
Anxiolytic-like Effects and Quantitative EEG Profile of Palmitone Induces Responses Like Buspirone Rather Than Diazepam as Clinical Drugs
 , , , and
, , , and 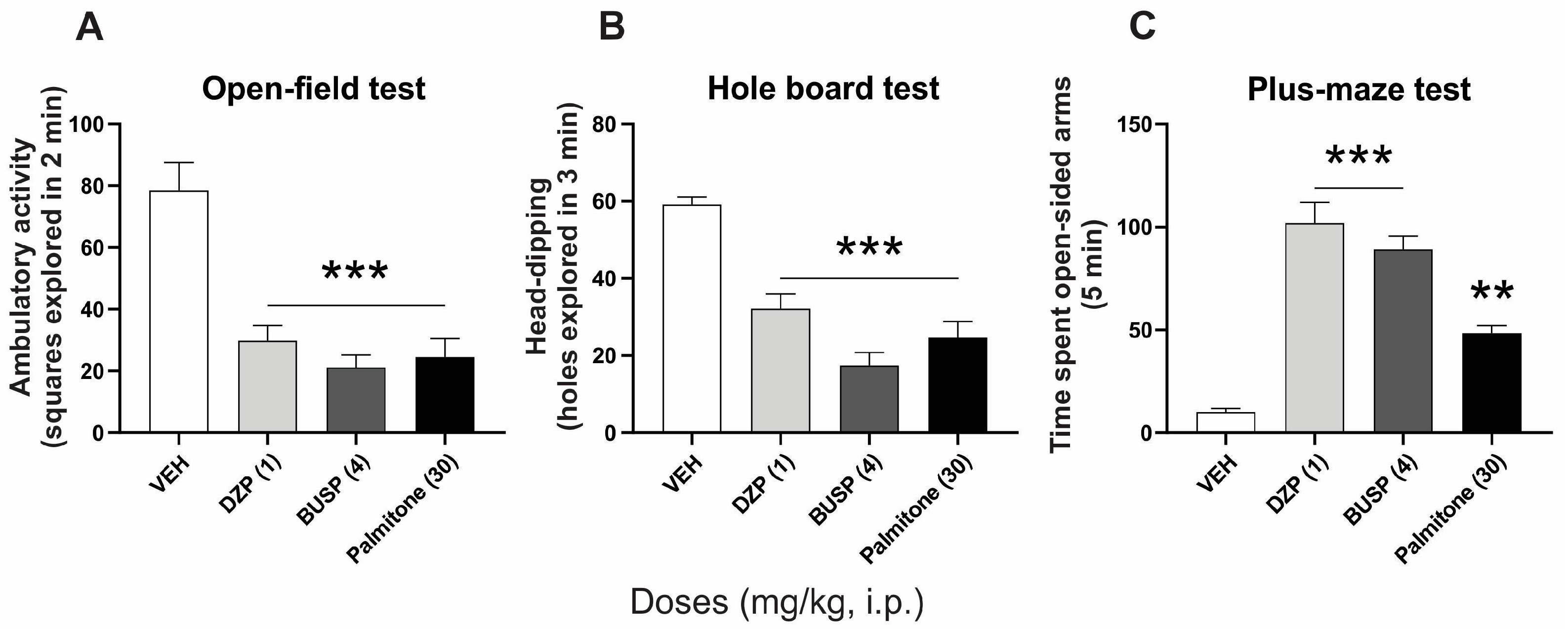{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anxiolytic-like Effects of Palmitone and Clinical Anxiolytics Drugs
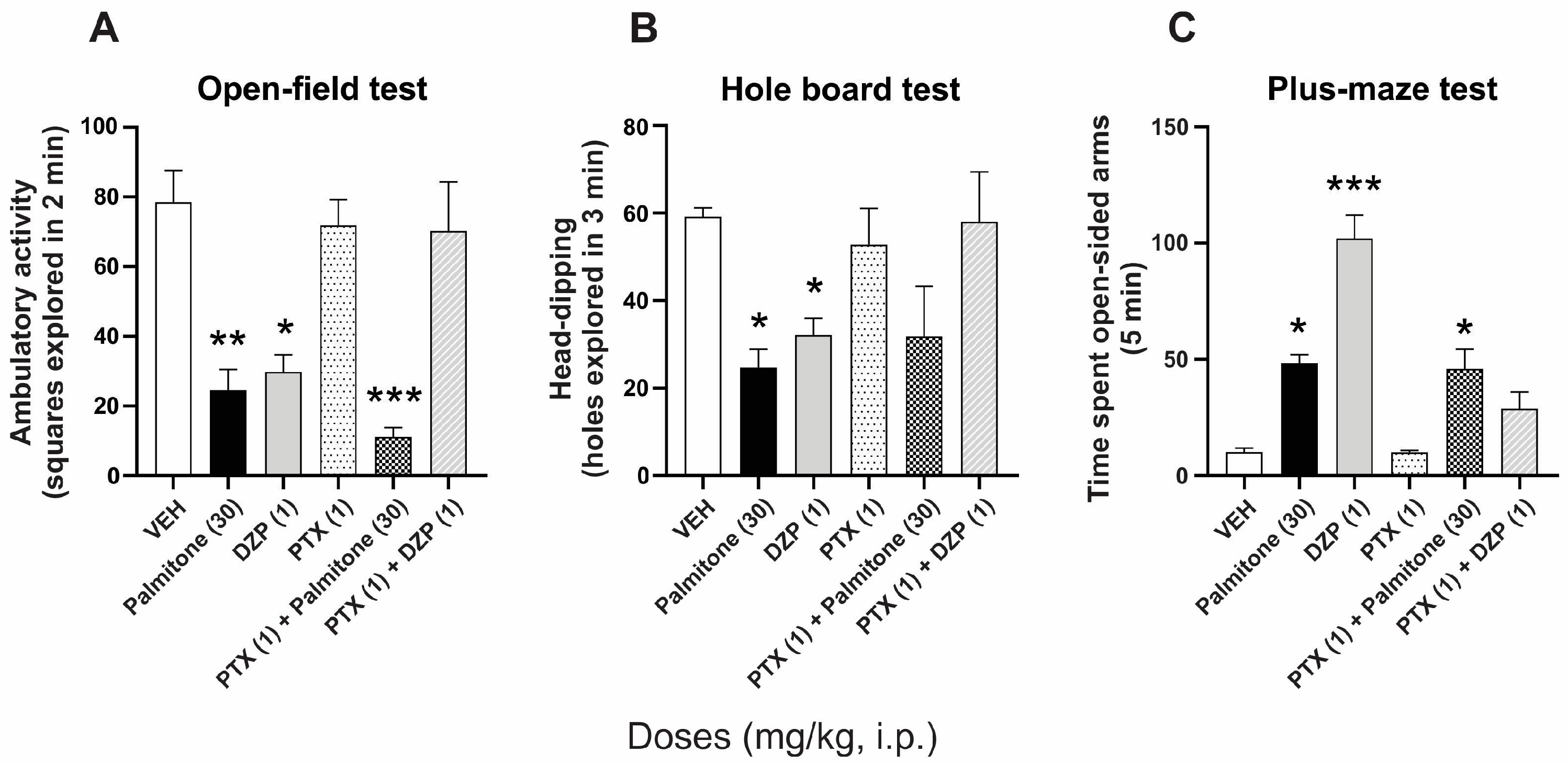
2.2. GABAA Receptors Involvement in the Anxiolytic-like Effects of Palmitone
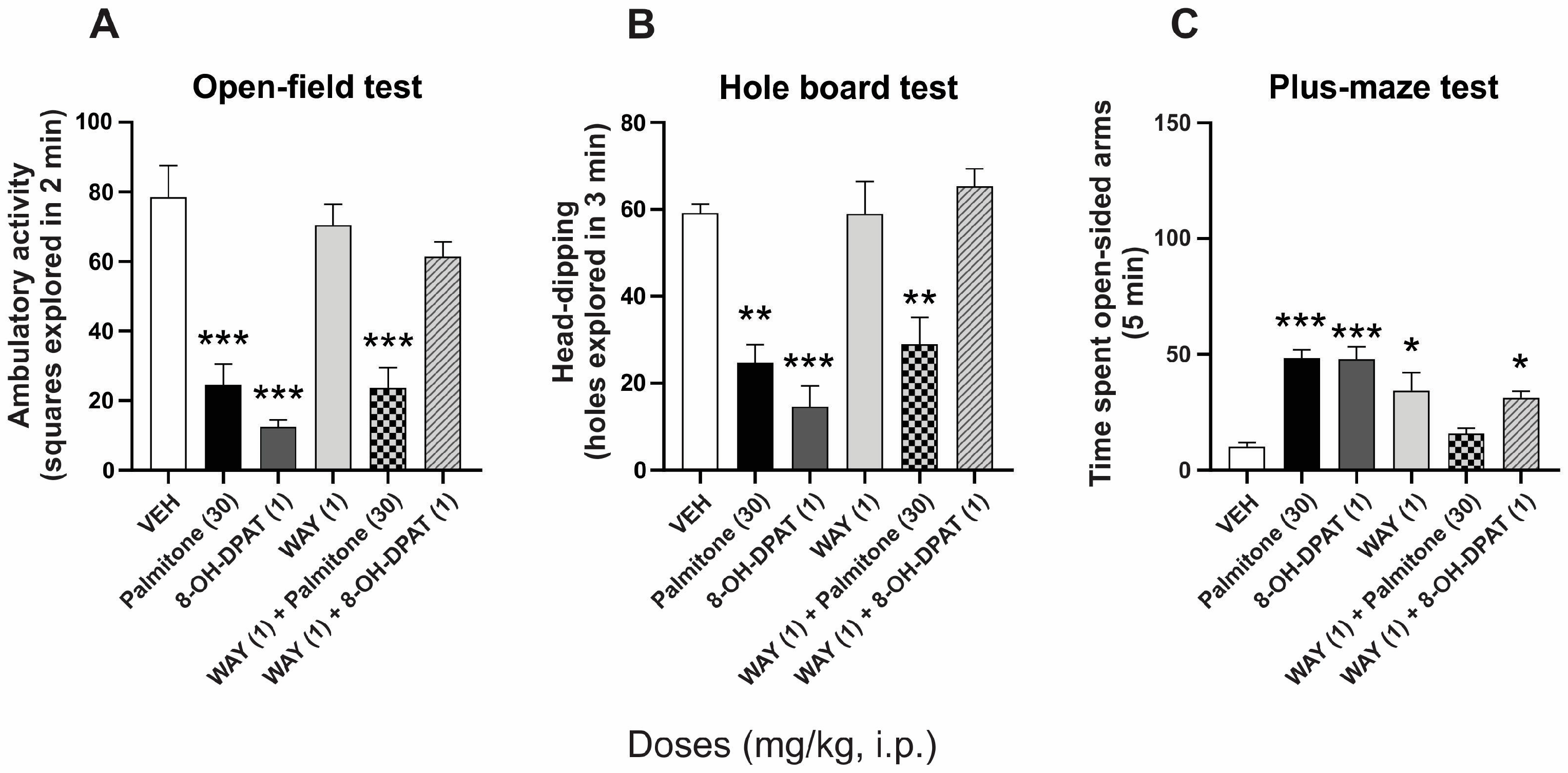
2.3. Serotonin 5-HT1A Receptors Involvement in the Anxiolytic-like Effects of Palmitone
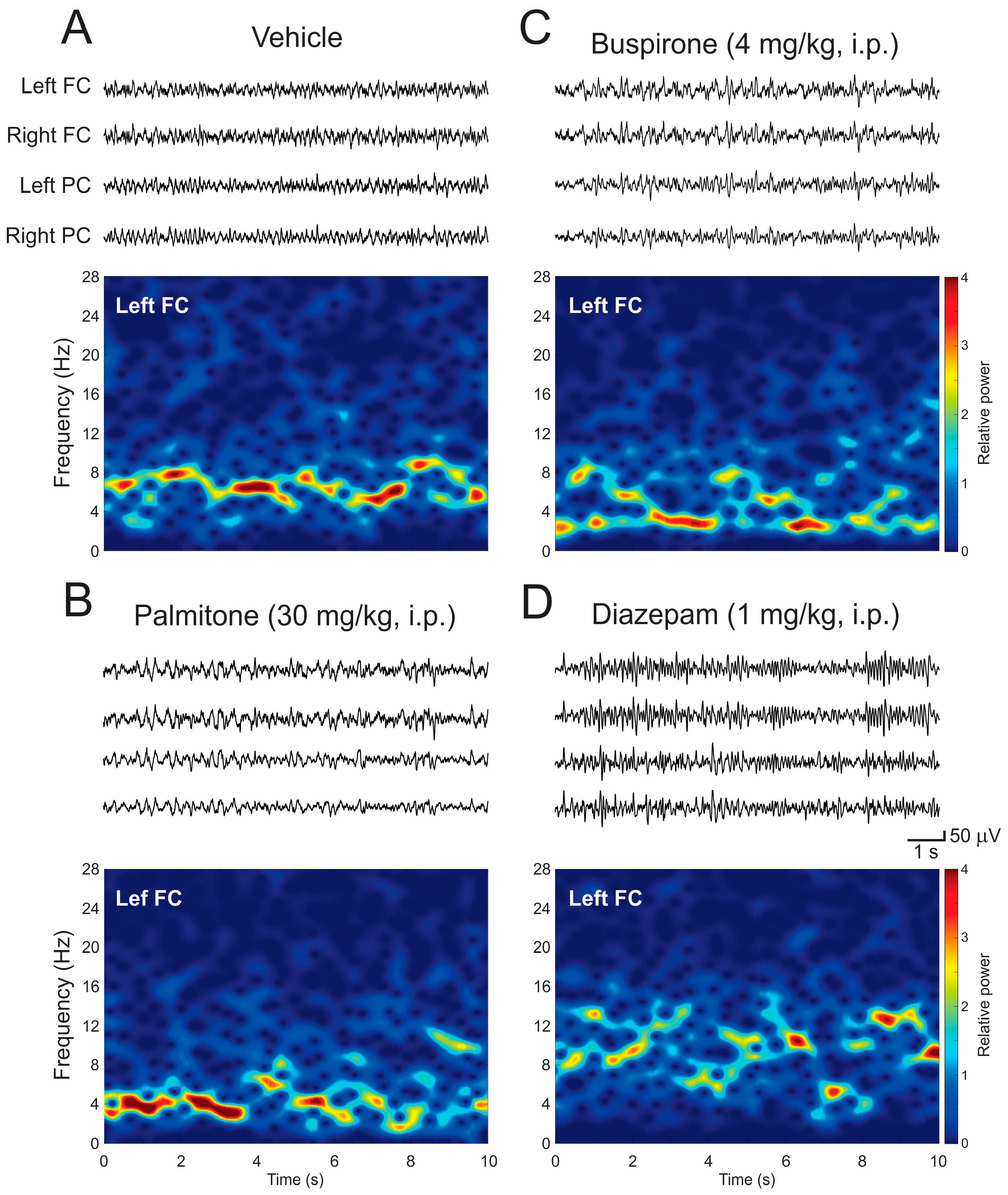
2.4. EEG Patterns and Spectral Power Analysis
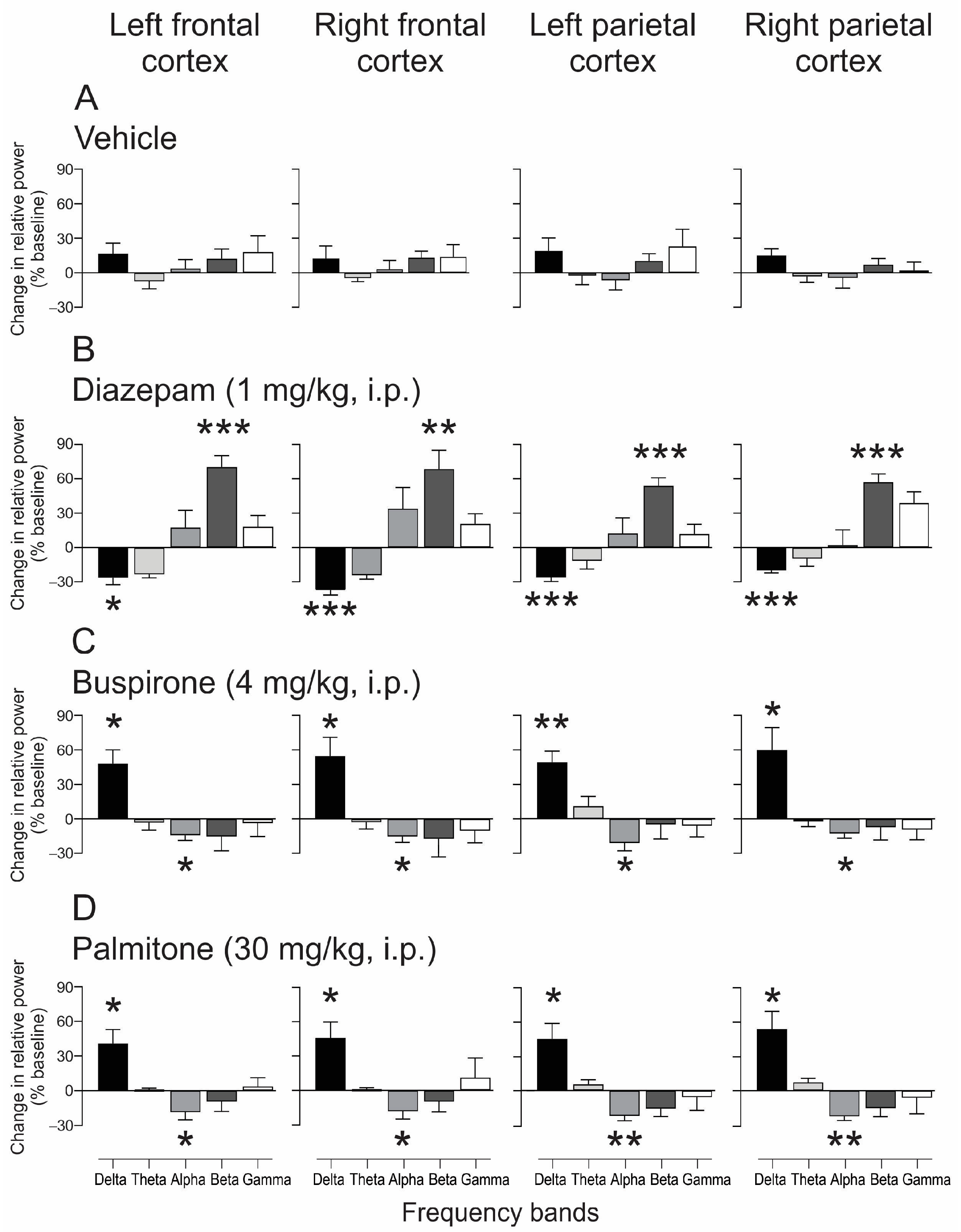
2.4.1. EEG Changes after Treatment of Palmitone and Anxiolytics Drugs
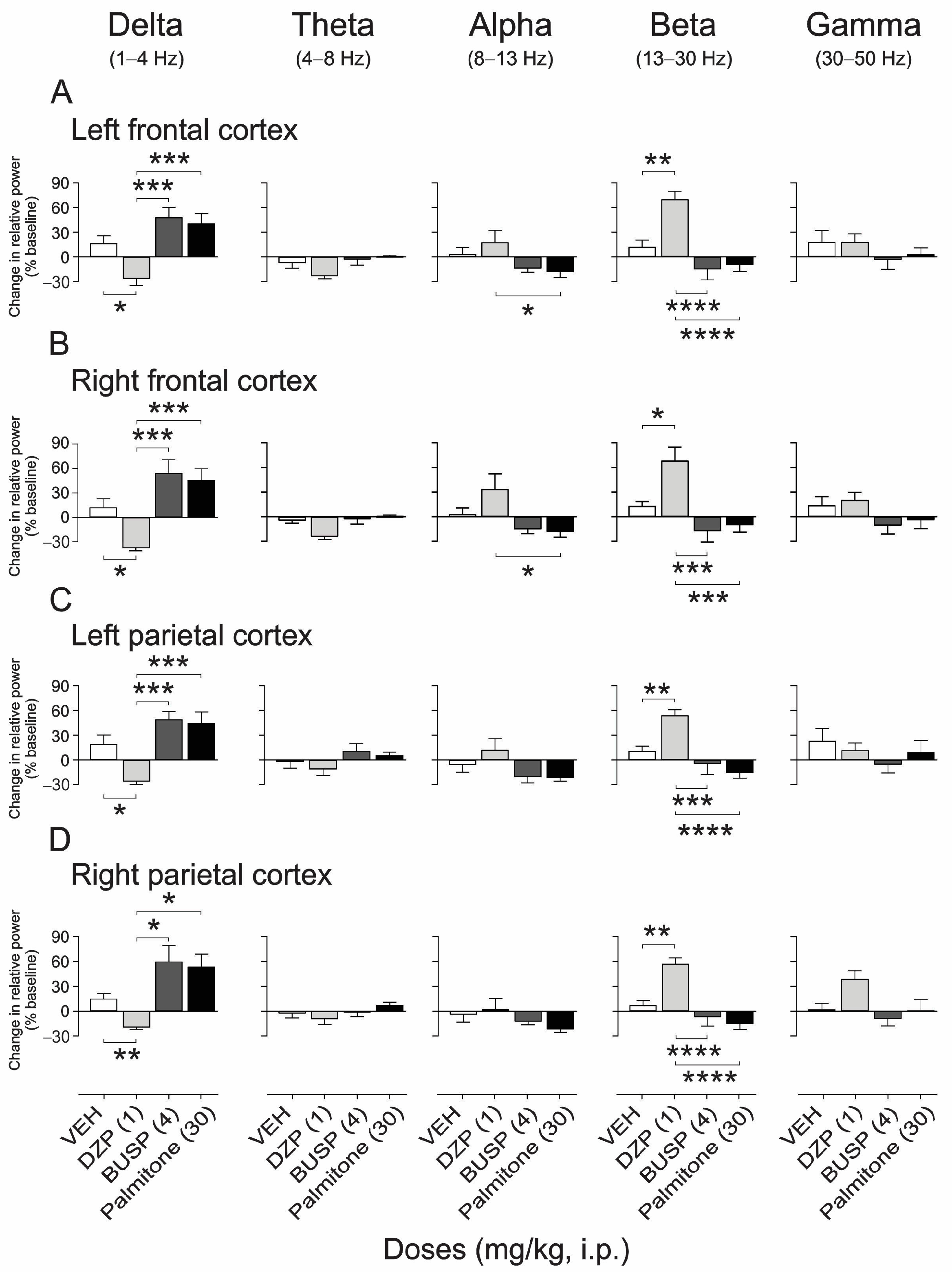
2.4.2. EEG Spectral Power Analysis after Palmitone and Anxiolytic Drugs
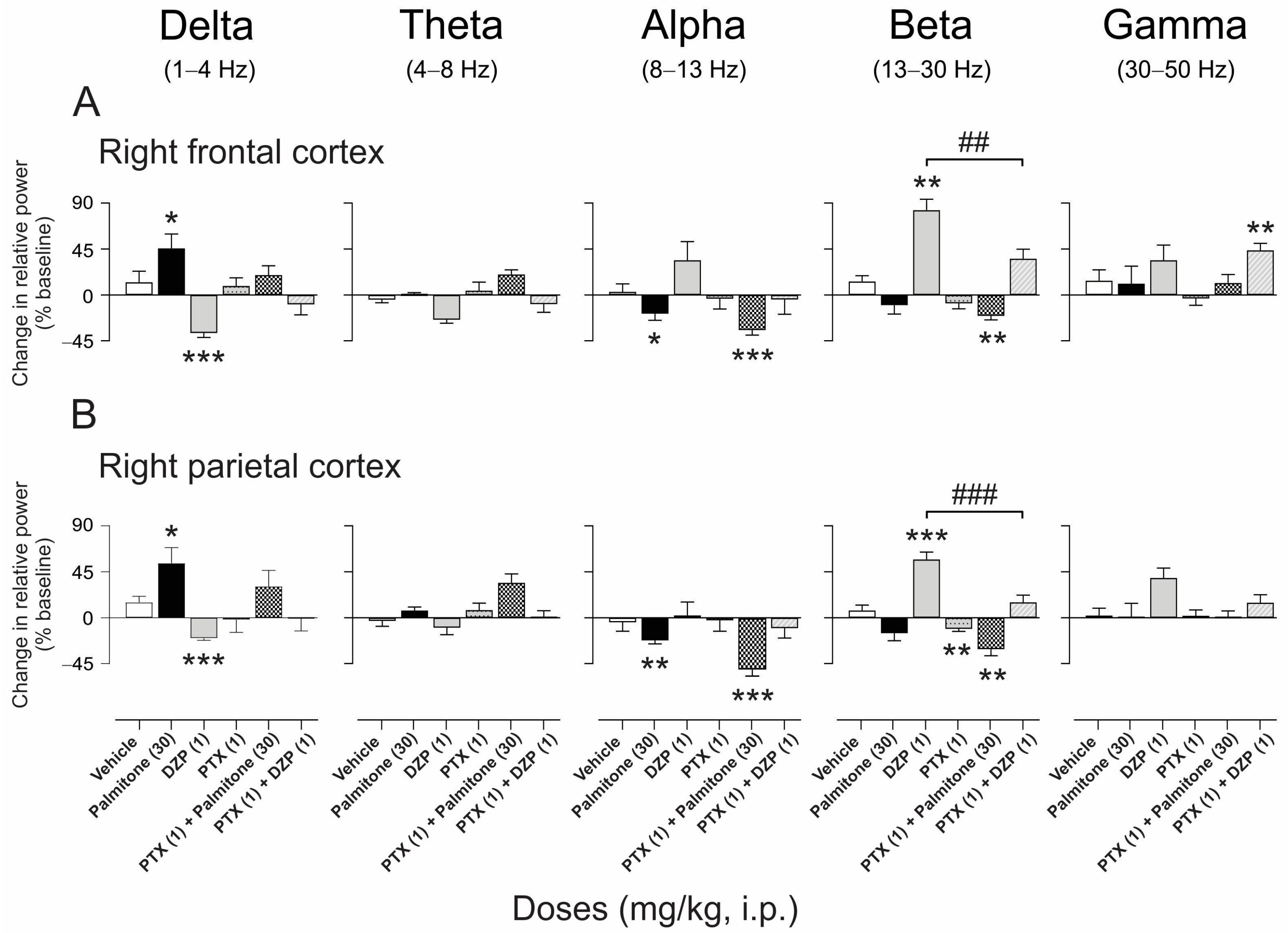
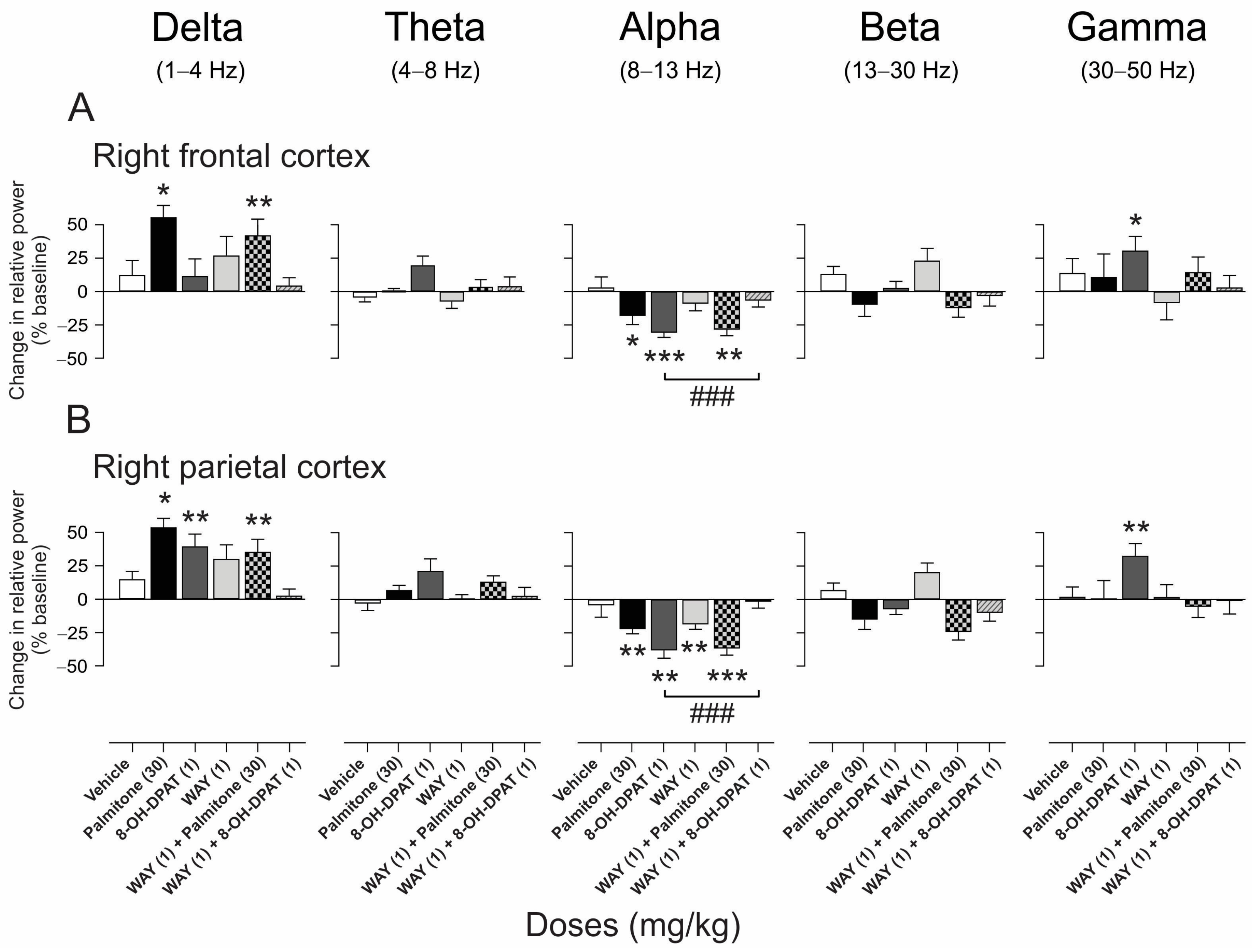
2.4.3. EEG Spectral Power Analysis after Palmitone Compared with Agonist and Antagonist of GABAA and 5-HT1A Receptors
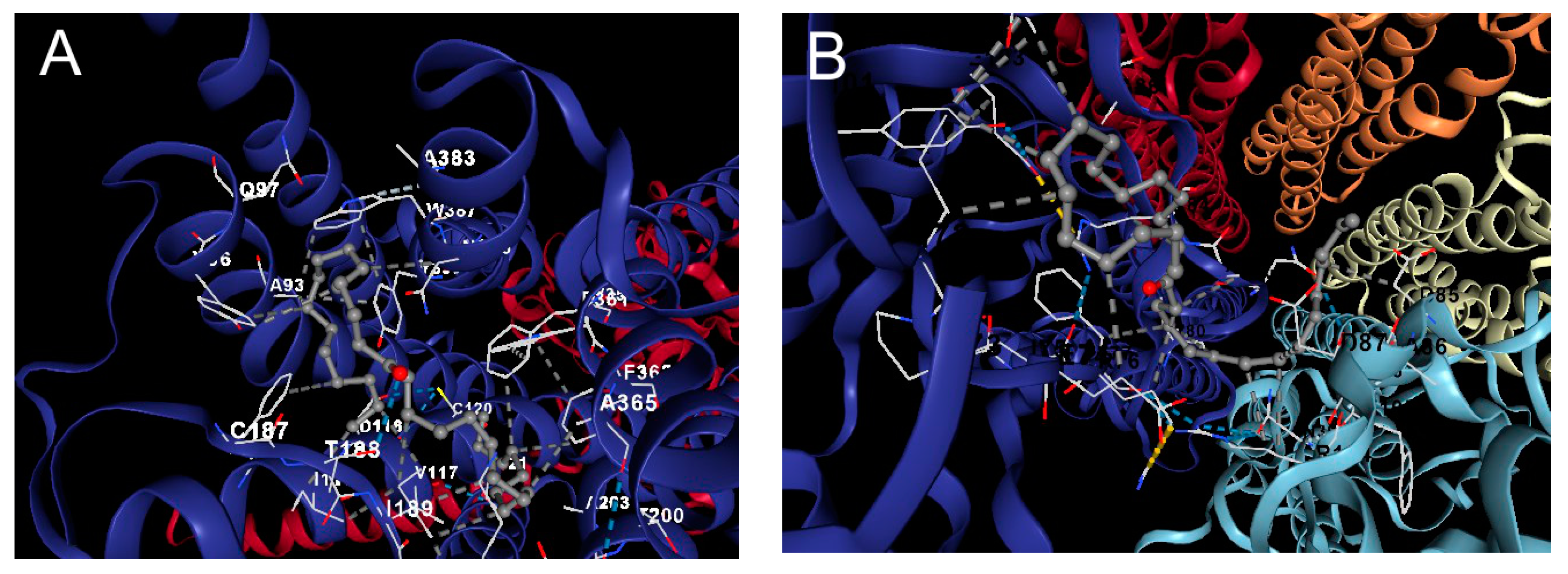
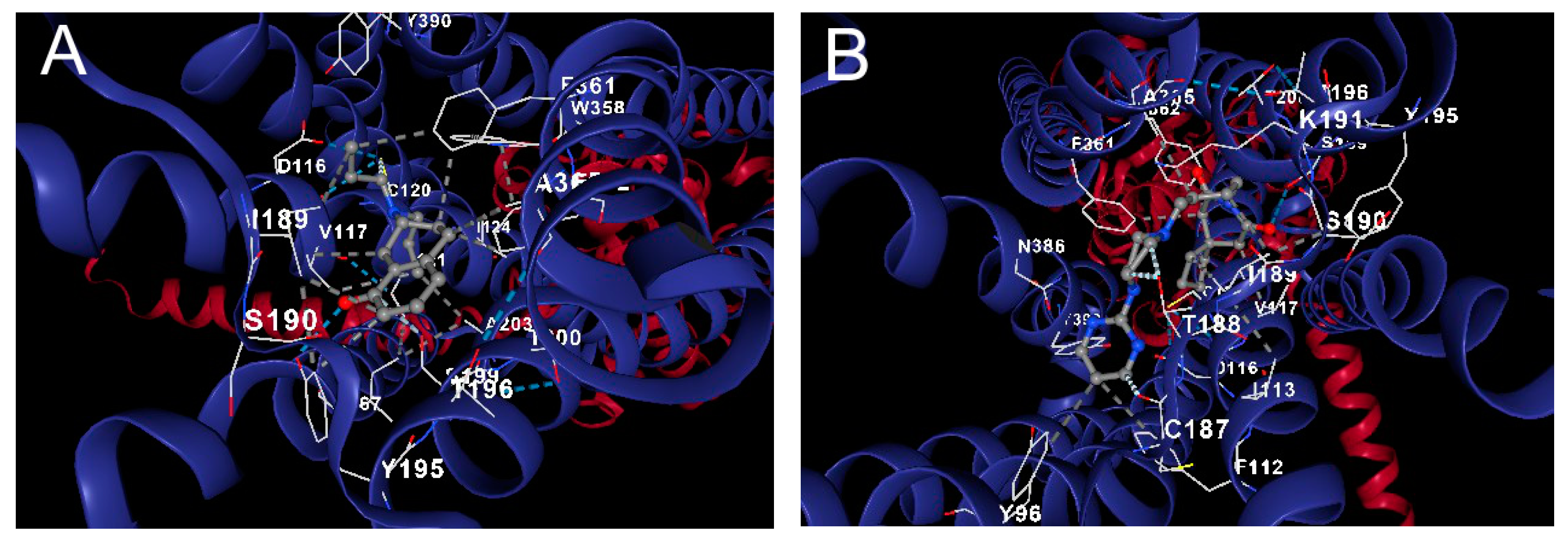
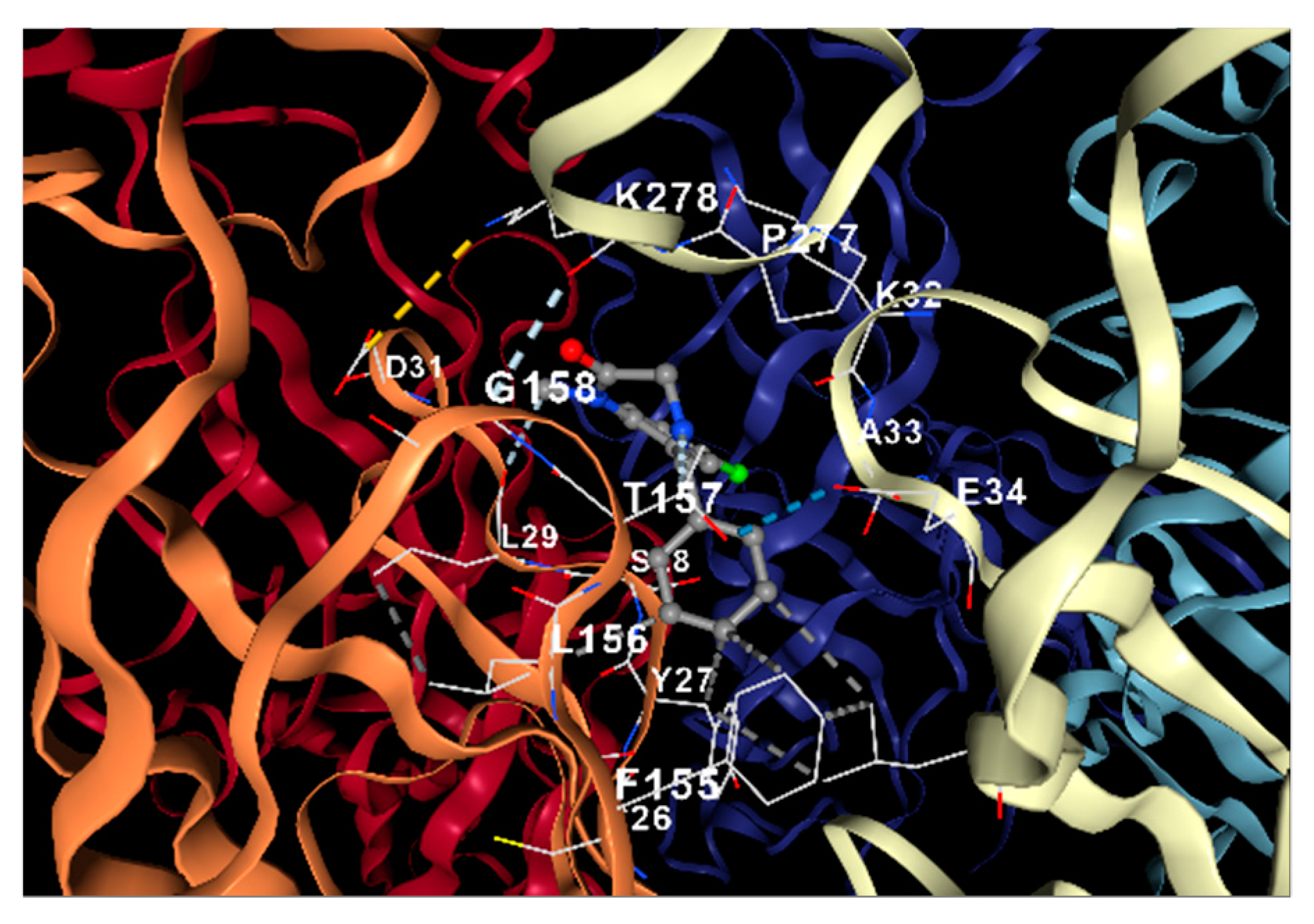
2.4.4. Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
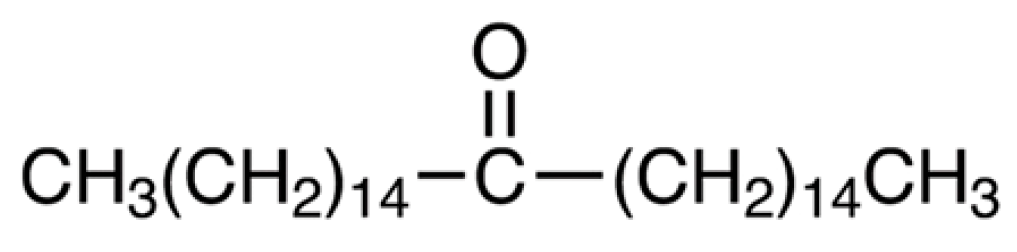
4.2. Reagents and Drugs
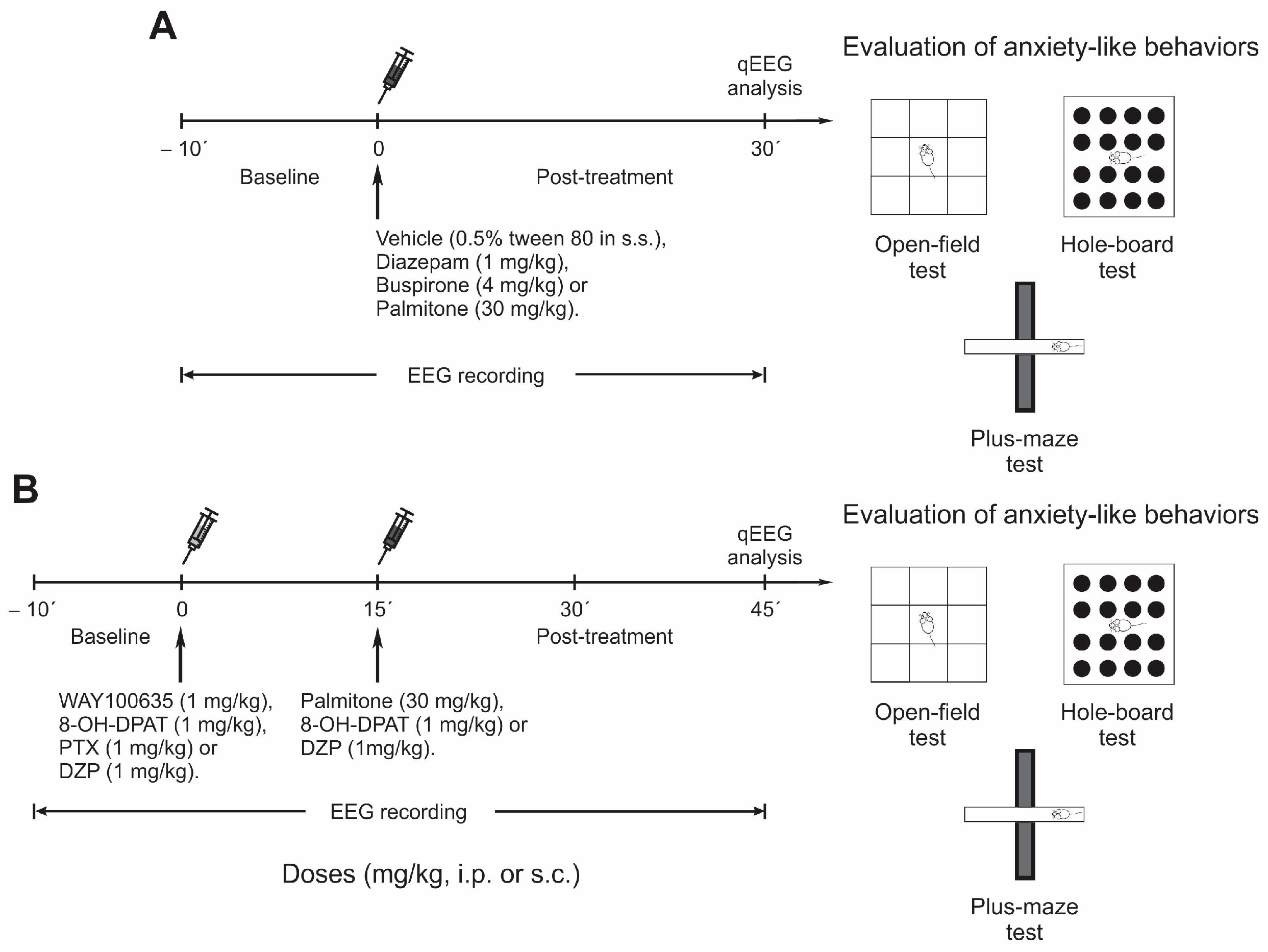
4.3. Experimental Design
- -
- Group 1 was injected with vehicle (0.5% tween 80 in s.s.).
- -
- Group 2 was administered palmitone (30 mg/kg, i.p.).
- -
- Groups 3 and 4 were injected with DZP (1 mg/kg, i.p.) or BUSP (4 mg/kg, i.p.), respectively.
- -
- Group 5 received DZP (GABAA agonist, 1 mg/kg, i.p.) plus PTX (blocker of GABAA receptor chloride channel, 1 mg/kg, i.p.).
- -
- Groups 6 and 7 received 8-OH-DPAT (5-HT1A agonist, 1 mg/kg, i.p.) alone, or 8-OH-DPAT plus WAY (5-HT1A antagonist, 1 mg/kg, i.p.), respectively.
- -
- Groups 8 and 9 were administered with each antagonist alone: PTX (1 mg/kg, i.p.) or WAY (1 mg/kg, i.p.), respectively.
- -
- Groups 10 and 11 received a combination of palmitone (30 mg/kg, i.p.) plus PTX (1 mg/kg, i.p.) or palmitone plus WAY.
4.4. Anxiety Behavioral Tests
4.4.1. Open-Field
4.4.2. Hole-Board
4.4.3. Plus-Maze
4.5. Surgery for EEG and Recordings
4.5.1. Electrode Implantation
4.5.2. EEG Recordings
4.5.3. EEG Spectral Power Analysis
4.6. Docking Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Remes, O.; Brayne, C.; van der Linde, R.; Lafortune, L. A systematic review of reviews on the prevalence of anxiety disorders in adult populations. Brain Behav. 2016, 6, e00497. [Google Scholar] [CrossRef] [PubMed]
