
Bioactivity Guided Study for the Isolation and Identification of Antidiabetic Compounds from Edible Seaweed—Ulva reticulata


Abstract
:1. Introduction
2. Results
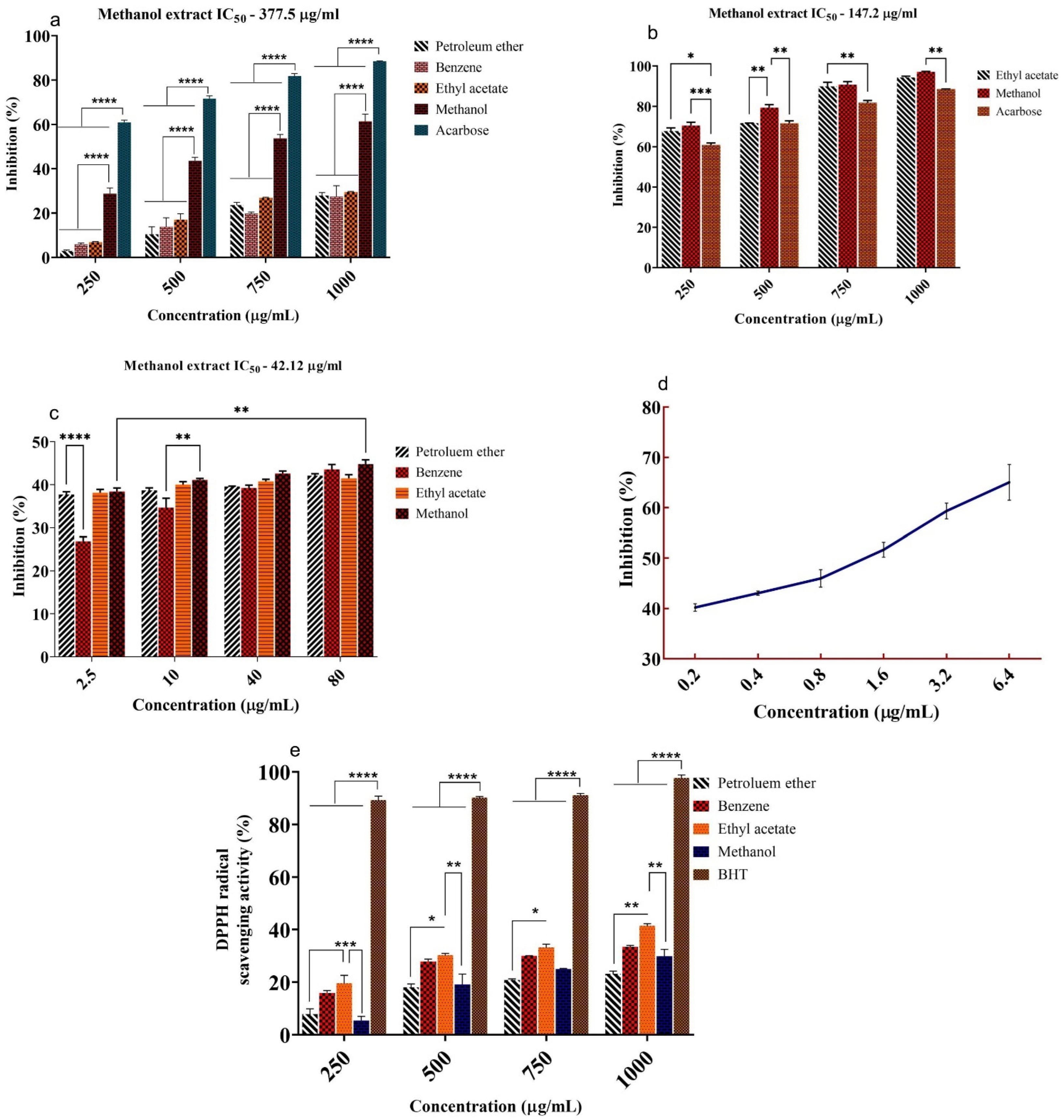
2.1. In Vitro α-Amylase, α-Glucosidase, and DPP-IV Inhibition Study
2.2. Free Radical Scavenging Activity (DPPH)
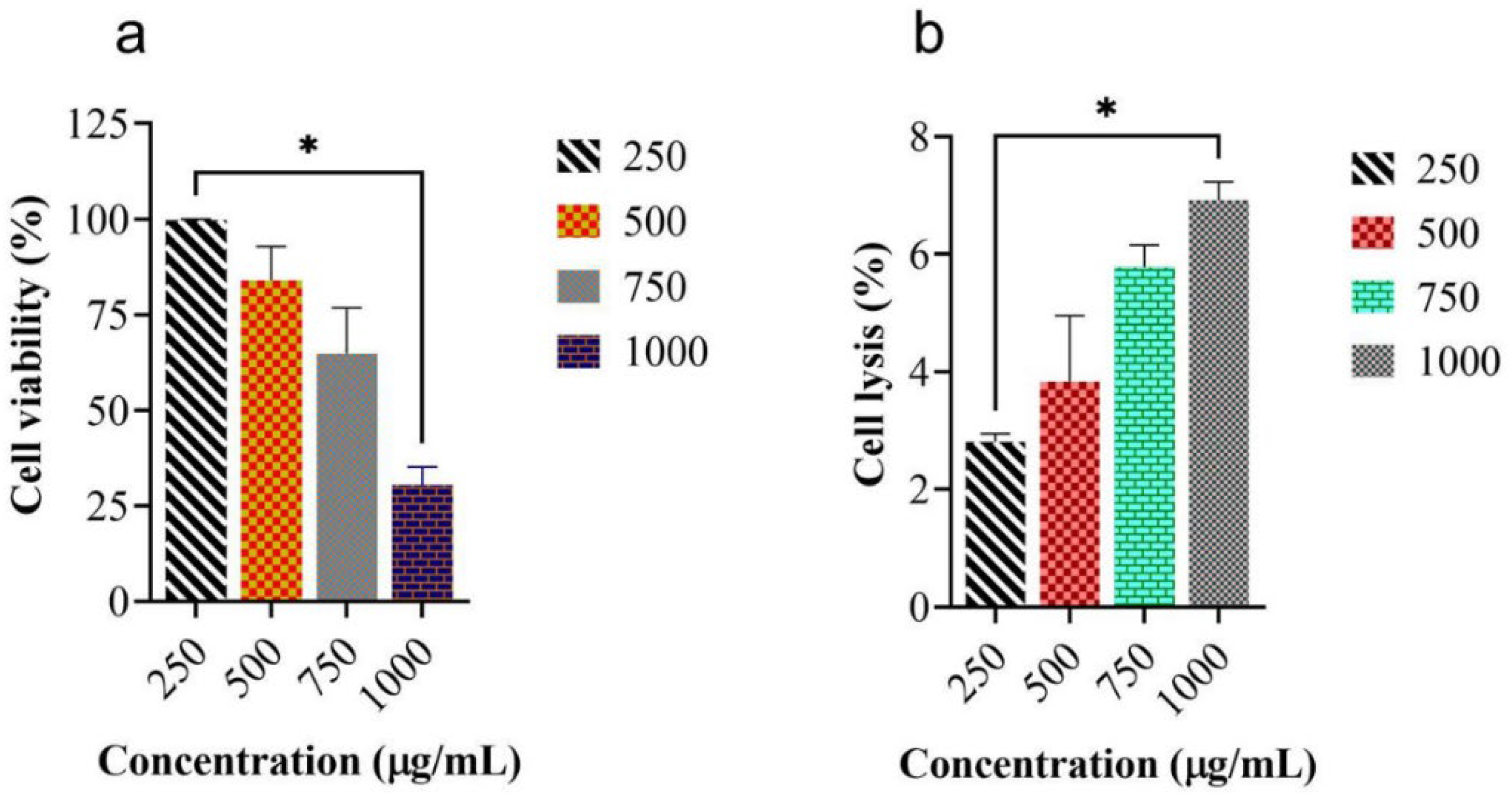
2.3. Measurement of Cell Viability and In Vitro Hemolytic Activity of MEUR
