

Functional Axis of PDE5/cGMP Mediates Timosaponin-AIII-Elicited Growth Suppression of Glioblastoma U87MG Cells
 , , , , , , and
, , , , , , and 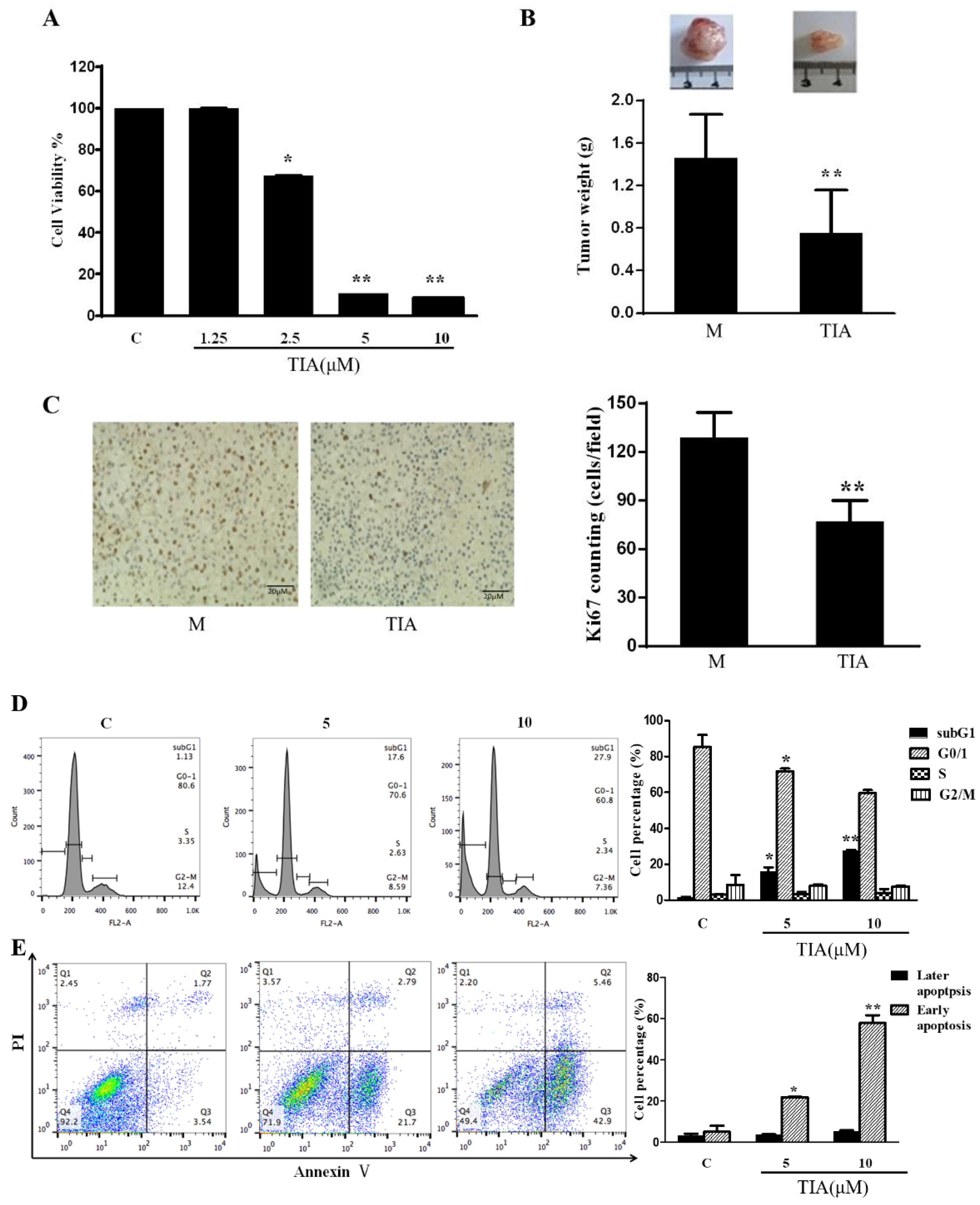{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. TIA Inhibited In Vitro Cell Growth and In Vivo Tumor Development of U87MG Cells
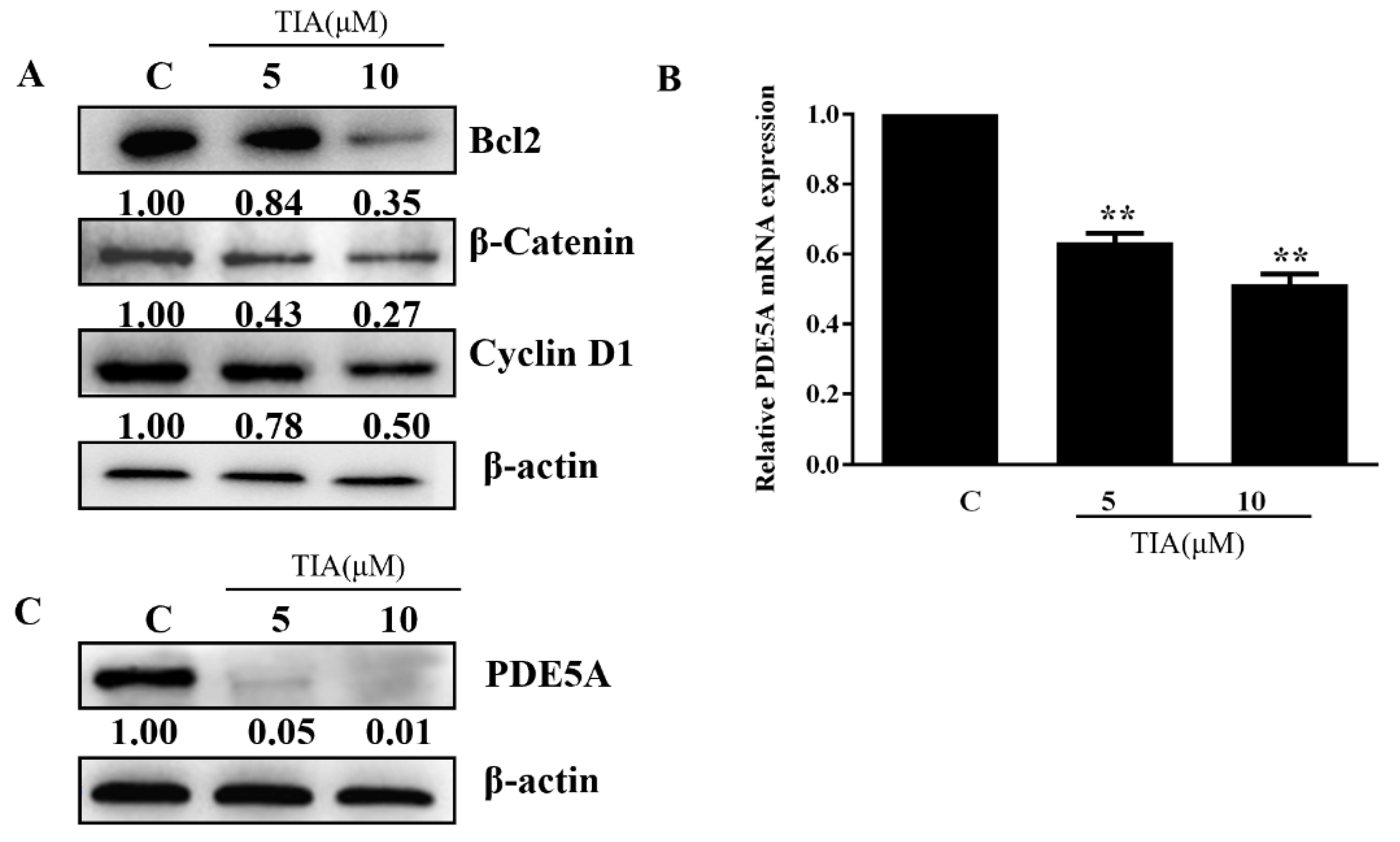
2.2. TIA Downregulated the Abundance of β-Catenin, Cyclin D1, Bcl-2, and PDE5 in U87MG Cells
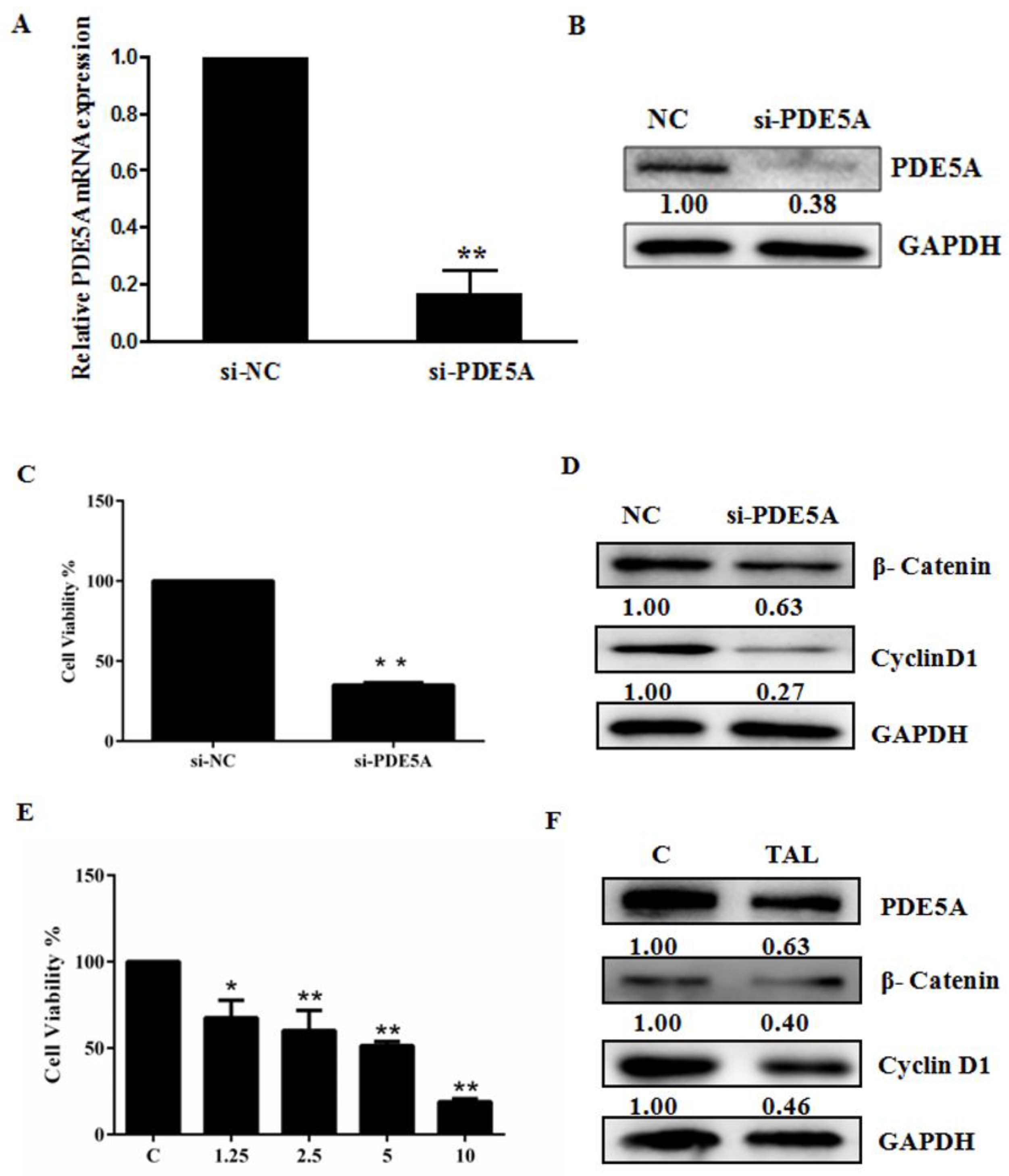
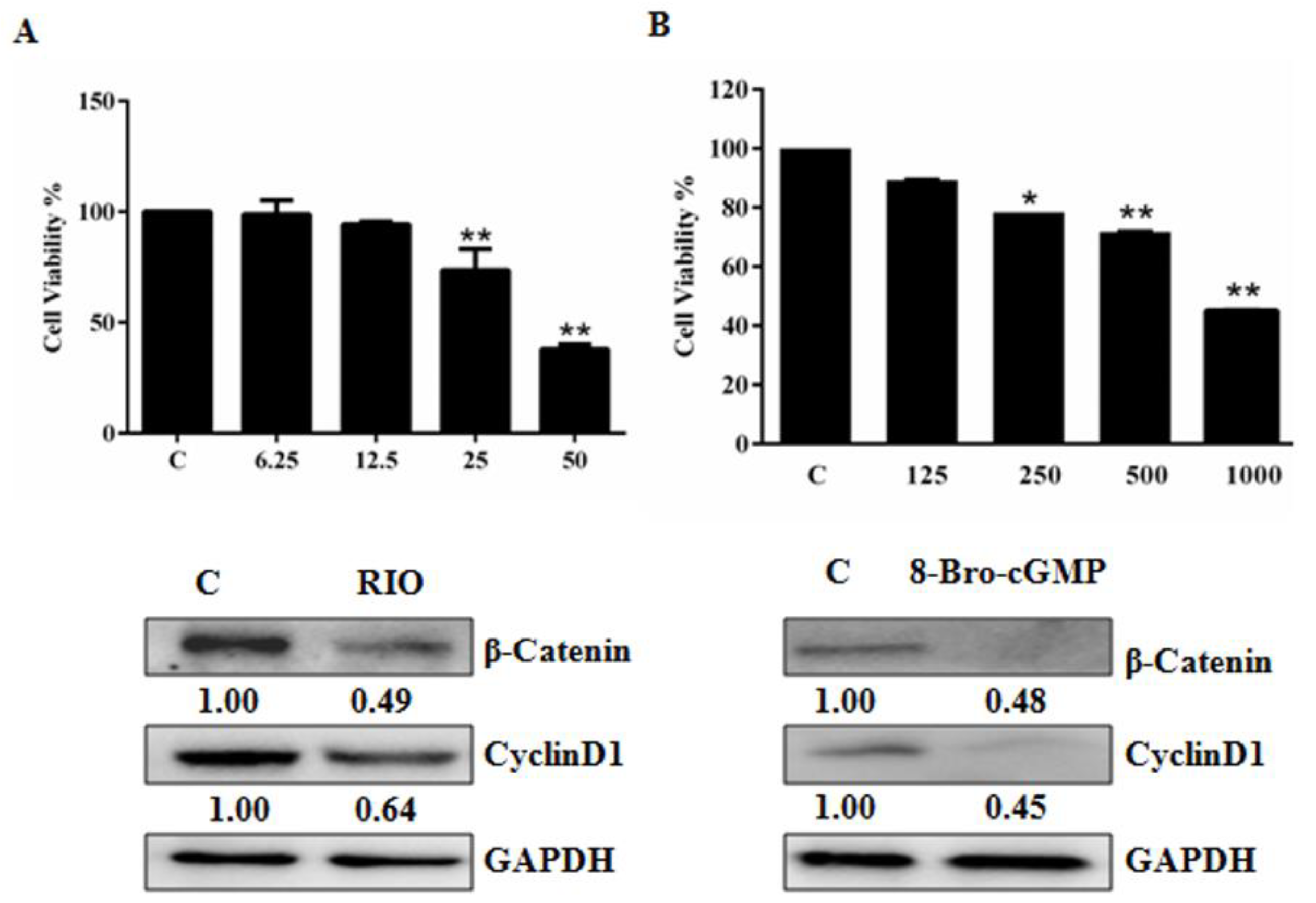
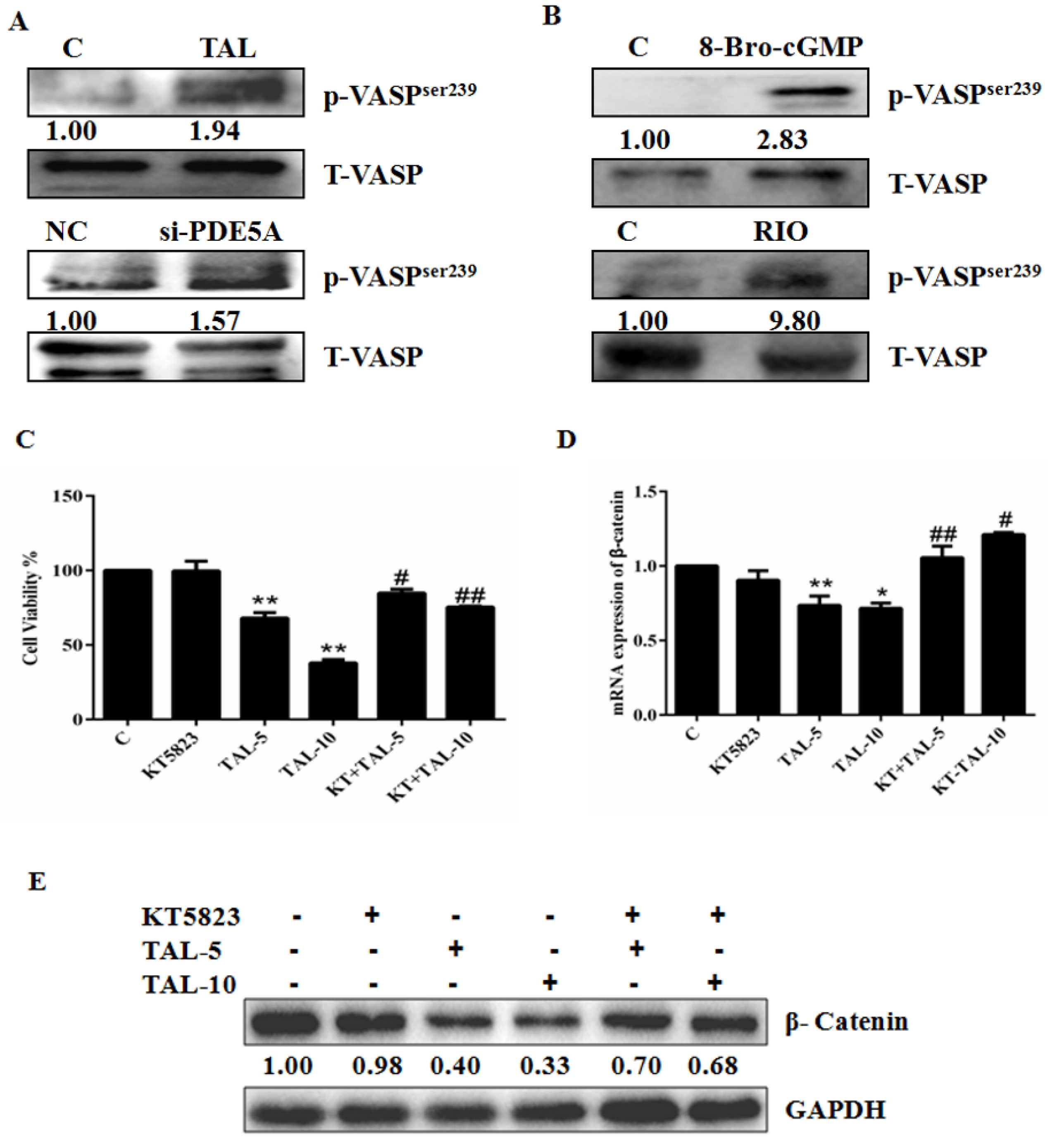
2.3. Activation of the cGMP Pathway Inhibits U87MG Cell Growth by Interfering with β-Catenin
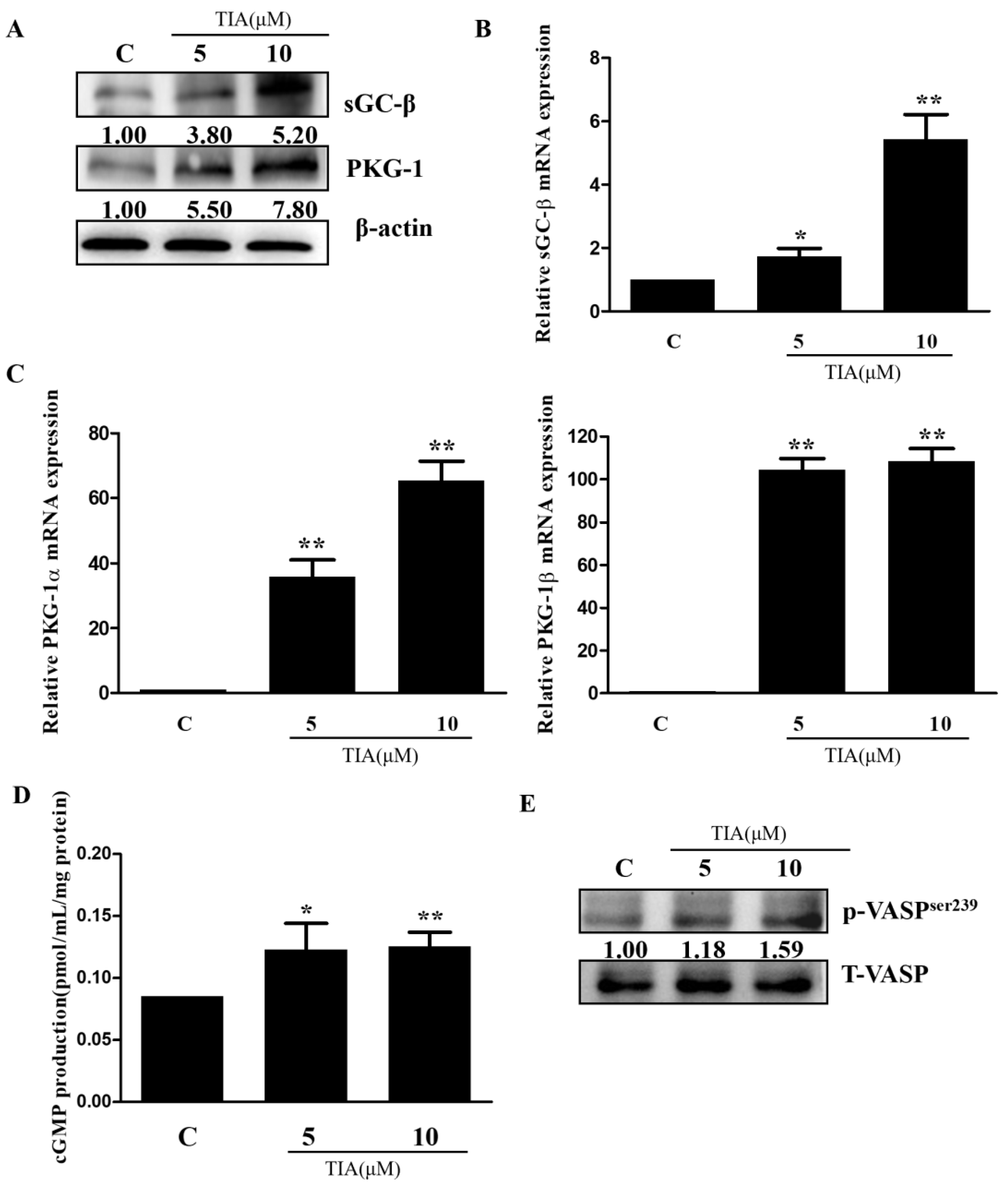
2.4. TIA Regulates the PDE5/cGMP Axis in U87MG Cells
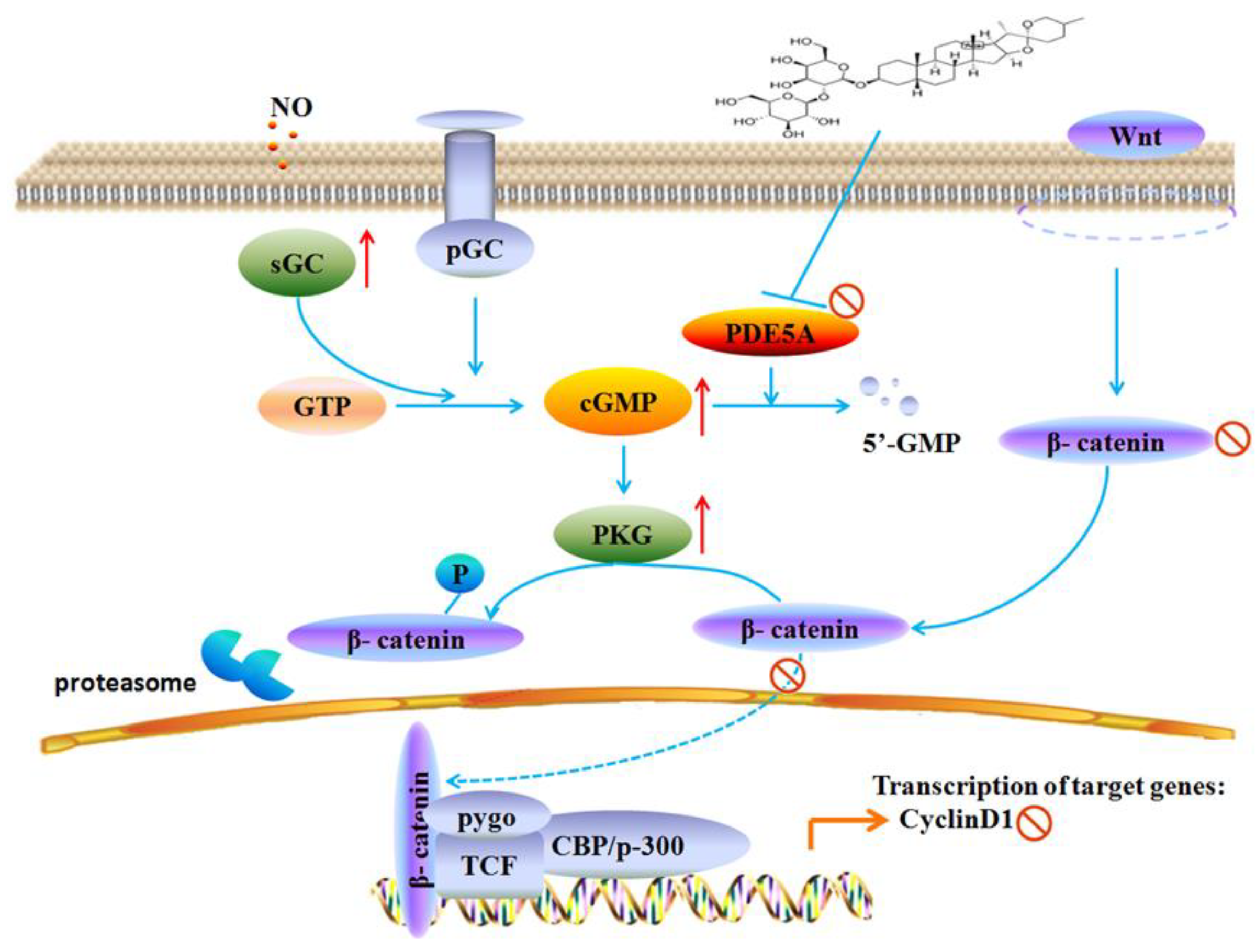
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Transfection
4.3. Cell Proliferation Assay
4.4. Cell Cycle and Apoptosis Analyses
4.5. qRT-PCR
4.6. Western Blotting Analysis
4.7. cGMP Analysis
4.8. Animals and Xenograft Studies
4.9. Immunohistochemistry
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- De Silva, M.I.; Stringer, B.W.; Bardy, C. Neuronal and tumourigenic boundaries of glioblastoma plasticity. Trends Cancer 2022, 9, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Messali, A.; Villacorta, R.; Hay, J.W. A review of the economic burden of glioblastoma and the cost effectiveness of pharmacologic treatments. Pharmacoeconomics 2014, 32, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Bian, K.; Murad, F. sGC-cGMP signaling: Target for anticancer therapy. Adv. Exp. Med. Biol. 2014, 814, 5–13. [Google Scholar]