- Santomauro, D.F.; Herrera, A.M.M.; Shadid, J.; Zheng, P.; Ashbaugh, C.; Pigott, D.M.; Abbafati, C.; Adolph, C.; Amlag, J.O.; Aravkin, A.Y.; et al. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B.; Michaelis, S.; Wedekind, D. Treatment of anxiety disorders. Dialog. Clin. Neurosci. 2017, 19, 93–107. [Google Scholar] [CrossRef]
- Murrough, J.W.; Yaqubi, S.; Sayed, S.; Charney, D.S. Emerging drugs for the treatment of anxiety. Expert Opin. Emerg. Drugs 2015, 20, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, E.J.; Mathew, S.J. Anxiety disorders: A comprehensive review of pharmacotherapies. Mt. Sinai J. Med. A J. Transl. Pers. Med. 2008, 75, 248–262. [Google Scholar] [CrossRef]
- Rickels, K. Antidepressants for the Treatment of Generalized Anxiety Disorder: A Placebo-Controlled Comparison of Imipramine, Trazodone, and Diazepam. Arch. Gen. Psychiatry 1993, 50, 884. [Google Scholar] [CrossRef]
- Eison, M.S. Serotonin: A common neurobiologic substrate in anxiety and depression. J. Clin. Psychopharmacol. 1990, 10, 26S–30S. [Google Scholar] [CrossRef]
- Su, K.-P.; Tseng, P.-T.; Lin, P.-Y.; Okubo, R.; Chen, T.-Y.; Chen, Y.-W.; Matsuoka, Y.J. Association of Use of Omega-3 Polyunsaturated Fatty Acids with Changes in Severity of Anxiety Symptoms: A Systematic Review and Meta-Analysis. JAMA Netw. Open 2018, 1, e182327. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Morales, B.; Cueto-Escobedo, J.; Guillén-Ruiz, G.; Rodríguez-Landa, J.F.; Contreras, C.M. A Fatty Acids Mixture Reduces Anxiety-Like Behaviors in Infant Rats Mediated by GABAA Receptors. BioMed Res. Int. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Frausto-González, O.; Bautista, C.J.; Narváez-González, F.; Hernández-León, A.; Estrada-Camarena, E.; Rivero-Cruz, F.; González-Trujano, M.E. Bertholletia excelsa Seeds Reduce Anxiety-Like Behavior, Lipids, and Overweight in Mice. Molecules 2021, 26, 3212. [Google Scholar] [CrossRef]
- Rodríguez-Landa, J.F.; García-Ríos, R.I.; Cueto-Escobedo, J.; Bernal-Morales, B.; Contreras, C.M. Participation of GABAAChloride Channels in the Anxiolytic-Like Effects of a Fatty Acid Mixture. BioMed Res. Int. 2013, 2013, 121794. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Hernández, E.; Martínez, A.L.; González-Trujano, M.E.; Moreno, J.; Vibrans, H.; Soto-Hernández, M. Pharmacological evaluation of the anxiolytic and sedative effects of Tilia americana L. var. mexicana in mice. J. Ethnopharmacol. 2007, 109, 140–145. [Google Scholar] [CrossRef]