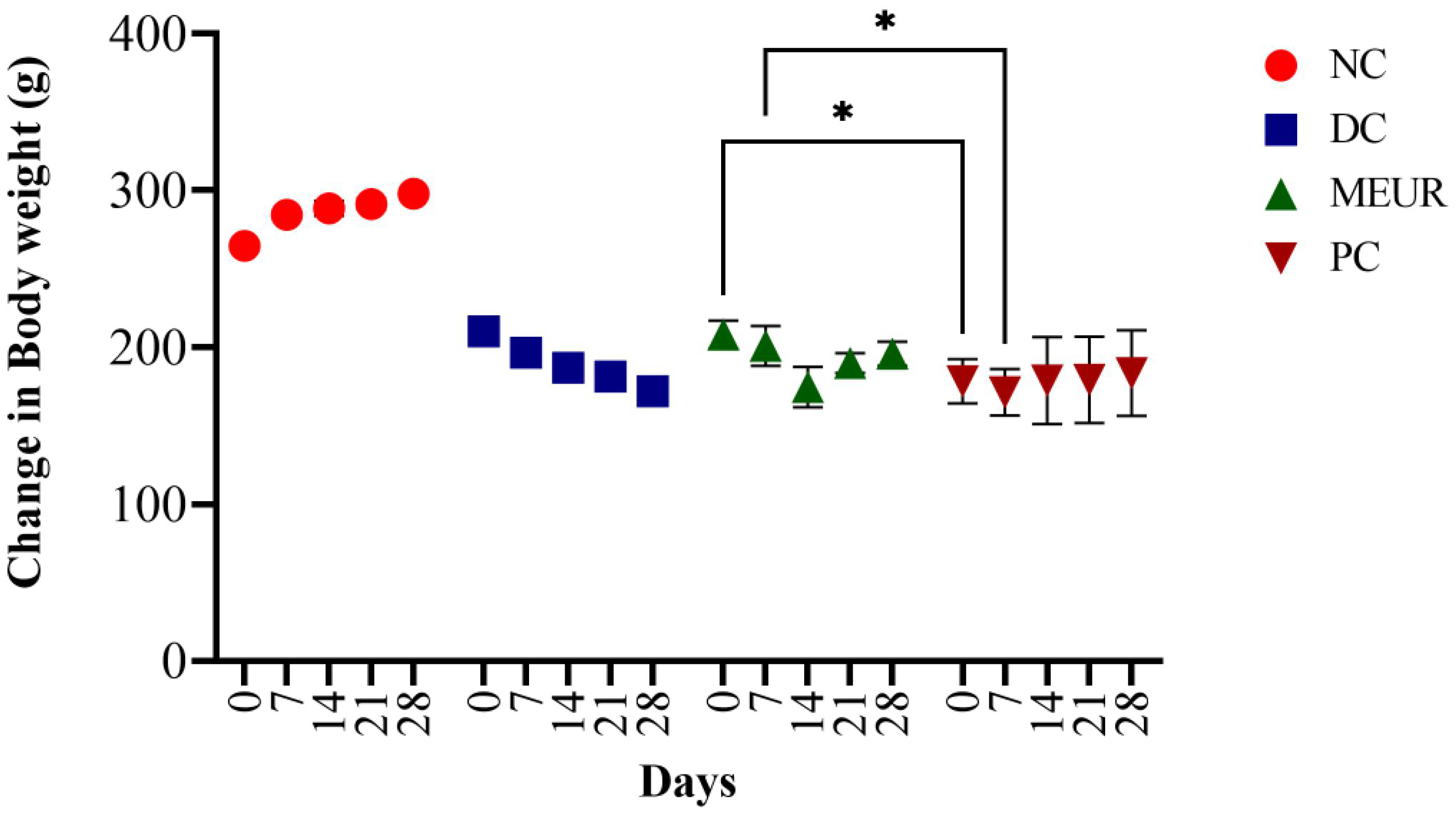
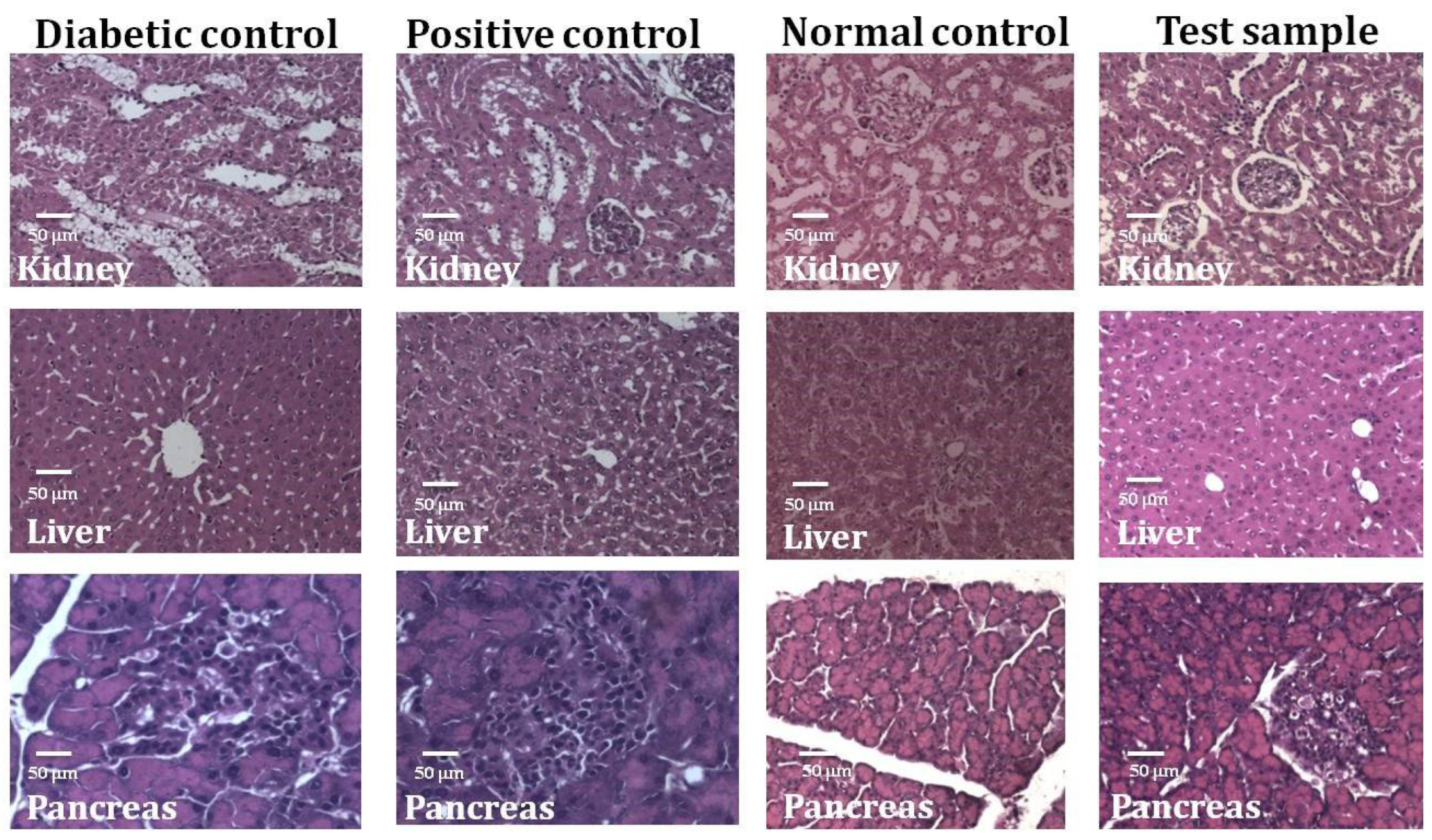
2.4. Effect of MEUR on STZ-Induced Diabetic Rats
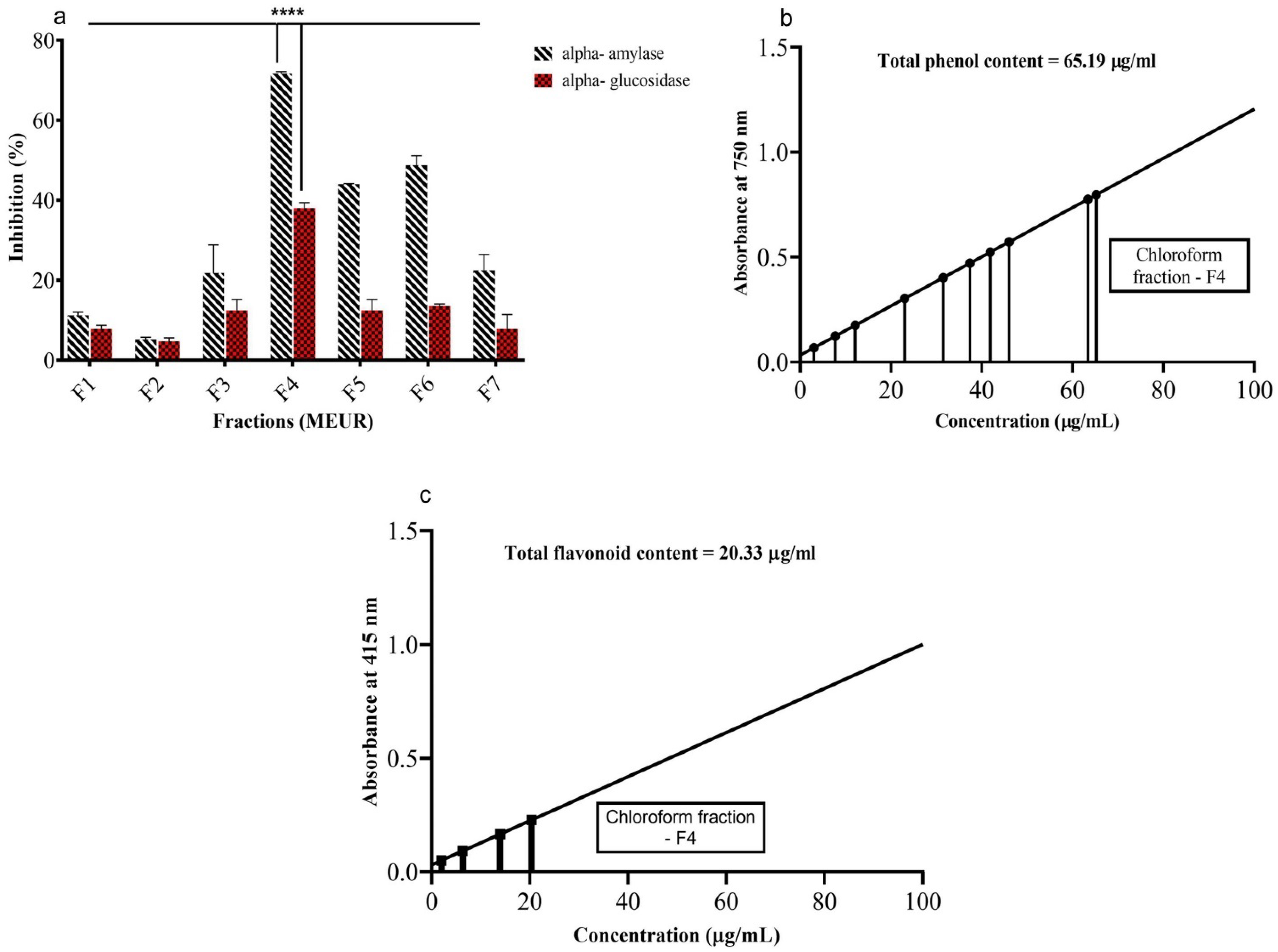
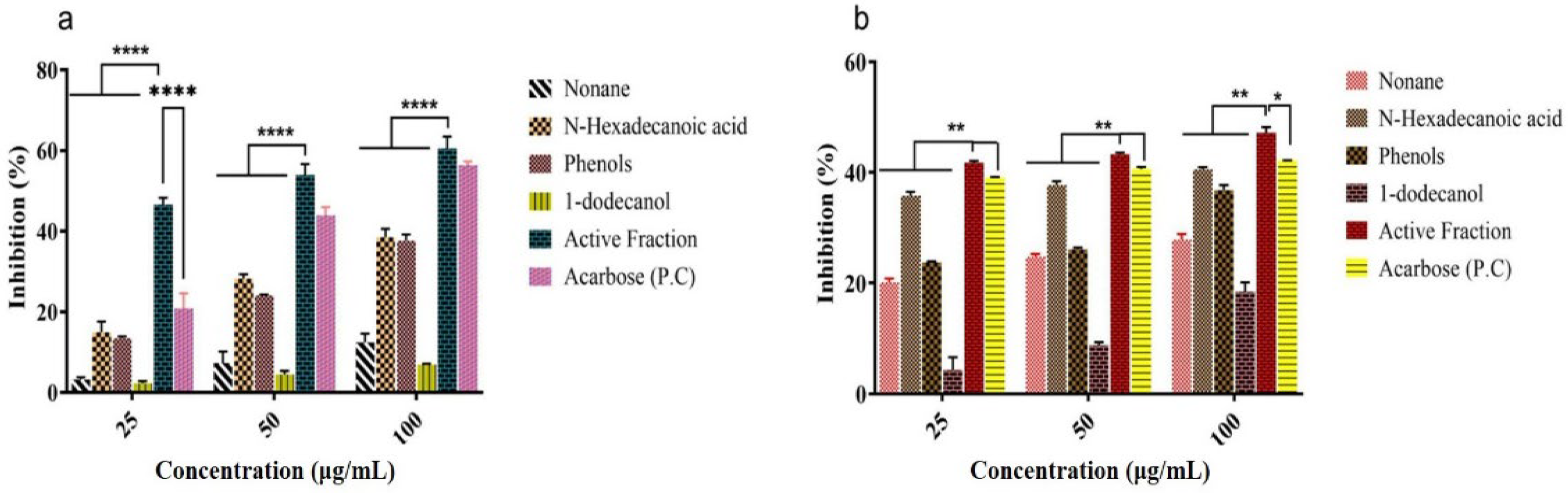
2.5. In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Isolated Fractions from MEUR
2.6. Determination of Total Phenols and Flavonoids
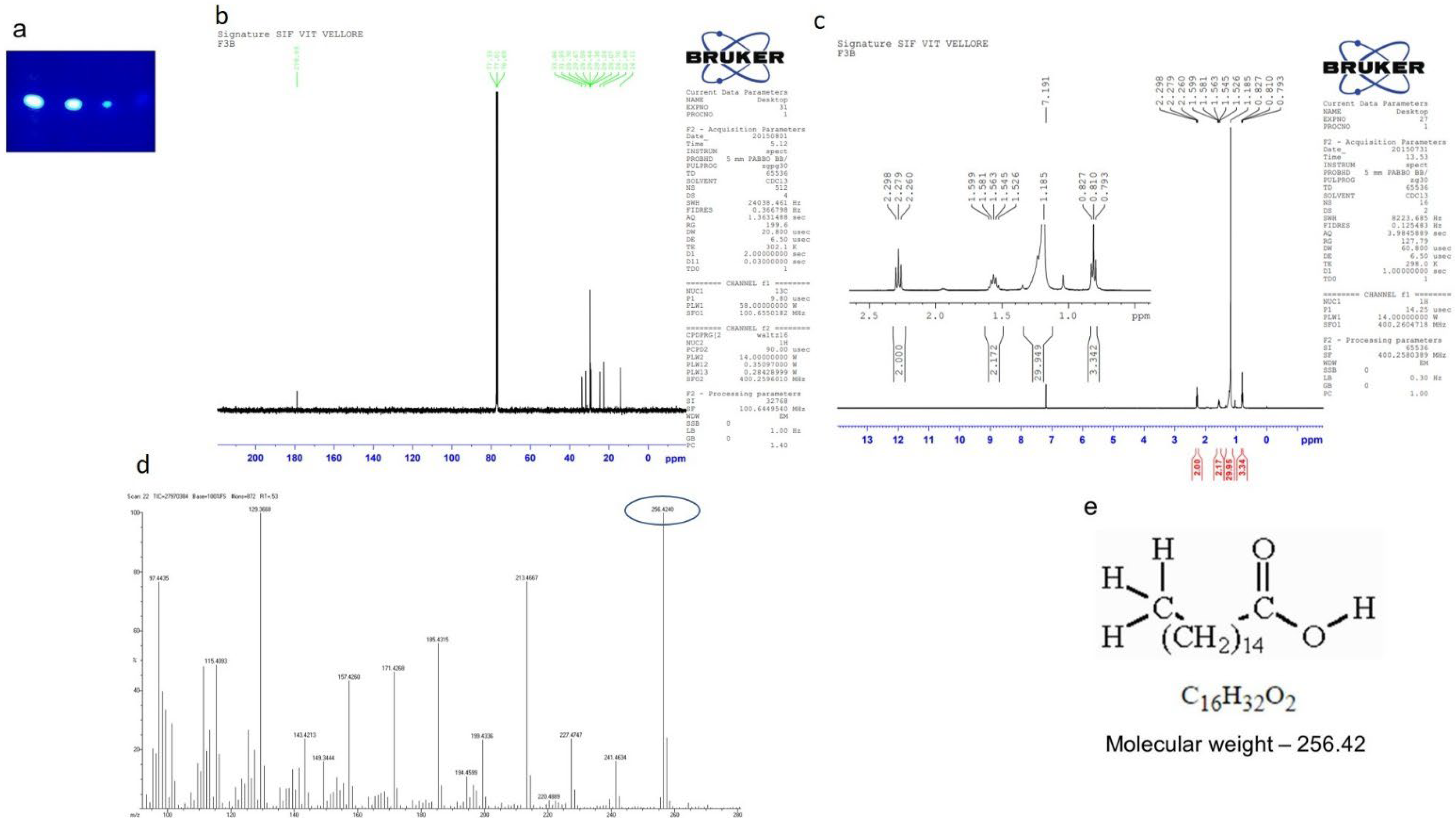
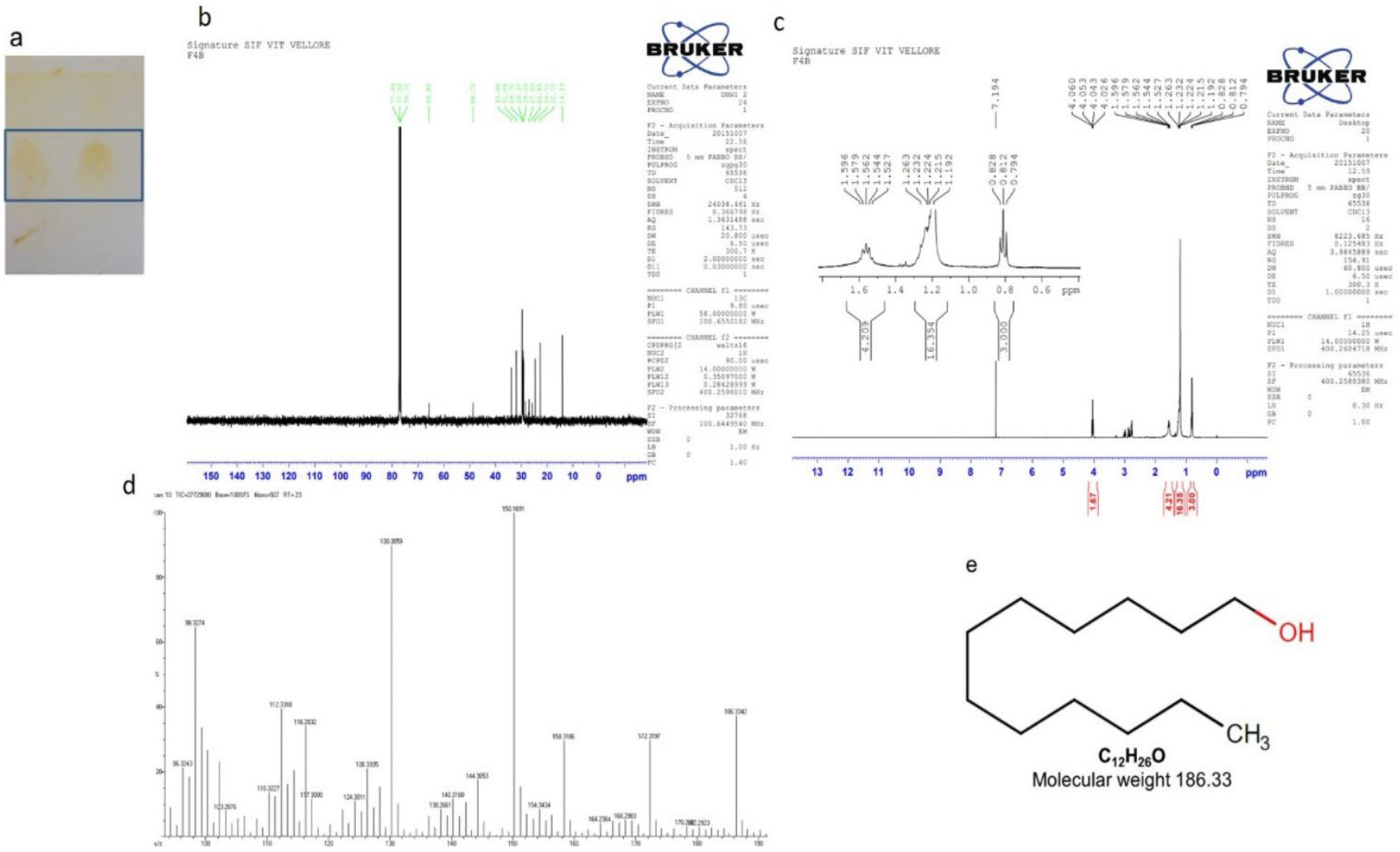