- Windham, P.F.; Tinsley, H.N. cGMP signaling as a target for the prevention and treatment of breast cancer. Semin. Cancer Biol. 2015, 31, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zheng, J.; Yao, X.; Weng, G.; Wu, L. Essential role of the cGMP/PKG signaling pathway in regulating the proliferation and survival of human renal carcinoma cells. Int. J. Mol. Med. 2014, 34, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, T.R.; Mierzwa, M.L.; Wells, S.I.; Fox, S.R.; Ben-Jonathan, N. The cyclic GMP/protein kinase G pathway as a therapeutic target in head and neck squamous cell carcinoma. Cancer Lett. 2016, 370, 279–285. [Google Scholar] [CrossRef]
- Lou, W.; Ding, B.; Xu, L.; Fan, W. Construction of Potential Glioblastoma Multiforme-Related miRNA-mRNA Regulatory Network. Front. Mol. Neurosci. 2019, 12, 66. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Kumar, M.S.; Lu, J.; Mercer, K.L.; Golub, T.R.; Jacks, T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat. Genet. 2007, 39, 673–677. [Google Scholar] [CrossRef]
- Zhu, H.; Li, J.T.; Zheng, F.; Martin, E.; Kots, A.Y.; Krumenacker, J.S.; Choi, B.K.; McCutcheon, I.E.; Weisbrodt, N.; Bögler, O.; et al. Restoring soluble guanylyl cyclase expression and function blocks the aggressive course of glioma. Mol. Pharmacol. 2011, 80, 1076–1084. [Google Scholar] [CrossRef]
- Catalano, S.; Campana, A.; Giordano, C.; Győrffy, B.; Tarallo, R.; Rinaldi, A.; Bruno, G.; Ferraro, A.; Romeo, F.; Lanzino, M.; et al. Expression and Function of Phosphodiesterase Type 5 in Human Breast Cancer Cell Lines and Tissues: Implications for Targeted Therapy. Clin. Cancer Res. 2016, 9, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Karami-Tehrani, F.; Moeinifard, M.; Aghaei, M.; Atri, M. Evaluation of PDE5 and PDE9 expression in