- Elyasi, H.; Sepahvand, A.; Rahimi, H.; Nafari, A.; Azizi, S.; Khadem, E.; Zamani, A.; Behnaminia, N.; Bahmani, M. Fatty Acids and Herbal Medicine. Curr. Tradit. Med. 2019, 5, 246–256. [Google Scholar] [CrossRef]
- Swastika; Chaturvedi, S.; Kaul, A.; Hazari, P.P.; Jha, P.; Pal, S.; Lal, S.; Singh, B.; Barthélémy, P.; Mishra, A.K. Evaluation of BBB permeable nucleolipid (NLDPU): A di-C15-ketalised palmitone appended uridine as neuro-tracer for SPECT. Int. J. Pharm. 2019, 565, 269–282. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Tapia, E.; López-Meraz, L.; Navarrete, A.; Reyes-Ramírez, A.; Martínez, A. Anticonvulsant Effect of Annona diversifolia Saff. and Palmitone on Penicillin-induced Convulsive Activity. A Behavioral and EEG Study in Rats. Epilepsia 2006, 47, 1810–1817. [Google Scholar] [CrossRef]
- Cano-Europa, E.; González-Trujano, M.E.; Reyes-Ramírez, A.; Hernández-García, A.; Blas-Valdivia, V.; Ortiz-Butrón, R. Palmitone prevents pentylenetetrazole-caused neuronal damage in the CA3 hippocampal region of prepubertal rats. Neurosci. Lett. 2010, 470, 111–114. [Google Scholar] [CrossRef]
- Carballo, A.I.; Martínez, A.L.; González-Trujano, M.E.; Pellicer, F.; Ventura-Martínez, R.; Díaz-Reval, M.I.; López-Muñoz, F.J. Antinociceptive activity of Annona diversifolia Saff. leaf extracts and palmitone as a bioactive compound. Pharmacol. Biochem. Behav. 2010, 95, 6–12. [Google Scholar] [CrossRef]
- Shanker, K.S.; Kanjilal, S.; Rao, B.V.S.K.; Kishore, K.H.; Misra, S.; Prasad, R.B.N. Isolation and antimicrobial evaluation of isomeric hydroxy ketones in leaf cuticular waxes of Annona squamosa. Phytochem. Anal. 2007, 18, 7–12. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Martínez, A.; Reyes-Ramírez, A.; Reyes-Trejo, B.; Navarrete, A. Palmitone Isolated from Annona diversifolia Induces an Anxiolytic-Like Effect in Mice. Planta Med. 2006, 72, 703–707. [Google Scholar] [CrossRef]
- López-Rubalcava, C.; Piña-Medina, B.; Estrada-Reyes, R.; Heinze, G.; Martínez-Vázquez, M. Anxiolytic-like actions of the hexane extract from leaves of Annona cherimolia in two anxiety paradigms: Possible involvement of the GABA/benzodiazepine receptor complex. Life Sci. 2006, 78, 730–737. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Navarrete, A.; Reyes, B.; Cedillo-Portugal, E.; Hong, E. Anticonvulsant Properties and Bio-Guided Isolation of Palmitone from Leaves of Annona diversifolia. Planta Medica 2001, 67, 136–141. [Google Scholar] [CrossRef]
