty Acid Profile and Vitamin E Content in Maize Hybrids (Zea mays L.). J. Agr. Food Chem. 2001, 49, 4990–4994. [Google Scholar] [CrossRef]
- Bramley, P.M.; Elmadfa, I.; Kafatos, A.; Kelly, F.J.; Manios, Y.; Roxborough, H.E.; Schuch, W.; Sheehy, P.J.A.; Wagner, K.H. Vitamin E. J. Sci. Food Agr. 2000, 80, 913–938. [Google Scholar] [CrossRef]
- Azzi, A. Many Tocopherols, One Vitamin E. Mol. Aspects Med. 2018, 61, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Miller, H.E.; Rigelhof, F.; Marquart, L.; Prakash, A.; Kanter, M. Antioxidant Content of Whole Grain Breakfast Cereals, Fruits and Vegetables. J. Am. Coll. Nutr. 2000, 19, 312S–319S. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Niki, E.; Noguchi, N. Comparative Study on the Action of Tocopherols and Tocotrienols as Antioxidant: Chemical and Physical Effects. Chem. Phys. Lipids 2003, 123, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Theriault, A.; Chao, J.T.; Wang, Q.; Gapor, A.; Adeli, K. Tocotrienol: A Review of Its Therapeutic Potential. Clin. Biochem. 1999, 32, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Zieliński, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. Antioxidant Capacity and Polyphenolic Content of High-Protein Lupin Products. Food Chem. 2009, 112, 84–88. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Manunza, P.; Arnoldi, A.; Boschin, G. Quality of Lupinus albus L. (White Lupin) Seed: Extent of Genotypic and Environmental Effects. J. Agr. Food Chem. 2014, 62, 6539–6545. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free Radicals, Natural Antioxidants, and Their Reaction Mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Quiñones, M.; Miguel, M.; Aleixandre, A. Beneficial Effects of Polyphenols on Cardiovascular Disease. Pharmacol. Res. 2013, 68, 125–131. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and Analysis of Phenolics in Food. J. Chromatogr. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and Metabolism of Polyphenols in the Gut and Impact on Health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound Phenolics in Foods, a Review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Tirdiľová, I.; Vollmannová, A.; Čéryová, S.; Obtulovič, P.; Árvay, J.; Zetochová, E. Impact of 3-Year Period as a Factor on the Content of Biologically Valuable Substances in Seeds of White Lupin. Plants 2022, 11, 2087. [Google Scholar] [CrossRef] [PubMed]
- Ben Hassine, A.; Rocchetti, G.; Zhang, L.; Senizza, B.; Zengin, G.; Mahomoodally, M.F.; Ben-Attia, M.; Rouphael, Y.; Lucini, L.; El-Bok, S. Untargeted Phytochemical Profile, Antioxidant Capacity and Enzyme Inhibitory Activity of Cultivated and Wild Lupin Seeds from Tunisia. Molecules 2021, 26, 3452. [Google Scholar] [CrossRef] [PubMed]
- Ferchichi, N.; Toukabri, W.; Vrhovsek, U.; Nouairi, I.; Angeli, A.; Masuero, D.; Mhamdi, R.; Trabelsi, D. Proximate Composition, Lipid and Phenolic Profiles, and Antioxidant Activity of Different Ecotypes of Lupinus albus, Lupinus luteus and Lupinus angustifolius. J. Food Meas. Charact. 2021, 15, 1241–1257. [Google Scholar] [CrossRef]
- Vollmannova, A.; Lidikova, J.; Musilova, J.; Snirc, M.; Bojnanska, T.; Urminska, D.; Tirdilova, I.; Zetochova, E. White Lupin as a Promising Source of Antioxidant Phenolics for Functional Food Production. J. Food Qual. 2021, 2021, 5512236. [Google Scholar] [CrossRef]
- Karamać, M.; Orak, H.H.; Amarowicz, R.; Orak, A.; Piekoszewski, W. Phenolic Contents and Antioxidant Capacities of Wild and Cultivated White Lupin (Lupinus albus L.) Seeds. Food Chem. 2018, 258, 1–7. [Google Scholar] [CrossRef]
- Magalhães, S.C.Q.; Taveira, M.; Cabrita, A.R.J.; Fonseca, A.J.M.; Valentão, P.; Andrade, P.B. European Marketable Grain Legume Seeds: Further Insight into Phenolic Compounds Profiles. Food Chem. 2017, 215, 177–184. [Google Scholar] [CrossRef]
- Siger, A.; Czubinski, J.; Kachlicki, P.; Dwiecki, K.; Lampart-Szczapa, E.; Nogala-Kalucka, M. Antioxidant Activity and Phenolic Content in Three Lupin Species. J. Food Compos. Anal. 2012, 25, 190–197. [Google Scholar] [CrossRef]
