
Chemical Composition and Antioxidant Potential of Five Algae Cultivated in Fully Controlled Closed Systems
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Proximate Composition
2.2. Elemental Composition of Seaweeds
2.3. Dietary Fiber Determination and Polysaccharide Characterization
2.4. Fatty Acid Composition
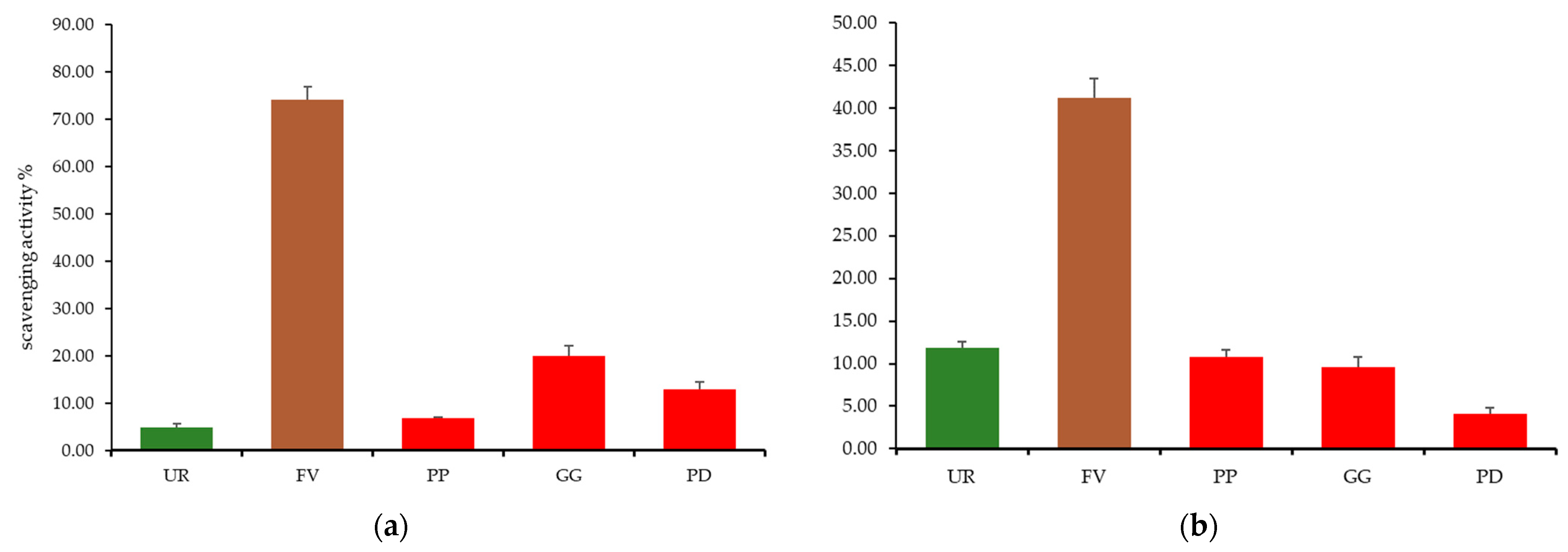
2.5. Total Phenolic Content, Free Radical Scavenging Activity, and Total Antioxidant Activity
3. Materials and Methods
3.1. Specimens of Seaweeds
3.2. Chemical Characterization of Seaweeds
3.2.1. Proximate Composition
3.2.2. Elemental Composition
3.2.3. Analysis of Fatty Acids
3.3. Polysaccharide Characterization
3.4. Determination of Total Phenolic Content and Antioxidant Activity
3.4.1. Extract Preparation
3.4.2. Phenolic Content
3.4.3. DPPH Free Radical Scavenging Activity
3.4.4. Total Antioxidant Capacity
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Chapman, R.L. Algae: The world’s most important “plants”—An introduction. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 5–12. [Google Scholar] [CrossRef]
- Pérez-Lloréns, J.L.; Mouritsen, O.G.; Rhatigan, P.; Cornish, M.L.; Critchley, A.T. Seaweeds in mythology, folklore, poetry, and life. J. Appl. Phycol. 2020, 32, 3157–3182. [Google Scholar] [CrossRef]
- Umen, J.; Coelho, S. Algal Sex Determination and the Evolution of Anisogamy. Annu. Rev. Microbiol. 2019, 73, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Rodríguez, A.G.; Juárez-Portilla, C.; Olivares-Bañuelos, T.; Zepeda, R.C. Anticancer activity of seaweeds. Drug Discov. Today 2018, 23, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Dini, I. The Potential of Algae in the Nutricosmetic Sector. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Freitas, A.C.; Pereira, L.; Rodrigues, D.; Carvalho, A.; Panteleitchouk, T.; Gomes, A.; Duarte, A. Marine Functional Foods. In Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 969–994. ISBN 978-3-642-53970-1. [Google Scholar]
- Rodrigues, D.; Costa-pinto, A.R.; Sousa, S.; Vasconcelos, M.W.; Pintado, M.M.; Pereira, L.; Rocha-santos, T.A.P. Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Mar. Drugs 2019, 17, 209. [Google Scholar] [CrossRef]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with α-Amylase Inhibitory Potential from Enzymatic Protein Hydrolysates of Red Seaweed (Porphyra spp.). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in Microalgae Incorporation Into Innovative Food Products With Potential Health Benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine Bioactive Compounds and Their Health Benefits: A Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Nova, P.; Pimenta-martins, A.; Silva, J.L.; Machado, A.; Gomes, A.M.; Freitas, A.C. Health benefits and bioavailability of marine resources components that contribute to health—What’ s new ? Crit. Rev. Food Sci. Nutr. 2020, 60, 3680–3692. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Nova, P.; Gomes, A.M.; Costa-Pinto, A.R. It comes from the sea: Macroalgae-derived bioactive compounds with anti-cancer potential. Crit. Rev. Biotechnol. 2023, 12, 1310. [Google Scholar] [CrossRef]
- Markets and Markets Algae Products Market| Size, Share and Market Forecast to 2023 | MarketsandMarkets. Available online: https://www.marketsandmarkets.com/Market-Reports/algae-product-market-250538721.html (accessed on 8 June 2020).