- Lezak, K.R.; Missig, G.; Carlezon, W.A., Jr. Behavioral methods to study anxiety in rodents. Dialog- Clin. Neurosci. 2017, 19, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Mathersul, D.; Williams, L.M.; Hopkinson, P.J.; Kemp, A.H. Investigating models of affect: Relationships among EEG alpha asymmetry, depression, and anxiety. Emotion 2008, 8, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, R.; Jorgensen, R.S.; Kim, S. Depression, anxiety, and resting frontal EEG asymmetry: A meta-analytic review. J. Abnorm. Psychol. 2006, 115, 715–729. [Google Scholar] [CrossRef] [Green Version]
- González-Trujano, M.E.; Martínez-González, C.L.; Flores-Carrillo, M.; Luna-Nophal, S.I.; Contreras-Murillo, G.; Magdaleno-Madrigal, V.M. Behavioral and electroencephalographic evaluation of the anticonvulsive activity of Moringa oleifera leaf non-polar extracts and one metabolite in PTZ-induced seizures. Phytomedicine 2018, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- González-Trujano, M.E.; Urbina-Trejo, E.; Santos-Valencia, F.; Villasana-Salazar, B.; Carmona-Aparicio, L.; Martínez-Vargas, D. Pharmacological and toxicological effects of Ruta chalepensis L. on experimentally induced seizures and electroencephalographic spectral power in mice. J. Ethnopharmacol. 2021, 271, 113866. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Krengel, F.; Reyes-Chilpa, R.; Villasana-Salazar, B.; González-Gómez, J.D.; Santos-Valencia, F.; Urbina-Trejo, E.; Martínez, A.; Martínez-Vargas, D. Tabernaemontana arborea and ibogaine induce paroxysmal EEG activity in freely moving mice: Involvement of serotonin 5-HT1A receptors. Neurotoxicology 2022, 89, 79–91. [Google Scholar] [CrossRef]
- Pereira, M.; da Silva, T.; Aguiar, A.; Oliva, G.; Guido, R.; Yokoyama-Yasunaka, J.; Uliana, S.; Lopes, L. Chemical Composition, Antiprotozoal and Cytotoxic Activities of Indole Alkaloids and Benzofuran Neolignan of Aristolochia cordigera. Planta Med. 2017, 83, 912–920. [Google Scholar] [CrossRef]
- Pooja-Reddy, C.; Amreen-Sultana, S.; Eswar-Tony, D.; Rama-Rao, N. Preclinical Evaluation of Anxiolytic Activity of Aristolochia indica (Linn) Leaf Extract in Swiss Albino Mice. SAJP 2016, 5, 6–8. [Google Scholar]
- Li, S.; Guo, C.; Zhang, X.; Liu, X.; Mu, J.; Liu, C.; Peng, Y.; Chang, M. Self-assembling modified neuropeptide S enhances nose-to-brain penetration and exerts a prolonged anxiolytic-like effect. Biomater. Sci. 2021, 9, 4765–4777. [Google Scholar] [CrossRef]