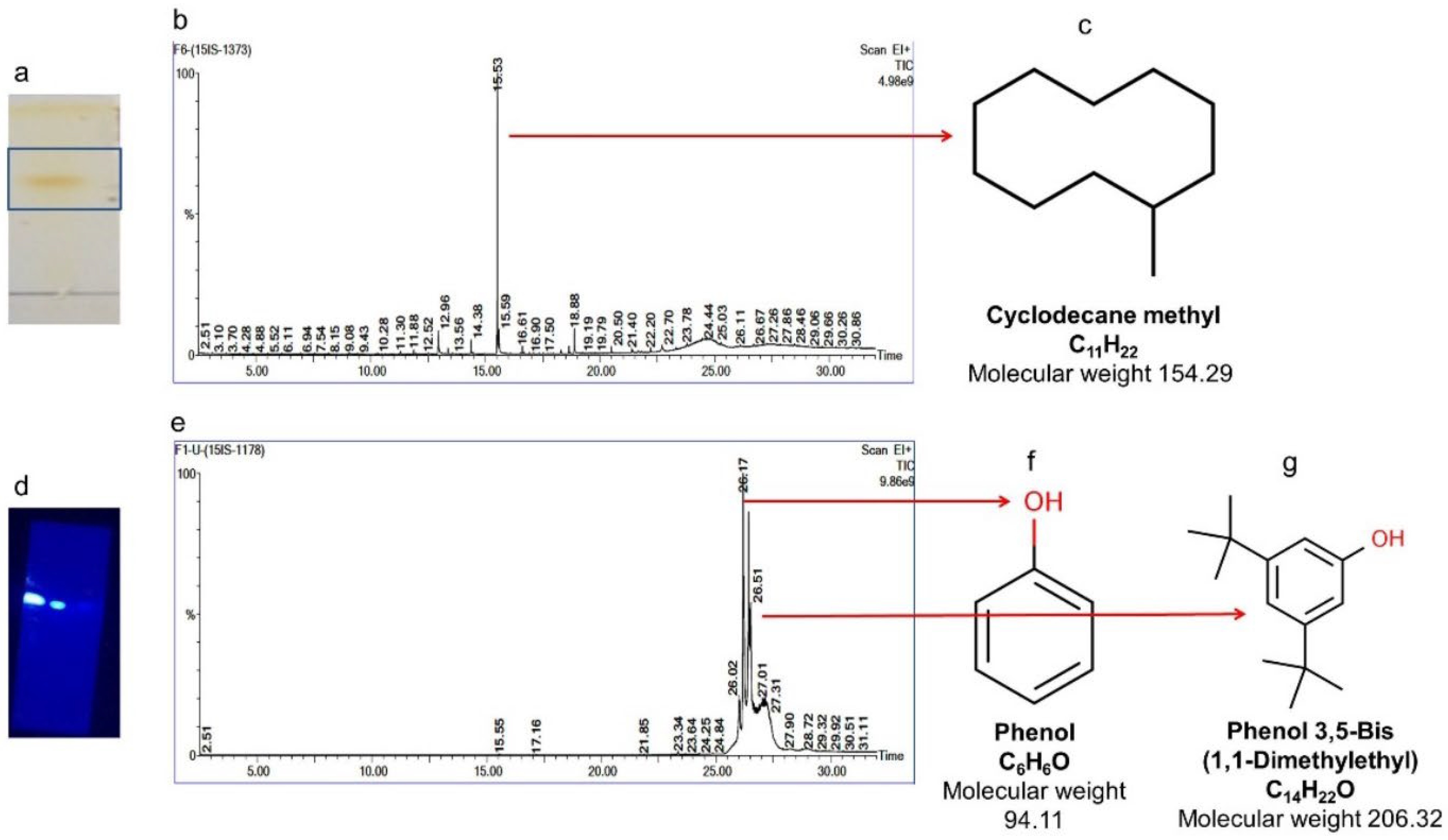
2.7. Purification of Compounds from the Active Fraction (F4)
2.7.1. Isolation of Compound 1 (Nonane)
2.7.2. Isolation of Compound 2 (Hexadecanoic Acid)
2.7.3. Isolation of Compound 3 (1-dodecanol)
2.7.4. Isolation of Compounds 4 and 5 (Cyclodecane Methyl and Phenol, Phenol, 3,5-bis(1,1-dimethylethyl))
2.7.5. In Vitro α-Amylase and α-Glucosidase Inhibition Study of Isolated Compounds and Its Active Fraction (F4)