- Nova, P.; Martins, A.P.; Teixeira, C.; Abreu, H.; Silva, J.G.; Silva, A.M.; Freitas, A.C.; Gomes, A.M. Foods with microalgae and seaweeds fostering consumers health: A review on scientific and market innovations. J. Appl. Phycol. 2020, 32, 1789–1802. [Google Scholar] [CrossRef]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Cherry, P.; O’hara, C.; Magee, P.J.; Mcsorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Wang, H.M.D.; Li, X.C.; Lee, D.J.; Chang, J.S. Potential biomedical applications of marine algae. Bioresour. Technol. 2017, 244, 1407–1415. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S. Natural Products from Marine Algae: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2015; ISBN 9781493926848. [Google Scholar]
- Christaki, E.; Bonos, E.; Florou-Paneri, P. Chapter 14—Innovative Microalgae Pigments as Functional Ingredients in Nutrition. In Handbook of Marine Microalgae: Biotechnology Advances; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 233–243. ISBN 978-0-12-800776-1. [Google Scholar]
- Neves, F.d.F.; Demarco, M.; Tribuzi, G. Drying and Quality of Microalgal Powders for Human Alimentation. In Microalgae—From Physiology to Application; Vítová, M., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Fernández-Segovia, I.; Lerma-García, M.J.; Fuentes, A.; Barat, J.M. Characterization of Spanish powdered seaweeds: Composition, antioxidant capacity and technological properties. Food Res. Int. 2018, 111, 212–219. [Google Scholar] [CrossRef]
- Bjarnadóttir, M.; Aðalbjörnsson, B.V.; Nilsson, A.; Slizyte, R.; Roleda, M.Y.; Hreggviðsson, G.Ó.; Friðjónsson, Ó.H.; Jónsdóttir, R. Palmaria palmata as an alternative protein source: Enzymatic protein extraction, amino acid composition, and nitrogen-to-protein conversion factor. J. Appl. Phycol. 2018, 30, 2061–2070. [Google Scholar] [CrossRef]
- Gadberry, B.A.; Colt, J.; Maynard, D.; Boratyn, D.C.; Webb, K.; Johnson, R.B.; Saunders, G.W.; Boyer, R.H. Intensive land-based production of red and green macroalgae for human consumption in the pacific northwest: An evaluation of seasonal growth, yield, nutritional composition, and contaminant levels. Algae 2018, 33, 109–125. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [PubMed]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Van Krimpen, M.; Bikker, P.; van der Meer, I.; van der Peet-Schwering, C.; Vereijken, J. Cultivation, Processing and Nutritional Aspects for Pigs and Poultry of European Protein Sources as Alternatives for Imported Soybean Products; Wageningen UR Livestock Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Cian, R.E.; Drago, S.R.; De Medina, F.S.; Martínez-Augustin, O. Proteins and carbohydrates from red seaweeds: Evidence for beneficial effects on gut function and microbiota. Mar. Drugs 2015, 13, 5358–5383. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. In Seaweed: Ecology, Nutrient Composition and Medicinal Uses; Nova Science Publishers: Coimbra, Portugal, 2011; pp. 15–47. ISBN 978-1-61470-878-0. [Google Scholar]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef]
- Meng, Y.; Sun, J.; Yu, J.; Wang, C.; Su, J. Dietary Intakes of Calcium, Iron, Magnesium, and Potassium Elements and the Risk of Colorectal Cancer: A Meta-Analysis. Biol. Trace Elem. Res. 2019, 189, 325–335. [Google Scholar] [CrossRef]
- Tako, E. Dietary trace minerals. Nutrients 2019, 11, 2823. [Google Scholar] [CrossRef]
- Sales, C.H.; Fontanelli, M.D.M.; Vieira, D.A.S.; Marchioni, D.M.; Fisberg, R.M. Inadequate dietary intake of minerals: Prevalence and association with socio-demographic and lifestyle factors. Br. J. Nutr. 2017, 117, 267–277. [Google Scholar] [CrossRef]
- Mišurcová, L.; Škrovánková, S.; Samek, D.; Ambrožová, J.; Machů, L. Health Benefits of Algal Polysaccharides in Human Nutrition; Academic Press: Burlington, ON, Canada, 2012; Volume 66, ISBN 9780123945976. [Google Scholar]