benign and malignant breast tumors. Arch. Med. Res. 2012, 43, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, T.; Onda, T.; Honda, H.; Hayashi, K.; Shibahara, T.; Nomura, T.; Takano, M. Over-expression of PDE5 in Oral Squamous Cell Carcinoma-Effect of Sildenafil Citrate. Anticancer. Res. 2021, 41, 2297–2306. [Google Scholar] [CrossRef]
- Klutzny, S.; Anurin, A.; Nicke, B.; Regan, J.L.; Lange, M.; Schulze, L.; Parczyk, K.; Steigemann, P. PDE5 inhibition eliminates cancer stem cells via induction of PKA signaling. Cell Death Dis. 2018, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Piazza, G.A.; Ward, A.; Chen, X.; Maxuitenko, Y.; Coley, A.; Aboelella, N.S.; Buchsbaum, D.J.; Boyd, M.R.; Keeton, A.B.; Zhou, G. PDE5 and PDE10 inhibition activates cGMP/PKG signaling to block Wnt/β-catenin transcription, cancer cell growth, and tumor immunity. Drug Discov. Today 2020, 25, 1521–1527. [Google Scholar] [CrossRef]
- Das, A.; Durrant, D.; Salloum, F.N.; Xi, L.; Kukreja, R.C. PDE5 inhibitors as therapeutics for heart disease, diabetes and cancer. Pharmacol. Ther. 2015, 147, 12–21. [Google Scholar] [CrossRef]
- Peak, T.C.; Richman, A.; Gur, S.; Yafi, F.A.; Hellstrom, W.J. The Role of PDE5 Inhibitors and the NO/cGMP Pathway in Cancer. Sex. Med. Rev. 2016, 4, 74–84. [Google Scholar] [CrossRef]
- Mei, X.L.; Yang, Y.; Zhang, Y.J.; Li, Y.; Zhao, J.M.; Qiu, J.G.; Zhang, W.J.; Jiang, Q.W.; Xue, Y.Q.; Zheng, D.W.; et al. Sildenafil inhibits the growth of human colorectal cancer in vitro and in vivo. Am. J. Cancer Res. 2015, 5, 3311–3324. [Google Scholar] [PubMed]
- Huang, W.; Sundquist, J.; Sundquist, K.; Ji, J. Use of Phosphodiesterase 5 Inhibitors Is Associated With Lower Risk of Colorectal Cancer in Men With Benign Colorectal Neoplasms. Gastroenterology 2019, 157, 672–681.e4. [Google Scholar] [CrossRef]