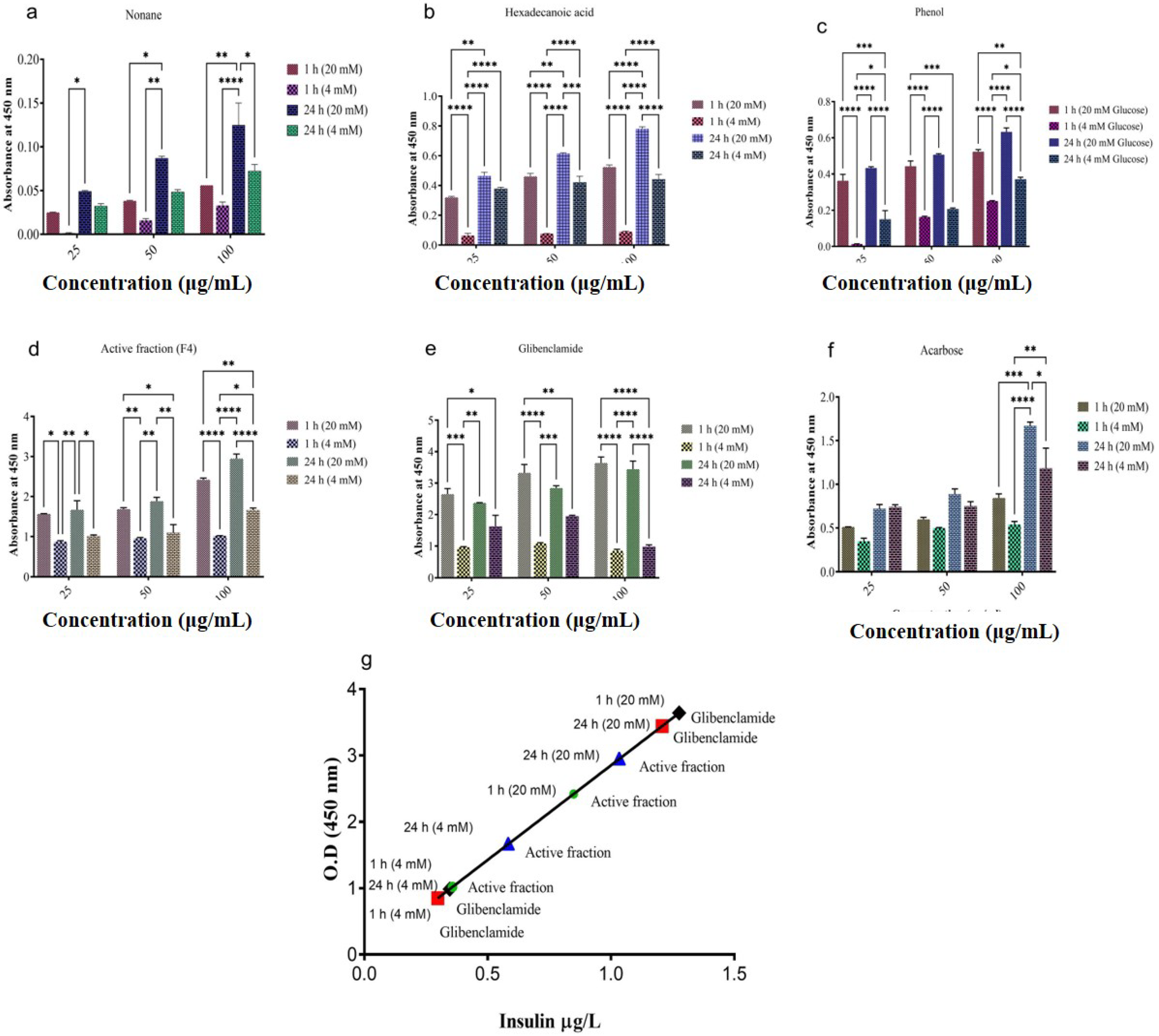
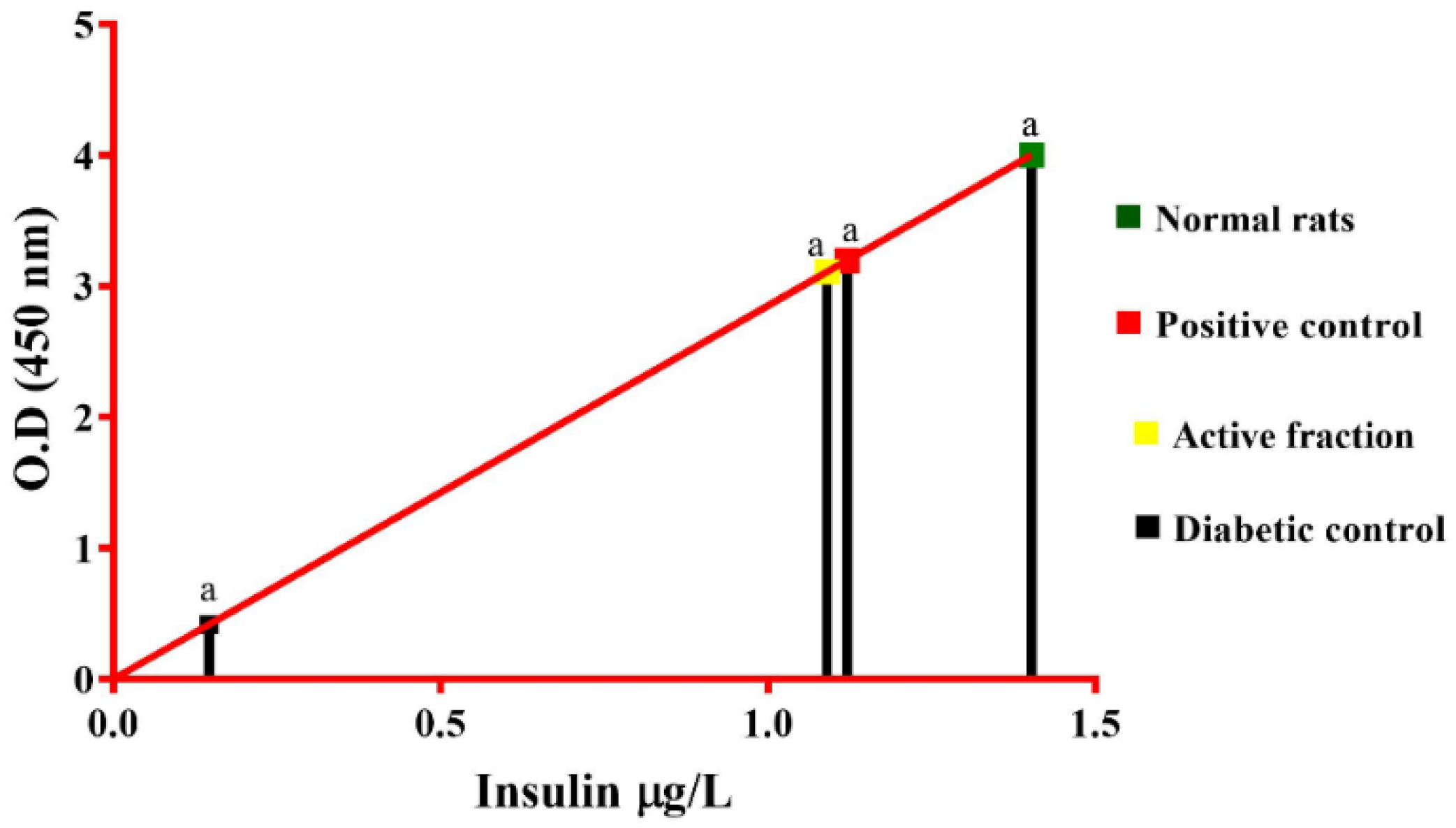
2.8. In Vitro Insulin Secretion Studies
2.9. In Vivo Antidiabetic Mechanism of Active Fraction (F4)
3. Discussion
4. Materials and Methods
4.1. Collection of Seaweeds
4.2. Preparation of Seaweed Extracts
4.3. In Vitro α-Amylase and α-Glucosidase Inhibition Study
4.4. In Vitro Dipeptidyl Peptidase-IV (DPP-IV) Inhibition Study
4.5. Free Radical Scavenging Activity (DPPH)
4.6. Maintenance of Cell Line
4.7. Cytotoxicity Assay
4.8. In Vitro Hemolytic Activity
4.9. Experimental Animal
4.10. Experimental Design
- (1)
- Normal control: Rats fed with normal food and water
- (2)
- Diabetic control: Rats were made diabetic by a single intraperitoneal injection of streptozotocin (STZ) at a concentration of 45 mg kg−1 body weight.