- Orsavova, J. Seaweed Minerals as Nutraceuticals. Adv. Food Nutr. Res. 2011, 64, 371–390. [Google Scholar] [CrossRef]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from macroalgae origin: Health benefits and risks for consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef]
- Morrissey, E.; Giltinan, M.; Kehoe, L.; Nugent, A.P.; McNulty, B.A.; Flynn, A.; Walton, J. Sodium and potassium intakes and their ratio in adults (18–90 y): Findings from the Irish national adult nutrition survey. Nutrients 2020, 12, 938. [Google Scholar] [CrossRef] [PubMed]
- Iwahori, T.; Miura, K.; Ueshima, H. Time to consider use of the sodium-to-potassium ratio for practical sodium reduction and potassium increase. Nutrients 2017, 9, 700. [Google Scholar] [CrossRef] [PubMed]
- Stambler, J. The INTERSALT and implications Study: Background, methods, findings, and implications. Am. J. Clin. Nutr. 1997, 65, 626S–642S. [Google Scholar] [CrossRef] [PubMed]
- De Morais, I.L.; Lunet, N.; Albuquerque, G.; Gelormini, M.; Casal, S.; Damasceno, A.; Pinho, O.; Moreira, P.; Jewell, J.; Breda, J.; et al. The sodium and potassium content of the most commonly available street foods in Tajikistan and Kyrgyzstan in the context of the FEEDcities project. Nutrients 2018, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Malavolti, M.; Naska, A.; Fairweather-Tait, S.J.; Malagoli, C.; Vescovi, L.; Marchesi, C.; Vinceti, M.; Filippini, T. Sodium and potassium content of foods consumed in an italian population and the impact of adherence to a mediterranean diet on their intake. Nutrients 2021, 13, 2681. [Google Scholar] [CrossRef]
- Cormick, G.; Belizán, J.M. Calcium intake and health. Nutrients 2019, 11, 1606. [Google Scholar] [CrossRef]
- Theobald, H. Dietary calcium and health. Nutr Bull. 2005, 30, 237–277. [Google Scholar] [CrossRef]
- Shkembi, B.; Huppertz, T. Calcium absorption from food products: Food matrix effects. Nutrients 2022, 14, 180. [Google Scholar] [CrossRef]
- Volpe, S.L. Magnesium in Disease Prevention and Overall Health. Adv. Nutr. 2013, 4, 378S–383S. [Google Scholar] [CrossRef]
- Razzaque, M.S. Magnesium: Are we consuming enough? Nutrients 2018, 10, 1863. [Google Scholar] [CrossRef]
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The role of magnesium in neurological disorders. Nutrients 2018, 10, 730. [Google Scholar] [CrossRef]
- Chen, F.; Du, M.; Blumberg, J.B.; Chui, K.K.H.; Ruan, M.; Rogers, G.; Shan, Z.; Zeng, L.; Zhang, F.F. Association Among Dietary Supplement Use, Nutrient Intake, and Mortality Among U.S. Adults. Ann. Intern. Med. 2019, 170, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Costello, R.B.; Rosanoff, A.; Dai, Q.; Saldanha, L.G.; Potischman, N.A. Perspective: Characterization of Dietary Supplements Containing Calcium and Magnesium and Their Respective Ratio-Is a Rising Ratio a Cause for Concern? Adv. Nutr. 2021, 12, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Sliwinska, A.; Luty, J.; Aleksandrowicz-Wrona, E.; Małgorzewicz, S. Iron status and dietary iron intake in vegetarians. Adv. Clin. Exp. Med. 2018, 27, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M.; Erikson, K. Manganese. Adv. Nutr. 2017, 8, 520–521. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Roy, C.I.L. Effect of diet on the gut microbiota: Rethinking intervention duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; De Morais, A.M.M.B.; De Morais, R.M.S.C. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Patil, N.P.; Le, V.; Sligar, A.D.; Mei, L.; Chavarria, D.; Yang, E.Y.; Baker, A.B. Algal Polysaccharides as Therapeutic Agents for Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 153. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal. Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical constituents and biological activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, P.; Crowe, W.; Bahar, B.; Harnedy, P.A.; Brown, E.S.; Taylor, S.S.; Smyth, T.J.; Soler-Vila, A.; Magee, P.J.; Gill, C.I.R.; et al. The effect of consuming Palmaria palmata-enriched bread on inflammatory markers, antioxidant status, lipid profile and thyroid function in a randomised placebo-controlled intervention trial in healthy adults. Eur. J. Nutr. 2016, 55, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Saeedi, M.; Nabavi, S.M.; Mubarak, M.S.; Bishayee, A. Glycosides from Medicinal Plants as Potential Anticancer Agents: Emerging Trends Towards Future Drugs. Curr. Med. Chem. 2019, 26, 2389–2406. [Google Scholar] [CrossRef] [PubMed]
- Viana, A.G.; Noseda, M.D.; Gonçalves, A.G.; Duarte, M.E.R.; Yokoya, N.; Matulewicz, M.C.; Cerezo, A.S. β-D-(1→4), β-D-(1→3) “mixed linkage” xylans from red seaweeds of the order Nemaliales and Palmariales. Carbohydr. Res. 2011, 346, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Alba, K.; Kontogiorgos, V. Seaweed Polysaccharides (Agar, Alginate Carrageenan); Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128140451. [Google Scholar]
- Yun, E.J.; Yu, S.; Park, N.J.; Cho, Y.; Han, N.R.; Jin, Y.S.; Kim, K.H. Metabolic and enzymatic elucidation of cooperative degradation of red seaweed agarose by two human gut bacteria. Sci. Rep. 2021, 11, 13955. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Yu, S.; Kim, Y.A.; Liu, J.J.; Kang, N.J.; Jin, Y.S.; Kim, K.H. In vitro prebiotic and anti-colon cancer activities of agar-derived sugars from red seaweeds. Mar. Drugs 2021, 19, 213. [Google Scholar] [CrossRef]
- Xie, X.T.; Zhang, X.; Liu, Y.; Chen, X.Q.; Cheong, K.L. Quantification of 3,6-anhydro-galactose in red seaweed polysaccharides and their potential skin-whitening activity. 3 Biotech. 2020, 10, 189. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R.; Caramujo, M.J. The various roles of fatty acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Sakamoto, A.; Saotome, M.; Iguchi, K.; Maekawa, Y. Marine-derived omega-3 polyunsaturated fatty acids and heart failure: Current understanding for basic to clinical relevance. Int. J. Mol. Sci. 2019, 20, 4025. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Ranard, K.M.; Jeon, S.; Mohn, E.S.; Griffiths, J.C.; Johnson, E.J.; Erdman, J.W. Dietary guidance for lutein: Consideration for intake recommendations is scientifically supported. Eur. J. Nutr. 2017, 56, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bi, X.; Wang, S.; Zhang, Z.; Li, F.; Zhao, A.Z. Therapeutic potential of ω-3 polyunsaturated fatty acids in human autoimmune diseases. Front. Immunol. 2019, 10, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.B.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds ulva armoricana, and solieria chordalis from brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef] [PubMed]
- De Alencar, D.B.; Diniz, J.C.; Rocha, S.A.S.; Pires-Cavalcante, K.M.S.; De Lima, R.L.; De Sousa, K.C.; Freitas, J.O.; Bezerra, R.M.; Baracho, B.M.; Sampaio, A.H.; et al. Fatty acid composition from the marine red algae pterocladiella capillacea (S. G. gmelin) santelices & hommersand 1997 and osmundaria obtusiloba (C. agardh) R. E. norris 1991 and its antioxidant activity. An. Acad. Bras. Cienc. 2018, 90, 449–459. [Google Scholar] [CrossRef]
- El Maghraby, D.M.; Fakhry, E.M. Lipid content and fatty acid composition of Mediterranean macro-algae as dynamic factors for biodiesel production. Oceanologia 2015, 57, 86–92. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Queirós, R.; Rocha-Santos, T.A.P.; Saraiva, J.A.; Gomes, A.M.P.; Duarte, A.C. Bioactive Polysaccharides Extracts from Sargassum muticum by High Hydrostatic Pressure. J. Food Process. Preserv. 2017, 41, e12977. [Google Scholar] [CrossRef]