- Hu, J.; Ljubimova, J.Y.; Inoue, S.; Konda, B.; Patil, R.; Ding, H.; Espinoza, A.; Wawrowsky, K.A.; Patil, C.; Ljubimov, A.V.; et al. Phosphodiesterase type 5 inhibitors increase Herceptin transport and treatment efficacy in mouse metastatic brain tumor models. PLoS ONE 2010, 5, e10108. [Google Scholar] [CrossRef]
- Kong, D.; Jiang, Y.; Miao, X.; Wu, Z.; Liu, H.; Gong, W. Tadalafil enhances the therapeutic efficacy of BET inhibitors in hepatocellular carcinoma through activating Hippo pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166267. [Google Scholar] [CrossRef] [PubMed]
- Muniyan, S.; Rachagani, S.; Parte, S.; Halder, S.; Seshacharyulu, P.; Kshirsagar, P.; Siddiqui, J.A.; Vengoji, R.; Rauth, S.; Islam, R.; et al. Sildenafil Potentiates the Therapeutic Efficacy of Docetaxel in Advanced Prostate Cancer by Stimulating NO-cGMP Signaling. Clin. Cancer Res. 2020, 26, 5720–5734. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dan, Y.; Yang, D.; Hu, Y.; Zhang, L.; Zhang, C.; Zhu, H.; Cui, Z.; Li, M.; Liu, Y. The genus Anemarrhena Bunge: A review on ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2014, 153, 42–60. [Google Scholar] [CrossRef]
- Han, F.Y.; Song, X.Y.; Chen, J.J.; Yao, G.D.; Song, S.J. Timosaponin AIII: A novel potential antitumor compound from Anemarrhena asphodeloides. Steroids 2018, 140, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, J.; Han, L.; Pu, P.; Kang, C. Wnt/beta-catenin signaling in glioma. J. Neuroimmune Pharmacol. 2012, 7, 740–749. [Google Scholar] [PubMed]