- (3)
- Diabetic rats treated with glibenclamide (Positive control): Rats were made diabetic by STZ (45 mg kg−1 body weight) and treated orally with standard antidiabetic drug glibenclamide (0.25 mg kg−1 body weight) once daily for 28 days.
- (4)
- Diabetic rats treated with methanolic extract of U. reticulata (MEUR): Rats were made diabetic by STZ (45 mg kg−1 body weight) and treated orally with methanolic extract of U. reticulata (250 mg kg−1 body weight) once daily for 28 days.
4.11. Bioassay-Guided Fractionation and Isolation of Active Compounds from MEUR
4.12. Determination of Total Phenols and Flavonoids
4.13. In Vitro α-Amylase and α-Glucosidase Inhibition Study of Isolated Compounds and Their Active Fraction (F4)
4.14. In Vitro Insulin Secretion Studies Using Isolated Pancreatic Islets
4.15. In Vivo Antidiabetic Mechanism of Active Fraction (F4)
4.15.1. Effect in Normoglycemic Animals
4.15.2. Effect in Glucose-Loaded Hyperglycemic Animals
4.15.3. Effect in STZ-Induced Diabetic Animals
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| MEUR | Methanolic extract of Ulva reticulata |
| S.T.Z. | streptozotocin |
| PPHG | Postprandial hyperglycemia |
| B.wt | Body weight |
| FBG | Fasting Blood Glucose levels |
| ALT | Alanine transaminase |
| AST | Aspartate transaminase |
References
- Marcovecchio, M.; Mohn, A.; Chiarelli, F. Type 2 Diabetes Mellitus in Children and Adolescents. J. Endocrinol. Investig. 2005, 28, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Sun, J.; Shan, W.; Xiao, W.; Wang, B.; Ma, X.; Hu, W.; Wang, X.; Xia, Y. Environmental Chemical Exposure Dynamics and Machine Learning-Based Prediction of Diabetes Mellitus. Sci. Total Environ. 2022, 806, 150674. [Google Scholar] [CrossRef] [PubMed]
- The American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2009, 32, S62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.H.; Ju, J.W.; Park, M.J.; Han, J.S. Daidzein Inhibits Carbohydrate Digestive Enzymes in Vitro and Alleviates Postprandial Hyperglycemia in Diabetic Mice. Eur. J. Pharmacol. 2013, 712, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.R.; Standl, E.; Tong, N.; Shah, P.; Kalra, S.; Rathod, R. Therapeutic Potential of α-Glucosidase Inhibitors in Type 2 Diabetes Mellitus: An Evidence-Based Review. Expert Opin. Pharmacother. 2015, 16, 1959–1981. [Google Scholar] [CrossRef]
- Akmal, M.; Wadhwa, R. Alpha Glucosidase Inhibitors—StatPearls—NCBI Bookshelf. Available online: https://www.ncbi.nlm.nih.gov/books/NBK557848/#_NBK557848_pubdet_ (accessed on 3 August 2022).
- Kang, S.M.; Park, J.H. Pleiotropic Benefits of DPP-4 Inhibitors Beyond Glycemic Control. Clin. Med. Insights Endocrinol. Diabetes 2021, 14. [Google Scholar] [CrossRef]
- Fehmann, H.C.; Habener, J.F. Insulinotropic Hormone Glucagon-like Peptide-i (7-37) Stimulation of Proinsulin Gene Expression and Proinsulin Biosynthesis in Insulinoma Βtc-1 Cells. Endocrinology 1992, 130, 159–166. [Google Scholar] [CrossRef]
- Ganesan, K.; Sultan, S. Oral Hypoglycemic Medications; StatPearls—NCBI Bookshelf. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482386/ (accessed on 30 October 2022).
- Ling, F.; Guo, D.; Zhu, L. Pneumatosis Cystoides Intestinalis: A Case Report and Literature Review. BMC Gastroenterol. 2019, 19, 176. [Google Scholar] [CrossRef]
- Monami, M.; Iacomelli, I.; Marchionni, N.; Mannucci, E. Dipeptydil Peptidase-4 Inhibitors in Type 2 Diabetes: A Meta-Analysis of Randomized Clinical Trials. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 224–235. [Google Scholar] [CrossRef]
- Rajasekaran, P.; Ande, C.; Vankar, Y.D. Synthesis of (5,6 & 6,6)-Oxa-Oxa Annulated Sugars as Glycosidase Inhibitors from 2-Formyl Galactal Using Iodocyclization as a Key Step. Arkivoc 2022, 2022, 5–23. [Google Scholar] [CrossRef]
- Nash, R.J.; Kato, A.; Yu, C.-Y.; Fleet, G.W. Iminosugars as Therapeutic Agents: Recent Advances and Promising Trends. Future Med. Chem. 2011, 3, 1513–1521. [Google Scholar] [CrossRef]
- Tyagi, T.; Agarwal, M. GC-MS ANALYSIS OF INVASIVE AQUATIC WEED, PISTIA STRATIOTES L. AND EICHHORNIA CRASSIPES (MART.) SOLMS. Int. J. Curr. Pharm. Res. 2017, 9, 111. [Google Scholar] [CrossRef]
- Veluchamy, C.; Palaniswamy, R. A Review on Marine Algae and Its Applications. Asian J. Pharm. Clin. Res. 2020, 13, 21–27. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, S.M.; Sok, C.H.; Hong, J.T.; Oh, J.Y.; Jeon, Y.J. Cellular Activities and Docking Studies of Eckol Isolated from Ecklonia Cava (Laminariales, Phaeophyceae) as Potential Tyrosinase Inhibitor. Algae 2015, 30, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.C.; Jeon, Y.J. Anti-Inflammatory Effect of Enzymatic Hydrolysates from Styela Clava Flesh Tissue in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages and in Vivo Zebrafish Model. Nutr. Res. Pract. 2015, 9, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, K.W.; Elvitigala, D.A.S.; Lakmal, H.H.C.; Kim, Y.M.; Jeon, Y.J. Future Prospects and Health Benefits of Functional Ingredients from Marine Bio-Resources: A Review. Fish. Aquat. Sci. 2014, 17, 275–290. [Google Scholar] [CrossRef]
- El Shafay, S.; El-Sheekh, M.; Bases, E.; El-Shenody, R. Antioxidant, Antidiabetic, Anti-Inflammatory and Anticancer Potential of Some Seaweed Extracts. Food Sci. Technol. 2022, 42. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of Antioxidant Potential of Seaweed Extracts for Enrichment of Convenience Food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Namvar, F.; Mohamed, S.; Fard, S.G.; Behravan, J.; Mustapha, N.M.; Alitheen, N.B.M.; Othman, F. Polyphenol-Rich Seaweed (Eucheuma Cottonii) Extract Suppresses Breast Tumour via Hormone Modulation and Apoptosis Induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- Iwai, K. Antidiabetic and Antioxidant Effects of Polyphenols in Brown Alga Ecklonia Stolonifera in Genetically Diabetic KK-Ay Mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef]