- Aminina, N.M.; Karaulova, E.P.; Vishnevskaya, T.I.; Yakush, E.V.; Kim, Y.K.; Nam, K.H.; Son, K.T. Characteristics of Polyphenolic Content in Brown Algae of the Pacific Coast of Russia. Molecules 2020, 25, 3909. [Google Scholar] [CrossRef]
- Barbosa, M.; Valentão, P.; Ferreres, F.; Gil-Izquierdo, Á.; Andrade, P.B. In vitro multifunctionality of phlorotannin extracts from edible Fucus species on targets underpinning neurodegeneration. Food Chem. 2020, 333, 127456. [Google Scholar] [CrossRef]
- Ford, L.; Theodoridou, K.; Sheldrake, G.N.; Walsh, P.J. A critical review of analytical methods used for the chemical characterisation and quantification of phlorotannin compounds in brown seaweeds. Phytochem. Anal. 2019, 30, 587–599. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. An emerging trend in functional foods for the prevention of cardiovascular disease and diabetes: Marine algal polyphenols. Crit. Rev. Food Sci. Nutr. 2018, 58, 1342–1358. [Google Scholar] [CrossRef] [PubMed]
- Mahato, N.; Sinha, M.; Sharma, K.; Koteswararao, R.; Cho, M.H. Modern Extraction and Purification Techniques for Obtaining High Purity Food-Grade Bioactive Compounds and Value-Added Co-Products from Citrus Wastes. Foods 2019, 8, 523. [Google Scholar] [CrossRef]
- Pruteanu, L.L.; Bailey, D.S.; Cristian, A. The Biochemistry and Effectiveness of Antioxidants in Food, Fruits, and Marine Algae. Antioxidants 2023, 12, 860. [Google Scholar] [CrossRef] [PubMed]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Tenorio-Rodriguez, P.A.; Murillo-Álvarez, J.I.; Campa-Cordova, Á.I.; Angulo, C. Antioxidant screening and phenolic content of ethanol extracts of selected Baja California Peninsula macroalgae. J. Food Sci. Technol. 2017, 54, 422–429. [Google Scholar] [CrossRef]
- Castejón, N.; Parailloux, M.; Izdebska, A.; Lobinski, R.; Fernandes, S.C.M. Valorization of the red algae gelidium sesquipedale by extracting a broad spectrum of minor compounds using green approaches. Mar. Drugs 2021, 19, 574. [Google Scholar] [CrossRef] [PubMed]
- Surget, G.; Roberto, V.P.; Le Lann, K.; Mira, S.; Guérard, F.; Laizé, V.; Poupart, N.; Cancela, M.L.; Stiger-Pouvreau, V. Marine green macroalgae: A source of natural compounds with mineralogenic and antioxidant activities. J. Appl. Phycol. 2017, 29, 575–584. [Google Scholar] [CrossRef]
- Fradinho, P.; Flórez-Fernández, N.; Sousa, I.; Raymundo, A.; Domínguez, H.; Torres, M.D. Environmentally friendly processing of Laminaria ochroleuca for soft food applications with bioactive properties. J. Appl. Phycol. 2020, 32, 1455–1465. [Google Scholar] [CrossRef]
- Pereira, J.M.; Lopes-Rodrigues, V.; Xavier, C.P.R.; Lima, M.J.; Lima, R.T.; Ferreira, I.C.F.R.; Vasconcelos, M.H. An aqueous extract of Tuberaria lignosa inhibits cell growth, alters the cell cycle profile, and induces apoptosis of NCI-H460 tumor cells. Molecules 2016, 21, 595. [Google Scholar] [CrossRef]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef]
- Sajadimajd, S.; Momtaz, S.; Haratipour, P.; El-Senduny, F.F.; Panah, A.I.; Navabi, J.; Soheilikhah, Z.; Farzaei, M.H.; Rahimi, R. Molecular Mechanisms Underlying Cancer Preventive and Therapeutic Potential of Algal Polysaccharides. Curr. Pharm. Des. 2019, 25, 1210–1235. [Google Scholar] [CrossRef] [PubMed]
- Geisen, U.; Zenthoefer, M.; Peipp, M.; Kerber, J.; Plenge, J.; Managò, A.; Fuhrmann, M.; Geyer, R.; Hennig, S.; Adam, D.; et al. Molecular mechanisms by which a Fucus vesiculosus extract mediates cell cycle inhibition and cell death in pancreatic cancer cells. Mar. Drugs 2015, 13, 4470–4491. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.; Guiry, G.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 9 June 2020).