- Estivi, L.; Grassi, S.; Briceño-Berrú, L.; Glorio-Paulet, P.; Camarena, F.; Hidalgo, A.; Brandolini, A. Free Phenolic Compounds, Antioxidant Capacity and FT-NIR Survey of Debittered Lupinus mutabilis Seeds. Processes 2022, 10, 1637. [Google Scholar] [CrossRef]
- Angelino, D.; Cossu, M.; Marti, A.; Zanoletti, M.; Chiavaroli, L.; Brighenti, F.; Del Rio, D.; Martini, D. Bioaccessibility and Bioavailability of Phenolic Compounds in Bread: A Review. Food Funct. 2017, 8, 2368–2393. [Google Scholar] [CrossRef] [PubMed]
- Lampart-Szczapa, E.; Korczak, J.; Nogala-Kalucka, M.; Zawirska-Wojtasiak, R. Antioxidant Properties of Lupin Seed Products. Food Chem. 2003, 83, 279–285. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Estrella, I.; Fernández, D. Germination as a Process to Increase the Polyphenol Content and Antioxidant Activity of Lupin Seeds (Lupinus angustifolius L.). Food Chem. 2009, 117, 599–607. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. Content of Phenolic Compounds and Antioxidant Properties in Seeds of Sweet and Bitter Cultivars of Lupine (Lupinus angustifolius). Nat. Prod. Commun. 2018, 13, 1341–1344. [Google Scholar] [CrossRef]
- Dagnia, S.G.; Petterson, D.S.; Bell, R.R.; Flanagan, F.V. Germination Alters the Chemical Composition and Protein Quality of Lupin Seeds. J. Sci. Food Agr. 1992, 60, 419–423. [Google Scholar] [CrossRef]
- Rumiyati; Jayasena, V.; James, A.P. Total Phenolic and Phytosterol Compounds and the Radical Scavenging Activity of Germinated Australian Sweet Lupin Flour. Plant Foods Hum. Nutr. 2013, 68, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Andor, B.; Danciu, C.; Alexa, E.; Zupko, I.; Hogea, E.; Cioca, A.; Coricovac, D.; Pinzaru, I.; Pǎtraşcu, J.M.; Mioc, M.; et al. Germinated and Ungerminated Seeds Extract from Two Lupinus Species: Biological Compounds Characterization and In Vitro and In Vivo Evaluations. Evid. Based Complement. Alternat. Med. 2016, 2016, 7638542. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Piskula, M.K.; Zielinski, H.; Kozlowska, H.; Frias, J.; Vidal-Valverde, C. Germination as a Process to Improve the Antioxidant Capacity of Lupinus angustifolius L. Var. Zapaton. Eur. Food Res. Technol. 2006, 223, 495–502. [Google Scholar] [CrossRef]
- Frias, J.; Miranda, M.L.; Doblado, R.; Vidal-Valverde, C. Effect of Germination and Fermentation on the Antioxidant Vitamin Content and Antioxidant Capacity of Lupinus albus L. Var. Multolupa. Food Chem. 2005, 92, 211–220. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In Vitro Investigation of Bioactivities of Solid-State Fermented Lupin, Quinoa and Wheat Using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Al-Mheiri, A.; Abushelaibi, A. Cytotoxicity, Antihypertensive, Antidiabetic and Antioxidant Activities of Solid-State Fermented Lupin, Quinoa and Wheat by Bifidobacterium Species: In-Vitro Investigations. LWT 2018, 95, 295–302. [Google Scholar] [CrossRef]
- Łopusiewicz, Ł.; Drozłowska, E.; Trocer, P.; Kwiatkowski, P.; Bartkowiak, A.; Gefrom, A.; Sienkiewicz, M. The Effect of Fermentation with Kefir Grains on the Physicochemical and Antioxidant Properties of Beverages from Blue Lupin (Lupinus angustifolius L.) Seeds. Molecules 2020, 25, 5791. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.E.; Jayasena, V. Development of a Fermented Product with Higher Phenolic Compounds and Lower Anti-Nutritional Factors from Germinated Lupin (Lupinus angustifolius L.). J. Food Process Preserv. 2018, 42, e13843. [Google Scholar] [CrossRef]
- Rumiyati, R.; James, A.P.; Jayasena, V. Effects of Lupin Incorporation on the Physical Properties and Stability of Bioactive Constituents in Muffins. Int. J. Food Sci. Technol. 2015, 50, 103–110. [Google Scholar] [CrossRef]
- Villarino, C.B.J.; Jayasena, V.; Coorey, R.; Chakrabarti-Bell, S.; Johnson, S.K. Nutritional, Health, and Technological Functionality of Lupin Flour Addition to Bread and Other Baked Products: Benefits and Challenges. Crit. Rev. Food Sci. Nutr. 2016, 56, 835–857. [Google Scholar] [CrossRef]
Table 1.