- Tinsley, H.N.; Gary, B.D.; Keeton, A.B.; Lu, W.; Li, Y.; Piazza, G.A. Inhibition of PDE5 by sulindac sulfide selectively induces apoptosis and attenuates oncogenic Wnt/β-catenin-mediated transcription in human breast tumor cells. Cancer Prev. Res. 2011, 4, 1275–1284. [Google Scholar] [CrossRef]
- Deguchi, A.; Thompson, W.J.; Weinstein, I.B. Activation of protein kinase G is sufficient to induce apoptosis and inhibit cell migration in colon cancer cells. Cancer Res. 2004, 64, 3966–3973. [Google Scholar] [CrossRef]
- Kwon, I.K.; Wang, R.; Thangaraju, M.; Shuang, H.; Liu, K.; Dashwood, R.; Dulin, N.; Ganapathy, V.; Browning, D.D. PKG inhibits TCF signaling in colon cancer cells by blocking beta-catenin expression and activating FOXO4. Oncogene 2010, 29, 3423–3434. [Google Scholar] [CrossRef]
- Lin, Y.; Zhao, W.R.; Shi, W.T.; Zhang, J.; Zhang, K.Y.; Ding, Q.; Chen, X.L.; Tang, J.Y.; Zhou, Z.Y. Pharmacological Activity, Pharmacokinetics, and Toxicity of Timosaponin AIII, a Natural Product Isolated From Anemarrhena asphodeloides Bunge: A Review. Front. Pharmacol. 2020, 11, 764. [Google Scholar] [CrossRef]
- Kim, K.M.; Im, A.R.; Kim, S.H.; Hyun, J.W.; Chae, S. Timosaponin AIII inhibits melanoma cell migration by suppressing COX-2 and in vivo tumor metastasis. Cancer Sci. 2016, 107, 181–188. [Google Scholar] [CrossRef]
- Chiang, K.C.; Lai, C.Y.; Chiou, H.L.; Lin, C.L.; Chen, Y.S.; Kao, S.H.; Hsieh, Y.H. Timosaponin AIII inhibits metastasis of renal carcinoma cells through suppressing cathepsin C expression by AKT/miR-129-5p axis. J. Cell. Physiol. 2019, 234, 13332–13341. [Google Scholar] [CrossRef] [PubMed]