- Almeida, C.C.; Alvares, T.S.; Costa, M.P.; Conte-Junior, C.A. Protein and Amino Acid Profiles of Different Whey Protein Supplements. J. Diet. Suppl. 2016, 13, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Al Juhaimi, F.; Özcan, M.M.; Ghafoor, K.; Babiker, E.E.; Hussain, S. Comparison of cold-pressing and soxhlet extraction systems for bioactive compounds, antioxidant properties, polyphenols, fatty acids and tocopherols in eight nut oils. J. Food Sci. Technol. 2018, 55, 3163–3173. [Google Scholar] [CrossRef] [PubMed]
- Goering, H.K.; Van Soest, P. Forage fiber analyses. US Dep. Agric.379 1975, 387–598. [Google Scholar]
- Voss, G.B.; Rodríguez-Alcalá, L.M.; Valente, L.M.P.; Pintado, M.M. Impact of different thermal treatments and storage conditions on the stability of soybean byproduct (okara). J. Food Meas. Charact. 2018, 12, 1981–1996. [Google Scholar] [CrossRef]
- Fontes, A.L.; Pimentel, L.; Rodríguez-Alcalá, L.M.; Gomes, A. Effect of Pufa Substrates on Fatty Acid Profile of Bifidobacterium breve Ncimb 702258 and CLA/CLNA Production in Commercial Semi-Skimmed Milk. Sci. Rep. 2018, 8, 15591. [Google Scholar] [CrossRef]
- Coimbra, M.A.; Waldron, K.W.; Selvendran, R.R. Isolation and characterisation of cell wall polymers from olive pulp (Olea europaea L.). Carbohydr. Res. 1994, 252, 245–262. [Google Scholar] [CrossRef]
- Stevenson, T.T.; Furneaux, R.H. Chemical methods for the analysis of sulphated galactans from red algae. Carbohydr. Res. 1991, 210, 277–298. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Marinho, G.S.; Sørensen, A.D.M.; Safafar, H.; Pedersen, A.H.; Holdt, S.L. Antioxidant content and activity of the seaweed Saccharina latissima: A seasonal perspective. J. Appl. Phycol. 2019, 31, 1343–1354. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.P.; Duarte, A.C.; Freitas, A.C. Impact of enzyme- and ultrasound-assisted extraction methods on biological properties of red, brown, and green seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Suresh, V.; Senthilkumar, N.; Thangam, R.; Rajkumar, M.; Anbazhagan, C.; Rengasamy, R.; Gunasekaran, P.; Kannan, S.; Palani, P. Separation, purification and preliminary characterization of sulfated polysaccharides from Sargassum plagiophyllum and its in vitro anticancer and antioxidant activity. Process. Biochem. 2013, 48, 364–373. [Google Scholar] [CrossRef]
- Gião, M.S.; González-Sanjosé, M.L.; Rivero-Pérez, M.D.; Pereira, C.I.; Pintado, M.E.; Malcata, F.X. Infusions of Portuguese medicinal plants: Dependence of final antioxidant capacity and phenol content on extraction features. J. Sci. Food Agric. 2007, 87, 2638–2647. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameter | F. vesiculosus | G. gracilis | P. palmata | P. dioica | U. rigida |
|---|---|---|---|---|---|
| % moisture (g/100 g *) | 12.4 ± 0.2 a | 4.1 ± 0.2 b | 3.2 ± 0.1 c | 10.4 ± 0.2 d | 16.2 ± 0.1 e |
| % protein (g/100 g *) | 12.4 ± 0.01 a | 28.8 ± 0.01 b | 41.8 ± 0.1 c | 35.7 ± 0.1 d | 19.5 ± 0.1 e |
| % sugars (g/100 g *) | 30.2 ± 1.3 | 26.8 ± 2.7 | 9.3 ± 0.1 | 29.8 ± 0.6 | 27.9 ± 0.4 |
| % total fat (g/100 g *) | 3.4 ± 0.2 a | 0.68 ± 0.01 b | 0.32 ± 0.02 b,c | 1.59 ± 0.01 d | 0.08 ± 0.004 c |
| % ash (g/100 g *) | 29.8 ± 0.1 a | 29.0 ± 0.2 a | 27.1 ± 0.6 b | 23.4 ± 0.03 c | 26.6 ± 0.4 b |