Carotenoid content (mg/kg DM) of bitter, sweet or debittered seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
Table 1.
Carotenoid content (mg/kg DM) of bitter, sweet or debittered seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
| N° Samples | (α + β)-Carotene | β-Cryptoxanthin | Lutein | Violaxanthin | Zeaxanthin | Total Carotenoids | Reference | |
|---|---|---|---|---|---|---|---|---|
| L. albus | ||||||||
| sweet | n = 2 | 6.65–8.77 | 0.06–0.11 | 2.23–3.82 | 8.94–12.70 | [31] | ||
| bitter | n = 3 | 0.39–0.82 | 0.01–0.02 | 0.85–3.74 | 0.65–1.02 | 1.90–5.26 | [19] | |
| debittered * | n = 2 | 2.25–7.86 | 0.18–1.58 | 7.91–9.33 | 1.86–2.30 | 12.20–21.07 | [70] | |
| debittered ** | n = 2 | 2.44–6.89 | 0.11–1.21 | 8.74–9.35 | 1.89–1.95 | 13.23–19.34 | [70] | |
| debittered *** | n = 2 | 2.49–12.01 | 0.42–3.05 | 7.97–8.05 | 1.74–2.06 | 12.7–25.09 | [70] | |
| debittered | n = 3 | 0.39–0.89 | 0.03–0–06 | 0.84–5.07 | 0.34–1.17 | 1.60–7.14 | [19] | |
| L. luteus | ||||||||
| sweet | n = 2 | 0.18–0.25 | 4.30–4–50 | 0.04–0.06 | 1.31–1.81 | 6.12–6.33 | [31] | |
| bitter | n = 1 | 0.14 | 0.01 | 0.75 | 0.09 | 0.99 | [19] | |
| bitter | n = 1 | 0.12 | 3.02 | 0.99 | 4.13 | [66] | ||
| debittered | n = 1 | 0.22 | 0.03 | 0.73 | 0.07 | 1.05 | [19] | |
| L. angustifolius | ||||||||
| sweet | n = 4 | 0.81–6.44 | 11.17–35.95 | 4.90–16.70 | 0.30–3.44 | 17.20–62.52 | [31] | |
| bitter/sweet | n = 55 | 3.6–7.7 | 13.8–30.1 | 2.1–10.8 | 5.6–15.6 | 30.6–56.5 | [68] | |
| bitter | n = 1 | 1.1 | 0.02 | 3.93 | 1.45 | 6.49 | [19] | |
| debittered | 0.86 | 0.05 | 3.39 | 1.25 | 5.55 | [19] | ||
| L. mutabilis | ||||||||
| bitter | n = 2 | 0.10–0.20 | 1.45–1.48 | 0.11–0.13 | 1.70–1.81 | [69] | ||
| bitter | n = 3 | 1.06–1.39 | 0.09–0.11 | 1.15–1.50 | [44] | |||
| bitter | n = 33 | 0.06–0.60 | 0.00–0.03 | 0.43–2.20 | 0.03–0.21 | 0.69–2.89 | [19] | |
| bitter | n = 1 | 0.26 | 3.3 | 0.71 | 4.51 | [41] | ||
| debittered | n = 33 | 0.09–1.22 | 0.02–0.07 | 0.43–2.28 | 0.04–0.19 | 0.68–3.25 | [19] | |
| debittered | n = 3 | 1.04–1.33 | 0.11–0.12 | 1.15–1.45 | [44] |
* debittered with H2O; ** debittered with 1%NaCl; *** debittered with 1% citric acid.
Table 2.