- Moon, M.L.; Joesting, J.J.; Lawson, M.A.; Chiu, G.S.; Blevins, N.A.; Kwakwa, K.A.; Freund, G.G. The saturated fatty acid, palmitic acid, induces anxiety-like behavior in mice. Metabolism 2014, 63, 1131–1140. [Google Scholar] [CrossRef] [Green Version]
- Hoehn-Saric, R. Neurotransmitters in Anxiety. Arch. Gen. Psychiatry 1982, 39, 735. [Google Scholar] [CrossRef]
- Mamalakis, G.; Kafatos, A.; Tornaritis, M.; Alevizos, B. Anxiety and adipose essential fatty acid precursors for prostaglandin E1 and E2. J. Am. Coll. Nutr. 1998, 17, 239–243. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; López-Meraz, L.; Reyes-Ramírez, A.; Aguillón, M.; Martínez, A. Effect of repeated administration of Annona diversifolia Saff. (ilama) extracts and palmitone on rat amygdala kindling. Epilepsy Behav. 2009, 16, 590–595. [Google Scholar] [CrossRef]
- Majewska, M.D.; Harrison, N.L.; Schwartz, R.D.; Barker, J.L.; Paul, S.M. Steroid Hormone Metabolites Are Barbiturate-Like Modulators of the GABA Receptor. Science 1986, 232, 1004–1007. [Google Scholar] [CrossRef] [Green Version]
- Hernández-León, A.; González-Trujano, M.E.; Fernández-Guasti, A. The anxiolytic-like effect of rutin in rats involves GABAA receptors in the basolateral amygdala. Behav. Pharmacol. 2017, 28, 303–312. [Google Scholar] [CrossRef]
- Newson, J.J.; Thiagarajan, T.C. EEG Frequency Bands in Psychiatric Disorders: A Review of Resting State Studies. Front. Hum. Neurosci. 2019, 12, 521. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.X. Where Does EEG Come from and What Does It Mean? Trends Neurosci. 2017, 40, 208–218. [Google Scholar] [CrossRef]
- Drevets, W.C.; Price, J.L.; Furey, M.L. Brain structural and functional abnormalities in mood disorders: Implications for neurocircuitry models of depression. Brain Struct. Funct. 2008, 213, 93–118. [Google Scholar] [CrossRef] [Green Version]
- Liskiewicz, A.; Przybyła, M.; Wojakowska, A.; Marczak, Ł.; Bogus, K.; Nowacka-Chmielewska, M.; Liśkiewicz, D.; Małecki, A.; Barski, J.; Lewin-Kowalik, J.; et al. Physical activity reduces anxiety and regulates brain fatty acid synthesis. Mol. Brain 2020, 13, 62. [Google Scholar] [CrossRef]
- Manor, R.; Kumarnsit, E.; Samerphob, N.; Rujiralai, T.; Puangpairote, T.; Cheaha, D. Characterization of pharmaco-EEG fingerprint and sleep-wake profiles of Lavandula angustifolia Mill. essential oil inhalation and diazepam administration in rats. J. Ethnopharmacol. 2021, 276, 114193. [Google Scholar] [CrossRef] [PubMed]