| Elements | mg/ day | F. vesiculosus mg/10 g Portion | % R D I | G. gracilis mg/10 g Portion | % R D I | P. palmata mg/10 g Portion | % R D I | P. dioica mg/10 g Portion | % R D I | U. rigida mg/10 g Portion | % R D I |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Calcium (Ca) | 800 | 77.9 ± 1.6 a | 9.7 | 62.4 ± 0.3 b | 7.8 | 159.1 ± 1.4 c | 19.9 | 22.5 ± 2.3 d | 2.8 | 35.4 ± 2.5 e | 4.4 |
| Potassium (K) | 2000 | 369.6 ± 5.5 a | 18.5 | 892.0 ± 7.5 b | 44.6 | 345.2 ± 7.2 a | 17.3 | 286.3 ± 4.3c | 14.3 | 136.4 ± 7.4 d | 6.8 |
| Sodium (Na) | -- | 476.4 ± 7.6 a | -- | 261.3 ± 1.7 b | -- | 175.9 ± 4.2 c | -- | 425.6 ± 6.2 d | -- | 105.7 ± 3.3 e | -- |
| Magnesium (Mg) | 375 | 81.3 ± 2.4 a | 21.7 | 40.2 ± 1.4 b | 10.7 | 22.20 ± 0.04 b | 5.9 | 61.7 ± 2.3 a,b | 16.5 | 186.0± c | 49.6 |
| Phosphorus (P) | 700 | 11.7 ± 0.2 a | 1.7 | 41.5 ± 1.7 b | 5.9 | 11.0 ± 0.18 a | 1.6 | 39.3 ± 1.4 c | 5.6 | 12.7 ± 0.2 d | 1.8 |
| Iron (Fe) | 14 | 10.3 ± 0.3 a | 73.6 | 16.5 ± 0.8 b | 117.9 | 4.6 ± 0.2 c | 32.9 | 6.2 ± 0.2 c | 44.4 | 13.1 ± 0.4 b | 93.6 |
| Zinc (Zn) | 10 | 0.56 ± 0.5 a | 5.6 | 0.41 ± 0.03 b | 4.1 | 0.18 ± 0.01 c | 1.8 | 0.73 ± 0.02 d | 7.3 | 0.14 ± 0.02 c | 1.4 |
| Copper (Cu) | 1 | 0.080 ± 0.001a | 8.0 | 0.26 ± 0.01 b | 26.0 | 0.050 ± 0.001a | 5.0 | 0.70 ± 0.03 c | 70.0 | 0.08 ± 0.01 a | 8.0 |
| Manganese (Mn) | 2 | 2.94 ± 0.03 a | 145.0 | 2.40 ± 0.03 b | 120.0 | 2.10 ± 0.02 c | 105.0 | 1.90 ± 0.01 c | 95.6 | 0.60 ± 0.02 d | 30.0 |
| Na/K ratio (mmol) | -- | 2.19 | -- | 0.50 | -- | 0.86 | -- | 2.52 | -- | 1.31 | -- |
| Ca/Mg ratio (mg) | -- | 0.96 | -- | 1.56 | -- | 7.16 | -- | 0.36 | -- | 0.19 | -- |
| Parameter | F. vesiculosus | G. gracilis | P. palmata | P. dioica | U. rigida | |
|---|---|---|---|---|---|---|
| % fiber (g/100 g *) | Total | 37.0 ± 0.4 a | 15.0 ± 0.03 b | 26.5 ± 0.7 c | 21.7 ± 0.8 d | 27.3 ± 1.6 c |
| Soluble | 15.6 ± 1.1 a | 7.1 ± 0.1 b | 9.8 ± 0.4 c | 20.4 ± 0.9 d | 8.9 ± 1.4 b,c | |
| Insoluble | 21.4 ± 1.6 a | 7.9 ± 0.2 b | 16.7 ± 0.8 c | 1.3 ± 0.05 d | 18.4 ± 0.2 c | |
| mol % | F. vesiculosus | U. rigida | P. palmata | P. dioica | G. gracilis |
|---|---|---|---|---|---|
| Rha | 0.4 | 23.1 | 0.3 | 0.1 | 0.5 |
| Fuc | 14.3 | -- | 2.3 | 0.1 | 0.7 |
| Rib | 2.1 | 0.2 | 1.1 | 0.7 | 0.6 |
| Ara | 0.4 | -- | 0.2 | 0.2 | -- |
| 3,6-AnGal | -- | -- | 0.2 | 14.4 | 24.6 |
| Xyl | 2.0 | 6.5 | 56.5 | 6.5 | 2.0 |
| 6-O-Me-Gal | -- | -- | -- | 10.9 | 7.3 |
| 4-O-Me-Gal | -- | -- | -- | -- | 4.5 |
| Man | 17.2 | 1.3 | 3.3 | 7.4 | 0.4 |
| Gal | 3.0 | 3.3 | 21.9 | 53.8 | 34.1 |
| Glc | 9.4 | 37.3 | 6.0 | 2.6 | 19.3 |
| UA | 51.2 | 28.4 | 8.1 | 3.3 | 5.9 |
| Seaweed | F. vesiculosus | G. gracilis | P. palmata | P. dioica | U. rigida | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acids | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| C14i | 0.29 a | 0.01 | 0.18 b,c | 0.02 | 0.12 c | 0.01 | 0.23 a,b | <0.01 | 0.24 a,b | 0.02 | |
| C14 | 3.76 a | 0.03 | 0.71 b | 0.01 | 0.54 c | 0.01 | 0.80 b | 0.01 | 0.08 d | 0.00 | |
| C15 | 0.17 | <0.01 | -- | -- | -- | -- | -- | -- | -- | -- | |
| C15:1 | 0.20 a | 0.01 | 0.13 a,b | 0.01 | 0.09 b | 0.01 | 0.17 a,b | <0.01 | 0.18 a,b | 0.02 | |
| C16 | 6.39 a | 0.06 | 3.66 b | 0.02 | 1.99 c | 0.06 | 3.38 b,c | 0.03 | 2.94 c | 0.21 | |
| C16:1 | -- | -- | -- | -- | -- | -- | 0.16 | <0.01 | -- | -- | |