Tocopherol content (mg/kg DM) of bitter, sweet or debittered seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
Table 2.
Tocopherol content (mg/kg DM) of bitter, sweet or debittered seeds of L. albus, L. luteus, L. angustifolius and L. mutabilis.
| N° Samples | α-Tocopherol | β-Tocopherol | γ-Tocopherol | δ-Tocopherol | Total Tocopherols | Reference | |
|---|---|---|---|---|---|---|---|
| L. albus | |||||||
| sweet | n = 2 | 126–130 | 126–130 | [31] | |||
| bitter | n = 3 | 0.88–1.22 | 0.34–0.50 | 109–151 | 1.38–1.94 | 112–154 | [19] |
| bitter/sweet | n = 8 | 121–131 | [79] | ||||
| n = 6 | 0.92–1.65 | 61–130 | 1.05–2.55 | 63–134 | [32] | ||
| bitter | n = 2 | 1.9–4.7 | 201–516 | 2.5–4.1 | 205–525 | [78] | |
| debittered | n = 3 | 1.45–1.76 | 0.29–0.99 | 131–218 | 2.08–2.44 | 136–223 | [19] |
| debittered * | n = 2 | 0.26–0.45 | 1.13–1.36 | 192–234 | 1.99–2.09 | 195–241 | [70] |
| debittered ** | n = 2 | 0.50–0.56 | 0.52–0.85 | 176–214 | 1.61–2.26 | 179–218 | [70] |
| debittered *** | n = 2 | 0.29–0.50 | 0.99–2.64 | 183–208 | 1.56–2.45 | 188–212 | [70] |
| L. luteus | |||||||
| sweet | n = 2 | 86–87 | 86–87 | [31] | |||
| bitter | n = 1 | 0.83 | 1.07 | 187 | 1.26 | 191 | [19] |
| bitter | n = 2 | 2.7–4.8 | 94–112 | 2.4–3.8 | 99–121 | [78] | |
| debittered | n = 1 | 0.5 | 0.89 | 257 | 2.26 | 261 | [19] |
| L. angustifolius | |||||||
| bitter | [69] | ||||||
| sweet | n = 4 | 2.67–5.16 | 61.24–91.52 | 65–97 | [31] | ||
| bitter/sweet | n = 55 | 2.3–7.3 | 82.4–116.7 | 85–123 | [68] | ||
| bitter | n = 1 | 2.08 | 0.24 | 89 | 0.67 | 92 | [19] |
| debittered | n = 1 | 1.6 | 0.97 | 100.4 | 0.27 | 103 | [19] |
| L. mutabilis | |||||||
| bitter | n = 2 | 0.66–0.82 | 0.90–1.47 | 165.8–213.6 | 1.86–2.24 | 169–218 | [69] |
| bitter | n = 33 | 0.27–1.24 | 0.70–3.77 | 169–245 | 0.43–3.36 | 172–250 | [19] |
| bitter | n = 3 | 0.51–0.61 | 0.93–1.60 | 224–228 | 1.66–2.81 | 227–233 | [44] |
| bitter | n = 1 | 2 | 171 | 173 | [41] | ||
| n = 7 | 1.42–5.51 | 69–90 | 74–95 | [32] | |||
| debittered | n = 33 | 0.18–1.72 | 0.75–5.06 | 223–370 | 1.33–5.01 | 227–378 | [19] |
| debittered | n = 3 | 0.63–0.72 | 1.55–2.72 | 275–363 | 2.37–3.02 | 280–379 | [44] |
* debittered with H2O; ** debittered with 1%NaCl; *** debittered with 1% citric acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Estivi, L.; Brandolini, A.; Gasparini, A.; Hidalgo, A. Lupin as a Source of Bioactive Antioxidant Compounds for Food Products. Molecules 2023, 28, 7529. https://doi.org/10.3390/molecules28227529
AMA Style
Estivi L, Brandolini A, Gasparini A, Hidalgo A. Lupin as a Source of Bioactive Antioxidant Compounds for Food Products. Molecules. 2023; 28(22):7529. https://doi.org/10.3390/molecules28227529
Chicago/Turabian StyleEstivi, Lorenzo, Andrea Brandolini, Andrea Gasparini, and Alyssa Hidalgo. 2023. "Lupin as a Source of Bioactive Antioxidant Compounds for Food Products" Molecules 28, no. 22: 7529. https://doi.org/10.3390/molecules28227529