- Jongsma, M.L.A.; van Rijn, C.M.; van Egmond, J.; van Schaijk, W.J.; Sambeth, A.; Coenen, A.M.L. The influence of diazepam on the electroencephalogram-evoked potential interrelation in rats. Neurosci. Lett. 2000, 293, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.S. The value of genetic and pharmacological approaches to understanding the complexities of GABAA receptor subtype functions: The anxiolytic effects of benzodiazepines. Pharmacol. Biochem. Behav. 2008, 90, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Leiser, S.C.; Dunlop, J.; Bowlby, M.R.; Devilbiss, D.M. Aligning strategies for using EEG as a surrogate biomarker: A review of preclinical and clinical research. Biochem. Pharmacol. 2011, 81, 1408–1421. [Google Scholar] [CrossRef]
- Christian, E.P.; Snyder, D.H.; Song, W.; Gurley, D.A.; Smolka, J.; Maier, D.L.; Ding, M.; Gharahdaghi, F.; Liu, X.F.; Chopra, M.; et al. EEG-β/γ spectral power elevation in rat: A translatable biomarker elicited by GABAAα2/3-positive allosteric modulators at nonsedating anxiolytic doses. J. Neurophysiol. 2015, 113, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Coenen, A.M.L.; Van Luijtelaar, E.L.J.M. Pharmacological Dissociation of EEG and Behavior: A Basic Problem in Sleep-Wake Classification. Sleep 1991, 14, 464–465. [Google Scholar]
- van Lier, H.; Drinkenburg, W.H.I.M.; van Eeten, Y.J.W.; Coenen, A.M.L. Effects of diazepam and zolpidem on EEG beta frequencies are behavior-specific in rats. Neuropharmacology 2004, 47, 163–174. [Google Scholar] [CrossRef]
- Gilles, C.; Luthringer, R. Pharmacological models in healthy volunteers: Their use in the clinical development of psychotropic drugs. J. Psychopharmacol. 2007, 21, 272–282. [Google Scholar] [CrossRef]
- Saletu, B.; Anderer, P.; Saletu-Zyhlarz, G.M. EEG Topography and Tomography (LORETA) in the Classification and Evaluation of the Pharmacodynamics of Psychotropic Drugs. Clin. EEG Neurosci. 2006, 37, 66–80. [Google Scholar] [CrossRef]
- VVisser, S.A.G.; Wolters, F.L.C.; Gubbens-Stibbe, J.M.; Tukker, E.; van der Graaf, P.H.; Peletier, L.A.; Danhof, M. Mechanism-Based Pharmacokinetic/Pharmacodynamic Modeling of the Electroencephalogram Effects of GABAAReceptor Modulators: In Vitro-in Vivo Correlations. J. Pharmacol. Exp. Ther. 2003, 304, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Buchsbaum, M.S.; Hazlett, E.; Sicotte, N.; Stein, M.; Wu, J.; Zetin, M. Topographic EEG changes with benzodiazepine administration in generalized anxiety disorder. Biol. Psychiatry 1985, 20, 832–842. [Google Scholar] [CrossRef]