- Khan, M.N.A.; Cho, J.Y.; Lee, M.C.; Kang, J.Y.; Nam, G.P.; Fujii, H.; Hong, Y.K. Isolation of Two Anti-Inflammatory and One pro-Inflammatory Polyunsaturated Fatty Acids from the Brown Seaweed Undaria Pinnatifida. J. Agric. Food Chem. 2007, 55, 6984–6988. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, P.S.; Jayasri, M.A. Antidiabetic Studies of Chaetomorpha Antennina Extract Using Experimental Models. J. Appl. Phycol. 2017, 29, 1047–1056. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Inhibitory Potential of Turbinaria Ornata against Key Metabolic Enzymes Linked to Diabetes. BioMed Res. Int. 2014, 2014, 783895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wu, W.; Ni, X.; Farag, M.A.; Capanoglu, E.; Zhao, C. Regulatory Mechanisms of the Green Alga Ulva Lactuca Oligosaccharide via the Metabolomics and Gut Microbiome in Diabetic Mice. Curr. Res. Food Sci. 2022, 5, 1127–1139. [Google Scholar] [CrossRef]
- Ratana-arporn, P.; Chirapart, A. Nutritional Evaluation of Tropical Green Seaweeds Caulerpa Lentillifera and Ulva Reticulata. Kasetsart J. Nat. Sci. 2006, 40, 75–83. [Google Scholar]
- Anh, H.T.L.; Kawata, Y.; Tam, L.T.; Thom, L.T.; Ha, N.C.; Hien, H.T.M.; Thu, N.T.H.; Huy, P.Q.; Hong, D.D. Production of Pyruvate from Ulva Reticulata Using the Alkaliphilic, Halophilic Bacterium Halomonas Sp. BL6. J. Appl. Phycol. 2020, 32, 2283–2293. [Google Scholar] [CrossRef]
- Thahira Banu, A.; Uma Mageswari, S. Nutritional Status and Effect of Seaweed Chocolate on Anemic Adolescent Girls. Food Sci. Hum. Wellness 2015, 4, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Chee, S.Y.; Wong, P.K.; Wong, C.L. Extraction and Characterisation of Alginate from Brown Seaweeds (Fucales, Phaeophyceae) Collected from Port Dickson, Peninsular Malaysia. J. Appl. Phycol. 2011, 23, 191–196. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M. Natural Products, a Potential Source of New Drugs Discovery to Combat Obesity and Diabetes: Their Efficacy and Multi-Targets Actions in Treatment of These Diseases. In Natural Products in Obesity and Diabetes; Springer: Cham, Switzerland, 2022; pp. 101–275. [Google Scholar]
- Dinda, B.; Dinda, S.; Chakraborty, M. Pharmacology of Anti-Obesity and Antidiabetic Phytochemicals Isolated from Various Natural Sources (Plants, Seaweeds, Mushrooms, Marine Animals, and Microorganisms). In Natural Products in Obesity and Diabetes; Springer: Cham, Switzerland, 2022; pp. 277–467. [Google Scholar]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Antidiabetic Potential of Marine Algae by Inhibiting Key Metabolic Enzymes. Front. Life Sci. 2015, 8, 148–159. [Google Scholar] [CrossRef]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant Activity of Extracts from Marine Macroalgae, Wild-Collected and Cultivated, in an Integrated Multi-Trophic Aquaculture System. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; El-Shenody, R.A.E.K.; Bases, E.A.; El Shafay, S.M. Comparative Assessment of Antioxidant Activity and Biochemical Composition of Four Seaweeds, Rocky Bay of Abu Qir in Alexandria, Egypt. Food Sci. Technol. 2021, 41, 29–40. [Google Scholar] [CrossRef]
- Al-Araby, S.Q.; Rahman, M.A.; Chowdhury, M.A.H.; Das, R.R.; Chowdhury, T.A.; Hasan, C.M.M.; Afroze, M.; Hashem, M.A.; Hajjar, D.; Alelwani, W.; et al. Padina Tenuis (Marine Alga) Attenuates Oxidative Stress and Streptozotocin-Induced Type 2 Diabetic Indices in Wistar Albino Rats. S. Afr. J. Bot. 2020, 128, 87–100. [Google Scholar] [CrossRef]
- Okechukwu, P.; Sharma, M.; Tan, W.H.; Chan, H.K.; Chirara, K.; Gaurav, A.; Al-Nema, M. In-Vitro Anti-Diabetic Activity and in-Silico Studies of Binding Energies of Palmatine with Alpha-Amylase, Alpha-Glucosidase and DPP-IV Enzymes. Pharmacia 2020, 67, 363–371. [Google Scholar] [CrossRef]
- Lankatillake, C.; Huynh, T.; Dias, D.A. Understanding Glycaemic Control and Current Approaches for Screening Antidiabetic Natural Products from Evidence-Based Medicinal Plants. Plant Methods 2019, 15, 105. [Google Scholar] [CrossRef] [Green Version]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic Potential of Marine Brown Algae—A Mini Review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef] [Green Version]
- Unnikrishnan, P.; Jayasri, M.; Suthindhiran, K.; Jayasri, M. Alpha-Amylase Inhibition and Antioxidant Activity of Marine Green Algae and Its Possible Role in Diabetes Management. Pharmacogn. Mag. 2015, 11, 511. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Nguyen, T.V.L.; Nguyen, Q.D.; Nguyen, N.N.; Nguyen, T.T.D. Comparison of Phytochemical Contents, Antioxidant and Antibacterial Activities of Various Solvent Extracts Obtained from ‘Maluma’ Avocado Pulp Powder. Molecules 2021, 26, 7693. [Google Scholar] [CrossRef]
- Stoddart, M.J. Cell Viability Assays: Introduction. Methods Mol. Biol. 2011, 740, 1–6. [Google Scholar]
- Garg, S.; Huifu, H.; Kaul, S.C.; Wadhwa, R. Integration of Conventional Cell Viability Assays for Reliable and Reproducible Read-Outs: Experimental Evidence. BMC Res. Notes 2018, 11, 403. [Google Scholar] [CrossRef]
- Kottaisamy, C.P.D.; Raj, D.S.; Prasanth Kumar, V.; Sankaran, U. Experimental Animal Models for Diabetes and Its Related Complications—A Review. Lab. Anim. Res. 2021, 37, 23. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.R. Diabetic Dyslipidemia. Atheroscler. Suppl. 2002, 3, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-Glucosidase and α-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding Phenolic Acids Inhibition of α-Amylase and α-Glucosidase and Influence of Reaction Conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, C.; Zhang, B.; Huang, Q. The Inhibitory Effects of Flavonoids on α-Amylase and α-Glucosidase. Crit. Rev. Food Sci. Nutr. 2020, 60, 695–708. [Google Scholar] [CrossRef]
- Akinyede, K.A.; Oyewusi, H.A.; Hughes, G.D.; Ekpo, O.E.; Oguntibeju, O.O. In Vitro Evaluation of the Anti-Diabetic Potential of Aqueous Acetone Helichrysum Petiolare Extract (AAHPE) with Molecular Docking Relevance in Diabetes Mellitus. Molecules 2022, 27, 155. [Google Scholar] [CrossRef]
- Hemmingsen, B.; Sonne, D.P.; Metzendorf, M.; Richter, B. Insulin Secretagogues for Prevention or Delay of Type 2 Diabetes Mellitus and Its Associated Complications in Persons at Increased Risk for the Development of Type 2 Diabetes Mellitus. Cochrane Database Syst. Rev. 2016, 2016, CD012151. [Google Scholar] [CrossRef]
- Skelin, M.; Rupnik, M.; Cencic, A. Pancreatic Beta Cell Lines and Their Applications in Diabetes Mellitus Research. ALTEX 2010, 27, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Al-Masri, I.M.; Mohammad, M.K.; Tahaa, M.O. Inhibition of Dipeptidyl Peptidase IV (DPP IV) Is One of the Mechanisms Explaining the Hypoglycemic Effect of Berberine. J. Enzyme Inhib. Med. Chem. 2009, 24, 1061–1066. [Google Scholar] [CrossRef]