- Jung, O.; Lee, J.; Lee, Y.J.; Yun, J.M.; Son, Y.J.; Cho, J.Y.; Ryou, C.; Lee, S.Y. Timosaponin AIII inhibits migration and invasion of A549 human non-small-cell lung cancer cells via attenuations of MMP-2 and MMP-9 by inhibitions of ERK1/2, Src/FAK and β-catenin signaling pathways. Bioorg. Med. Chem. Lett. 2016, 26, 3963–3967. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Yang, C.W.; Wang, J.Y.; Tsai, Y.F.; Tseng, L.M.; King, K.L.; Chen, W.S.; Chiu, J.H.; Shyr, Y.M. Timosaponin AIII Suppresses Hepatocyte Growth Factor-Induced Invasive Activity through Sustained ERK Activation in Breast Cancer MDA-MB-231 Cells. Evid. Based Complement. Altern. Med. 2013, 2013, 421051. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.J.; Liu, C.J.; Ying, T.H.; Wu, P.J.; Wang, J.W.; Ting, Y.H.; Hsieh, Y.H.; Wang, S.C. Timosaponin AIII Inhibits Migration and Invasion Abilities in Human Cervical Cancer Cells through Inactivation of p38 MAPK-Mediated uPA Expression In Vitro and In Vivo. Cancers 2022, 15, 37. [Google Scholar] [CrossRef]
- Sy, L.K.; Yan, S.C.; Lok, C.N.; Man, R.Y.; Che, C.M. Timosaponin A-III induces autophagy preceding mitochondria-mediated apoptosis in HeLa cancer cells. Cancer Res. 2008, 68, 10229–10237. [Google Scholar] [CrossRef]
- Wang, H.; Dong, R.; Fan, W.W.; Zheng, X.C.; Li, A.M.; Wang, W.D. Timosaponin A-III induces autophagy of T-cell acute lymphoblastic leukemia Jurkat cells via inhibition of the PI3K/Akt/mTOR pathway. Oncol. Rep. 2019, 41, 2937–2944. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Lou, L.L.; Song, S.J.; Yao, G.D.; Ge, M.Y.; Hayashi, T.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Timosaponin AIII induces apoptosis and autophagy in human melanoma A375-S2 cells. Arch. Pharm. Res. 2017, 40, 69–78. [Google Scholar] [CrossRef]