- Victorino, D.B.; Pinheiro, D.J.L.L.; Scott-McKean, J.J.; Barker, S.; Stasko, M.R.; Faber, J.; Scorza, C.A.; Costa, A.C.S. Atypical electrophysiological and behavioral responses to diazepam in a leading mouse model of Down syndrome. Sci. Rep. 2021, 11, 9521. [Google Scholar] [CrossRef]
- Nishida, M.; Zestos, M.M.; Asano, E. Spatial–temporal patterns of electrocorticographic spectral changes during midazolam sedation. Clin. Neurophysiol. 2016, 127, 1223–1232. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, K.A.A.L.; dos Santos, J.R.; Melo, T.C.D.S.; de Souza, M.F.; Santos, L.D.G.; de Gois, A.M.; Cintra, R.R.; Lins, L.C.R.; Ribeiro, A.M.; Marchioro, M. Depressant effect of geraniol on the central nervous system of rats: Behavior and ECoG power spectra. Biomed. J. 2018, 41, 298–305. [Google Scholar] [CrossRef]
- Cheaha, D.; Sawangjaroen, K.; Kumarnsit, E. Characterization of fluoxetine effects on ethanol withdrawal-induced cortical hyperexcitability by EEG spectral power in rats. Neuropharmacology 2014, 77, 49–56. [Google Scholar] [CrossRef]
- Dimpfel, W. Pharmacological modulation of cholinergic brain activity and its reflection in special EEG frequency ranges from various brain areas in the freely moving rat (Tele-Stereo-EEG). Eur. Neuropsychopharmacol. 2005, 15, 673–682. [Google Scholar] [CrossRef]
- Wilfried, D.; Nina, C.D.G.; Silvia, B. Effectiveness of Menosan® Salvia officinalis in the treatment of a wide spectrum of menopausal complaints. A double-blind, randomized, placebo-controlled, clinical trial. Heliyon 2021, 7, e05910. [Google Scholar] [CrossRef]
- Dimpfel, W. Pharmacological Modulation of Dopaminergic Brain Activity and Its Reflection in Spectral Frequencies of the Rat Electropharmacogram. Neuropsychobiology 2008, 58, 178–186. [Google Scholar] [CrossRef]
- Cadieux, R.J. Azapirones: An alternative to benzodiazepines for anxiety. Am. Fam. Physician 1996, 53, 2349–2353. [Google Scholar]
- Estrada-Camarena, E.; Sollozo-Dupont, I.; Islas-Preciado, D.; González-Trujano, M.E.; Carro-Juárez, M.; López-Rubalcava, C. Anxiolytic- and anxiogenic-like effects of Montanoa tomentosa (Asteraceae): Dependence on the endocrine condition. J. Ethnopharmacol. 2019, 241, 112006. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Carrera, D.; Ventura-Martinez, R.; Cedillo-Portugal, E.; Navarrete, A. Neuropharmacological profile of an ethanol extract of Ruta chalepensis L. in mice. J. Ethnopharmacol. 