- Mensor, L.L.; Menezes, F.S.; Leitão, G.G.; Reis, A.S.; dos Santos, T.C.; Coube, C.S.; Leitão, S.G. Screening of Brazilian Plant Extracts for Antioxidant Activity by the Use of DPPH Free Radical Method. Phyther. Res. 2001, 15, 127–130. [Google Scholar] [CrossRef]
- Malagoli, D. A Full-Length Protocol to Test Hemolytic Activity of Palytoxin on Human Erythrocytes. Invertebr. Surviv. J. 2007, 4, 92–94. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the Total Phenolic, Flavonoid and Proline Contents in Burkina Fasan Honey, as Well as Their Radical Scavenging Activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colometric Methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Nampoothiri, S.V.; Prathapan, A.; Cherian, O.L.; Raghu, K.G.; Venugopalan, V.V.; Sundaresan, A. In Vitro Antioxidant and Inhibitory Potential of Terminalia Bellerica and Emblica Officinalis Fruits against LDL Oxidation and Key Enzymes Linked to Type 2 Diabetes. Food Chem. Toxicol. 2011, 49, 125–131. [Google Scholar] [CrossRef]
- Howell, S.L.; Taylor, K.W. Effects of Glucose Concentration on Incorporation of [3H]Leucine into Insulin Using Isolated Mammalian Islets of Langerhans. BBA—Gen. Subj. 1966, 130, 519–521. [Google Scholar] [CrossRef]
- Ansari, P.; Tabasumma, N.; Snigdha, N.N.; Siam, N.H.; Panduru, R.V.N.R.S.; Azam, S.; Hannan, J.M.A.; Abdel-Wahab, Y.H.A. Diabetic Retinopathy: An Overview on Mechanisms, Pathophysiology and Pharmacotherapy. Diabetology 2022, 3, 159–175. [Google Scholar] [CrossRef]
- Animaw, W.; Seyoum, Y. Increasing Prevalence of Diabetes Mellitus in a Developing Country and Its Related Factors. PLoS ONE 2017, 12, e0187670. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Jayasri, M.A. Marine Algae As A Prospective Source For Antidiabetic Compounds—A Brief Review. Curr. Diabetes Rev. 2018, 14, 237–245. [Google Scholar] [CrossRef]
- Li, X.; Yu, Z.; Long, S.; Guo, Y.; Duan, D. Hypoglycemic Effect of Laminaria Japonica Polysaccharide in a Type 2 Diabetes Mellitus Mouse Model. ISRN Endocrinol. 2012, 2012, 507462. [Google Scholar] [CrossRef]
- Rajakani, M.; Muthulingam, M.; Karuppasamy, R. Padina Gymnospora (Kutzing) on Streptozotocin Induced Diabetics in Male Albino Wistar Rats. Int. J. Pharm. Biol. Sci. 2018, 8, 845–850. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (n = 6) | Experimental Group | Fasting Blood Glucose Level (mg/dL) | ||||
|---|---|---|---|---|---|---|
| 0th Day | 7th Day | 14th Day | 21st Day | 28th Day | ||
| I | Normal control | 96 ± 3.7 | 97 ± 4.06 | 74 ± 1.78 | 83 ± 3.14 | 85 ± 4.69 |
| II | Diabetic control | 389 ± 20.08 | 359 ± 25.84 | 345 ± 12.7 | 334 ± 21.72 | 329 ± 14.75 |
| III | Diabetic + glibenclamide (0.25 mg/kg) | 349 ± 57.37 | 267 ± 75.32 | 197 ± 28.2 | 131 ± 24.59 | 87 ± 4.83 |
| IV | Diabetic + MEUR (MEUR extract, 250 mg/kg) | 450 ± 12.01 | 430 ± 23.34 | 333 ± 20.81 | 242 ± 31.09 | 177 ± 27.3 |
| Group (n = 6) | Experimental Group | Serum Parameters | ||||||
|---|---|---|---|---|---|---|---|---|
| Total Protein (g/dL) | Triglycerides (mg/dL) | Urea (mg/dL) | ALT (IU/L) | AST (IU/L) | Cholesterol (mg/dL) | Albumin (g/dL) | ||
| I | Normal control | 6.8 ± 0.67 | 102.4 ± 3.86 | 8.22 ± 0.2 | 6.75 ± 0.83 | 13.43 ± 1.68 | 54.45 ± 2.24 | 3.17 ± 0.04 |
| II | Diabetic control | 7.04 ± 1.2 | 221 ± 72.54 | 17.9 ± 1.8 | 25.04 ± 3.37 | 36.42 ± 6.01 | 82.87 ± 8.5 | 2.91 ± 0.04 |
| III | Diabetic + Glibenclamide (0.25 mg/kg) | 7.15 ± 1.16 | 128.91 ± 5.64 | 16.43 ± 2.86 | 17.38 ± 2.39 | 14.84 ± 2.73 | 59.8 ± 8.12 | 2.82 ± 0.1 |
| IV | Diabetic + MEUR (MEUR extract, 250 mg/kg) | 6.14 ± 0.57 | 120 ± 27.79 | 16.97 ± 1.28 | 13.55 ± 1.17 | 10.01 ± 2.56 | 65.69 ± 2.24 | 2.38 ± 0.17 |
| Parameters | Time Point | Normal Control | Test (Active Fraction) | Acarbose | Glibenclamide |
|---|---|---|---|---|---|
| FBG (mg/dL) | 30 min | 71 ± 5.46 | 81 ± 5.57 | 77 ± 2.6 | 71 ± 1.85 |
| 60 min | 72 ± 3.53 | 64 ± 0.9 | 68 ± 3.05 | 66 ± 1.73 | |
| 120 min | 67 ± 1.45 | 58 ± 1.52 | 66 ± 1.32 | 55 ± 0.87 | |
| 240 min | 69 ± 7.84 | 54 ± 1.73 | 61 ± 0.28 | 47 ± 2.51 |
| Parameters | Time Point | Normal Control | Test (Active Fraction) | Acarbose |
|---|---|---|---|---|
| FBG (mg/dL) | 30 min | 89 ± 3.21 | 87 ± 1.45 | 77 ± 5.36 |
| 60 min | 114 ± 2.33 | 105 ± 3.6 | 117 ± 4.78 | |
| 120 min | 122 ± 2.31 | 77 ± 5.57 | 72 ± 1.2 | |
| 240 min | 96 ± 1.73 | 61 ± 4.67 | 56 ± 4.37 |
| Parameters | Normal Control | Test (Active Fraction) | Glibenclamide | Diabetic Control | |
|---|---|---|---|---|---|
| FBG (mg/dL) | 1st day | 95 ± 3.60 | 257 ± 9.24 | 301 ± 20.71 | 292 ± 11.85 |
| 7th day | 90 ± 2.08 | 212 ± 7.00 | 229 ± 44.08 | 363 ± 15.6 | |
| 14th day | 88 ± 1.2 | 141 ± 14.82 | 115 ± 7.67 | 356 ± 19.17 | |
| Net reduction = 61% | Net reduction = 68% | ||||
| Total protein (g/dL) | 9.22 ± 0.249 | 9.32 ± 0.05 | 11.37 ± 0.18 | 4.54 ± 0.37 | |
| Triglycerides (mg/dL) | 65.91 ± 7.53 | 103.81 ± 1.24 | 117 ± 5.35 | 215 ± 21.73 | |
| Urea (mg/dL) | 8.52 ± 0.39 | 17 ± 2.23 | 17.76 ± 0.72 | 21.71 ± 1.63 | |
| ALT (IU/L) | 7.65 ± 1.55 | 14.13 ± 2.56 | 15.9 ± 2.04 | 24.16 ± 2.79 | |
| AST (IU/L) | 7.65 ± 1.55 | 22.36 ± 0.57 | 17.67 ± 1.01 | 31.82 ± 3.60 | |
| Cholesterol (mg/dL) | 59.22 ± 2.95 | 79.56 ± 0.75 | 72.63 ± 3.02 | 99.22 ± 2.83 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unnikrishnan, P.S.; Animish, A.; Madhumitha, G.; Suthindhiran, K.; Jayasri, M.A. Bioactivity Guided Study for the Isolation and Identification of Antidiabetic Compounds from Edible Seaweed—Ulva reticulata. Molecules 2022, 27, 8827. https://doi.org/10.3390/molecules27248827
Unnikrishnan PS, Animish A, Madhumitha G, Suthindhiran K, Jayasri MA. Bioactivity Guided Study for the Isolation and Identification of Antidiabetic Compounds from Edible Seaweed—Ulva reticulata. Molecules. 2022; 27(24):8827. https://doi.org/10.3390/molecules27248827
Chicago/Turabian StyleUnnikrishnan, Pullikaparambil Sasidharan, Andhere Animish, Gunabalan Madhumitha, Krishnamurthy Suthindhiran, and Mangalam Achuthananthan Jayasri. 2022. "Bioactivity Guided Study for the Isolation and Identification of Antidiabetic Compounds from Edible Seaweed—Ulva reticulata" Molecules 27, no. 24: 8827. https://doi.org/10.3390/molecules27248827