| C16:1 c7 | 0.12 a | <0.01 | 0.08 b | 0.01 | 0.16 c | <0.01 | -- | -- | -- | -- | |
| C16:1 c9 | 0.45 a | <0.01 | 0.58 b | 0.01 | -- | -- | -- | -- | -- | -- | |
| C17 | 0.09 a | <0.01 | -- | -- | -- | -- | 0.65 b | 0.01 | 0.27 c | 0.02 | |
| C17:1 c10 | 0.09 a | <0.01 | -- | -- | -- | -- | -- | -- | 0.24 b | 0.02 | |
| C18 | 0.71 a | 0.01 | 0.16 b | 0.01 | 0.15 b | 0.01 | 0.11 c | <0.01 | 0.14 b | 0.04 | |
| C18:1 t4 | 0.21 a | <0.01 | -- | -- | -- | -- | -- | -- | 0.16 a | 0.01 | |
| C18:1 c9 | 8.83 a | 0.05 | 0.66 b | <0.01 | 0.71 b | 0.04 | 0.14 c | <0.01 | -- | -- | |
| C18:1 c11 | 0.17 a | <0.01 | 0.22 b | <0.01 | 0.17 a | 0.01 | 0.21 b | <0.01 | 1.22 c | 0.11 | |
| C18:2 c9c12 | 1.38 a | <0.01 | 0.10 b | <0.01 | 0.07 b | 0.01 | -- | -- | 0.12 b | 0.02 | |
| C18:3 c9c12c15 | 0.91 a | 0.01 | -- | -- | -- | -- | -- | -- | 0.18 b | 0.03 | |
| C20 | 0.17 a | <0.01 | -- | -- | -- | -- | -- | -- | 0.13 b | 0.02 | |
| C18:2 c9t11 | 0.07 a | <0.01 | -- | -- | -- | -- | 0.18 b | <0.01 | -- | -- | |
| C21 | 0.08 | <0.01 | -- | -- | -- | -- | -- | -- | -- | -- | |
| C20:4 n6 | 1.63 a | 0.01 | 0.50 b | 0.05 | -- | -- | 0.07 c | <0.01 | -- | -- | |
| C22 | 0.14 a | <0.01 | -- | -- | -- | -- | -- | -- | -- | -- | |
| C20:5 n3 | 0.53 a | <0.01 | -- | -- | 0.10 b | 0.01 | -- | -- | -- | -- | |
| Total µg/mg (dry seaweed) | 26.39 | 6.99 | 4.10 | 6.11 | 5.91 | ||||||
| SFA | 11.79 | 4.72 | 2.81 | 5.17 | 3.80 | ||||||
| MUFA | 10.08 | 1.66 | 1.12 | 0.68 | 1.80 | ||||||
| PUFAS | 4.52 | 0.61 | 0.17 | 0.26 | 0.30 | ||||||
| MUFA/SFA ratio | 0.85 | 0.35 | 0.40 | 0.13 | 0.47 | ||||||
| SFA/MUFA ratio | 1.17 | 2.84 | 2.51 | 7.60 | 2.11 | ||||||
| SFA/(MUFA + PUFAS) ratio | 0.81 | 2.08 | 2.18 | 5.50 | 1.81 | ||||||
| Radical Scavenging Activity | Total Antioxidant Capacity | Total Phenolic Content | |
|---|---|---|---|
| SEAWEED | DPPH, mg Trolox equiv/g Dry Algae Extract | ABTS, mg Ascorbic Acid equiv/g Dry Algae Extract | Folin–Ciocalteu, mg Gallic acid equiv/g % Dry Algae Extract |
| F. vesiculosus | 0.033 ± 0.002 a | 8.20 ± 0.07 a | 10.89 ± 0.61 a |
| U. rigida | 0.129 ± 0.002 b | 9.89 ± 0.70 b | 3.27 ± 0.25 b |
| P. palmata | 0.127 ± 0.010 b | 8.35 ± 0.20 a | 5.52 ± 0.52 c |
| P. dioica | 0.126 ± 0.003 b | 9.67 ± 0.06 b | 3.87 ± 0.41 b,c |
| G. gracilis | 0.115 ± 0.005 c | 8.90 ± 0.11 c | 4.41 ± 0.21 b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nova, P.; Pimenta-Martins, A.; Maricato, É.; Nunes, C.; Abreu, H.; Coimbra, M.A.; Freitas, A.C.; Gomes, A.M. Chemical Composition and Antioxidant Potential of Five Algae Cultivated in Fully Controlled Closed Systems. Molecules 2023, 28, 4588. https://doi.org/10.3390/molecules28124588
Nova P, Pimenta-Martins A, Maricato É, Nunes C, Abreu H, Coimbra MA, Freitas AC, Gomes AM. Chemical Composition and Antioxidant Potential of Five Algae Cultivated in Fully Controlled Closed Systems. Molecules. 2023; 28(12):4588. https://doi.org/10.3390/molecules28124588
Chicago/Turabian StyleNova, Paulo, Ana Pimenta-Martins, Élia Maricato, Cláudia Nunes, Helena Abreu, Manuel A. Coimbra, Ana Cristina Freitas, and Ana Maria Gomes. 2023. "Chemical Composition and Antioxidant Potential of Five Algae Cultivated in Fully Controlled Closed Systems" Molecules 28, no. 12: 4588. https://doi.org/10.3390/molecules28124588