2006, 106, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.; Koester, A.G.; Pearson, D.W. Exploratory behavior in chronic disulfoton poisoning in mice. Psychopharmacology 1971, 20, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.G. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 1987, 92, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates, 5th ed.; Academic Press: London, UK, 2019; ISBN 978-0-12-816158-6. [Google Scholar]
- Valdés-Cruz, A.; Negrete-Díaz, J.V.; Magdaleno-Madrigal, V.M.; Martínez-Vargas, D.; Fernández-Mas, R.; Almazán-Alvarado, S.; Torres-García, M.E.; Flores, G. Electroencephalographic activity in neonatal ventral hippocampus lesion in adult rats. Synapse 2012, 66, 738–746. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.-X.; Cao, Y. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Schüssler, P.; Kluge, M.; Yassouridis, A.; Dresler, M.; Held, K.; Zihl, J.; Steiger, A. Progesterone reduces wakefulness in sleep EEG and has no effect on cognition in healthy postmenopausal women. Psychoneuroendocrinology 2008, 33, 1124–1131. [Google Scholar] [CrossRef]
- Rihs, T.A.; Michel, C.M.; Thut, G. Mechanisms of selective inhibition in visual spatial attention are indexed by-band EEG synchronization. Eur. J. Neurosci. 2007, 25, 603–610. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onofre-Campos, D.; González-Trujano, M.E.; Moreno-Pérez, G.F.; Narváez-González, F.; González-Gómez, J.D.; Villasana-Salazar, B.; Martínez-Vargas, D. Anxiolytic-like Effects and Quantitative EEG Profile of Palmitone Induces Responses Like Buspirone Rather Than Diazepam as Clinical Drugs. Molecules 2023, 28, 3680. https://doi.org/10.3390/molecules28093680
Onofre-Campos D, González-Trujano ME, Moreno-Pérez GF, Narváez-González F, González-Gómez JD, Villasana-Salazar B, Martínez-Vargas D. Anxiolytic-like Effects and Quantitative EEG Profile of Palmitone Induces Responses Like Buspirone Rather Than Diazepam as Clinical Drugs. Molecules. 2023; 28(9):3680. https://doi.org/10.3390/molecules28093680
Chicago/Turabian StyleOnofre-Campos, Daniela, María Eva González-Trujano, Gabriel Fernando Moreno-Pérez, Fernando Narváez-González, José David González-Gómez, Benjamín Villasana-Salazar, and David Martínez-Vargas. 2023. "Anxiolytic-like Effects and Quantitative EEG Profile of Palmitone Induces Responses Like Buspirone Rather Than Diazepam as Clinical Drugs" Molecules 28, no. 9: 3680. https://doi.org/10.3390/molecules28093680