- Chen, J.R.; Jia, X.H.; Wang, H.; Yi, Y.J.; Wang, J.Y.; Li, Y.J. Timosaponin A-III reverses multi-drug resistance in human chronic myelogenous leukemia K562/ADM cells via downregulation of MDR1 and MRP1 expression by inhibiting PI3K/Akt signaling pathway. Int. J. Oncol. 2016, 48, 2063–2070. [Google Scholar] [CrossRef]
- Song, X.Y.; Han, F.Y.; Chen, J.J.; Wang, W.; Zhang, Y.; Yao, G.D.; Song, S.J. Timosaponin AIII, a steroidal saponin, exhibits antitumor effect on taxol-resistant cells in vitro and in vivo. Steroids 2019, 146, 57–64. [Google Scholar] [CrossRef]
- Ko, H.M.; Jee, W.; Park, D.I.; Kim, K.I.; Jung, J.H.; Jang, H.J. The Antitumor Effect of Timosaponin A3 through c-Myc Inhibition in Colorectal Cancer Cells and Combined Treatment Effect with 5-FU or Doxorubicin. Int. J. Mol. Sci. 2022, 23, 11900. [Google Scholar] [CrossRef]
- Li, X.; Lu, W.; Zhou, T.; Zhao, F.; Yang, L. Timosaponin AIII Suppresses RAP1 Signaling Pathway to Enhance the Inhibitory Effect of Paclitaxel on Nasopharyngeal Carcinoma. Comput. Math. Methods Med. 2022, 2022, 6756676. [Google Scholar] [CrossRef]
- King, F.W.; Fong, S.; Griffin, C.; Shoemaker, M.; Staub, R.; Zhang, Y.L.; Cohen, I.; Shtivelman, E. Timosaponin AIII is preferentially cytotoxic to tumor cells through inhibition of mTOR and induction of ER stress. PLoS ONE 2009, 4, e7283. [Google Scholar] [CrossRef]
- Nho, K.J.; Chun, J.M.; Kim, H.K. Induction of mitochondria-dependent apoptosis in HepG2 human hepatocellular carcinoma cells by timosaponin A-III. Environ. Toxicol. Pharmacol. 2016, 45, 295–301. [Google Scholar] [CrossRef]
- MarElia, C.B.; Sharp, A.E.; Shemwell, T.A.; Clare Zhang, Y.; Burkhardt, B.R. Anemarrhena asphodeloides Bunge and its constituent timosaponin-AIII induce cell cycle arrest and apoptosis in pancreatic cancer cells. FEBS Open Bio 2018, 8, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, K.H.; Lee, I.S.; Park, J.Y.; Na, Y.C.; Chung, W.S.; Jang, H.J. Apoptosis and G2/M cell cycle arrest induced by a timosaponin A3 from Anemarrhena asphodeloides Bunge on AsPC-1 pancreatic cancer cells. Phytomedicine 2019, 56, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Deng, X.; Sun, X.; Dong, J.; Huang, J. Inhibition of autophagy enhances timosaponin AIII-induced lung cancer cell apoptosis and antitumor effect in vitro and in vivo. Life Sci. 2020, 257, 118040. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Qu, J.; Gao, Z.; Qi, Q.; Yin, H.; Zhu, L.; Wu, Y.; Liu, W.; Yang, J.; Huang, X. Timosaponin AIII Induces G2/M Arrest and Apoptosis in Breast Cancer by Activating the ATM/Chk2 and p38 MAPK Signaling Pathways. Front. Pharmacol. 2021, 11, 601468. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.-F.; Pan, H.-J.; Abudurezeke, N.; Yuan, C.-L.; Yuan, Y.-L.; Zhao, S.-D.; Zhang, D.-D.; Huang, S. Functional Axis of PDE5/cGMP Mediates Timosaponin-AIII-Elicited Growth Suppression of Glioblastoma U87MG Cells. Molecules 2023, 28, 3795. https://doi.org/10.3390/molecules28093795
Liao Y-F, Pan H-J, Abudurezeke N, Yuan C-L, Yuan Y-L, Zhao S-D, Zhang D-D, Huang S. Functional Axis of PDE5/cGMP Mediates Timosaponin-AIII-Elicited Growth Suppression of Glioblastoma U87MG Cells. Molecules. 2023; 28(9):3795. https://doi.org/10.3390/molecules28093795
Chicago/Turabian StyleLiao, Ya-Fang, Hui-Jun Pan, Nuerziba Abudurezeke, Chun-Lu Yuan, Yan-Li Yuan, Shu-Da Zhao, Dan-Dan Zhang, and Shuang Huang. 2023. "Functional Axis of PDE5/cGMP Mediates Timosaponin-AIII-Elicited Growth Suppression of Glioblastoma U87MG Cells" Molecules 28, no. 9: 3795. https://doi.org/10.3390/molecules28093795